
Metaproteomic and Metagenomic-Coupled Approach to Investigate Microbial Response to Electrochemical Conditions in Microbial Fuel Cells


Abstract
:1. Introduction
2. Materials and Methods
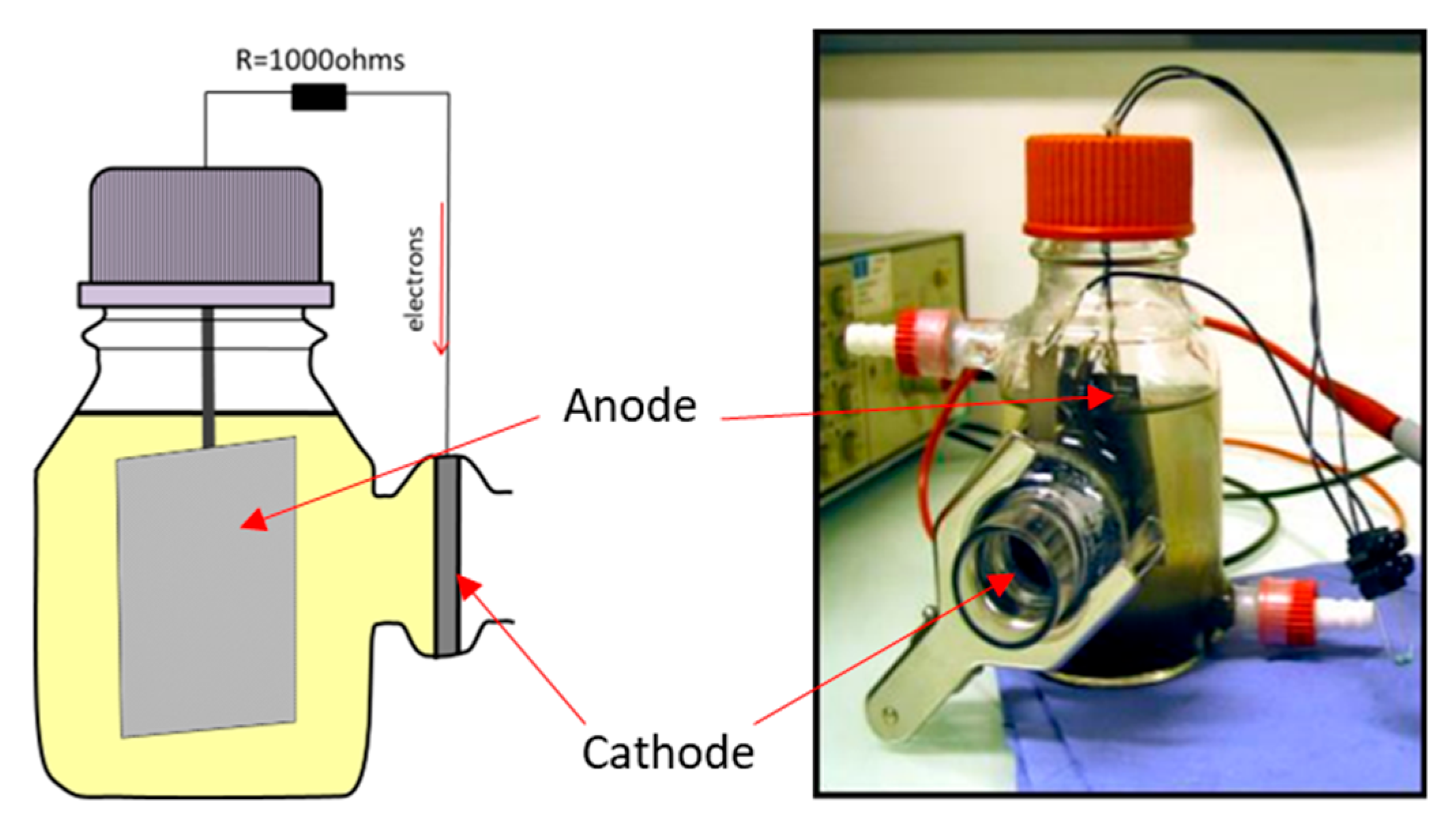
2.1. MFC Setup
2.2. Electrochemical Characterization
2.3. DNA Extraction and Sequencing
2.4. Proteins Extraction and Sequencing
2.5. Sequence Analyses
2.5.1. 16S Ribosomal RNA Gene Amplicon Sequence Analysis
2.5.2. Metagenomic and Metaproteomic Sequence Analysis
3. Results
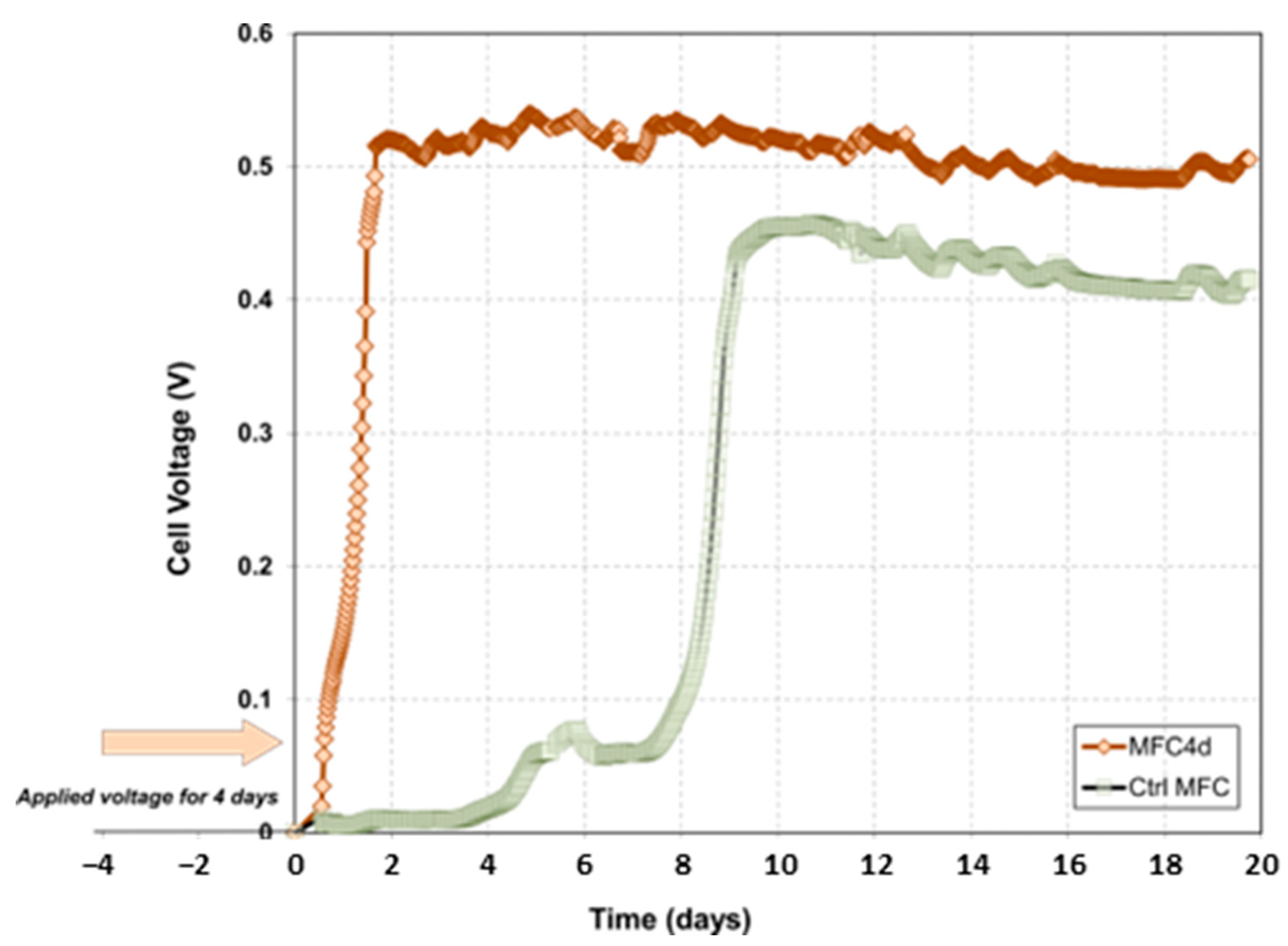
3.1. Effect of the Applied Voltage on Electricity Production
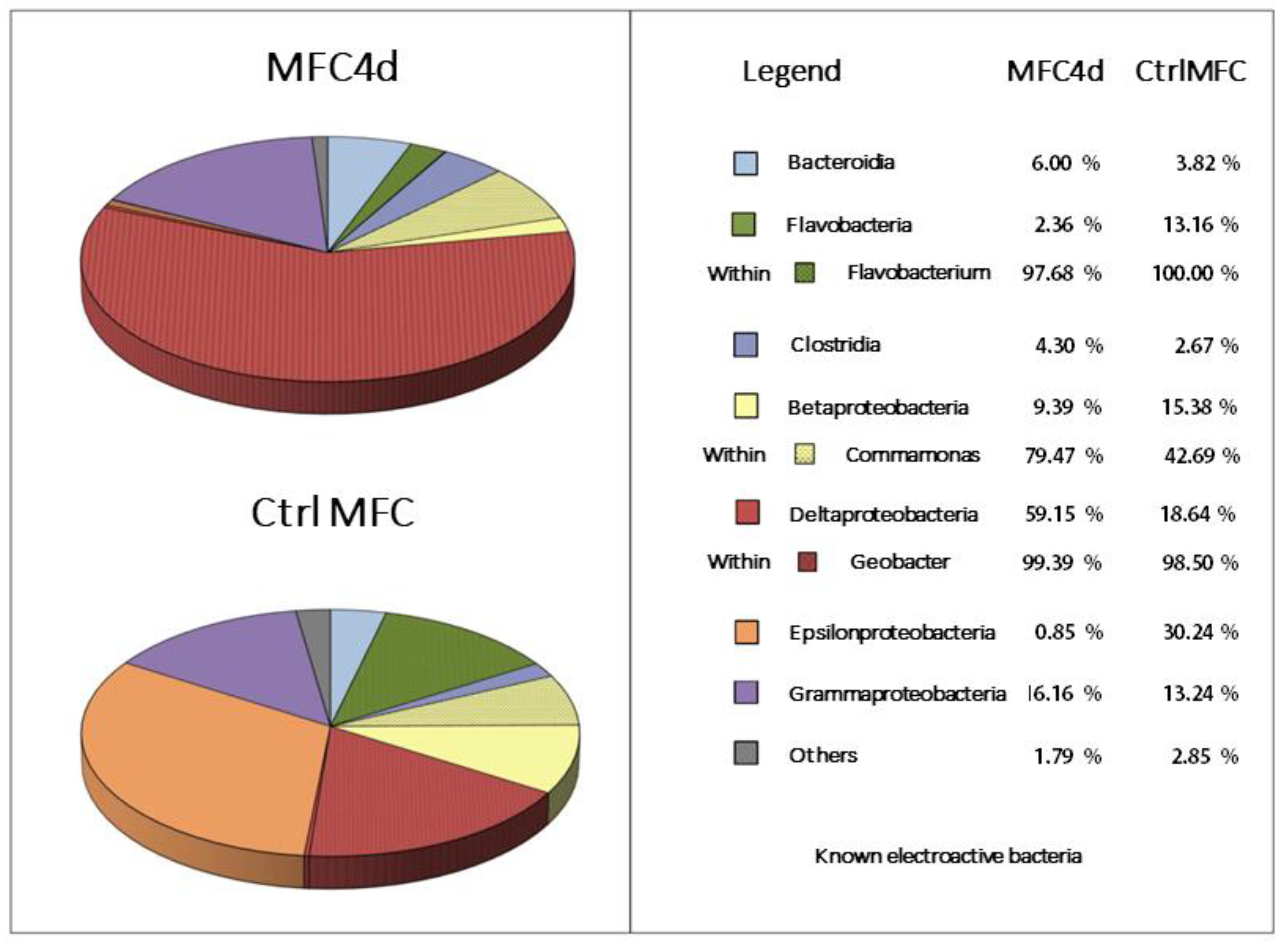
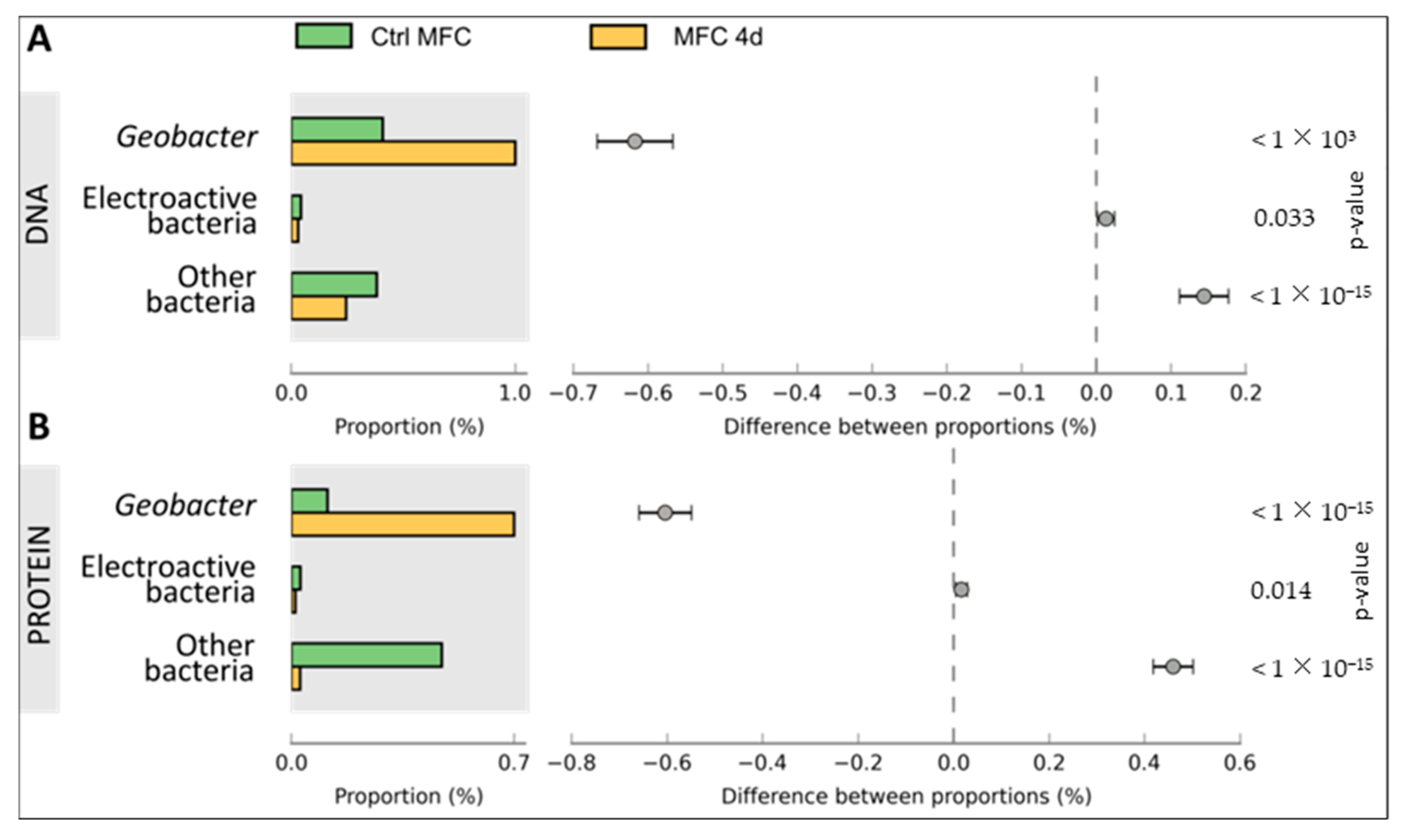
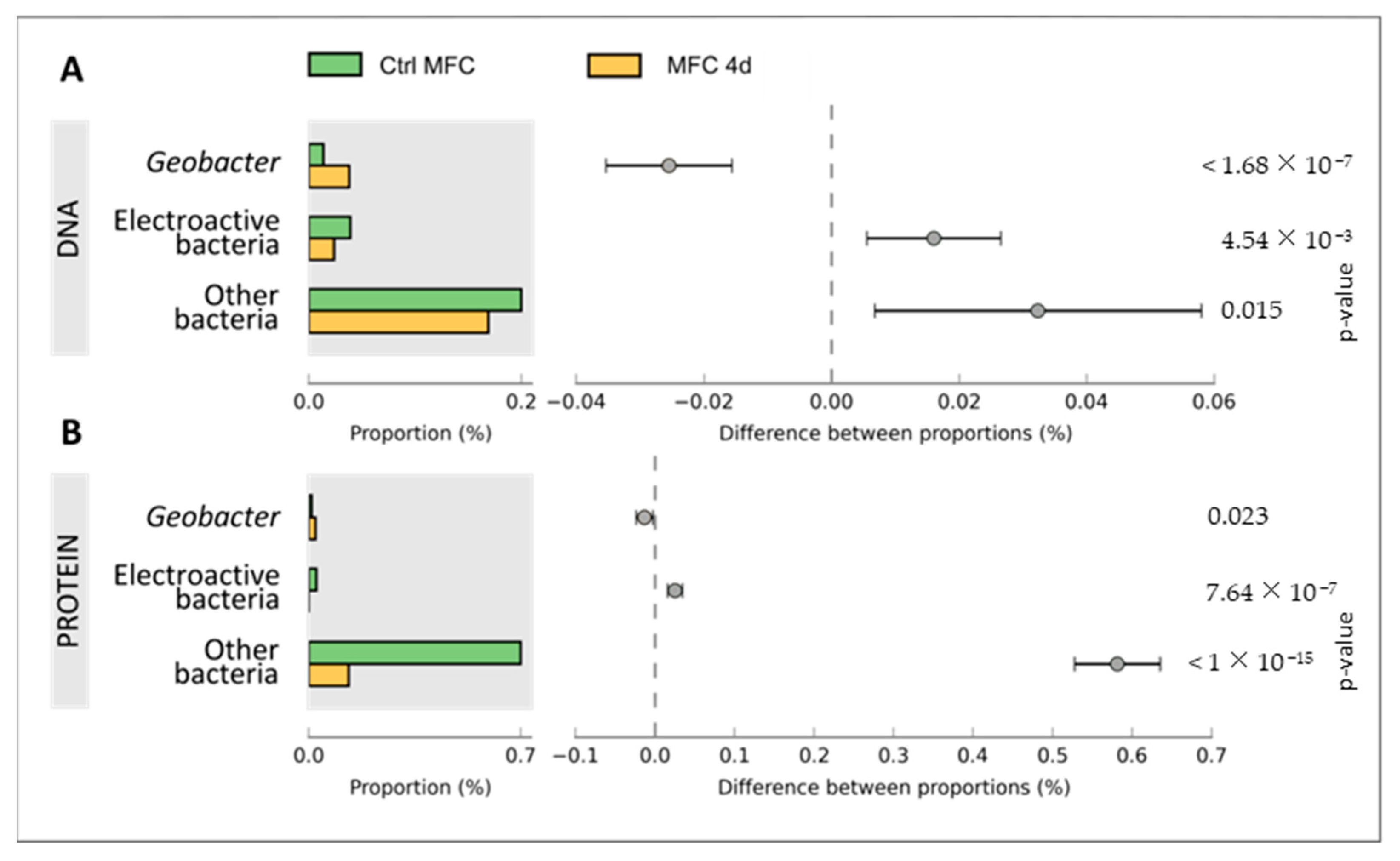
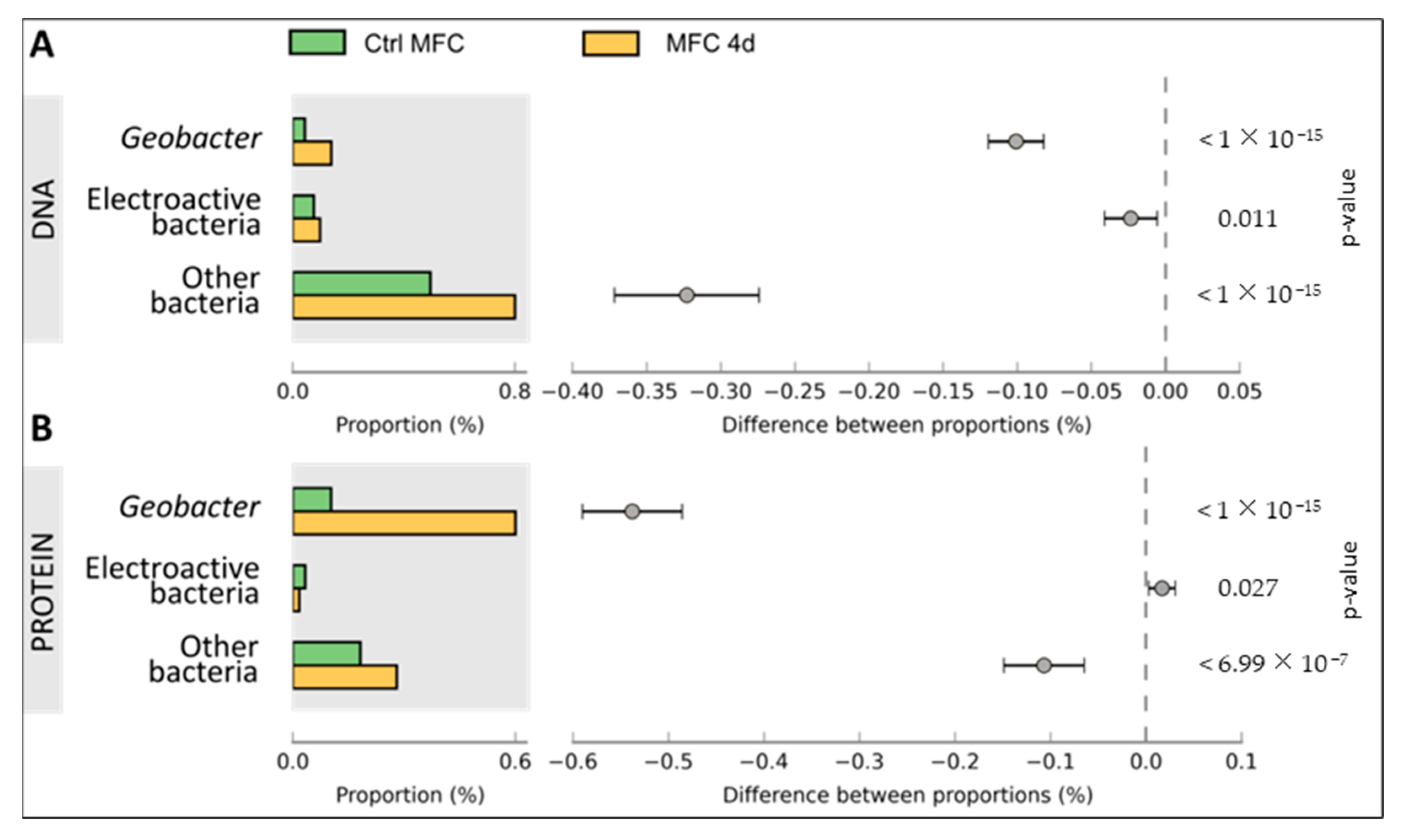
3.2. Bacterial Composition of Biofilms
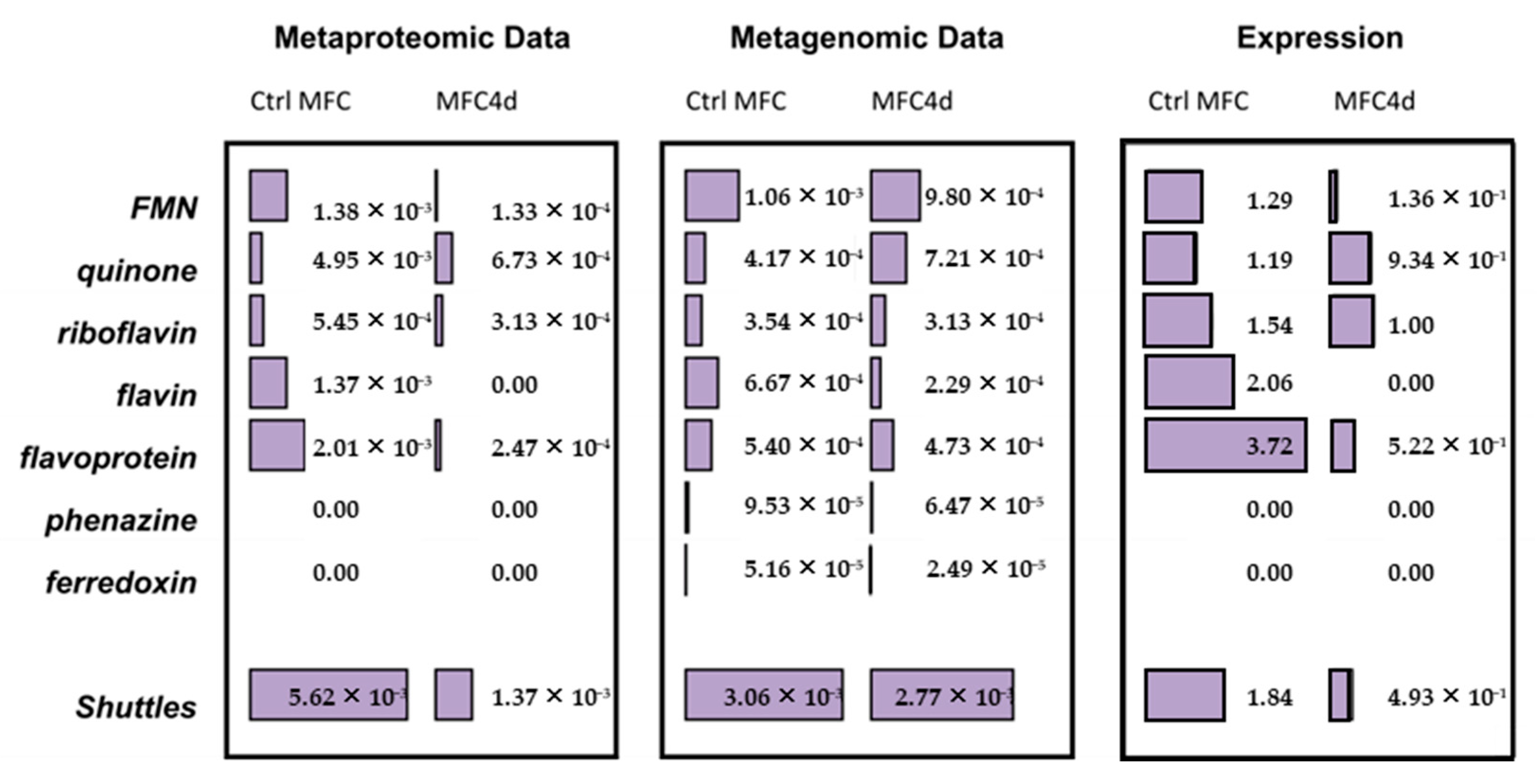
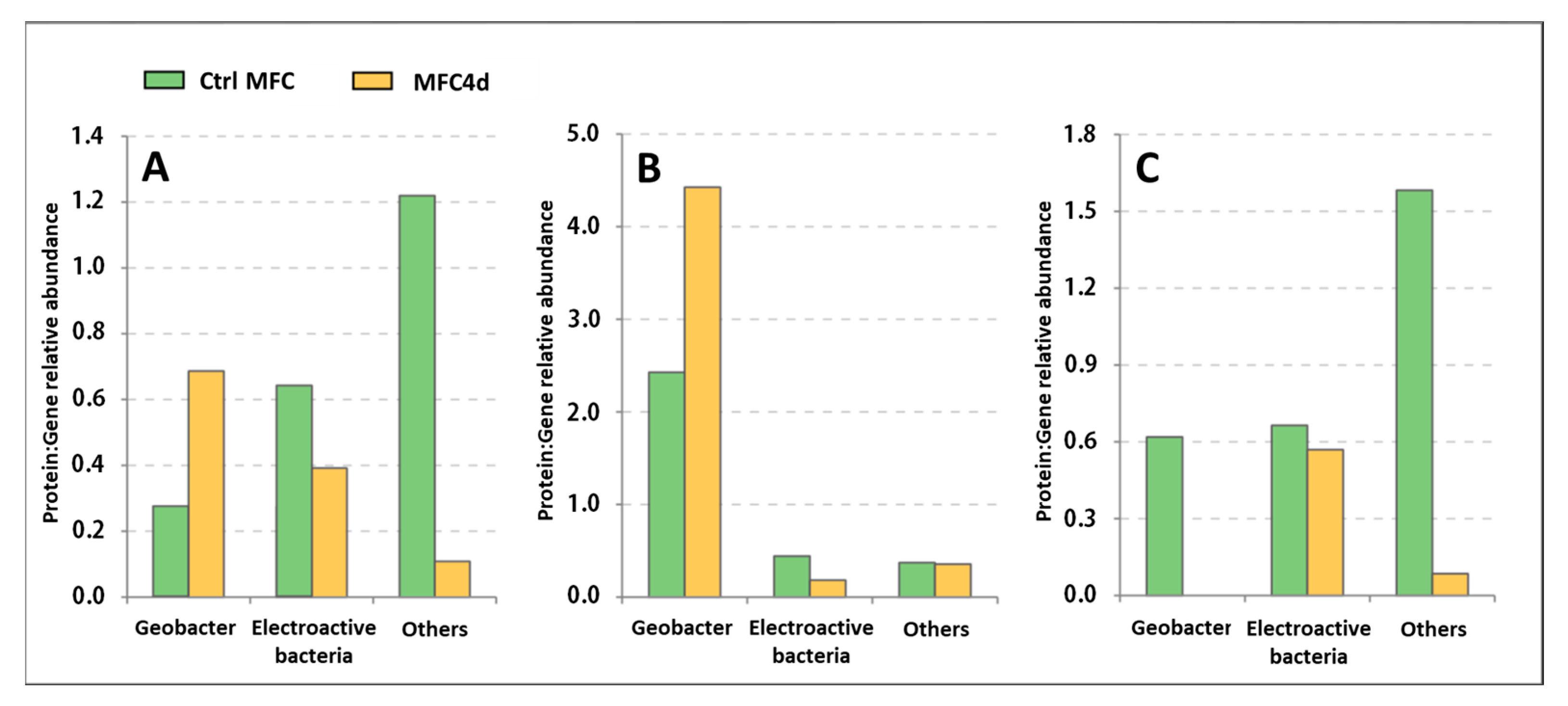
3.3. Comparison of Functional Gene Expression
4. Discussion
4.1. Electron Transfer Mechanisms in the Biofilms
4.2. Taxonomic and Functional Diversity in Microbial Biofilm
4.3. Effects of the Applied Voltage on Geobacter selection
4.4. Advantages and Limits of Metaproteomic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bataillou, G.; Lee, C.; Monnier, V.; Gerges, T.; Sabac, A.; Vollaire, C.; Haddour, N. Cedar Wood-Based Biochar: Properties, Characterization, and Applications as Anodes in Microbial Fuel Cell. Appl. Biochem. Biotechnol. 2022, 194, 4169–4186. [Google Scholar] [CrossRef] [PubMed]
- Paitier, A.; Haddour, N.; Gondran, C. Effect of Contact Area and Shape of Anode Current Collectors on Bacterial Community Structure in Microbial Fuel Cells. Molecules 2022, 27, 2245. [Google Scholar] [CrossRef] [PubMed]
- Godain, A.; Haddour, N.; Fongarland, P. Bacterial Competition for the Anode Colonization under Different External Resistances in Microbial Fuel Cells. Catalysts 2022, 12, 176. [Google Scholar] [CrossRef]
- Paitier, A.; Godain, A.; Lyon, D.; Haddour, N.; Vogel, T.M.; Monier, J.M. Microbial Fuel Cell Anodic Microbial Population Dynamics during MFC Start-Up. Biosens. Bioelectron. 2017, 92, 357–363. [Google Scholar] [CrossRef]
- Starowicz, A.; Zieliński, M.; Rusanowska, P.; Dębowski, M. Microbial Fuel Cell Performance Boost through the Use of Graphene and Its Modifications—Review. Energies 2023, 16, 576. [Google Scholar] [CrossRef]
- Malik, S.; Kishore, S.; Dhasmana, A.; Kumari, P.; Mitra, T.; Chaudhary, V.; Kumari, R.; Bora, J.; Ranjan, A.; Minkina, T.; et al. A Perspective Review on Microbial Fuel Cells in Treatment and Product Recovery from Wastewater. Water 2023, 15, 316. [Google Scholar] [CrossRef]
- Okamoto, A.; Hashimoto, K.; Nealson, K.H.; Nakamura, R. Rate Enhancement of Bacterial Extracellular Electron Transport Involves Bound Flavin Semiquinones. Proc. Natl. Acad. Sci. USA 2013, 110, 7856–7861. [Google Scholar] [CrossRef]
- Park, H.S.; Kim, B.H.; Kim, H.S.; Kim, H.J.; Kim, G.T.; Kim, M.; Chang, I.S.; Park, Y.K.; Chang, H.I. A Novel Electrochemically Active and Fe(III)-Reducing Bacterium Phylogenetically Related to Clostridium Butyricum Isolated from a Microbial Fuel Cell. Anaerobe 2001, 7, 297–306. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S.; et al. Electrically Conductive Bacterial Nanowires Produced by Shewanella Oneidensis Strain MR-1 and Other Microorganisms. Proc. Natl. Acad. Sci. USA 2006, 103, 11358–11363. [Google Scholar] [CrossRef]
- Chae, K.J.; Choi, M.J.; Lee, J.W.; Kim, K.Y.; Kim, I.S. Effect of Different Substrates on the Performance, Bacterial Diversity, and Bacterial Viability in Microbial Fuel Cells. Bioresour. Technol. 2009, 100, 3518–3525. [Google Scholar] [CrossRef]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.J.; Tappu, R. MEGAN Community Edition-Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Keller, J.; Verstraete, W.; Freguia, S.; Aelterman, P. The Anode Potential Regulates Bacterial Activity in Microbial Fuel Cells. Appl. Microbiol. Biotechnol. 2008, 78, 409–418. [Google Scholar] [CrossRef]
- Torres, C.I.; Krajmalnik-Brown, R. Selecting Anode-Respiring Bacteria Based on Anode Potentiall: Phylogenetic, electrochemical, and microscopic characterization. Environ. Sci. Technol. 2009, 43, 9519–9524. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Feng, Y.; Ren, N.; Wang, H.; Lee, H.; Li, N.; Zhao, Q. Accelerated Start-up of Two-Chambered Microbial Fuel Cells: Effect of Anodic Positive Poised Potential. Electrochim. Acta 2009, 54, 1109–1114. [Google Scholar] [CrossRef]
- Wei, J.; Liang, P.; Cao, X.; Huang, X. A New Insight into Potential Regulation on Growth and Power Generation of Geobacter Sulfurreducens in Microbial Fuel Cells Based on Energy Viewpoint. Environ. Sci. Technol. 2010, 44, 3187–3191. [Google Scholar] [CrossRef]
- Aelterman, P.; Versichele, M.; Marzorati, M.; Boon, N.; Verstraete, W. Loading Rate and External Resistance Control the Electricity Generation of Microbial Fuel Cells with Different Three-Dimensional Anodes. Bioresour. Technol. 2008, 99, 8895–8902. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, X.; Li, J.; Liao, Q.; Ye, D. Biofilm Formation and Electricity Generation of a Microbial Fuel Cell Started up under Different External Resistances. J. Power Sources 2011, 196, 6029–6035. [Google Scholar] [CrossRef]
- Yang, Y.; Li, X.; Yang, X.; He, Z. Enhanced Nitrogen Removal by Membrane-Aerated Nitritation-Anammox in a Bioelectrochemical System. Bioresour. Technol. 2017, 238, 22–29. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, M.; Ailijiang, N.; Mamat, A.; Chang, J.; Pu, M.; He, C. Impact of Voltage Application on Degradation of Biorefractory Pharmaceuticals in an Anaerobic–Aerobic Coupled Upflow Bioelectrochemical Reactor. Int. J. Environ. Res. Public Health 2022, 19, 15364. [Google Scholar] [CrossRef]
- Carrillo-Peña, D.; Escapa, A.; Hijosa-Valsero, M.; Paniagua-García, A.I.; Díez-Antolínez, R.; Mateos, R. Bioelectrochemical Enhancement of Methane Production from Exhausted Vine Shoot Fermentation Broth by Integration of MEC with Anaerobic Digestion. Biomass Convers. Biorefinery 2022. [Google Scholar] [CrossRef]
- Patil, S.A.; Surakasi, V.P.; Koul, S.; Ijmulwar, S.; Vivek, A.; Shouche, Y.S.; Kapadnis, B.P. Electricity Generation Using Chocolate Industry Wastewater and Its Treatment in Activated Sludge Based Microbial Fuel Cell and Analysis of Developed Microbial Community in the Anode Chamber. Bioresour. Technol. 2009, 100, 5132–5139. [Google Scholar] [CrossRef]
- Commault, A.S.; Lear, G.; Weld, R.J. Maintenance of Geobacter-Dominated Biofilms in Microbial Fuel Cells Treating Synthetic Wastewater. Bioelectrochemistry 2015, 106, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Regan, J.M. Comparison of Anode Bacterial Communities and Performance in Microbial Fuel Cells with Different Electron Donors. Appl. Microbiol. Biotechnol. 2007, 77, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Ki, D.; Park, J.; Lee, J.; Yoo, K. Microbial Diversity and Population Dynamics of Activated Sludge Microbial Communities Participating in Electricity Generation in Microbial Fuel Cells. Water Sci. Technol. 2008, 58, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Stratford, J.P.; Beecroft, N.J.; Slade, R.C.T.; Grüning, A.; Avignone-Rossa, C. Anodic Microbial Community Diversity as a Predictor of the Power Output of Microbial Fuel Cells. Bioresour. Technol. 2014, 156, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wei, J.; Liang, P.; Huang, X. Electricity Generation and Microbial Community Changes in Microbial Fuel Cells Packed with Different Anodic Materials. Bioresour. Technol. 2011, 102, 10886–10891. [Google Scholar] [CrossRef] [PubMed]
- Yates, M.D.; Kiely, P.D.; Call, D.F.; Rismani-Yazdi, H.; Bibby, K.; Peccia, J.; Regan, J.M.; Logan, B.E. Convergent Development of Anodic Bacterial Communities in Microbial Fuel Cells. ISME J. 2012, 6, 2002–2013. [Google Scholar] [CrossRef]
- Kiseleva, L.; Garushyants, S.K.; Ma, H.; Simpson, D.J.W.; Fedorovich, V.; Cohen, M.F.; Goryanin, I. Taxonomic and Functional Metagenomic Analysis of Anodic Communities in Two Pilot-Scale Microbial Fuel Cells Treating Different Industrial Wastewaters. J. Integr. Bioinform. 2015, 12, 273. [Google Scholar] [CrossRef]
- Osburn, E.D.; Yang, G.; Rillig, M.C.; Strickland, M.S. Evaluating the Role of Bacterial Diversity in Supporting Soil Ecosystem Functions under Anthropogenic Stress. ISME Commun. 2023, 3, 66. [Google Scholar] [CrossRef]
- Stamboulian, M.; Canderan, J.; Ye, Y. Metaproteomics as a Tool for Studying the Protein Landscape of Human-Gut Bacterial Species. PLoS Comput. Biol. 2022, 18, e1009397. [Google Scholar] [CrossRef]
- Leary, D.H.; Hervey, W.J.; Malanoski, A.P.; Wang, Z.; Eddie, B.J.; Tender, G.S.; Vora, G.J.; Tender, L.M.; Lin, B.; Strycharz-Glaven, S.M. Metaproteomic Evidence of Changes in Protein Expression Following a Change in Electrode Potential in a Robust Biocathode Microbiome. Proteomics 2015, 15, 3486–3496. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Liu, H.; Logan, B.E. Increased Performance of Single-Chamber Microbial Fuel Cells Using an Improved Cathode Structure. Electrochem. Commun. 2006, 8, 489–494. [Google Scholar] [CrossRef]
- VerBerkmoes, N.C.; Shah, M.B.; Lankford, P.K.; Pelletier, D.A.; Strader, M.B.; Tabb, D.L.; McDonald, W.H.; Barton, J.W.; Hurst, G.B.; Hauser, L.; et al. Determination and Comparison of the Baseline Proteomes of the Versatile Microbe Rhodopseudomonas Palustris under Its Major Metabolic States. J. Proteome Res. 2006, 5, 287–298. [Google Scholar] [CrossRef]
- Niessen, J.; Schröder, U.; Scholz, F. Exploiting Complex Carbohydrates for Microbial Electricity Generation-A Bacterial Fuel Cell Operating on Starch. Electrochem. Commun. 2004, 6, 955–958. [Google Scholar] [CrossRef]
- Massela, A.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: PAired-END Assembler for Illumina Sequences. Gut 2012, 13, 1–7. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Correspondence QIIME Allows Analysis of High-Throughput Community Sequencing Data Intensity Normalization Improves Color Calling in SOLiD Sequencing. Nat. Publ. Gr. 2010, 7, 335–336. [Google Scholar]
- Koch, C.; Harnisch, F. Is There a Specific Ecological Niche for Electroactive Microorganisms? ChemElectroChem 2016, 3, 1282–1295. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical Analysis of Taxonomic and Functional Profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Boon, N.; Maeyer, K. De Use of Pseudomonas Species Producing Phenazine-Based Metabolites in the Anodes of Microbial Fuel Cells to Improve Electricity Generation. Appl. Microbiol. Biotechnol. 2008, 80, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Borole, A.P.; Hamilton, C.Y.; Vishnivetskaya, T.; Leak, D.; Andras, C. Improving Power Production in Acetate-Fed Microbial Fuel Cells via Enrichment of Exoelectrogenic Organisms in Flow-through Systems. Biochem. Eng. J. 2009, 48, 71–80. [Google Scholar] [CrossRef]
- Sun, Y.; Wei, J.; Liang, P.; Huang, X. Microbial Community Analysis in Biocathode Microbial Fuel Cells Packed with Different Materials. AMB Express 2012, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; Young, N.D.; Lovley, D.R. Evolution of Electron Transfer out of the Cell. BMC Genom. 2010, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Coppi, M.V. The Hydrogenases of Geobacter Sulfurreducens: A Comparative Genomic Perspective. Microbiology 2005, 151, 1239–1254. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhuang, X.; Dong, W.; Xin, F.; Jia, H.; Wu, X. Advances in the Application of Quorum Sensing to Regulate Electrode Biofilms in Bioelectrochemical Systems. Fermentation 2023, 9, 625. [Google Scholar] [CrossRef]
- Smith, J.A.; Holmes, D.E.; Woodard, T.L.; Li, Y.; Liu, X.; Wang, L.-Y.; Meier, D.; Schwarz, I.A.; Lovley, D.R. Detrimental Impact of a Type VI Secretion System on Direct Interspecies Electron Transfer. bioRxiv 2023. [Google Scholar] [CrossRef]
- Material, S.O.; Ram, R.J.; VerBerkmoes, N.C.N.C.; Thelen, M.P.M.P.; Tyson, G.W.G.W.; Baker, B.J.; Shah, M.; Hettich, R.L.; Banfield, J.F.; Blake, R.C. Community Proteomics of a Natural Microbial Biofilm. Science 2005, 308, 1915. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functions | Metaproteomic Data | Metagenomic Data | ||
|---|---|---|---|---|
| Crtl MFC | MFC 4d | Crtl MFC | MFC 4d | |
| Cytochrome c oxidase | 2.575 | 1.602 | 5.836 | 5.277 |
| Hydrogenase | 2.457 | 1.944 | 5.517 | 5.439 |
| Function associated to acetate | 2.862 | 1.661 | 4.613 | 4.477 |
| Shuttles | 3.310 | 2.656 | 6.125 | 5.453 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godain, A.; Vogel, T.M.; Monnier, J.-M.; Paitier, A.; Haddour, N. Metaproteomic and Metagenomic-Coupled Approach to Investigate Microbial Response to Electrochemical Conditions in Microbial Fuel Cells. Microorganisms 2023, 11, 2695. https://doi.org/10.3390/microorganisms11112695
Godain A, Vogel TM, Monnier J-M, Paitier A, Haddour N. Metaproteomic and Metagenomic-Coupled Approach to Investigate Microbial Response to Electrochemical Conditions in Microbial Fuel Cells. Microorganisms. 2023; 11(11):2695. https://doi.org/10.3390/microorganisms11112695
Chicago/Turabian StyleGodain, Alexiane, Timothy M. Vogel, Jean-Michel Monnier, Agathe Paitier, and Naoufel Haddour. 2023. "Metaproteomic and Metagenomic-Coupled Approach to Investigate Microbial Response to Electrochemical Conditions in Microbial Fuel Cells" Microorganisms 11, no. 11: 2695. https://doi.org/10.3390/microorganisms11112695