Genomic Sequence Analysis of the Multidrug-Resistance Region of Avian Salmonella enterica serovar Indiana Strain MHYL


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Drugs Tested
2.2. Kits and Reagents
2.3. Concentration and Purification
2.4. Amplification of Drug-Resistance Genes and Southern Blot Restriction Mapping
2.5. Sequence Analysis of the MDR Region of the Strain MHYL Genome
2.6. Prediction of Strain MHYL Genome-Encoding Genes and Non-Coding Genes
3. Results
3.1. Southern Blot Hybridization of Drug-Resistance Genes
3.2. Statistics of the Strain MHYL Genome Sequencing Data
3.3. Screening and Clustering Statistics of Genes Related to Drug Resistance
3.4. Assembly and Sequencing of the Strain MHYL Genome Gene Sequence
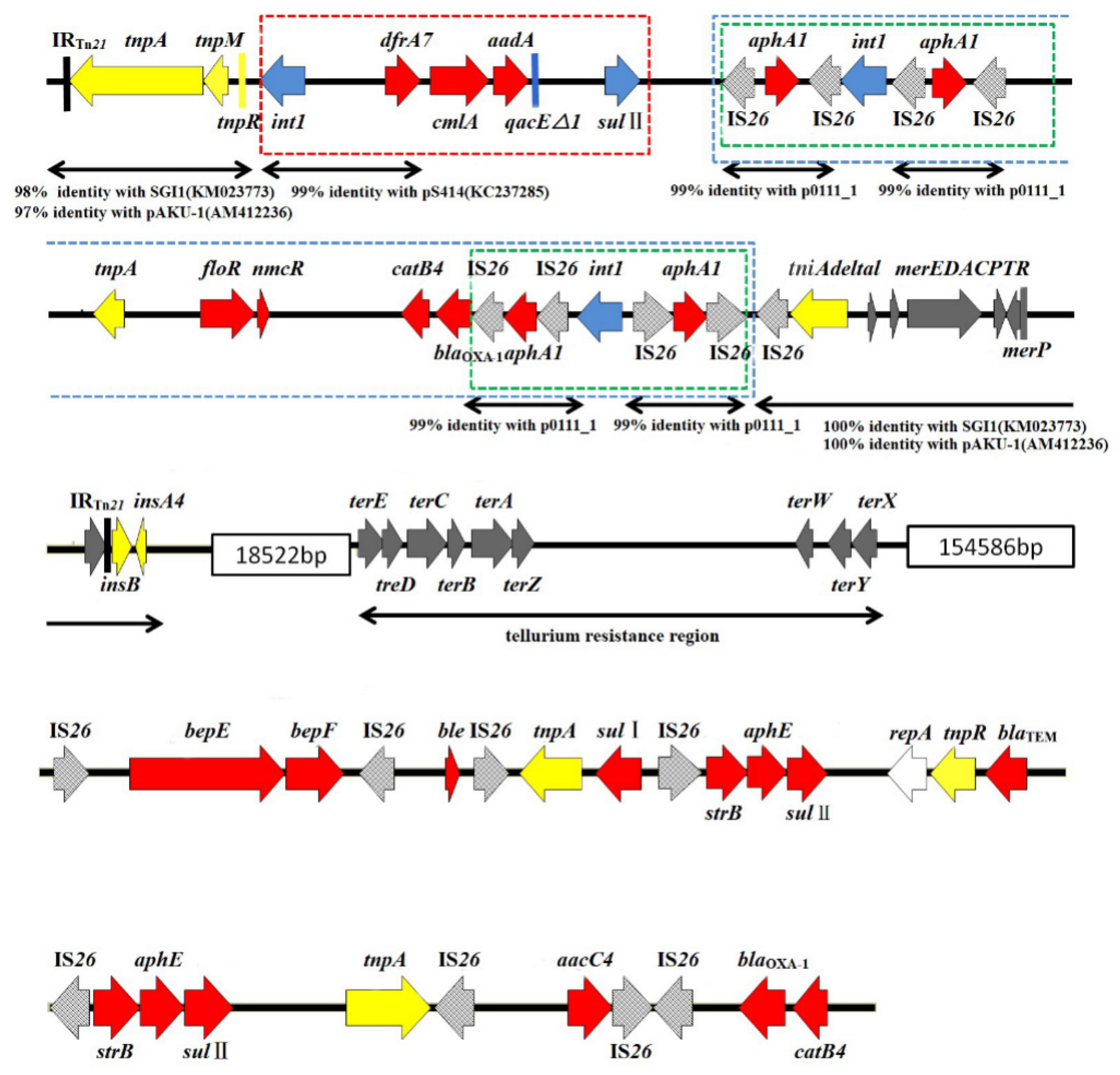
3.5. Mapping of the MDR Region of the Strain MHYL Genome
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbial. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Foodborne Diseases Active Surveillance Network (FoodNet): FoodNet Surveillance Report for 2012 (Final Report); Department of Health and Human Services: Atlanta, GA, USA, 2014; p. 9. Available online: https://www.cdc.gov/foodnet/PDFs/2012_annual_report_508c.pdf (accessed on 11 October 2018).
- Gong, J.; Kelly, P.; Wang, C. Prevalence and antimicrobial resistance of Salmonella enterica serovar Indiana in China (1984–2016). Zoonoses Public Health 2017, 64, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-S.; Wang, Y.-Y.; Zhu, Z.-Y.; Wang, H.-X.; Liu, D.-H.; Qiu, W.; Zheng, Y.; Zhang, F.-Q.; Fan, Q.-S. Identification and drug resistance of the first Salmonella Indiana in Yunnan military garrison. Chin. Trop. Med. 2013, 13, 1051–1053. [Google Scholar]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef]
- Shahada, F.; Sekizuka, T.; Kuroda, M.; Kusumoto, M.; Ohishi, D.; Matsumoto, A.; Okazaki, H.; Tanaka, K.; Uchida, I.; Izumiya, H.; et al. Characterization of Salmonella enterica serovar Typhimurium isolates harboring a chromosomally encoded CMY-2 β-lactamase gene located on a multidrug resistance genomic island. Antimicrob. Agents Chemother. 2011, 55, 4114–4121. [Google Scholar] [CrossRef] [PubMed]
- Boyd, D.A.; Peters, G.A.; Ng, L.-K.; Mulvey, M.R. Partial characterization of a genomic island associated with the multidrug resistance region of Salmonella enterica Typhymurium DT104. FEMS Microbiol. Lett. 2000, 189, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Pornsukarom, S.; Thakur, S. Horizontal dissemination of antimicrobial resistance determinants in multiple Salmonella serotypes following isolation from the commercial swine operation environment after manure application. Appl. Environ. Microbiol. 2017, 83, e01503-17. [Google Scholar] [CrossRef]
- Mulvey, M.R.; Boyd, D.A.; Olson, A.B.; Doublet, B.; Cloeckaert, A. The genetics of Salmonella genomic island 1. Microb. Infect. 2006, 8, 1915–1922. [Google Scholar] [CrossRef]
- Doublet, B.; Lailler, R.; Meunier, D.; Brisabois, A.; Boyd, D.; Mulvey, M.R.; Chasius-Dancia, E.; Cloeckaert, A. Variant Salmonella genomic island 1 antibiotic resistance gene cluster in Salmonella enterica serovar Albany. Emerg. Infect. Dis. 2003, 9, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standard Third Edition. Informational Supplement; CLSI document M31-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Lu, Y.; Zhao, H.; Sun, J.; Liu, Y.; Zhou, X.; Beier, R.C.; Wu, G.; Hou, X. Characterization of multidrug-resistant Salmonella enterica serovars Indiana and Enteritidis from chickens in Eastern China. PLoS ONE 2014, 9, e96050. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, C.-M.; Wu, G.-J.; Zhao, H.-Y.; He, T.; Cao, X.-Y.; Dai, L.; Xia, L.-N.; Qin, S.-S.; Shen, J.-Z. Prevalence of antimicrobial resistance among Salmonella isolates from chicken in China. Foodborne Pathog. Dis. 2011, 8, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.-Y.; Bao, H.-X.; Tang, L.; Zou, Q.-H.; Liu, W.-Q.; Zhu, D.-L.; Chin, J.; Dong, Y.-Y.; Li, Y.-G.; Cao, F.-L.; et al. Genomic comparison of Salmonella typhimurium DT104 with non-DT104 strains. Mol. Genet. Genom. 2013, 288, 549–557. [Google Scholar] [CrossRef]
- Lai, J.; Wang, Y.; Shen, J.; Li, R.; Han, J.; Foley, S.L.; Wu, C. Unique class 1 integron and multiple resistance genes co-located on IncHI2 plasmid is associated with the emerging multidrug resistance of Salmonella Indiana isolated from chicken in China. Foodborne Pathog. Dis. 2013, 10, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Chiou, C.-S.; Alam, M.; Kuo, J.-C.; Liu, Y.-Y.; Wang, P.-J. Chromosome-mediated multidrug resistance in Salmonella enterica serovar Typhi. Antimicrob. Agents Chemother. 2015, 59, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.E.; Thomson, N.R.; Wain, J.; Phan, M.D.; Nair, S.; Hasan, R.; Bhutta, Z.A.; Quail, M.A.; Norbeertczak, H.; Walker, D.; et al. Multidrug-resistant Salmonella enterica serovar Paratyphi A harbors IncHI1 plasmids similar to those found in serovar Typhi. J. Bacteriol. 2007, 189, 4257–4264. [Google Scholar] [CrossRef] [PubMed]
- Tauch, A.; Krieft, S.; Kalinowski, J.; Pühler, A. The 51,409-bp R-plasmid pTP10 from the multiresistant clinical isolate Corynebacterium striatum M82B is composed of DNA segments initially identified in soil bacteria and in plant, animal, and human pathogens. Mol. Gen. Genet. 2000, 263, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tauch, A.; Trost, E.; Tilker, A.; Ludewig, U.; Schneiker, S.; Goesmann, A.; Arnold, W.; Bekel, T.; Brinkrolf, K.; Brune, I.; et al. The lifestyle of Corynebacterium urealyticum derived from its complete genome sequence established by pyrosequencing. J. Biotechnol. 2008, 136, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.E.; Howe, M.M. Mobile DNA; ASM Press: Washington, DC, USA, 1989. [Google Scholar]
- Doublet, B.; Praud, K.; Bertrand, S.; Collard, J.-M.; Weill, F.-X.; Cloeckaert, A. Novel insertion sequence- and transposon-mediated genetic rearrangements in genomic island SGI1 of Salmonella enterica serovar Kentucky. Antimicrob. Agents Chemother. 2008, 52, 3745–3754. [Google Scholar] [CrossRef] [PubMed]
- Argüello, J.M.; Raimunda, D.; Padilla-Benavides, T. Mechanisms of copper homeostasis in bacteria. Front. Cell. Infect. Microbiol. 2013, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Hobman, J.L.; Crossman, L.C. Bacterial antimicrobial metal ion resistance. J. Med. Microbiol. 2014, 64, 471–497. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.-E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; et al. Comparative genomics of multidrug resistance in Acinetobacter baumannii. PloS Genet. 2006, 2, 62–72. [Google Scholar] [CrossRef]
- Barkay, T.; Miller, S.M.; Summers, A.O. Bacterial mercury resistance from atoms to ecosystems. FEMS Microbiol. Rev. 2003, 27, 355–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamlett, N.V.; Landale, E.C.; Davis, B.H.; Summers, A.O. Roles of the Tn21 merT, merP, and merC gene products in mercury resistance and mercury binding. J. Bacteriol. 1992, 174, 6377–6385. [Google Scholar] [CrossRef] [PubMed]
- Balabanov, V.P.; Kotova, V.Y.; Kholodii, G.Y.; Mindlin, S.Z.; Zavilgelsky, G.B. A novel gene, ardD, determines antirestriction activity of the non-conjugative transposon Tn5053 and is located antisense within the tniA gene. FEMS Microbiol. Lett. 2012, 337, 55–60. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antibacterial Drug Type | Gene | Upstream primer (5′–3′) Downstream primer (5′–3′) | Registration No. | Fragment Size (bp) |
|---|---|---|---|---|
| β-lactam | blaTEM | CAGCGGTAAGATCCTTGAGA ACTCCCCGTCGTGTAGATAA | AY463797 | 643 |
| Aminoglycosides | strA | CGACTTCTTACCGGACGAGGAC ACAGGTTGCGAAACGTGCCAAT | NC_009981 | 422 |
| Tetracycline | tetA | GCTACATCCTGCTTGCCTTC CATAGATCGCCGTGAAGAGG | X75761 | 210 |
| Amide alcohol | floR | TCCTGAACACGACGCCCGCTAT TCACCGCCAATGTCCCGACGAT | AJ251806 | 962 |
| Quinolones | aac(6’)-Ib-cr | TTGCGATGCTCTATGAGTGGCTA CTCGAATGCCTGGCGTGTTT | EU543272 | 482 |
| Gene | Initial Denaturation | Denaturation | Annealing | Extension | Cycles | Final Extension |
|---|---|---|---|---|---|---|
| blaTEM | 95 °C, 4 min | 94 °C, 30 s | 48 °C, 30 s | 72 °C, 1 min | 30 | 72 °C, 10 min |
| strA | 95 °C, 4 min | 94 °C, 30 s | 57 °C, 30 s | 72 °C, 1 min | 30 | 72 °C, 10 min |
| tetA | 95 °C, 4 min | 94 °C, 30 s | 57 °C, 30 s | 72 °C, 1 min | 30 | 72 °C, 10 min |
| floR | 95 °C, 4 min | 94 °C, 1 min | 63 °C, 1.5 min | 72 °C, 1.5 min | 30 | 72 °C, 10 min |
| aac(6’)-Ib-cr | 95 °C, 4 min | 94 °C, 45 s | 55 °C, 45 s | 72 °C, 45 s | 30 | 72 °C, 10 min |
| Categories | Resistance Genes | Resistance Phenotypes |
|---|---|---|
| Beta lactams | blaTEM, blaOXA, blaCTX-M-65, AmpC, nmcR | Ampicillin, amoxicillin-clavulanic acid, cefazolin, ceftiofur |
| Aminoglycosides | strA, aphA, aphE, strB, aacC4, aadA, aadA1 | Trimethoprim, gentamicin, kanamycin, amikacin |
| Tetracyclines | tetA, tetR | Tetracycline, doxycycline |
| Amide alcohols | cmlA, floR, catB4 | Chloramphenicol, florfenicol |
| Quinolones | aac(6’)-Ib-cr | Nalidixic acid, enrofloxacin, norfloxacin, dafloxacin |
| Tetracycline | sulI, sulII | Sulfisoxazole, sulfamethoxazole |
| Efflux pump membrane transport coding genes | bepD, bepE, bepF | |
| MDR protein-encoding genes | mdtA, mdtB, mdtC, mdtE, mdtG, mdtH, mdtK, mdtL, marA, marB, marC, marR, emr | |
| Fosfomycin resistance Protein-encoding gene | fsr | Fosfomycin |
| Acridine yellow resistance protein-encoding gene | acrA, acrB, acre, acrF | Acridine yellow |
| Macrolide efflux pump protein-encoding genes | macA, macB | Macrolide |
| Metal ion resistance protein encoding genes | merA, merB, merC, merE, merP,merT, merR, TerA, TerB, TerC, TerD, pcoA, pcoB, pcoC, pcoD, TerE, TerW, TerX, TerY, TerZ, zraP | Mercury, antimony, copper, zinc |
| Bleomycin resistance protein-encoding genes | Ble, bcr | Bleomycin |
| Quaternary ammonium salt-resistance protein-encoding genes | qacF, sugE | Quaternary ammonium compound |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Wen, Y.; Hu, G.; Liu, Y.; Beier, R.C.; Hou, X. Genomic Sequence Analysis of the Multidrug-Resistance Region of Avian Salmonella enterica serovar Indiana Strain MHYL. Microorganisms 2019, 7, 248. https://doi.org/10.3390/microorganisms7080248
Lu Y, Wen Y, Hu G, Liu Y, Beier RC, Hou X. Genomic Sequence Analysis of the Multidrug-Resistance Region of Avian Salmonella enterica serovar Indiana Strain MHYL. Microorganisms. 2019; 7(8):248. https://doi.org/10.3390/microorganisms7080248
Chicago/Turabian StyleLu, Yan, Yanjia Wen, Ge Hu, Yuqi Liu, Ross C. Beier, and Xiaolin Hou. 2019. "Genomic Sequence Analysis of the Multidrug-Resistance Region of Avian Salmonella enterica serovar Indiana Strain MHYL" Microorganisms 7, no. 8: 248. https://doi.org/10.3390/microorganisms7080248