
Metagenomic Approach Deciphers the Role of Community Composition of Mycobiome Structured by Bacillus velezensis VB7 and Trichoderma koningiopsis TK in Tomato Rhizosphere to Suppress Root-Knot Nematode Infecting Tomato
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Nematode Inoculum
2.2. Preparation of Liquid Formulations of B. velezensis VB7 and T. koningiopsis TK
2.3. Bioefficacy of the Liquid Formulation of B. velezensis VB7 and T. koningiopsis TK against M. incognita Infestation in Greenhouse Condition
2.4. Collection of Samples
2.5. PCR Amplification of 18S rRNA
2.6. Library Preparation and Sequencing of DNA Product
2.7. Processing of Sequencing Data and Taxonomic Profiling
2.8. Diversity Index Analysis
2.9. Taxonomic Abundance of the Microbial Population through Cluster Heatmap
2.10. Venn Diagram
2.11. Statistical Analysis
3. Results
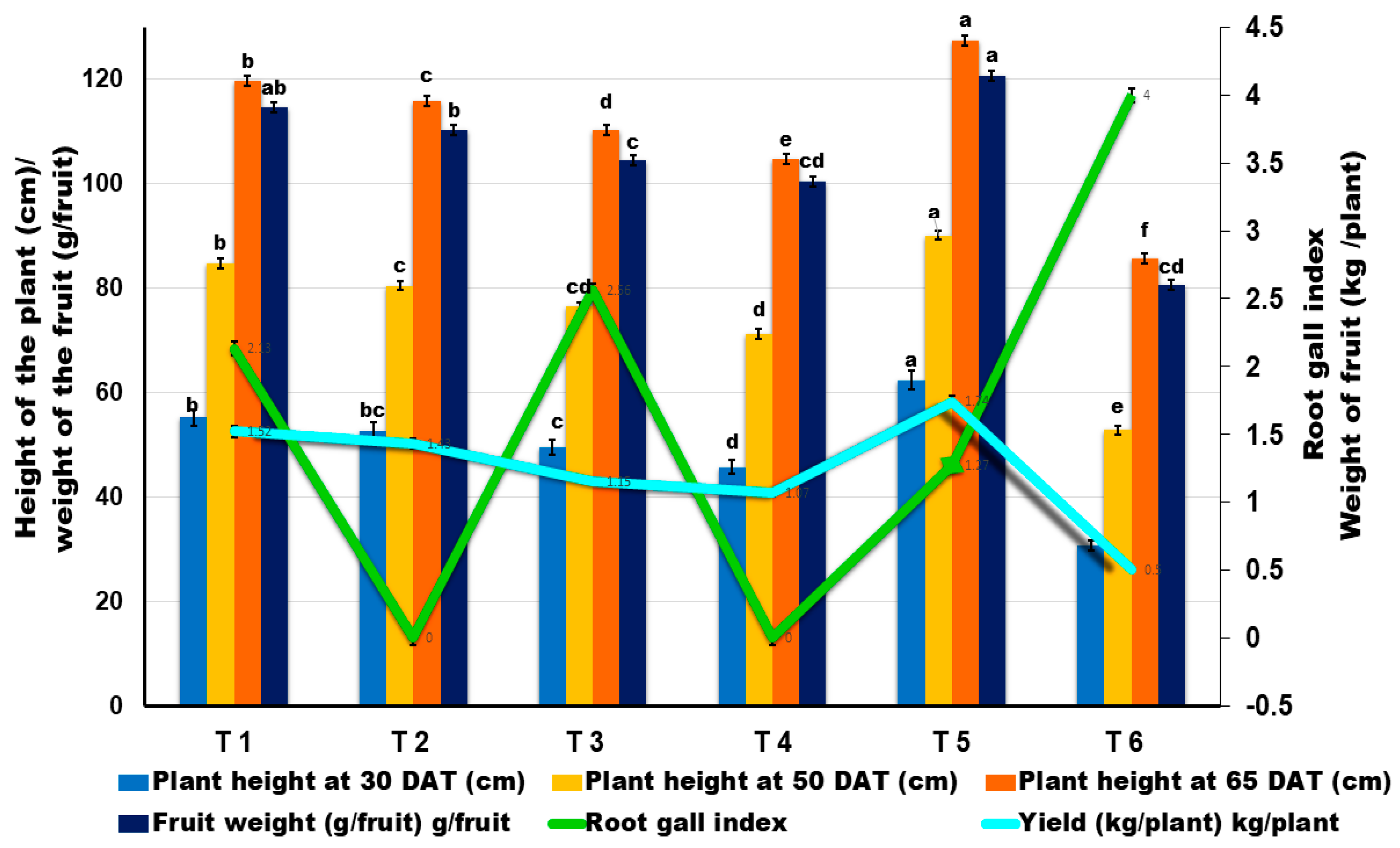
3.1. Effect of a Liquid Formulation of B. velezensis VB7 and T. koningiopsis TK against Root-Knot Nematode (M. incognita) in Tomato under Greenhouse Conditions
3.2. Identification of OTU and Taxonomic Annotation for Fungal Communities
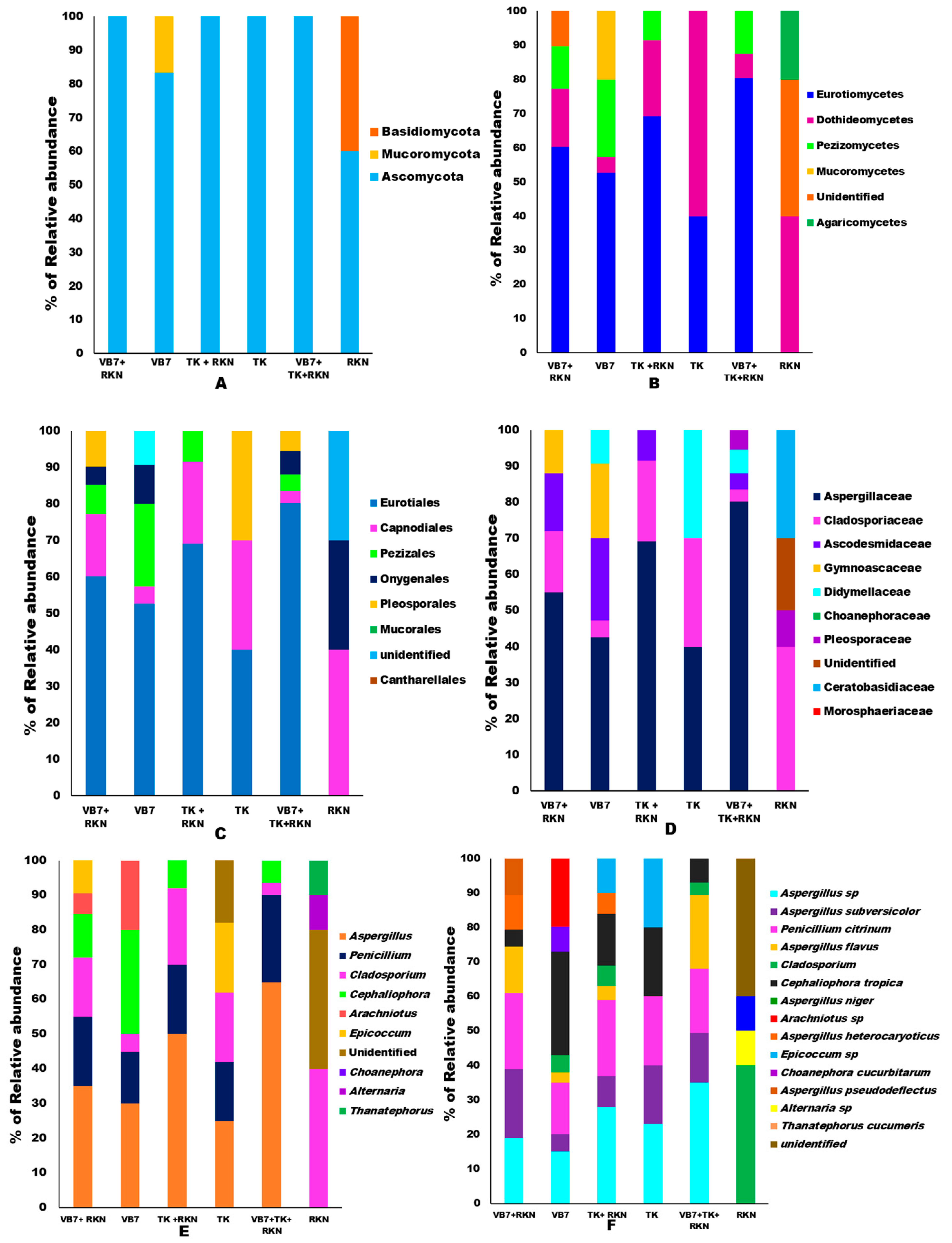
3.3. Analysis of the Microbial Community’s Composition
Relative Abundance
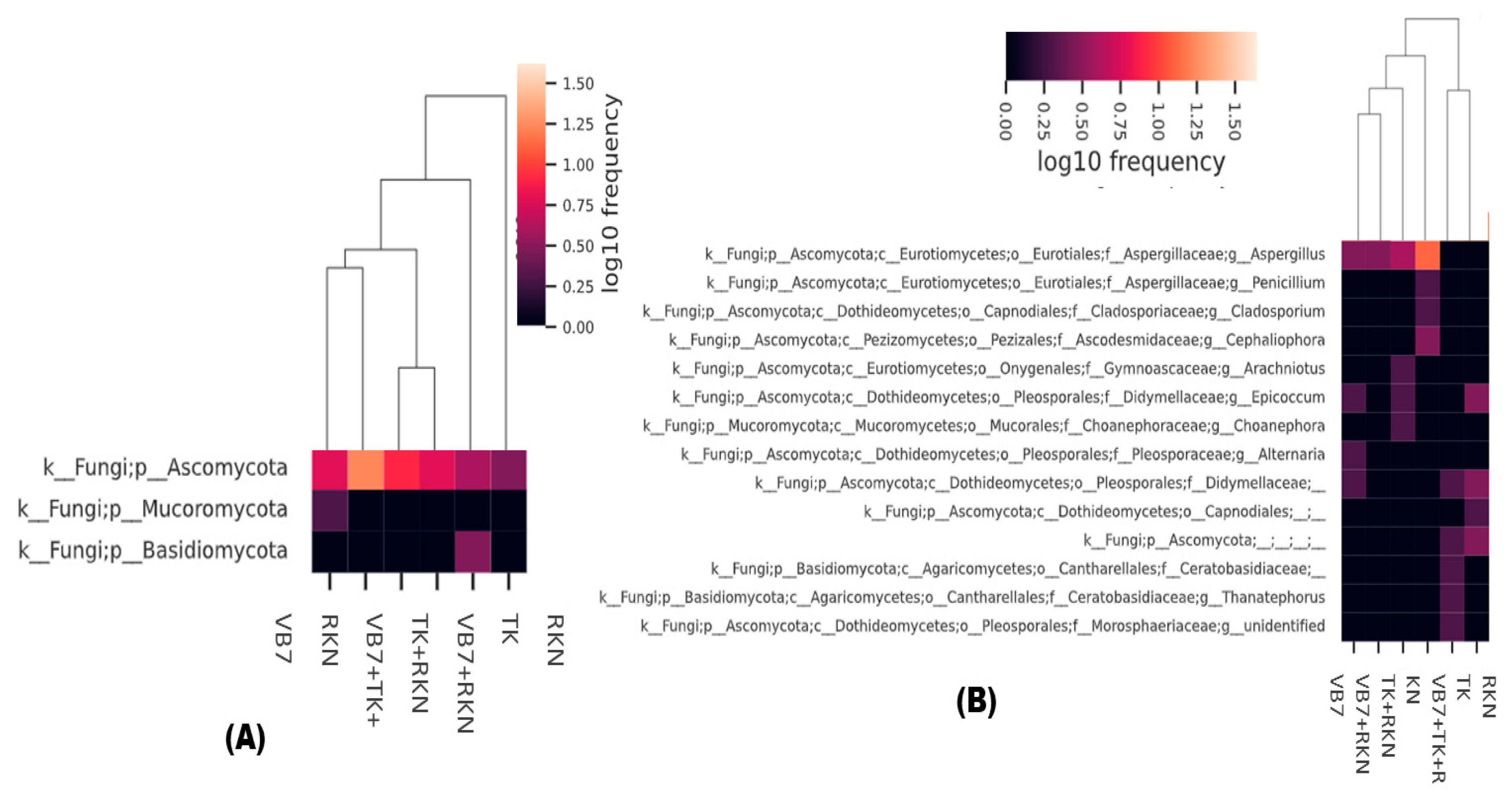
3.4. Taxonomic Abundance of Fungal Population Analyzed through a Cluster Heatmap
3.5. Comparison of OTUs in Different Treatments
3.6. Diversity of Fungal Communities in Different Rhizosphere Soils
3.6.1. Alpha Diversity Indexes
3.6.2. Beta Diversity Indexes
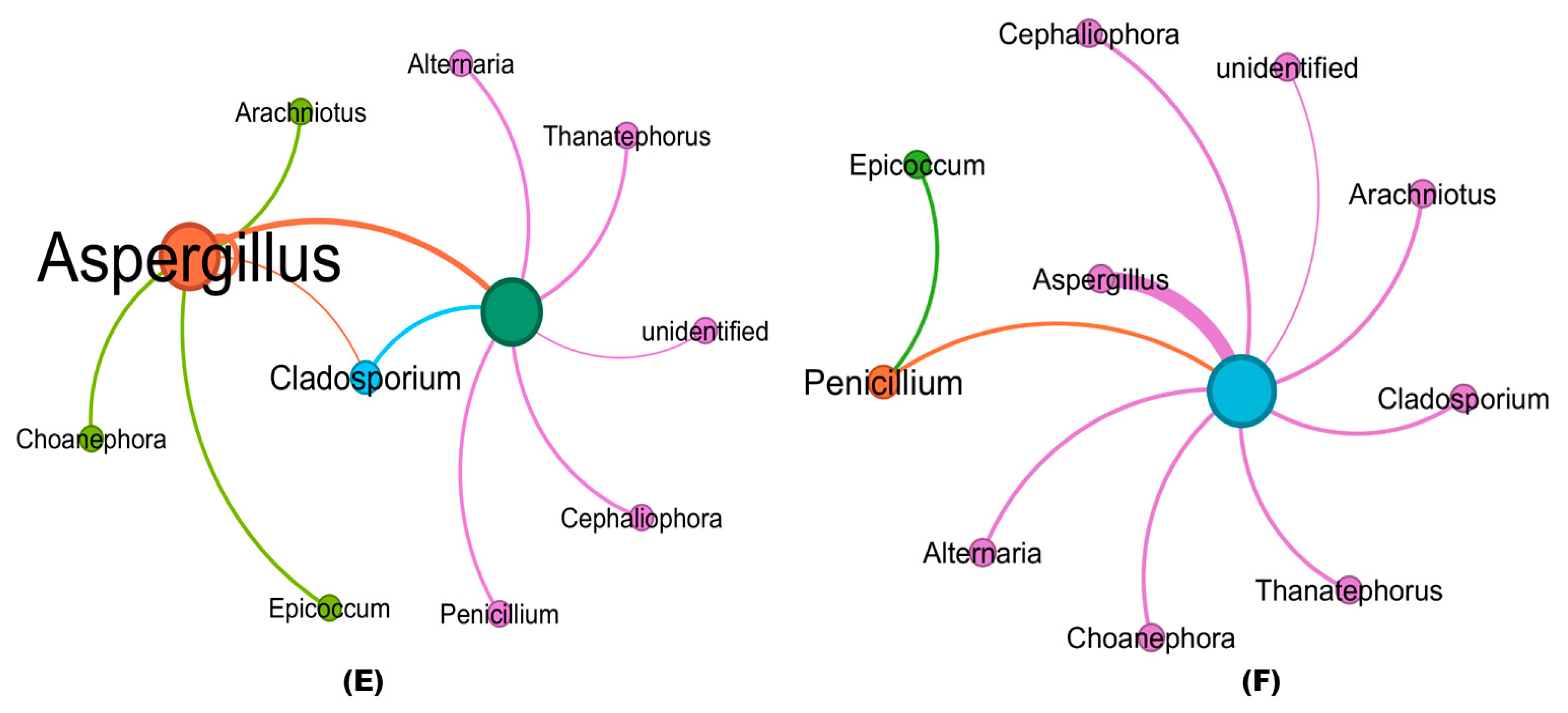
3.7. Co-Occurrence Clustering Coefficient Analysis of Fungal Communities in Treated and Untreated Soil Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simões, M.F.; Antunes, A.; Ottoni, C.A.; Amini, M.S.; Alam, I.; Alzubaidy, H.; Mokhtar, N.-A.; Archer, J.A.; Bajic, V.B. Soil and rhizosphere associated fungi in gray mangroves (Avicennia marina) from the Red Sea—A metagenomic approach. Genom. Proteom. Bioinform. 2015, 13, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Espenberg, M.; Pärn, J.; Lehtovirta-Morley, L.; Anslan, S.; Kasak, K.; Kõljalg, U.; Liira, J.; Maddison, M.; Moora, M. Structure and function of the soil microbiome underlying N2O emissions from global wetlands. Nat. Commun. 2022, 13, 1430. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef]
- Leff, J.W.; Fierer, N. Bacterial communities associated with the surfaces of fresh fruits and vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Tapia-Vázquez, I.; Montoya-Martínez, A.C.; De los Santos-Villalobos, S.; Ek-Ramos, M.J.; Montesinos-Matías, R.; Martínez-Anaya, C. Root-knot nematodes (Meloidogyne spp.) a threat to agriculture in Mexico: Biology, current control strategies, and perspectives. World J. Microbiol. Biotechnol. 2022, 38, 26. [Google Scholar] [CrossRef]
- Afriyie Boakye, T.; Kwadwo Anning, D.; Li, H.; Osei, R.; Boamah, S.; Anim, S. Mechanism of Antagonistic Bioagents in Controlling Root-Knot Nematodes (Meloidogyne sp.): A Review. Asian Res. J. Agric. 2022, 15, 27–44. [Google Scholar] [CrossRef]
- Liu, M.; Philp, J.; Wang, Y.; Hu, J.; Wei, Y.; Li, J.; Ryder, M.; Toh, R.; Zhou, Y.; Denton, M.D. Plant growth-promoting rhizobacteria Burkholderia vietnamiensis B418 inhibits root-knot nematode on watermelon by modifying the rhizosphere microbial community. Sci. Rep. 2022, 12, 8381. [Google Scholar] [CrossRef]
- Alberton, D.; Valdameri, G.; Moure, V.R.; Monteiro, R.A.; Pedrosa, F.d.O.; Müller-Santos, M.; de Souza, E.M. What did we learn from plant growth-promoting rhizobacteria (PGPR)-grass associations studies through proteomic and metabolomic approaches? Front. Sustain. Food Syst. 2020, 4, 607343. [Google Scholar] [CrossRef]
- Dehghanian, S.; Abdollahi, M.; Charehgani, H.; Niazi, A. Combined of salicylic acid and Pseudomonas fluorescens CHA0 on the expression of PR1 gene and control of Meloidogyne javanica in tomato. Biol. Control 2020, 141, 104134. [Google Scholar]
- Migunova, V.D.; Sasanelli, N. Bacteria as biocontrol tool against phytoparasitic nematodes. Plants 2021, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Tariq-Khan, M.; Mukhtar, T.; Munir, A.; Hallmann, J.; Heuer, H. Comprehensive report on the prevalence of root-knot nematodes in the Poonch division of Azad Jammu and Kashmir, Pakistan. J. Phytopathol. 2020, 168, 322–336. [Google Scholar] [CrossRef]
- Půža, V.; Tarasco, E. Interactions between Entomopathogenic Fungi and Entomopathogenic Nematodes. Microorganisms 2023, 11, 163. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.-q.; Wang, M.; Sun, J.; Gao, J.-x.; Chen, J. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium Stalk rot. Sci. Rep. 2017, 7, 1771. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Guo, C.; Zhao, P.; Yu, F.-Y.; Su, Y.; Qu, J.-P.; Wang, J.-L.; Lin, R.-S.; Wang, B.; Gao, Z. Biocontrol potential of Bacillus altitudinis AMCC1040 against root-knot nematode disease of ginger and its impact on rhizosphere microbial community. Biol. Control 2021, 158, 104598. [Google Scholar] [CrossRef]
- Tian, B.-Y.; Cao, Y.; Zhang, K.-Q. Metagenomic insights into communities, functions of endophytes and their associates with infection by root-knot nematode, Meloidogyne incognita, in tomato roots. Sci. Rep. 2015, 5, 17087. [Google Scholar] [CrossRef]
- Vinodkumar, S.; Nakkeeran, S.; Renukadevi, P.; Mohankumar, S. Diversity and antiviral potential of rhizospheric and endophytic Bacillus species and phyto-antiviral principles against tobacco streak virus in cotton. Agric. Ecosyst. Environ. 2018, 267, 42–51. [Google Scholar]
- Suneeta, P.; Aiyanathan, K.E.A.; Nakkeeran, S. Evaluation of Trichoderma spp. and Fungicides in the Management of Collar Rot of Gerbera Incited by Sclerotium rolfsii. J. Pure Appl. Microbiol. 2017, 11, 1161–1168. [Google Scholar] [CrossRef]
- Taylor, A.L.; Sasser, J.N. Biology, identification and control of root-knot nematodes (Meloidogyne species). In Biology, Identification and Control of Root-Knot Nematodes (Meloidogyne Species); Department of Plant Pathology, North Carolina State University: Raleigh, NC, USA, 1978. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Willis, A.D.; Martin, B.D. Estimating diversity in networked ecological communities. Biostatistics 2022, 23, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A. Soil multifunctionality is affected by the soil environment and by microbial community composition and diversity. Soil Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Malacrino, A.; Wisniewski, M.; Cacciola, S.O.; Schena, L. Metabarcoding: A powerful tool to investigate microbial communities and shape future plant protection strategies. Biol. Control 2018, 120, 1–10. [Google Scholar] [CrossRef]
- Cao, H.; Jiao, Y.; Yin, N.; Li, Y.; Ling, J.; Mao, Z.; Yang, Y.; Xie, B. Analysis of the activity and biological control efficacy of the Bacillus subtilis strain Bs-1 against Meloidogyne incognita. Crop Prot. 2019, 122, 125–135. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Z.; Xie, J.; Hesselberg-Thomsen, V.; Tan, T.; Zheng, D.; Strube, M.L.; Dragoš, A.; Shen, Q.; Zhang, R. Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 2022, 16, 774–787. [Google Scholar] [CrossRef]
- Yin, N.; Zhao, J.-L.; Liu, R.; Li, Y.; Ling, J.; Yang, Y.-H.; Xie, B.-Y.; Mao, Z.-C. Biocontrol efficacy of Bacillus cereus strain Bc-cm103 against Meloidogyne incognita. Plant Dis. 2021, 105, 2061–2070. [Google Scholar] [CrossRef]
- Fan, H.; Yao, M.; Wang, H.; Zhao, D.; Zhu, X.; Wang, Y.; Liu, X.; Duan, Y.; Chen, L. Isolation and effect of Trichoderma citrinoviride Snef1910 for the biological control of root-knot nematode, Meloidogyne incognita. BMC Microbiol. 2020, 20, 299. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Fernandez, I.; Lok, G.B.; Pozo, M.J.; Pieterse, C.M.; Van Wees, S.C. Shifting from priming of salicylic acid-to jasmonic acid-regulated defences by Trichoderma protects tomato against the root knot nematode Meloidogyne incognita. New Phytol. 2017, 213, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Tazi, H.; Hamza, M.A.; Hallouti, A.; Benjlil, H.; Idhmida, A.; Furze, J.N.; Paulitz, T.C.; Mayad, E.H.; Boubaker, H.; El Mousadik, A. Biocontrol potential of nematophagous fungi against Meloidogyne spp. infecting tomato. Org. Agric. 2021, 11, 63–71. [Google Scholar] [CrossRef]
- d’Errico, G.; Greco, N.; Vinale, F.; Marra, R.; Stillittano, V.; Davino, S.W.; Woo, S.L.; D’Addabbo, T. Synergistic Effects of Trichoderma harzianum, 1, 3 Dichloropropene and Organic Matter in Controlling the Root-Knot Nematode Meloidogyne incognita on Tomato. Plants 2022, 11, 2890. [Google Scholar] [CrossRef] [PubMed]
- Saharan, R.; Patil, J.; Yadav, S.; Kumar, A.; Goyal, V. The nematicidal potential of novel fungus, Trichoderma asperellum FbMi6 against Meloidogyne incognita. Sci. Rep. 2023, 13, 6603. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; You, J.; Wang, Y.; Long, Y.; Wang, S.; Pan, F.; Yu, Z. Biocontrol efficacy of Bacillus velezensis strain YS-AT-DS1 against the root-knot nematode Meloidogyne incognita in tomato plants. Front. Microbiol. 2022, 13, 1035748. [Google Scholar] [CrossRef]
- Tian, X.-l.; Zhao, X.-m.; Zhao, S.-y.; Zhao, J.-l.; Mao, Z.-c. The biocontrol functions of Bacillus velezensis strain Bv-25 against Meloidogyne incognita. Front. Microbiol. 2022, 13, 843041. [Google Scholar] [CrossRef]
- Mwangi, M.; Muiru, W.; Narla, R.; Kimenju, J.; Kariuki, G. Management of Fusarium oxysporum f. sp. lycopersici and root-knot nematode disease complex in tomato by use of antagonistic fungi, plant resistance and neem. Biocontrol Sci. Technol. 2019, 29, 229–238. [Google Scholar] [CrossRef]
- Sreenayana, B.; Vinodkumar, S.; Nakkeeran, S.; Muthulakshmi, P.; Poornima, K. Multitudinous potential of Trichoderma species in imparting resistance against F. oxysporum f. sp. cucumerinum and Meloidogyne incognita disease complex. J. Plant Growth Regul. 2022, 41, 1187–1206. [Google Scholar] [CrossRef]
- Dababat, A.; Sikora, R. Importance of application time and inoculum density of Fusarium oxysporum 162 for biological control of Meloidogyne incognita on tomato. Nematropica 2007, 37, 267–276. [Google Scholar]
- Lee, S.; Kim, Y.; Kim, J.M.; Chu, B.; Joa, J.-H.; Sang, M.K.; Song, J.; Weon, H.-Y. A preliminary examination of bacterial, archaeal, and fungal communities inhabiting different rhizocompartments of tomato plants under real-world environments. Sci. Rep. 2019, 9, 9300. [Google Scholar] [CrossRef]
- French, E.; Kaplan, I.; Iyer-Pascuzzi, A.; Nakatsu, C.H.; Enders, L. Emerging strategies for precision microbiome management in diverse agroecosystems. Nat. Plants 2021, 7, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Servat, S.; Pinyol-Escala, L.; Daura-Pich, O.; Almazán, M.; Hernández, I.; López-García, B.; Fernández, C. Characterization of Lysobacter enzymogenes B25, a potential biological control agent of plant-parasitic nematodes, and its mode of action. AIMS Microbiol. 2023, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Topalović, O.; Hussain, M.; Heuer, H. Plants and associated soil microbiota cooperatively suppress plant-parasitic nematodes. Front. Microbiol. 2020, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Topalović, O.; Vestergård, M. Can microorganisms assist the survival and parasitism of plant-parasitic nematodes? Trends Parasitol. 2021, 37, 947–958. [Google Scholar] [CrossRef]
- Masson, A.-S.; Ho Bich, H.; Simonin, M.; Nguyen Thi, H.; Czernic, P.; Moulin, L.; Bellafiore, S. Deep modifications of the microbiome of rice roots infected by the parasitic nematode Meloidogyne graminicola in highly infested fields in Vietnam. FEMS Microbiol. Ecol. 2020, 96, fiaa099. [Google Scholar] [CrossRef] [PubMed]
- Lamelas, A.; Desgarennes, D.; López-Lima, D.; Villain, L.; Alonso-Sánchez, A.; Artacho, A.; Latorre, A.; Moya, A.; Carrion, G. The bacterial microbiome of Meloidogyne-based disease complex in coffee and tomato. Front. Plant Sci. 2020, 11, 136. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.; Zhao, J.; Xun, W.; Li, R.; Zhang, R.; Wu, H.; Shen, Q. Different continuous cropping spans significantly affect microbial community membership and structure in a vanilla-grown soil as revealed by deep pyrosequencing. Microb. Ecol. 2015, 70, 209–218. [Google Scholar] [CrossRef]
- Parsa, S.; Ortiz, V.; Gómez-Jiménez, M.I.; Kramer, M.; Vega, F.E. Root environment is a key determinant of fungal entomopathogen endophytism following seed treatment in the common bean, Phaseolus vulgaris. Biol. Control 2018, 116, 74–81. [Google Scholar] [CrossRef]
- Jousset, A.; Becker, J.; Chatterjee, S.; Karlovsky, P.; Scheu, S.; Eisenhauer, N. Biodiversity and species identity shape the antifungal activity of bacterial communities. Ecology 2014, 95, 1184–1190. [Google Scholar] [CrossRef]
- Zhou, D.; Feng, H.; Schuelke, T.; De Santiago, A.; Zhang, Q.; Zhang, J.; Luo, C.; Wei, L. Rhizosphere microbiomes from root knot nematode non-infested plants suppress nematode infection. Microb. Ecol. 2019, 78, 470–481. [Google Scholar] [CrossRef]
- Watson, T.T.; Strauss, S.L.; Desaeger, J.A. Identification and characterization of Javanese root-knot nematode (Meloidogyne javanica) suppressive soils in Florida. Appl. Soil Ecol. 2020, 154, 103597. [Google Scholar] [CrossRef]
- Cao, C.; Huang, J.; Ge, L.; Li, T.; Bu, Z.-J.; Wang, S.; Wang, Z.; Liu, Z.; Liu, S.; Wang, M. Does Shift in Vegetation Abundance After Nitrogen and Phosphorus Additions Play a Key Role in Regulating Fungal Community Structure in a Northern Peatland? Front. Microbiol. 2022, 13, 920382. [Google Scholar] [CrossRef] [PubMed]
- Prestt, A.J.; Roberts, M.R. Specific suppression of vascular wilt disease of onion is linked with fungal soil community structure. Rhizosphere 2023, 25, 100671. [Google Scholar] [CrossRef]
- Rashidifard, M.; Ashrafi, S.; Claassens, S.; Thünen, T.; Fourie, H. A pilot approach investigating the potential of crop rotation with sainfoin to reduce Meloidogyne enterolobii infection of maize under greenhouse conditions. Front. Plant Sci. 2021, 12, 676. [Google Scholar] [CrossRef]
- Adedayo, A.A.; Fadiji, A.E.; Babalola, O.O. Plant Health Status Affects the Functional Diversity of the Rhizosphere Microbiome Associated with Solanum lycopersicum. Front. Sustain. Food Syst. 2022, 6, 894312. [Google Scholar] [CrossRef]
- Wu, H.; Xiang, W.; Ouyang, S.; Forrester, D.I.; Zhou, B.; Chen, L.; Ge, T.; Lei, P.; Chen, L.; Zeng, Y. Linkage between tree species richness and soil microbial diversity improves phosphorus bioavailability. Funct. Ecol. 2019, 33, 1549–1560. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.-T.; Wang, W.-H.; Tsui, C.K.; Cai, L. Changes in bacterial and fungal microbiomes associated with tomatoes of healthy and infected by Fusarium oxysporum f. sp. lycopersici. Microb. Ecol. 2021, 81, 1004–1017. [Google Scholar] [CrossRef]
- Naylor, D.; McClure, R.; Jansson, J. Trends in microbial community composition and function by soil depth. Microorganisms 2022, 10, 540. [Google Scholar] [CrossRef]
- Cui, R.; Fan, C.; Sun, X. Isolation and characterisation of Aspergillus awamori BS05, a root-knot-nematode-trapping fungus. Biocontrol Sci. Technol. 2015, 25, 1233–1240. [Google Scholar] [CrossRef]
- Jang, J.Y.; Choi, Y.H.; Shin, T.S.; Kim, T.H.; Shin, K.-S.; Park, H.W.; Kim, Y.H.; Kim, H.; Choi, G.J.; Jang, K.S. Biological control of Meloidogyne incognita by Aspergillus niger F22 producing oxalic acid. PLoS ONE 2016, 11, e0156230. [Google Scholar] [CrossRef]
- Ying, L.; Zhong, D.; PENG, D.-l.; LIU, S.-m.; KONG, L.-a.; Huan, P.; Xiang, C.; LI, Z.-c.; HUANG, W.-k. Evaluation of the biocontrol potential of Aspergillus welwitschiae against the root-knot nematode Meloidogyne graminicola in rice (Oryza sativa L.). J. Integr. Agric. 2019, 18, 2561–2570. [Google Scholar]
- Xiang, C.; Liu, Y.; Liu, S.-M.; Huang, Y.-F.; Kong, L.-A.; Peng, H.; Liu, M.-Y.; Liu, J.; Peng, D.-L.; Huang, W.-K. αβ-Dehydrocurvularin isolated from the fungus Aspergillus welwitschiae effectively inhibited the behaviour and development of the root-knot nematode Meloidogyne graminicola in rice roots. BMC Microbiol. 2020, 20, 48. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Wang, D.; Li, B.; Maqsood, A.; Wu, H. Nematicidal evaluation and active compounds isolation of Aspergillus japonicus ZW1 against root-knot nematodes Meloidogyne incognita. Agronomy 2020, 10, 1222. [Google Scholar] [CrossRef]
- Shemshura, O.N.; Bekmakhanova, N.E.; Mazunina, M.N.; Meyer, S.L.; Rice, C.P.; Masler, E.P. Isolation and identification of nematode-antagonistic compounds from the fungus Aspergillus candidus. FEMS Microbiol. Lett. 2016, 363, fnw026. [Google Scholar] [CrossRef] [PubMed]
- Bardhan, P.; Gohain, M.; Daimary, N.; Kishor, S.; Chattopadhyay, P.; Gupta, K.; Chaliha, C.; Kalita, E.; Deka, D.; Mandal, M. Microbial lipids from cellulolytic oleaginous fungus Penicillium citrinum PKB20 as a potential feedstock for biodiesel production. Ann. Microbiol. 2019, 69, 1135–1146. [Google Scholar] [CrossRef]
- Baazeem, A.; Alorabi, M.; Darwesh, H.; Alotaibi, S.S.; El-Deen, A.N.; Iqbal, S.; Naqvi, S.A.H. Biological control of Root-knot nematode (Meloidogyne javanica) by potential antagonism of endophytic fungi isolated from Taify roses. J. King Saud Univ. -Sci. 2022, 34, 102329. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, D.; Ling, N.; Chen, F.; Fang, W.; Shen, Q. Bio-organic fertilizer application significantly reduces the Fusarium oxysporum population and alters the composition of fungi communities of watermelon Fusarium wilt rhizosphere soil. Biol. Fertil. Soils 2014, 50, 765–774. [Google Scholar] [CrossRef]
- Huang, X.; Liu, L.; Wen, T.; Zhu, R.; Zhang, J.; Cai, Z. Illumina MiSeq investigations on the changes of microbial community in the Fusarium oxysporum f. sp. cubense infected soil during and after reductive soil disinfestation. Microbiol. Res. 2015, 181, 33–42. [Google Scholar]
- Vujanovic, V.; Mavragani, D.; Hamel, C. Fungal communities associated with durum wheat production system: A characterization by growth stage, plant organ and preceding crop. Crop Prot. 2012, 37, 26–34. [Google Scholar] [CrossRef]
- Köhl, J.; Lombaers, C.; Moretti, A.; Bandyopadhyay, R.; Somma, S.; Kastelein, P. Analysis of microbial taxonomical groups present in maize stalks suppressive to colonization by toxigenic Fusarium spp.: A strategy for the identification of potential antagonists. Biol. Control 2015, 83, 20–28. [Google Scholar] [CrossRef]
- de Azevedo Silva, F.; de Oliveira Vieira, V.; Carrenho, R.; Rodrigues, V.B.; Junior, M.L.; da Silva, G.F.; Soares, M.A. Influence of the biocontrol agents Trichoderma spp. on the structure and functionality of the edaphic microbial community in common bean cultivars (Phaseolus vulgaris L.) inoculated with Sclerotinia sclerotiorum (Lib.) de Bary. Appl. Soil Ecol. 2021, 168, 104190. [Google Scholar] [CrossRef]
- Back, M.; Haydock, P.; Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathol. 2002, 51, 683–697. [Google Scholar] [CrossRef]
- Van Gundy, S.; Kirkpatrick, J.; Golden, J. The nature and role of metabolic leakage from root-knot nematode galls and infection by Rhizoctonia solani. J. Nematol. 1977, 9, 113. [Google Scholar] [PubMed]
- Passera, A.; Vacchini, V.; Cocetta, G.; Arpanahi, A.A.; Casati, P.; Ferrante, A.; Piazza, L. Towards nutrition-sensitive agriculture: An evaluation of biocontrol effects, nutritional value, and ecological impact of bacterial inoculants. Sci. Total Environ. 2020, 724, 138127. [Google Scholar] [CrossRef] [PubMed]
- Patil, J.; Yadav, S.; Kumar, A. Management of root-knot nematode, Meloidogyne incognita and soil borne fungus, Fusarium oxysporum in cucumber using three bioagents under polyhouse conditions. Saudi J. Biol. Sci. 2021, 28, 7006–7011. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Balodi, R. Bio-management of soil borne pathogens infesting cucumber (Cucumis sativus L.) under protected cultivation system. Biol. Control 2021, 157, 104569. [Google Scholar] [CrossRef]
- Naz, I.; Khan, R.A.A.; Masood, T.; Baig, A.; Siddique, I.; Haq, S. Biological control of root knot nematode, Meloidogyne incognita, in vitro, greenhouse and field in cucumber. Biol. Control 2021, 152, 104429. [Google Scholar] [CrossRef]
- Chaudhari, P. Role of microbial bio-products in sustainable agriculture. Int. J. Innov. Hortic. 2017, 6, 25–47. [Google Scholar]
- Popescu, S.C.; Tomaso-Peterson, M.; Wilkerson, T.; Bronzato-Badial, A.; Wesser, U.; Popescu, G.V. Metagenomic Analyses of the Soybean Root Mycobiome and Microbiome Reveal Signatures of the Healthy and Diseased Plants Affected by Taproot Decline. Microorganisms 2022, 10, 856. [Google Scholar] [CrossRef]
- Zhao, X.; Lin, C.; Tan, J.; Yang, P.; Wang, R.; Qi, G. Changes of rhizosphere microbiome and metabolites in Meloidogyne incognita infested soil. Plant Soil 2023, 483, 331–353. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamalanathan, V.; Sevugapperumal, N.; Nallusamy, S.; Ashraf, S.; Kailasam, K.; Afzal, M. Metagenomic Approach Deciphers the Role of Community Composition of Mycobiome Structured by Bacillus velezensis VB7 and Trichoderma koningiopsis TK in Tomato Rhizosphere to Suppress Root-Knot Nematode Infecting Tomato. Microorganisms 2023, 11, 2467. https://doi.org/10.3390/microorganisms11102467
Kamalanathan V, Sevugapperumal N, Nallusamy S, Ashraf S, Kailasam K, Afzal M. Metagenomic Approach Deciphers the Role of Community Composition of Mycobiome Structured by Bacillus velezensis VB7 and Trichoderma koningiopsis TK in Tomato Rhizosphere to Suppress Root-Knot Nematode Infecting Tomato. Microorganisms. 2023; 11(10):2467. https://doi.org/10.3390/microorganisms11102467
Chicago/Turabian StyleKamalanathan, Vinothini, Nakkeeran Sevugapperumal, Saranya Nallusamy, Suhail Ashraf, Kumanan Kailasam, and Mohd Afzal. 2023. "Metagenomic Approach Deciphers the Role of Community Composition of Mycobiome Structured by Bacillus velezensis VB7 and Trichoderma koningiopsis TK in Tomato Rhizosphere to Suppress Root-Knot Nematode Infecting Tomato" Microorganisms 11, no. 10: 2467. https://doi.org/10.3390/microorganisms11102467