
Gastric Microbiota Gender Differences in Subjects with Healthy Stomachs and Autoimmune Atrophic Gastritis
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gastric Biopsy Sampling
2.2. Bacterial Microbiota Identification by 16SrRNA Gene Sequencing and Data Analysis
2.3. Statistical Analyses
3. Results
3.1. Gastric Bacterial Abundance
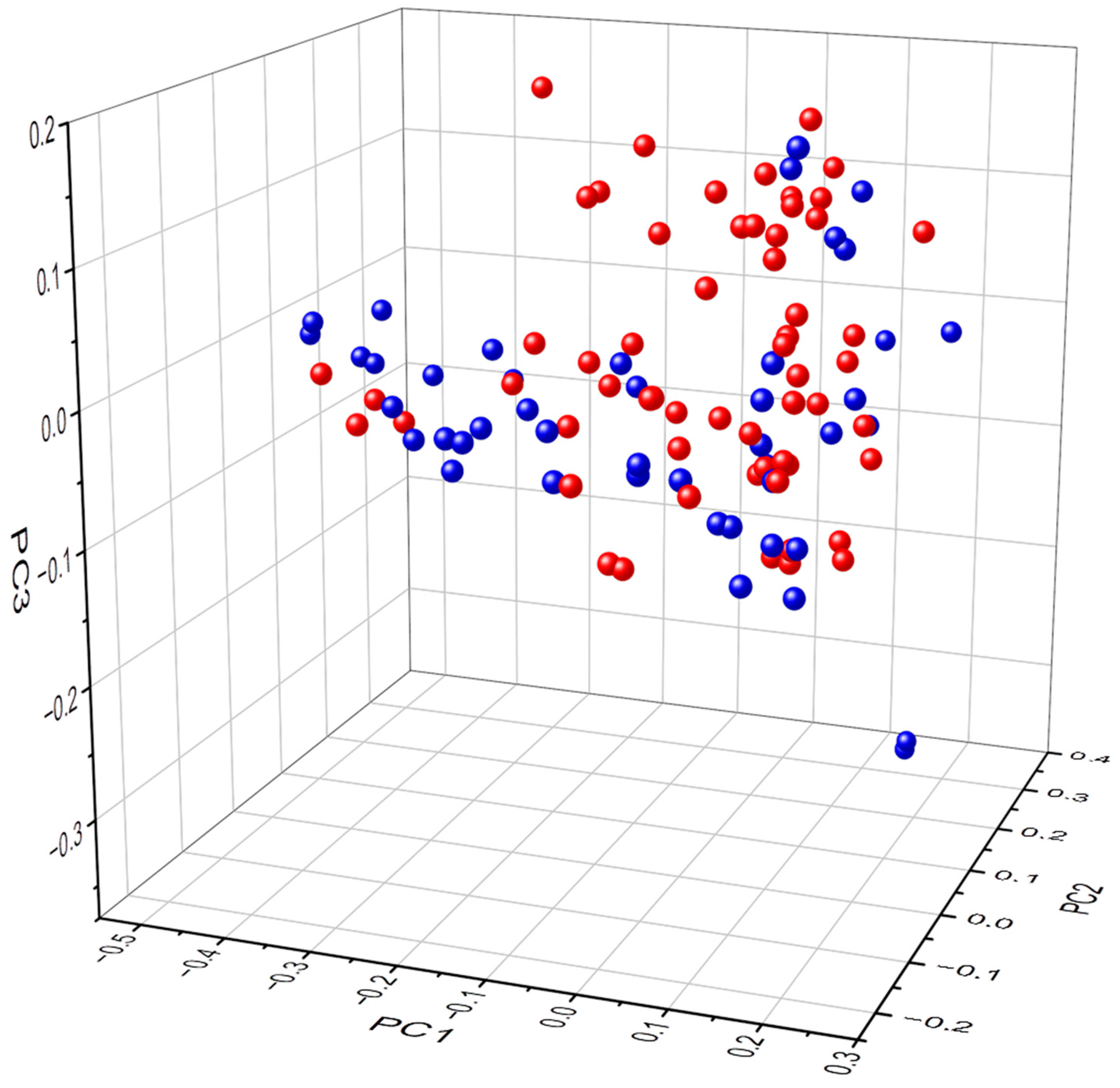
3.2. Biodiversity
3.3. Taxa Frequency at the Bacterial Phylum and Genus Level
3.4. Heatmap and LEfSe Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lenti, M.V.; Rugge, M.; Lahner, E.; Miceli, E.; Toh, B.H.; Genta, R.M.; De Block, C.; Hershko, C.; Di Sabatino, A. Autoimmune gastritis. Nat. Rev. Dis. Primers 2020, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Dottori, L.; Pivetta, G.; Annibale, B.; Lahner, E. Update on serum biomarkers in autoimmune atrophic gastritis. Clin. Chem. 2023, in press. [Google Scholar]
- Neumann, W.L.; Coss, E.; Rugge, M.; Genta, R.M. Autoimmune atrophic gastritis—Pathogenesis, pathology and management. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Dottori, L.; Pivetta, G.; Ligato, I.; Dilaghi, E.; Lahner, E. Pernicious Anemia: The Hematological Presentation of a Multifaceted Disorder Caused by Cobalamin Deficiency. Nutrients 2022, 14, 1672. [Google Scholar] [CrossRef]
- Miceli, E.; Vanoli, A.; Lenti, M.V.; Klersy, C.; Di Stefano, M.; Luinetti, O.; Caccia Dominioni, C.; Pisati, M.; Staiani, M.; Gentile, A.; et al. Natural history of autoimmune atrophic gastritis: A prospective, single centre, long-term experience. Aliment. Pharmacol. Ther. 2019, 50, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Lahner, E.; Galli, G.; Esposito, G.; Pilozzi, E.; Corleto, V.D.; Annibale, B. Updated features associated with type 1 gastric carcinoids patients: A single-center study. Scand. J. Gastroenterol. 2014, 49, 1447–1455. [Google Scholar] [CrossRef]
- Rugge, M.; Bricca, L.; Guzzinati, S.; Sacchi, D.; Pizzi, M.; Savarino, E.; Farinati, F.; Zorzi, M.; Fassan, M.; Dei Tos, A.P.; et al. Autoimmune gastritis: Long-term natural history in naïve Helicobacter pylori-negative patients. Gut 2023, 72, 30–38. [Google Scholar] [CrossRef]
- Esposito, G.; Dilaghi, E.; Cazzato, M.; Pilozzi, E.; Conti, L.; Carabotti, M.; Di Giulio, E.; Annibale, B.; Lahner, E. Endoscopic surveillance at 3 years after diagnosis, according to European guidelines, seems safe in patients with atrophic gastritis in a low-risk region. Dig. Liver Dis. 2021, 53, 467–473. [Google Scholar] [CrossRef]
- De Block, C.E.; De Leeuw, I.H.; Van Gaal, L.F. Autoimmune gastritis in type 1 diabetes: A clinically oriented review. J. Clin. Endocrinol. Metab. 2008, 93, 363–371. [Google Scholar] [CrossRef]
- Carabotti, M.; Lahner, E.; Esposito, G.; Sacchi, M.C.; Severi, C.; Annibale, B. Upper gastrointestinal symptoms in autoimmune gastritis: A cross-sectional study. Medicine 2017, 96, e5784. [Google Scholar] [CrossRef]
- Bouman, A.; Schipper, M.; Heineman, M.J.; Faas, M.M. Gender difference in the non-specific and specific immune response in humans. Am. J. Reprod. Immunol. 2004, 52, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Rizzetto, L.; Fava, F.; Tuohy, K.M.; Selmi, C. Connecting the immune system, systemic chronic inflammation and the gut microbiome: The role of sex. J. Autoimmun. 2018, 92, 12–34. [Google Scholar] [CrossRef]
- Conti, L.; Borro, M.; Milani, C.; Simmaco, M.; Esposito, G.; Canali, G.; Pilozzi, E.; Ventura, M.; Annibale, B.; Lahner, E. Gastric microbiota composition in patients with corpus atrophic gastritis. Dig. Liver Dis. 2021, 53, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Annibale, B.; Lahner, E. Autoimmune Gastritis and Gastric Microbiota. Microorganisms 2020, 8, 1827. [Google Scholar] [CrossRef] [PubMed]
- Parsons, B.N.; Ijaz, U.Z.; D’Amore, R.; Burkitt, M.D.; Eccles, R.; Lenzi, L.; Duckworth, C.A.; Moore, A.R.; Tiszlavicz, L.; Varro, A.; et al. Comparison of the human gastric microbiota in hypochlorhydric states arising as a result of Helicobacter pylori-induced atrophic gastritis, autoimmune atrophic gastritis and proton pump inhibitor use. PLOS Pathog. 2017, 13, e1006653. [Google Scholar] [CrossRef] [Green Version]
- Santos-Marcos, J.A.; Haro, C.; Vega-Rojas, A.; Alcala-Diaz, J.F.; Molina-Abril, H.; Leon-Acuña, A.; Lopez-Moreno, J.; Landa, B.B.; Tena-Sempere, M.; Perez-Martinez, P.; et al. Sex Differences in the Gut Microbiota as Potential Determinants of Gender Predisposition to Disease. Mol. Nutr. Food Res. 2019, 63, e1800870. [Google Scholar] [CrossRef]
- Dixon, M.F.; Genta, R.M.; Yardley, J.H.; Correa, P. Classification and grading of gastritis. The updated Sydney System. International Workshop on the Histopathology of Gastritis, Houston 1994. Am. J. Surg. Pathol. 1996, 20, 1161–1181. [Google Scholar] [CrossRef] [PubMed]
- Coati, I.; Fassan, M.; Farinati, F.; Graham, D.Y.; Genta, R.M.; Rugge, M. Autoimmune gastritis: Pathologist’s viewpoint. World J. Gastroenterol. 2015, 21, 12179–12189. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.C.; Piazuelo, M.B.; Kuipers, E.J.; Li, D. AGA Clinical Practice Update on the Diagnosis and Management of Atrophic Gastritis: Expert Review. Gastroenterology 2021, 161, 1325–1332.e7. [Google Scholar] [CrossRef]
- Park, S.Y.; Hwang, B.O.; Lim, M.; Ok, S.H.; Lee, S.K.; Chun, K.S.; Park, K.K.; Hu, Y.; Chung, W.Y.; Song, N.Y. Oral-Gut Microbiome Axis in Gastrointestinal Disease and Cancer. Cancers 2021, 13, 2124. [Google Scholar] [CrossRef]
- Di Giosia, P.; Stamerra, C.A.; Giorgini, P.; Jamialahamdi, T.; Butler, A.E.; Sahebkar, A. The role of nutrition in inflammaging. Ageing Res. Rev. 2022, 77, 101596. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.S.; Leys, E.J.; Bryk, J.M.; Martinez, F.J.; Moeschberger, M.L.; Griffen, A.L. Changes in periodontal health status are associated with bacterial community shifts as assessed by quantitative 16S cloning and sequencing. J. Clin. Microbiol. 2006, 44, 3665–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhou, M.; Salazar, C.R.; Hays, R.; Bedi, S.; Chen, Y.; Li, Y. Chronic Periodontal Disease, Periodontal Pathogen Colonization, and Increased Risk of Precancerous Gastric Lesions. J. Periodontol. 2017, 88, 1124–1134. [Google Scholar] [CrossRef]
- Gunathilake, M.; Lee, J.; Choi, I.J.; Kim, Y.I.; Yoon, J.; Sul, W.J.; Kim, J.F.; Kim, J. Alterations in Gastric Microbial Communities Are Associated with Risk of Gastric Cancer in a Korean Population: A Case-Control Study. Cancers 2020, 12, 2619. [Google Scholar] [CrossRef]
- Lahner, E.; Dilaghi, E.; Cingolani, S.; Pivetta, G.; Dottori, L.; Esposito, G.; Marzinotto, I.; Lampasona, V.; Buzzetti, R.; Annibale, B. Gender-sex differences in autoimmune atrophic gastritis. Transl. Res. 2022, 248, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Lenti, M.V.; Miceli, E.; Cococcia, S.; Klersy, C.; Staiani, M.; Guglielmi, F.; Giuffrida, P.; Vanoli, A.; Luinetti, O.; De Grazia, F.; et al. Determinants of diagnostic delay in autoimmune atrophic gastritis. Aliment. Pharmacol. Ther. 2019, 50, 167–175. [Google Scholar] [CrossRef]
- Cancer Today [Internet]. Available online: http://gco.iarc.fr/today/home (accessed on 21 January 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects with Healthy Stomachs n = 30 (57.7%) | Patients with AAG n = 22 (42.3%) | p | |
|---|---|---|---|
| Females/males, n (%) | 15 (50)/15 (50) | 17 (77.3)/5 (22.7) | Ns |
| Age, years, median (range) | 58 (18–83) | 64 (36–81) | 0.02 |
| Positivity to parietal cells antibodies | 0 | 17 (73.9) | <0.001 |
| Severe corpus atrophy | Na | 14 (60.9%) |
| Subjects with Healthy Stomachs | |||
| Reads | Females | Males | p |
| 30,000 | 867.20 (476.10–1365.60) | 709.40 (243.60–1250.60) | 0.05 |
| 26,666 | 835.20 (474.10–1334.00) | 639.20 (239.70–1240.30) | 0.04 |
| 23,333 | 796.40 (287.00–1308.30) | 533.85 (162.00–1220.80) | 0.06 |
| 20,000 | 766.60 (286.60–1264.50) | 486.20 (161.80–1196.90) | 0.04 |
| 16,667 | 752.40 (285.80–1213.30) | 485.00 (161.50–1172.40) | 0.04 |
| 13,333 | 704.65 (284.10–1145.00) | 470.40 (159.60–1116.60) | 0.03 |
| 10,000 | 668.00 (279.40–1062.90) | 452.20 (156.40–1051.20) | 0.04 |
| 6667 | 619.65 (269.60–921.00) | 438.40 (149.00–936.70) | 0.05 |
| Patients with autoimmune atrophic gastritis | |||
| Reads | Females | Males | p |
| 30.000 | 503.15 (263.80–1251.30) | 524.50 (415.20–611.00) | 0.89 |
| 26,666 | 498.85 (263.60–1228.00) | 519.50 (366.80–603.70) | 0.80 |
| 23,333 | 487.10 (260.90–1198.80) | 512.70 (365.50–598.00) | 0.94 |
| 20,000 | 482.25 (257.90–1152.70) | 482.45 (203.00–587.70) | 0.54 |
| 16,667 | 476.85 (256.40–1108.60) | 447.60 (161.00–578. 40) | 0.27 |
| 13,333 | 467.20 (174.60–1042.20) | 433.00 (160.60–560.50) | 0.32 |
| 10,000 | 456.30 (173.00–953.90) | 391.05 (140.00–539.70) | 0.14 |
| 6667 | 421.25 (169.60–837.30) | 361.70 (139.00–501.60) | 0.10 |
| Females | Males | p-Value | |
|---|---|---|---|
| Healthy stomachs | 3169 | 3414 | 0.003 |
| Autoimmune atrophic gastritis | 3505 | 2345 | <0.0001 |
| p-value | p = 0.002 | p < 0.001 |
| Taxonomy Phyla | HEALTHY STOMACHS n = 30 | p-Value | AAG n = 22 | p-Value | ||
|---|---|---|---|---|---|---|
| Females Median | Males Median | Females Median | Males Median | |||
| Firmicutes | 22.67 | 15.83 | 0.5677 | 27.46 | 33.11 | 0.0155 |
| Proteobacteria | 18.70 | 26.41 | 0.8489 | 19.13 | 19.32 | 0.1425 |
| Bacteroidetes | 10.55 | 10.46 | 0.1824 | 5.17 | 2.36 | 0.4042 |
| Actinobacteria | 2.97 | 1.75 | 0.6424 | 2.19 | 2.10 | 0.0042 |
| Fusobacteria | 0.19 | 1.31 | 0.0124 | 0.58 | 0.18 | 0.4634 |
| Taxonomy Genus | HEALTHY STOMACHS n = 30 | p-Value | AAG n = 22 | p-Value | |||
|---|---|---|---|---|---|---|---|
| Females Mean | Males Mean | Females Median | Males Median | ||||
| Streptococcus | 11.26 | 7.88 | 0.0358 | Streptococcus | 15.49 | 15.94 | 0.7751 |
| Prevotella 7 | 3.24 | 2.23 | 0.0269 | Haemophilus | 2.81 | 0.59 | 0.0223 |
| Neisseria | 1.59 | 1.36 | 0.5675 | Prevotella 7 | 1.21 | 0.85 | 0.4899 |
| Granulicatella | 1.03 | 0.73 | 0.0020 | Neisseria | 1.17 | 0.43 | 0.0156 |
| Gemella | 1.16 | 0.37 | 0.0057 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pivetta, G.; Dottori, L.; Fontana, F.; Cingolani, S.; Ligato, I.; Dilaghi, E.; Milani, C.; Ventura, M.; Borro, M.; Esposito, G.; et al. Gastric Microbiota Gender Differences in Subjects with Healthy Stomachs and Autoimmune Atrophic Gastritis. Microorganisms 2023, 11, 1938. https://doi.org/10.3390/microorganisms11081938
Pivetta G, Dottori L, Fontana F, Cingolani S, Ligato I, Dilaghi E, Milani C, Ventura M, Borro M, Esposito G, et al. Gastric Microbiota Gender Differences in Subjects with Healthy Stomachs and Autoimmune Atrophic Gastritis. Microorganisms. 2023; 11(8):1938. https://doi.org/10.3390/microorganisms11081938
Chicago/Turabian StylePivetta, Giulia, Ludovica Dottori, Federico Fontana, Sophia Cingolani, Irene Ligato, Emanuele Dilaghi, Christian Milani, Marco Ventura, Marina Borro, Gianluca Esposito, and et al. 2023. "Gastric Microbiota Gender Differences in Subjects with Healthy Stomachs and Autoimmune Atrophic Gastritis" Microorganisms 11, no. 8: 1938. https://doi.org/10.3390/microorganisms11081938