ZnO Modulates Swine Gut Microbiota and Improves Growth Performance of Nursery Pigs When Combined with Peptide Cocktail

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Method
2.1. Animal and Diets
2.2. Gut Microbiota Analyses
2.3. Growth Performance Data Analysis
3. Results
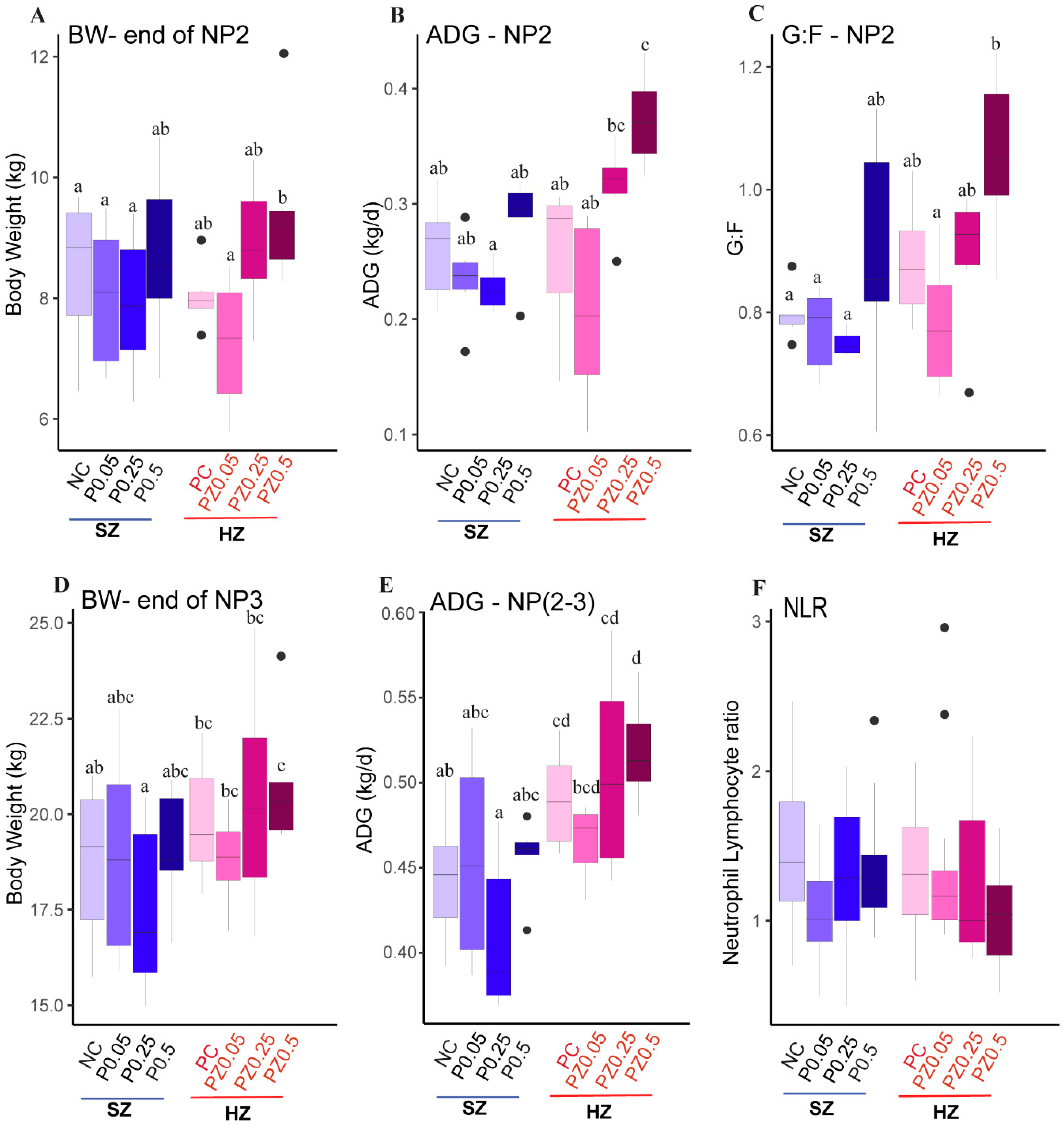
3.1. Growth Performance
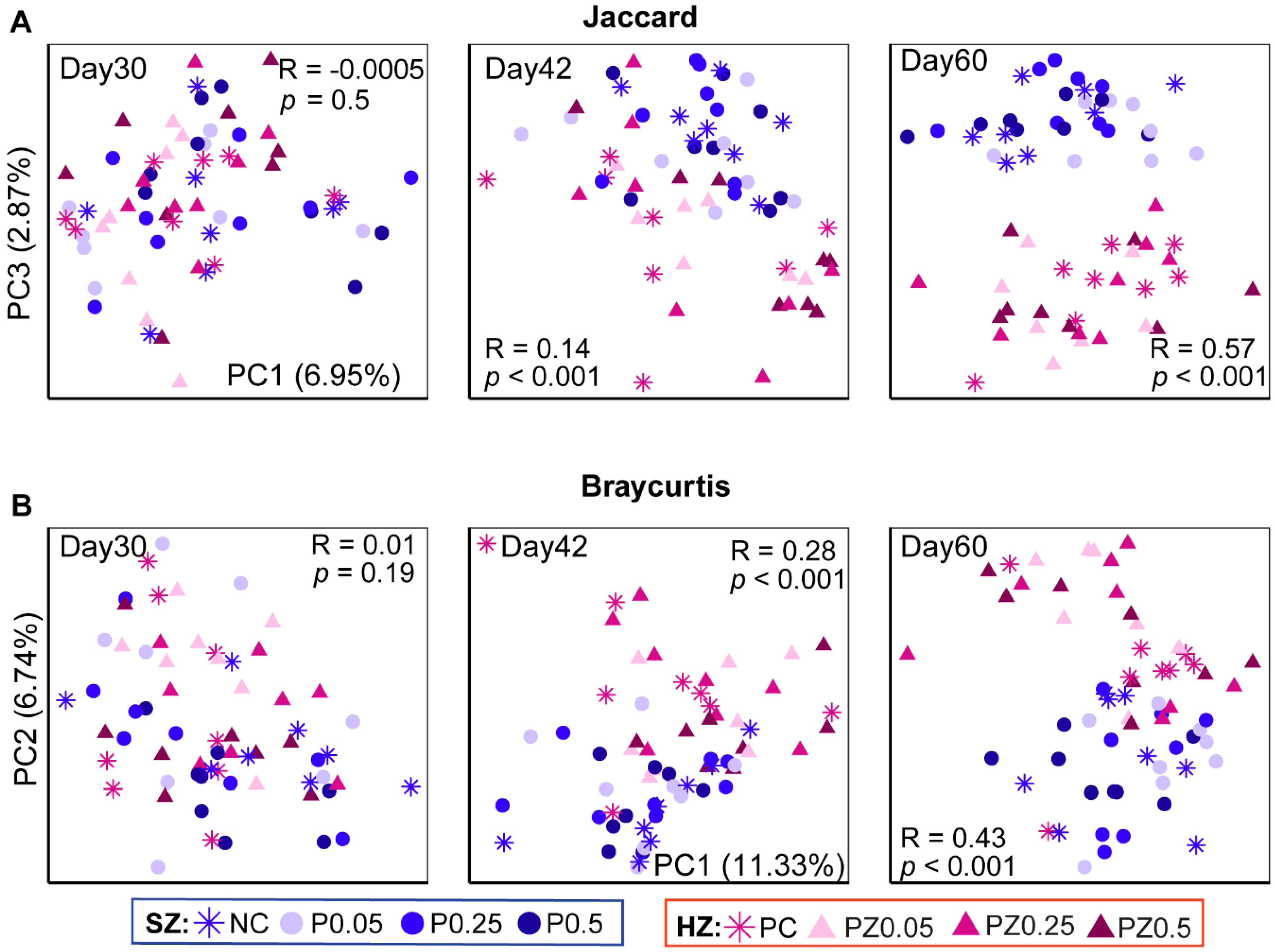
3.2. Effects of Peptide Plus ZnO on Alpha and Beta Diversity
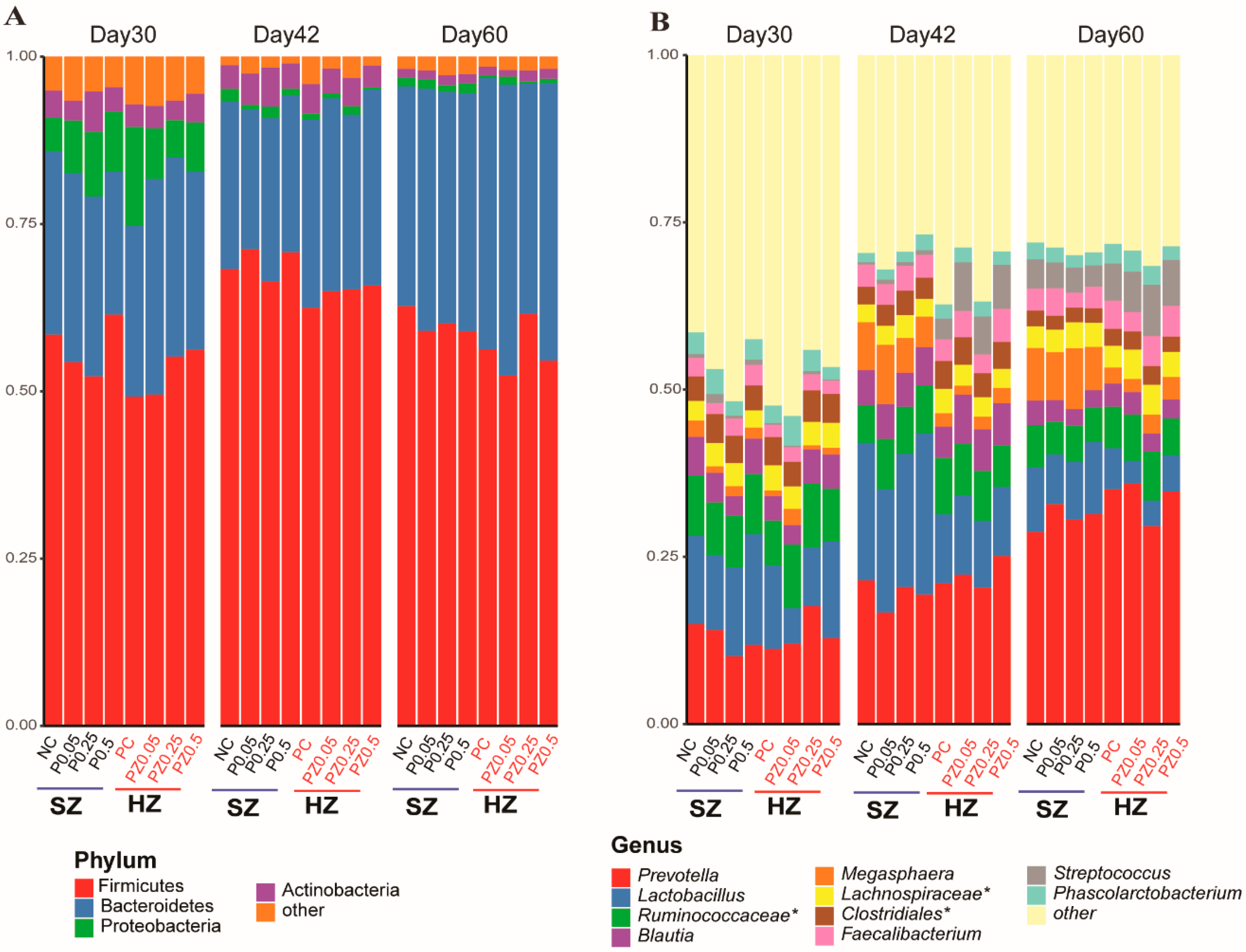
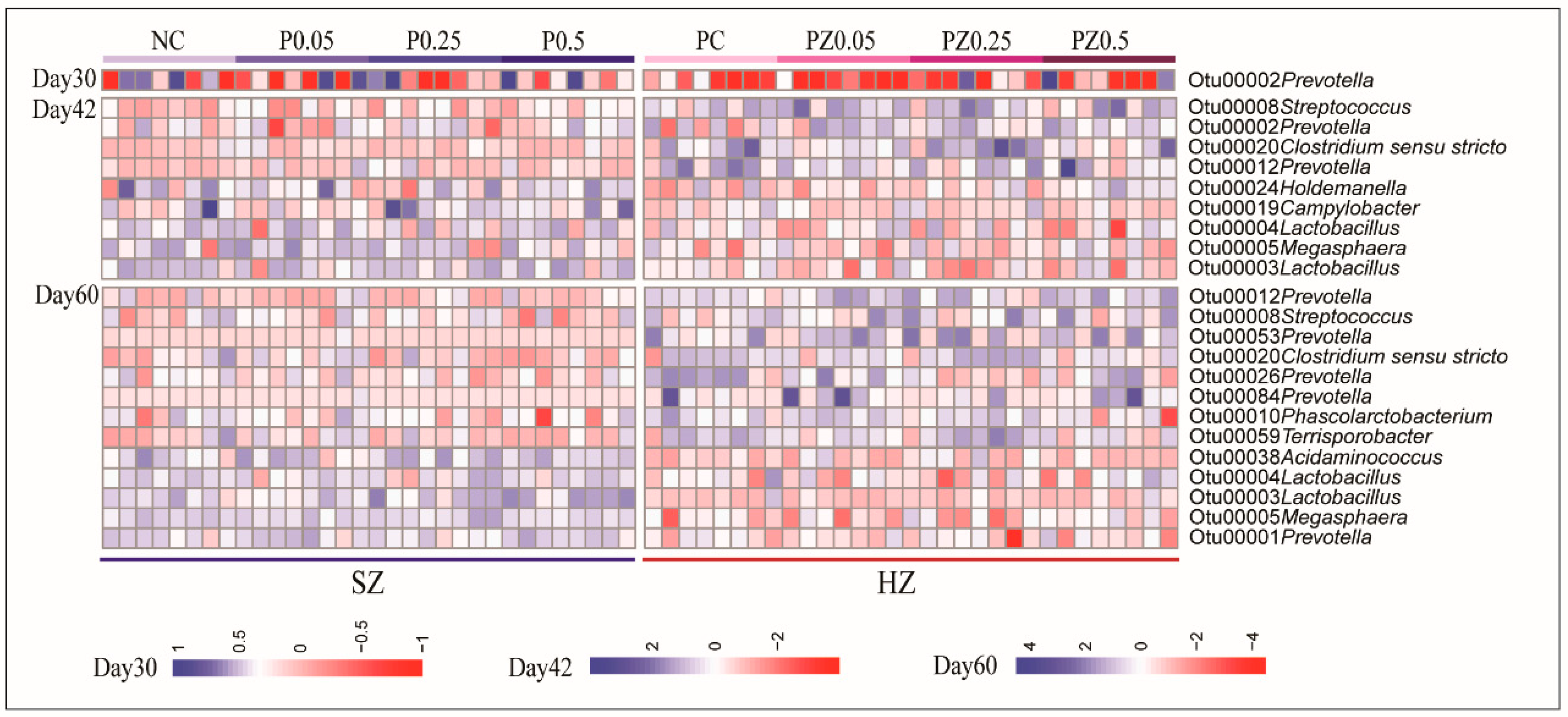
3.3. Effects of Peptide Plus ZnO on Gut Microbiota Composition
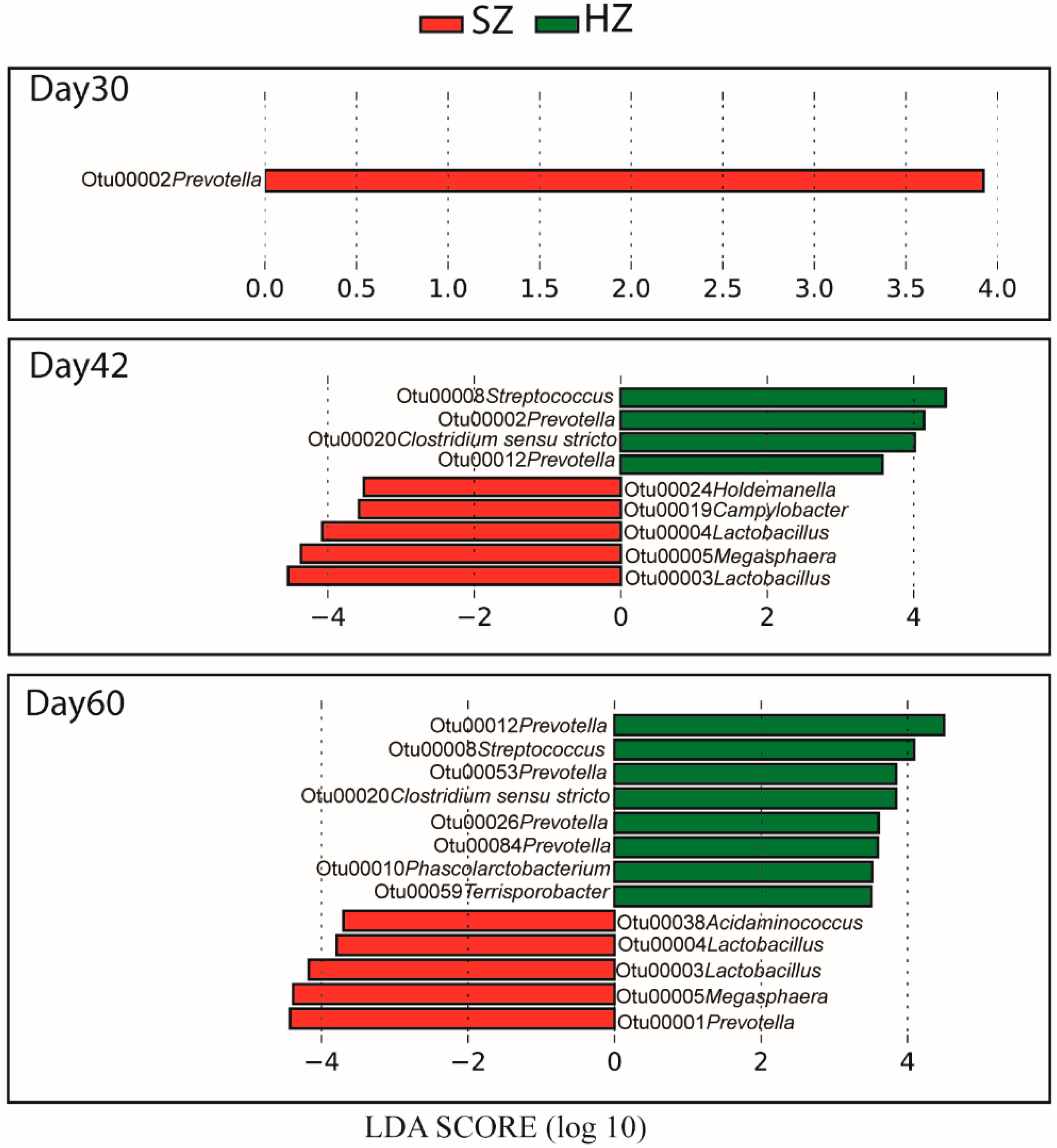
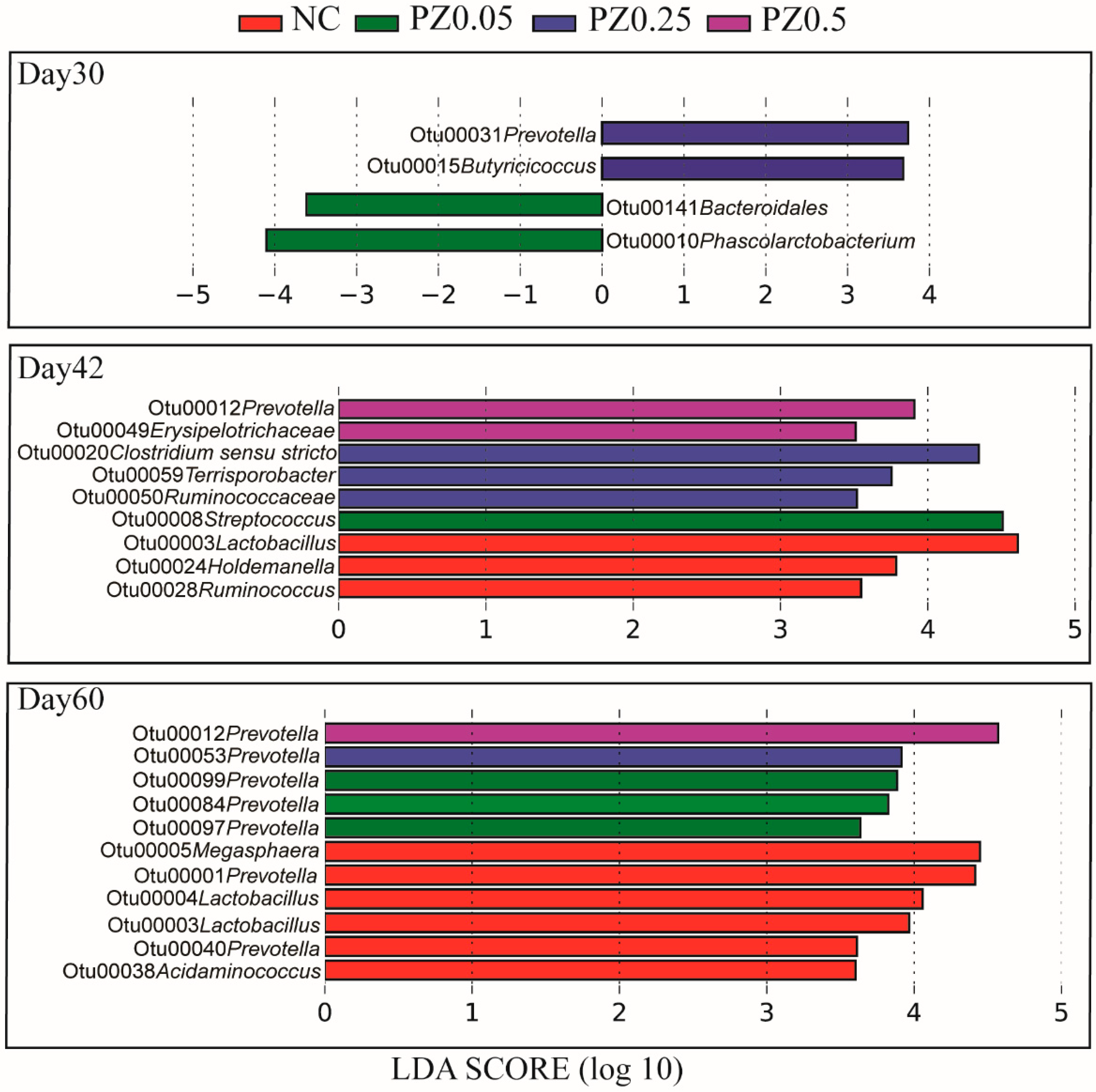
3.4. Linear Discriminant Analysis of Gut Bacteria
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Forbes, J.M. Voluntary Food Intake and Diet Selection in Farm Animals; Cabi: Wallingford, UK, 2007. [Google Scholar]
- Whittemore, C.; Green, D.J. Growth of the Young Weaned Pig; CAB International: Wallingford, UK, 2001; pp. 1–15. [Google Scholar]
- Boudry, G.; Péron, V.; Le Huërou-Luron, I.; Lallès, J.P.; Sève, B. Weaning induces both transient and long-lasting modifications of absorptive, secretory, and barrier properties of piglet intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [PubMed]
- Spreeuwenberg, M.; Verdonk, J.; Gaskins, H.; Verstegen, M.J.T.J. Small intestine epithelial barrier function is compromised in pigs with low feed intake at weaning. J. Nutr. 2001, 131, 1520–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkes, J.; Viswanathan, V.; Savkovic, S.; Hecht, G.J.G. Intestinal epithelial responses to enteric pathogens: Effects on the tight junction barrier, ion transport, and inflammation. Gut 2003, 52, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, E.; McLaughlin, J.T. Ageing and the gut. Proceed. Nutr. Soc. 2013, 72, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Soeters, P.B.; Luyer, M.D.; Greve, J.W.M.; Buurman, W.A.; Care, M. The significance of bowel permeability. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 632–638. [Google Scholar] [CrossRef]
- Modesto, M.; D’Aimmo, M.R.; Stefanini, I.; Trevisi, P.; De Filippi, S.; Casini, L.; Mazzoni, M.; Bosi, P.; Biavati, B.J.L.S. A novel strategy to select Bifidobacterium strains and prebiotics as natural growth promoters in newly weaned pigs. Livest. Sci. 2009, 122, 248–258. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Awati, A.A.; Williams, B.A.; Miller, B.G.; Jones, P.; Stokes, C.R.; Akkermans, A.D.; Smidt, H.; De Vos, W.M. Post-natal development of the porcine microbiota composition and activities. Environ. Microbiol. 2006, 8, 1191–1199. [Google Scholar] [CrossRef]
- Vondruskova, H.; Slamova, R.; Trckova, M.; Zraly, Z.; Pavlik, I. Alternatives to antibiotic growth promoters in prevention of diarrhoea in weaned piglets: A review. Vet. Med. 2010, 55, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Shankar, A.H.; Prasad, A.S. Zinc and immune function: The biological basis of altered resistance to infection. Am. J. Clin. Nutr. 1998, 68, 447S–463S. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.S. Discovery of human zinc deficiency and studies in an experimental human model. Am. J. Clin. Nutr. 1991, 53, 403–412. [Google Scholar] [CrossRef]
- Aggarwal, R.; Sentz, J.; Miller, M.A. Role of zinc administration in prevention of childhood diarrhea and respiratory illnesses: A meta-analysis. Pediatrics 2007, 119, 1120–1130. [Google Scholar] [CrossRef] [Green Version]
- Bhan, M.K.; Bhandari, N. The role of zinc and vitamin A in persistent diarrhea among infants and young children. J. Pediatr. Gastroenterol. Nutr. 1998, 26, 446–453. [Google Scholar] [CrossRef]
- Hahn, J.D.; Baker, D.H. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci. 1993, 71, 3020–3024. [Google Scholar] [CrossRef] [PubMed]
- Pettigrew, J. Reduced use of antibiotic growth promoters in diets fed to weanling pigs: Dietary tools, part 1. Anim. Biotechnol. 2006, 17, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Tokach, M.; Goodband, R.; Nelssen, J.; Richert, B.T. Effects of the interrelationship between zinc oxide and copper sulfate on growth performance of early-weaned pigs. J. Anim. Sci. 1997, 75, 1861–1866. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, H.D. Zinc and copper as feed additives, growth factors or unwanted environmental factors. J. Anim. Feed Sci. 1998, 7, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, S.; Wahab, R.; Khan, F.; Mishra, Y.K.; Musarrat, J.; Al-Khedhairy, A.A. Reactive oxygen species mediated bacterial biofilm inhibition via zinc oxide nanoparticles and their statistical determination. PLoS ONE 2014, 9, e111289. [Google Scholar] [CrossRef]
- Wang, W.; Van Noten, N.; Degroote, J.; Romeo, A.; Vermeir, P.; Michiels, J. Effect of zinc oxide sources and dosages on gut microbiota and integrity of weaned piglets. J. Anim. Physiol. Anim. Nutr. 2019, 103, 231–241. [Google Scholar] [CrossRef]
- Broom, L.; Miller, H.; Kerr, K.; Knapp, J. Effects of zinc oxide and Enterococcus faecium SF68 dietary supplementation on the performance, intestinal microbiota and immune status of weaned piglets. Res. Vet. Sci. 2006, 80, 45–54. [Google Scholar] [CrossRef]
- Højberg, O.; Canibe, N.; Poulsen, H.D.; Hedemann, M.S.; Jensen, B.B. Influence of dietary zinc oxide and copper sulfate on the gastrointestinal ecosystem in newly weaned piglets. Appl. Environ. Microbiol. 2005, 71, 2267–2277. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Song, Z.H.; Xiao, K.; Song, J.; Jiao, L.F.; Ke, Y.L. Zinc oxide influences intestinal integrity, the expressions of genes associated with inflammation and TLR4-myeloid differentiation factor 88 signaling pathways in weanling pigs. Innate Immun. 2014, 20, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, D.; Li, D.; Cao, Y.; Li, X.; Yin, J.; Qiao, S.; Wu, G. Dietary supplementation with zinc oxide decreases expression of the stem cell factor in the small intestine of weanling pigs. J. Nutr. Biochem. 2007, 18, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Song, J.; Li, Y.; Luan, Z.; Zhu, K. Diosmectite-zinc oxide composite improves intestinal barrier function, modulates expression of pro-inflammatory cytokines and tight junction protein in early weaned pigs. Br. J. Nutr. 2013, 110, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Xiao, K.; Song, J.; Luan, Z.S. Effects of zinc oxide supported on zeolite on growth performance, intestinal microflora and permeability, and cytokines expression of weaned pigs. Anim. Feed Sci. Technol. 2013, 181, 65–71. [Google Scholar] [CrossRef]
- Roselli, M.; Finamore, A.; Garaguso, I.; Britti, M.S.; Mengheri, E. Zinc oxide protects cultured enterocytes from the damage induced by Escherichia coli. J. Nutr. 2003, 133, 4077–4082. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-derived bioactive peptides on inflammation and oxidative stress. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, A.; Rai, P.; Marchbank, T.; Taylor, G.; Ghosh, S.; Ritz, B.; Playford, R.J.G. Reparative properties of a commercial fish protein hydrolysate preparation. Gut 2005, 54, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, D. Interaction of Intestinal Hydrolysate and Spray-Dried Plasma Fed to Weanling Pigs; Experiment; Iowa State University: Ames, Iowa, 1996; p. 9615. [Google Scholar]
- Zimmerman, D. The Duration of Carry-Over Growth Response to Intestinal Hydrolysate Fed to Weanling Pigs; Experiment; Iowa State University: Ames, Iowa, 1996; p. 9612. [Google Scholar]
- Frikha, M.; Mohiti-Asli, M.; Chetrit, C.; Mateos, G.G. Hydrolyzed porcine mucosa in broiler diets: Effects on growth performance, nutrient retention, and histomorphology of the small intestine. Poult. Sci. 2014, 93, 400–411. [Google Scholar] [CrossRef]
- Opheim, M.; Sterten, H.; Øverland, M.; Kjos, N.P. Atlantic salmon (Salmo salar) protein hydrolysate–Effect on growth performance and intestinal morphometry in broiler chickens. Livest. Sci. 2016, 187, 138–145. [Google Scholar] [CrossRef]
- Bui, H.T.D.; Khosravi, S.; Fournier, V.; Herault, M.; Lee, K.-J. Growth performance, feed utilization, innate immunity, digestibility and disease resistance of juvenile red seabream (Pagrus major) fed diets supplemented with protein hydrolysates. Aquaculture 2014, 418, 11–16. [Google Scholar] [CrossRef]
- Hevrøy, E.; Espe, M.; Waagbø, R.; Sandnes, K.; Ruud, M.; Hemre, G.I. Nutrient utilization in Atlantic salmon (Salmo salar L.) fed increased levels of fish protein hydrolysate during a period of fast growth. Aquac. Nutr. 2005, 11, 301–313. [Google Scholar] [CrossRef]
- Refstie, S.; Olli, J.J.; Standal, H.J.A. Feed intake, growth, and protein utilisation by post-smolt Atlantic salmon (Salmo salar) in response to graded levels of fish protein hydrolysate in the diet. Aquaculture 2004, 239, 331–349. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Ambalam, P.; Prajapati, J.; Dave, J.; Nair, B.M.; Ljungh, Å.; Vyas, B.J.M. Isolation and characterization of antimicrobial proteins produced by a potential probiotic strain of human Lactobacillus rhamnosus 231 and its effect on selected human pathogens and food spoilage organisms. Microb. Ecol. Health Dis. 2009, 21, 211–220. [Google Scholar] [CrossRef]
- Pithva, S.; Ambalam, P.; Dave, J.M.; Vyas, B.R. Potential of Probiotic Lactobacillus Strains as Food Additives; IntechOpen: London, UK, 2012. [Google Scholar]
- Lu, R.; Fasano, S.; Madayiputhiya, N.; Morin, N.P.; Nataro, J.; Fasano, A. Isolation, identification, and characterization of small bioactive peptides from Lactobacillus GG conditional media that exert both anti-Gram-negative and Gram-positive bactericidal activity. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Swine; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- The MIXED Procedure. SAS/STAT® 15.1 User’s Guide; SAS: Stockholm, Sweden, 2018; pp. 6534–6719. [Google Scholar]
- Wicklin, R. Statistical Programming with SAS/IML Software; SAS Institute: Stockholm, Sweden, 2010. [Google Scholar]
- Xiao, Y.; Kong, F.; Xiang, Y.; Zhou, W.; Wang, J.; Yang, H.; Zhang, G.; Zhao, J. Comparative biogeography of the gut microbiome between Jinhua and Landrace pigs. Sci. Rep. 2018, 8, 5985. [Google Scholar] [CrossRef]
- Coates, M.E.; Fuller, R.; Harrison, G.; Lev, M.; Suffolk, S.F. A comparision of the growth of chicks in the Gustafsson germ-free apparatus and in a conventional environment, with and without dietary supplements of penicillin. Br. J. Nutr. 1963, 17, 141–150. [Google Scholar] [CrossRef]
- Xie, Y.; He, Y.; Irwin, P.L.; Jin, T.; Shi, X. Antibacterial activity and mechanism of action of zinc oxide nanoparticles against Campylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Min, L.; Zhang, W.; Liu, J.; Hou, Z.; Chu, M.; Li, L.; Shen, W.; Zhao, Y.; Zhang, H. Zinc oxide nanoparticles influence microflora in ileal digesta and correlate well with blood metabolites. Front. Microbiol. 2017, 8, 992. [Google Scholar] [CrossRef] [PubMed]
- Imtiaz, F.; Shafique, K.; Mirza, S.S.; Ayoob, Z.; Vart, P.; Rao, S. Neutrophil lymphocyte ratio as a measure of systemic inflammation in prevalent chronic diseases in Asian population. Int. Arch. Med. 2012, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C.; Demaurex, N.; Lowell, C.A.; Uribe-Querol, E. Neutrophils: Their role in innate and adaptive immunity. J. Immunol. Res. 2016, 2016, 1469780. [Google Scholar] [CrossRef]
- Azab, B.; Zaher, M.; Weiserbs, K.F.; Torbey, E.; Lacossiere, K.; Gaddam, S.; Gobunsuy, R.; Jadonath, S.; Baldari, D.; McCord, J. Usefulness of neutrophil to lymphocyte ratio in predicting short-and long-term mortality after non–ST-elevation myocardial infarction. Am. J. Cardiol. 2010, 106, 470–476. [Google Scholar] [CrossRef]
- Boissier, R.; Campagna, J.; Branger, N.; Karsenty, G.; Lechevallier, E. The prognostic value of the neutrophil-lymphocyte ratio in renal oncology: A review. In Urologic Oncology: Seminars and Original Investigations; Elsevier: Amsterdam, The Netherlands, 2017; pp. 135–141. [Google Scholar]
- DiGangi, C. Neutrophil-lymphocyte ratio: Predicting cardiovascular and renal complications in patients with diabetes. J. Am. Assoc. Nurse Pract. 2016, 28, 410–414. [Google Scholar] [CrossRef]
- Gomez, D.; Farid, S.; Malik, H.; Young, A.; Toogood, G.; Lodge, J.; Prasad, K.R. Preoperative neutrophil-to-lymphocyte ratio as a prognostic predictor after curative resection for hepatocellular carcinoma. World J. Surg. 2008, 32, 1757–1762. [Google Scholar] [CrossRef]
- Melo, M.C.A.; Garcia, R.F.; de Araújo, C.F.C.; Abreu, R.L.C.; de Bruin, P.F.C.; de Bruin, V.M.S. Clinical significance of neutrophil-lymphocyte and platelet-lymphocyte ratios in bipolar patients: An 18-month prospective study. Psychiatry Res. 2019, 271, 8–14. [Google Scholar] [CrossRef]
- Xu, L.; Pan, Q.; Lin, R. Prevalence rate and influencing factors of preoperative anxiety and depression in gastric cancer patients in China: Preliminary study. J. Int. Med. Res. 2016, 44, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Al-Hussain, F.; Alfallaj, M.M.; Alahmari, A.N.; Almazyad, A.N.; Alsaeed, T.K.; Abdurrahman, A.A.; Murtaza, G.; Bashir, S. Relationship between neutrophil-to-lymphocyte ratio and stress in multiple sclerosis patients. J. Clin. Diagn. Res. JCDR 2017, 11, CC01. [Google Scholar]
- Akimova, E.; Lanzenberger, R.; Kasper, S. The serotonin-1A receptor in anxiety disorders. Biol. Psychiatry 2009, 66, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.; Sales, M.A.; Kim, H.; Erf, G.F.; Vo, N.; Carbonero, F.; Van Der Merwe, M.; Kegley, E.A.; Buddington, R.K.; Wang, X. Isolated rearing at lactation increases gut microbial diversity and post-weaning performance in pigs. Front. Microbiol. 2018, 9, 2889. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Huang, S.; Jiang, L.; Wang, W.; Li, T.; Zuo, B.; Li, Z.; Wang, J. Differences in the gut microbiota establishment and metabolome characteristics between low-and normal-birth-weight piglets during early life. Front. Microbiol. 2018, 9, 1798. [Google Scholar] [CrossRef] [PubMed]
- Tappenden, K.A.; Deutsch, A.S. The physiological relevance of the intestinal microbiota-contributions to human health. J. Am. Coll. Nutr. 2007, 26, 679S–683S. [Google Scholar] [CrossRef] [PubMed]
- Kaci, G.; Goudercourt, D.; Dennin, V.; Pot, B.; Doré, J.; Ehrlich, S.D.; Renault, P.; Blottière, H.M.; Daniel, C.; Delorme, C. Anti-inflammatory properties of Streptococcus salivarius, a commensal bacterium of the oral cavity and digestive tract. Appl. Environ. Microbiol. 2014, 80, 928–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mel’nikova, E.; Koroleva, N. Capacity of Lb. bulgaricus and Str. thermophilus starter to produce antibiotic substances. Dairy Sci. Abstr. 1975, 37, 4329. [Google Scholar]
- Akpinar, A.; Yerlikaya, O.; Kiliccedil, S. Antimicrobial activity and antibiotic resistance of Lactobacillus delbrueckii ssp. bulgaricus and Streptococcus thermophilus strains isolated from Turkish homemade yoghurts. Afr. J. Microbiol. Res. 2011, 5, 675–682. [Google Scholar]
- Kasraei, S.; Sami, L.; Hendi, S.; AliKhani, M.-Y.; Rezaei-Soufi, L.; Khamverdi, Z. Antibacterial properties of composite resins incorporating silver and zinc oxide nanoparticles on Streptococcus mutans and Lactobacillus. Restor. Dent. Endod. 2014, 39, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Starke, I.C.; Pieper, R.; Neumann, K.; Zentek, J.; Vahjen, W. The impact of high dietary zinc oxide on the development of the intestinal microbiota in weaned piglets. FEMS Microbiol. Ecol. 2014, 87, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Hunkapiller, A.A.; Layton, A.C.; Chang, Y.-J.; Robbins, K.R. Response of intestinal microbiota to antibiotic growth promoters in chickens. Foodborne Pathog. Dis. 2013, 10, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushida, K.; Kishimoto, A.; Piao, S.J.; Itoh, M.; Shiga, A.; Nakanishi, N.; Tsukahara, T.J. An epidemiological survey on pigs showing symptoms of infectious enteric diseases and dyspepsia in Japan. Anim. Sci. J. 2009, 80, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, K.; Tsukahara, T.; Yamada, K.; Koyama, H.; Ushida, K. Megasphaera elsdenii JCM1772T normalizes hyperlactate production in the large intestine of fructooligosaccharide-fed rats by stimulating butyrate production. J. Nutr. 2003, 133, 3187–3190. [Google Scholar] [CrossRef] [Green Version]
- Sakata, T. Influence of short chain fatty acids on intestinal growth and functions. In Dietary Fiber in Health and Disease; Springer: Berlin/Heidelberg, Germany, 1997; pp. 191–199. [Google Scholar]
- Tsukahara, T.; Iwasaki, Y.; Nakayama, K.; Ushida, K. Stimulation of butyrate production in the large intestine of weaning piglets by dietary fructooligosaccharides and its influence on the histological variables of the large intestinal mucosa. J. Nutr. Sci. Vitaminol. 2003, 49, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Shimotoyodome, A.; Meguro, S.; Hase, T.; Tokimitsu, I.; Sakata, T.J.C.B.; Molecular, P.P.A.; Physiology, I. Short chain fatty acids but not lactate or succinate stimulate mucus release in the rat colon. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2000, 125, 525–531. [Google Scholar] [CrossRef]
- Holtug, K.; Rasmussen, H.S.; Mortensen, P.B. An in vitro study of short-chain fatty acid concentrations, production and absorption in pig (Sus scrofa) colon. Comp. Biochem. Physiol. 1992, 103, 189–197. [Google Scholar] [CrossRef]
- Roediger, W.; Moore, A. Effect of short-chain fatty acid on sodium absorption in isolated human colon perfused through the vascular bed. Dig. Dis. Sci. 1981, 26, 100–106. [Google Scholar] [CrossRef]
- Starke, I.; Pieper, R.; Vahjen, W.; Zentek, J. The impact of dietary Zinc Oxide on the bacterial diversity of the small intestinal microbiota of weaned piglets. J. Vet. Sci. Technol. 2014. [Google Scholar]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial ecology along the gastrointestinal tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.; Tsai, T.; Knapp, J.; Bottoms, K.; Deng, F.; Story, R.; Maxwell, C.; Zhao, J. ZnO Modulates Swine Gut Microbiota and Improves Growth Performance of Nursery Pigs When Combined with Peptide Cocktail. Microorganisms 2020, 8, 146. https://doi.org/10.3390/microorganisms8020146
Wei X, Tsai T, Knapp J, Bottoms K, Deng F, Story R, Maxwell C, Zhao J. ZnO Modulates Swine Gut Microbiota and Improves Growth Performance of Nursery Pigs When Combined with Peptide Cocktail. Microorganisms. 2020; 8(2):146. https://doi.org/10.3390/microorganisms8020146
Chicago/Turabian StyleWei, Xiaoyuan, Tsungcheng Tsai, Joshua Knapp, Kristopher Bottoms, Feilong Deng, Robert Story, Charles Maxwell, and Jiangchao Zhao. 2020. "ZnO Modulates Swine Gut Microbiota and Improves Growth Performance of Nursery Pigs When Combined with Peptide Cocktail" Microorganisms 8, no. 2: 146. https://doi.org/10.3390/microorganisms8020146