Detection of Fungal Diseases in Lettuce by VIR-NIR Spectroscopy in Aquaponics

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
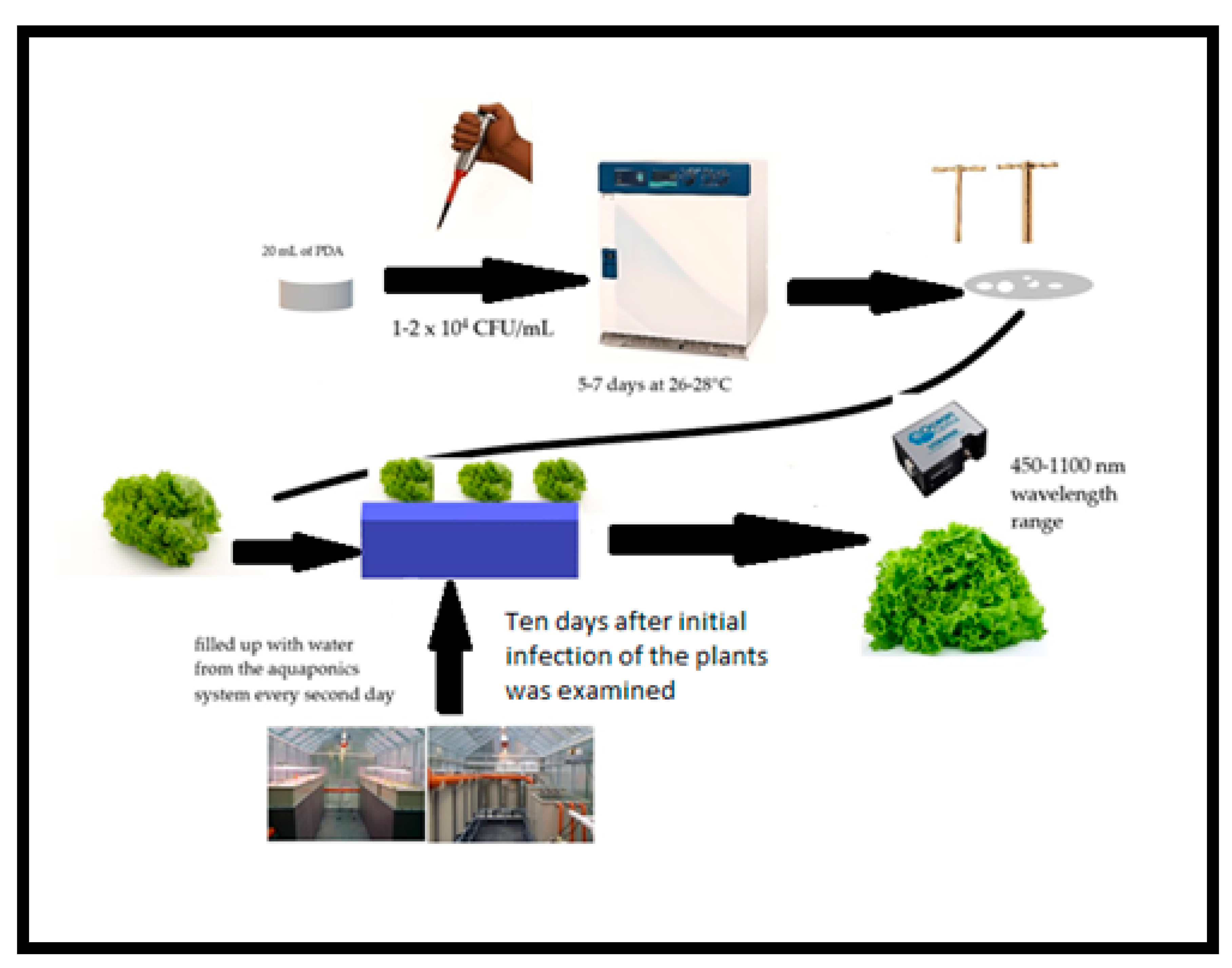
2.1. Experimental Design and Characterization of Aquaponic System and Vessels
2.2. Microorganisms Studied
2.3. NIR Spectroscopy Measurement
2.4. Statistical Data Analysis
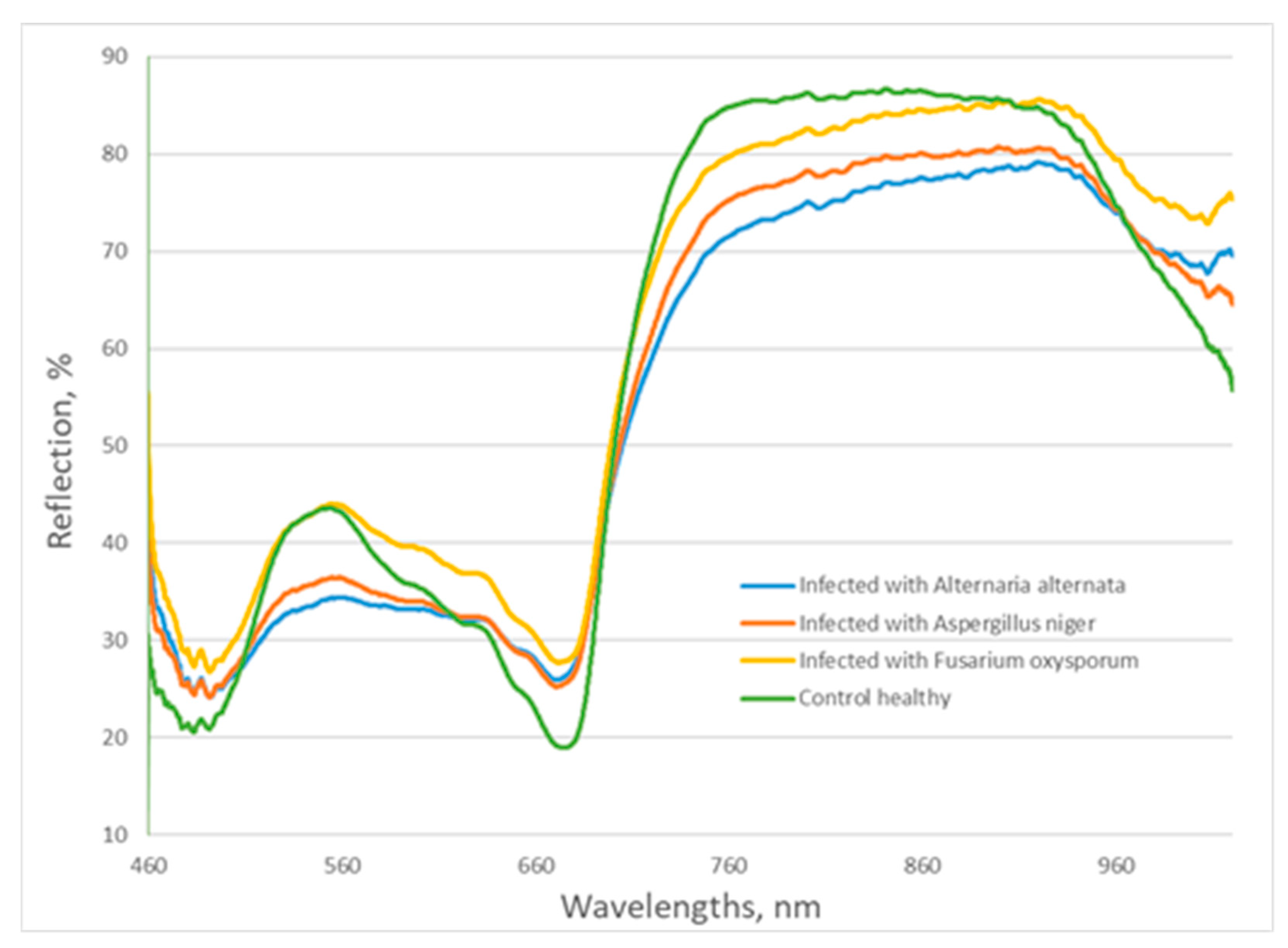
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Graber, A.; Junge, R. Aquaponic Systems: Nutrient recycling from fish wastewater by vegetable production. Desalination 2009, 246, 147–156. [Google Scholar] [CrossRef]
- Colt, J.; Schuur, A.; Weaver, D.; Semmens, K. Engineering design of aquaponics systems. Rev. Fish. Sci. Aquac. 2021, 30, 33–80. [Google Scholar] [CrossRef]
- Sirakov, I.; Lutz, M.; Graber, A.; Mathis, A.; Staykov, Y.; Smits, T.; Junge, R. Potential for combined biocontrol activity against fungal fish and plant pathogens by bacterial isolates from a model aquaponic system. Water 2016, 8, 518. [Google Scholar] [CrossRef]
- Suárez-Cáceres, G.; Pérez-Urrestarazu, L.; Avilés, M.; Borrero, C.; Eguíbar, J.; Fernández-Cabanás, V. Susceptibility to water-borne plant diseases of hydroponic vs. aquaponics systems. Aquaculture 2021, 544, 737093. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.; Pretorius, Z.; Hammond-Kosack, K. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [PubMed]
- Martinko, K.; Ivankovic, S.; Lazarevic, B.; Ðermic, E.; Ðermic, D. Control of early blight fungus (Alternaria alternata) in tomato by boric and phenylboronic acid. Antibiotics 2022, 11, 320. [Google Scholar] [CrossRef]
- Gravel, D.; Poisot, T.; Albouy, C.; Velez, L.; Mouillot, D. Inferring food web structure from predator–prey body size relationships. Methods Ecol. Evol. 2013, 4, 1083–1090. [Google Scholar] [CrossRef]
- Nemethy, S.; Bittsanszky, A.; Schmautz, Z.; Junge, R.; Komives, T. Protecting plants from pests and diseases in aquaponic systems. In Ecological Footprint in Central Europe, Multiauthorial Monograph; The University College of Tourism and Ecology Press: Sucha Beskidzka, Poland, 2016; pp. 166–179. [Google Scholar]
- Sirakov, I.; Velichkova, K.; Stoyanova, S.; Zhelyazkov, G.; Staykov, Y. The effect of diet supplemented with dandelion’s (Taraxacum officinale) extract on the productive and blood parameters of common carp (Cyprinus carpio L.), cultivated in the recirculation system. Maced. Vet. Rev. 2019, 42, 131–139. [Google Scholar] [CrossRef]
- Stouvenakers, G.; Massart, S.; Depireux, P.; Jijakli, M. Microbial origin of aquaponic water suppressiveness against Pythium aphanidermatum lettuce root rot disease. Microorganisms 2020, 8, 1683. [Google Scholar] [CrossRef]
- Rivas-García, S.; Bernal, J.; Bachiller-Corral, J. Rhabdomyolysis as the main manifestation of coronavirus disease 2019. Rheumatology 2020, 59, 2174–2176. [Google Scholar] [CrossRef]
- Mahlein, A.; Kuska, M.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral sensors and imaging technologies in phytopathology: State of the art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Kuska, M.; Bohnenkamp, D.; Brugger, A.; Alisaac, E.; Wahabzada, M.; Behmann, J.; Mahlein, A. Benefits of hyperspectral imaging for plant disease detection and plant protection: A technical perspective. J. Plant Dis. Prot. 2018, 125, 5–20. [Google Scholar] [CrossRef]
- Terentev, A.; Dolzhenko, V.; Fedotov, A.; Eremenko, D. Current State of Hyperspectral Remote Sensing for Early Plant Disease Detection: A Review. Sensors 2022, 22, 757. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Wan, L.; Li, H.; Li, C.; Wang, A.; Yang, Y.; Wang, P. Hyperspectral Sensing of Plant Diseases: Principle and Methods. Agronomy 2022, 12, 1451. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.; Pattey, E.; Zarco-Tejada, P.; Strachan, I. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Gitelson, A.; Viña, A.; Ciganda, V.; Rundquist, D.; Arkebauer, T. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, L08403. [Google Scholar] [CrossRef]
- Mahlein, A.; Rumpf, T.; Welke, P. Development of spectral indices for detecting and identifying plant diseases. Remote Sens. Environ. 2013, 128, 21–30. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.; Zur, Y.; Stark, R.; Gritz, U. Non-destructive and remote sensing techniques for estimation of vegetation status. In Proceedings of the Third European Conference on Precision Agriculture, Montpelier, France, 18–20 June 2001; Volume 1, pp. 301–306. [Google Scholar]
- Moshou, D.; Bravo, C.; Oberti, R.; West, J.; Bodria, L.; McCartney, A.; Ramon, H. Plant disease detection basedondata fusion of hyper-spectral and multi-spectral fluorescence imaging using Kohonen maps. Real-Time Imaging 2005, 1, 75–83. [Google Scholar] [CrossRef]
- Smith, R.; Adams, J.; Stephens, D.; Hick, P. Forecasting wheat yield in a Mediterranean-type environment from the NOAA satellite. Aust. J. Agric. Res. 1995, 46, 113–125. [Google Scholar] [CrossRef]
- Daughtry, C.; Walthall, C.; Kim, M.; Colstoun, E.; Murtrey, J. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Tucker, C.; Holben, B.; Elgin, J.; James, H.; McMurtrey, I.; James, E. Remote sensing of total dry-matter accumulation in winter wheat. Remote Sens. Environ. 1998, 11, 171–189. [Google Scholar] [CrossRef]
- Main, R.; Cho, M.; Moses, A.; Mathieu, R.; Kennedy, M.; Martha, M.; Ramoelo, A.; Koch, S. An investigation into robust spectral indices for leaf chlorophyll estimation. J. Photogramm. Remote Sens. 2011, 66, 751–761. [Google Scholar] [CrossRef]
- Brantley, S.; Zinnert, J.; Young, D. Application of hyperspectral vegetation indices to detect variations in high leaf area index temperate shrub thicket canopies. Remote Sens. Environ. 2011, 115, 514–523. [Google Scholar] [CrossRef]
- Datt, B. Remote sensing of chlorophyll a, chlorophyll b, chlorophyll a + b and total carotenoid content in eucalyptus leaves. Remote Sens. Environ. 1998, 66, 111–121. [Google Scholar] [CrossRef]
- Merzlyak, M.; Gitelson, A.; Chivkunova, O.; Rakitin, V. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Vogelmann, J.; Rock, B.; Moss, D. Red edge spectral measurements from sugar maple leaves. Int. J. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M. Remote estimation of chlorophyll content in higher plant leaves. Int. J. Remote Sens. 1997, 18, 2691–2697. [Google Scholar] [CrossRef]
- Broge, N.; Leblanc, E. Comparing prediction power and stability of broadband and hyperspectral vegetation indices for estimation of green leaf area index and canopy chlorophyll density. Remote Sens. Environ. 2001, 76, 156–172. [Google Scholar] [CrossRef]
- Wang, L.; Qu, J. NMDI: A normalized multi-band drought index for monitoring soil and vegetation moisture with satellite remote sensing. Geophys. Res. Lett. 2007, 34, L20405. [Google Scholar] [CrossRef]
- Miranda-Apodaca, J.; Artetxe, U.; Aguado, I.; Martin-Souto, L.; Ramirez-Garcia, A.; Lacuesta, M.; Becerril, J.; Estonba, A.; Ortiz-Barredo, A.; Hernández, A.; et al. Stress response to climate change and postharvest handling in two differently pigmented lettuce genotypes: Impact on Alternaria alternata invasion and mycotoxin production. Plants 2023, 12, 1304. [Google Scholar] [CrossRef] [PubMed]
- Yahaya, S.; Fagwalawa, L.; Ali, M.; Lawan, M.; Mahmud, S. Isolation and identification of pathogenic fungi causing deterioration of lettuce plant (Lactuca sativa). A case study of Yankaba and Sharada vegetables markets. J. Plant Sci. Res. 2016, 3, 141. [Google Scholar]
- Mukhtar, Y.; Muhammad, M.; Zubairu, S.; Galalain, A.; Ahmad, U. Isolation, identification and pathogenicity of fungal organisms causing postharvest rot of sweet oranges, cucumber and lettuce in Sharada Market, Kano State-Nigeria. Asian J. Med. Biol. Res. 2019, 5, 286–291. [Google Scholar] [CrossRef]
- Swarbrick, P.; Schulze-Lefert, P.; Scholes, J. Metabolic consequences of susceptibility and resistance in barley leaves challenged with powdery mildew. Plant Cell Environ. 2006, 29, 1061–1076. [Google Scholar] [CrossRef]
- Berger, S.; Benediktyová, Z.; Matous, K.; Bonfig, K.; Mueller, M.; Nedbal, L.; Roitsch, T. Visualization of dynamics of plant-pathogen interaction by novel combination of chlorophyll fluorescence imaging and statistical analysis: Differential effects of virulent and avirulent strains of P. syringae and of oxylipins on A. thaliana. J. Exp. Bot. 2007, 58, 797–806. [Google Scholar] [CrossRef]
- Zarger, M.; Rizvi, S.; Bagga, P. Biochemical response of mango (Mangifera indica) to the infection of leaf spot pathogen (Alternaria alternata). J. Environ. Biol. 2014, 35, 529–534. [Google Scholar]
- Tiwari, J.; Upadhyay, D. Correlation and path-coefficient studies in tomato (Lycopersicon esculentum Mill.). J. Agri. Sci. 2011, 2, 63–68. [Google Scholar]
- Berger, S.; Sinha, A.; Roitsch, T. Plant physiology meets phytopathology: Plant primary metabolism and plant–pathogen interactions. J. Exp. Bot. 2007, 58, 4019–4026. [Google Scholar] [CrossRef]
- Jackson, R.; Huete, A. Interpreting vegetation indices. Prev. Vet. Med. 1991, 11, 185–200. [Google Scholar] [CrossRef]
- Jones, H.; Vaughan, R. Remote Sensing of Vegetation; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Penuelas, J.; Frederic, B.; Filella, I. Semi-Empirical Indices to Assess Carotenoids/Chlorophyll-a Ratio from Leaf Spectral Reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Gamon, J.; Penuelas, J.; Field, C. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Agrios, G. Antifungal Activity of Selected Medicinal Plant Extract on Fusarium oxysporum Schlechtthe Causal Agent of Fusarium Wilt Disease in Tomato. Plant Pathol. 2005, 26–27, 398–401. [Google Scholar]
- Zarger, S.; Rizvi, G.; Parashar, R.; Paijwar, M. Impact of Alternaria alternata on organic components of mango leaves. Mycopath 2014, 12, 129–131. [Google Scholar]
- Zarco-Tejada, P.; González-Dugo, V.; Berni, J. Fluorescence, temperature and narrow-band indices acquired from a UAV platform for water stress detection using a micro-hyperspectral imager and a thermal camera. Remote Sens. Environ. 2012, 117, 322–337. [Google Scholar] [CrossRef]
- Furbank, R.; Tester, M. Phenomics–technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef]
- Thenkabail, P.; Lyon, J.; Huete, A. Hyperspectral Remote sensing of Vegetation; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Kira, K.; Rendell, L. A practical approach to feature selection. In Machine Learning Proceedings; Morgan Kaufmann: Burlington, MA, USA, 1992; pp. 249–256. [Google Scholar]



{kind=link}
{kind=link}
| Index | Definition | References |
|---|---|---|
| CARI Chlorophyll Absorption in Reflectance Index | Haboudane et al. [17] | |
| Cl green Clorophyll Index | Gitelson et al. [18] | |
| Cl red edge Clorophyll Index at red edge | Gitelson et al. [18] | |
| CLSI Cercospora leaf spot index | Mahlein et al. [19] | |
| CRI Carotenoids Reflectance index | Gitelson et al. [20] | |
| fD Index of disease | Moshou et al. [21] | |
| G Greennes index | Smith et al. [22] | |
| HI Healthy index | Mahlein et al. [19] | |
| MCARI Modified chlorophyll absorption in reflectance index | Daughtry et al. [23] | |
| mNDVI Modified normalized difference vegetation index | Tucker et al. [24] | |
| NDVI (1) Normalized difference vegetation index | Tucker et al. [24] | |
| NDVI (2) Normalized difference vegetation index | Main et al. [25] | |
| NDVI (3) Normalized difference vegetation index | Brantley et al. [26] | |
| PI Pigment index | Datt [27] | |
| PMI Powdery mildew index | Mahlein et al. [19] | |
| PRI Photochemical reflectance index | Sims and Gamon [15] | |
| PSRI Plant Senescence Reflectance Index | Merzlyak et al. [28] | |
| REI 1 Red Edge Index | Vogelmann et al. [29] | |
| REI 2 Red Edge Index | Vogelmann et al. [29] | |
| REI 3 Red Edge Index | Vogelmann et al. [29] | |
| SBRI Sugar beet rust index | Mahlein et al. [19] | |
| SR Simple ratio | Gitelson and Merzlyak [30] | |
| TVI triangular vegetation index | Broge and Leblanc [31] | |
| WBI Water band index | Wang and Qu [32] |
| x ± SD | |||||
|---|---|---|---|---|---|
| Vegetation Indices | Classes | R2 | |||
| Infected with Alternaria alternata (n = 8) | Infected with Aspergillus niger (n = 9) | Infected with Fusarium oxysporum (n = 7) | Uninfected (n = 5) | ||
| CARI | 1.975 ± 0.676 a | 2.356 ± 0.354 | 1.854 ± 0.251 c | 2.744 ± 0.535 a, c | 0.339 |
| Clgreen | 5.146 ± 2.228 | 3.909 ± 1.051 | 5.195 ± 1.260 | 3.457 ± 0.965 | 0.212 |
| Clred edge | 0.817 ± 0.208 | 0.682 ± 0.120 | 0.838 ± 0.010 c | 0.599 ± 0.133 c | 0.309 |
| CLSI | −10.957 ± 0.932 | −11.204 ± 0.393 | −11.408 ± 0.576 | −11.466 ± 0.419 | 0.099 |
| CRI | −1.661 ± 3.670 | −2.825 ± 0.550 | −4.117 ± 3.463 | −2.604 ± 1.110 | 0.116 |
| fD | 8.038 ± 30.228 | −2.741 ± 6.650 | 0.184 ± 2.914 | −2.362 ± 6.510 | 0.077 |
| G | 2.800 ± 0.706 | 3.291 ± 0.581 | 2.483 ± 0.783 c | 3.732 ± 0.580 c | 0.333 |
| HI | −2.168 ± 0.436 | −2.344 ± 0.174 | −2.184 ± 0.193 | −2.576 ± 0.307 | 0.225 |
| MCARI | 7.744 ± 3.630 a | 10.253 ± 2.614 | 6.681 ± 3.039 c | 13.665 ± 4.427 a, c | 0.378 |
| mNDVI | 0.486 ± 0.075 | 0.446 ± 0.040 | 0.497 ± 0.032 c | 0.412 ± 0.056 c | 0.285 |
| NDVI (1) | 0.921 ± 0.053 | 0.928 ± 0.017 | 0.913 ± 0.049 | 0.930 ± 0.025 | 0.032 |
| NDVI (2) | 0.844 ± 0.043 | 0.849 ± 0.012 | 0.837 ± 0.039 | 0.852 ± 0.018 | 0.032 |
| NDVI (3) | 0.874 ± 0.046 | 0.880 ± 0.014 | 0.867 ± 0.042 | 0.884 ± 0.021 | 0.038 |
| PI | 0.380 ± 0.103 | 0.315 ± 0.074 | 0.434 ± 0.119 c | 0.273 ± 0.041 c | 0.316 |
| PMI | −11.719 ± 4.662 | −10.247 ± 0.336 | −10.369 ± 0.292 | −10.669 ± 0.390 | 0.066 |
| PRI | −0.825 ± 0.877 | −0.436 ± 0.158 | −0.619 ± 0.276 | −0.405 ± 0.144 | 0.119 |
| PSRI | 0.207 ± 0.014 a | 0.215 ± 0.009 b | 0.220 ± 0.013 | 0.232 ± 0.009 a, b | 0.397 |
| REI 1 | 1.382 ± 0.090 | 1.324 ± 0.052 | 1.393 ± 0.042 c | 1.287 ± 0.059 c | 0.314 |
| REI 2 | 0.678 ± 0.090 a | 0.612 ± 0.063 | 0.682 ± 0.053 c | 0.557 ± 0.063 a, c | 0.352 |
| REI 3 | 0.336 ± 0.038 a | 0.308 ± 0.026 | 0.338 ± 0.020 c | 0.282 ± 0.029 a, c | 0.377 |
| SBRI | 1.383 ± 9.315 | 5.111 ± 16.361 | −4.683 ± 6.522 | 0.840 ± 3.952 | 0.110 |
| SR | 7.739 ± 2.622 | 6.532 ± 0.912 | 7.506 ± 1.413 | 5.999 ± 1.159 | 0.154 |
| TVI | 738.660 ± 46.810 | 762.604 ± 23.518 | 757.503 ± 19.201 | 785.200 ± 25.646 | 0.218 |
| WBI | 1.882 ± 0.063 a | 1.929 ± 0.044 b | 1.920 ± 0.058 c | 2.049 ± 0.046 a, b, c | 0.553 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirakov, I.; Velichkova, K.; Dinev, T.; Slavcheva-Sirakova, D.; Valkova, E.; Yorgov, D.; Veleva, P.; Atanasov, V.; Atanassova, S. Detection of Fungal Diseases in Lettuce by VIR-NIR Spectroscopy in Aquaponics. Microorganisms 2023, 11, 2348. https://doi.org/10.3390/microorganisms11092348
Sirakov I, Velichkova K, Dinev T, Slavcheva-Sirakova D, Valkova E, Yorgov D, Veleva P, Atanasov V, Atanassova S. Detection of Fungal Diseases in Lettuce by VIR-NIR Spectroscopy in Aquaponics. Microorganisms. 2023; 11(9):2348. https://doi.org/10.3390/microorganisms11092348
Chicago/Turabian StyleSirakov, Ivaylo, Katya Velichkova, Toncho Dinev, Desislava Slavcheva-Sirakova, Elica Valkova, Dimitar Yorgov, Petya Veleva, Vasil Atanasov, and Stefka Atanassova. 2023. "Detection of Fungal Diseases in Lettuce by VIR-NIR Spectroscopy in Aquaponics" Microorganisms 11, no. 9: 2348. https://doi.org/10.3390/microorganisms11092348