
Recent Trends and Applications of Nanoencapsulated Bacteriocins against Microbes in Food Quality and Safety

 ,
,  , , ,
, , ,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
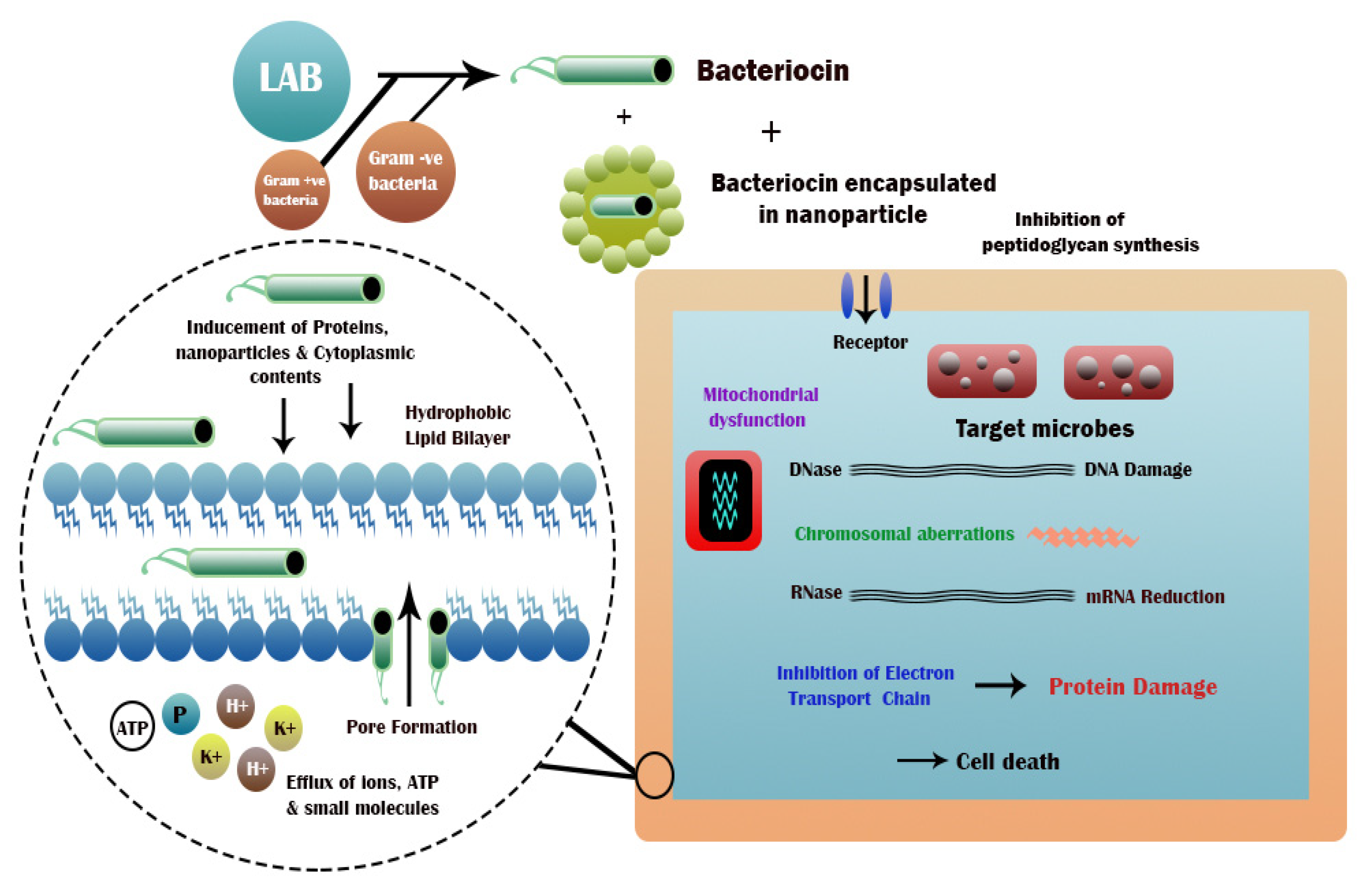
2. General Action Mechanism of Bacteriocins
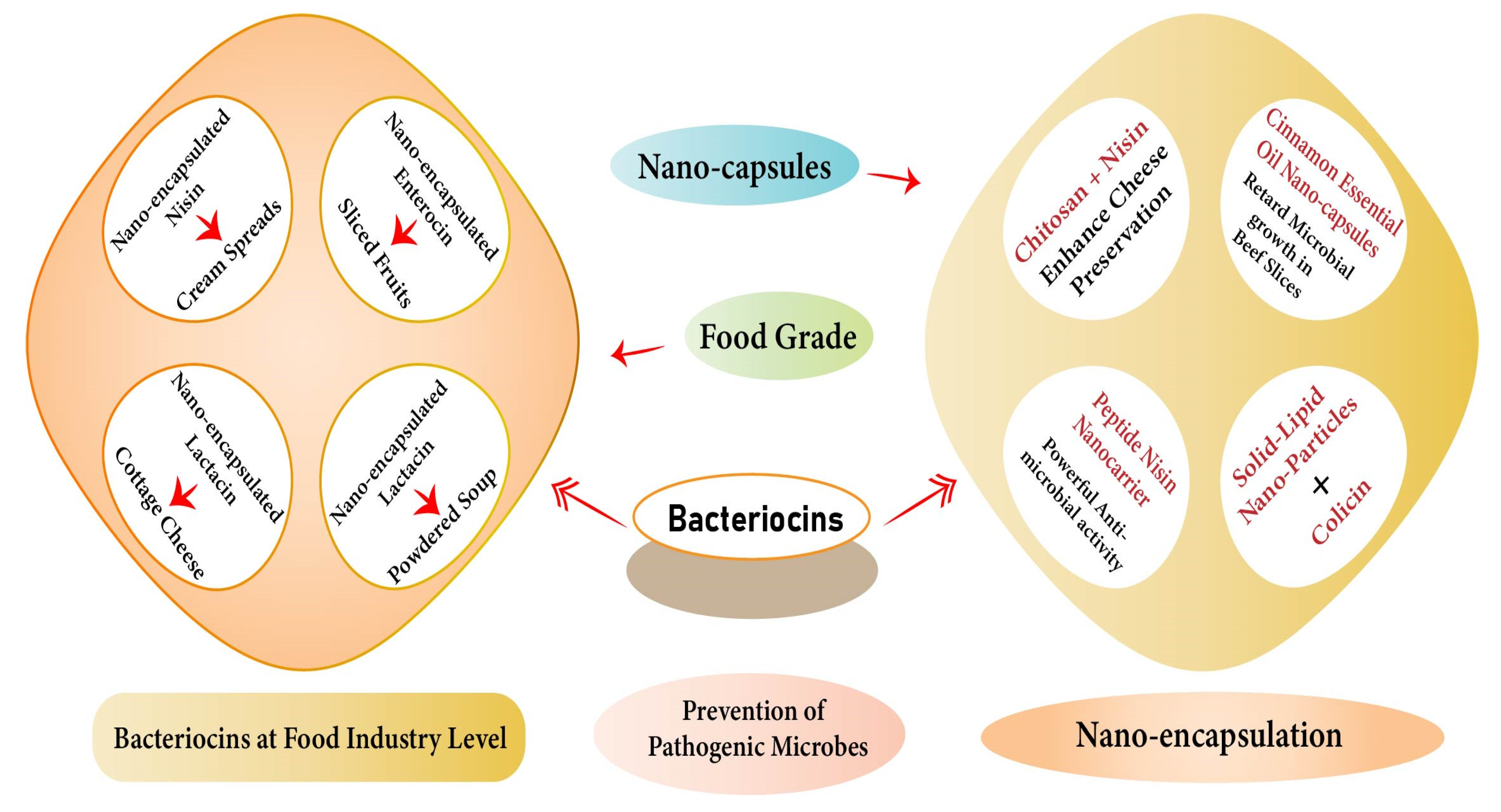
3. Formulation of Nano-Encapsulated Bacteriocins
3.1. Chitosan-Encapsulated Nisin
3.2. Liposome-Encapsulated Pediocin
3.3. Solid Lipid Nano-Encapsulated Colicin
3.4. Nano-Encapsulated Microcins
3.5. Nano-Encapsulated Lantibiotics
3.6. Nano-Encapsulated Peptides

| LAB Bacteriocins | Purification of Bacteriocins | Nanoparticles | Applied Encapsulation Technique | Nanoencapsulation Conditions | Effective against Bacteria | Effect of Nanoformulation and Applied Technique | References |
|---|---|---|---|---|---|---|---|
| Nisin | Milk fat globule membrane (MFGM) phospholipids-based nanostructures | Rhamnolipids (RLs) | Ultrasonication-assisted self-assembly method | Sonicated for 30 min | Prevent Escherichia coli and Listeria monocytogenes | Enhanced cheese preservation to prevent the foodborne pathogens | [81] |
| Nisin | ------------ | Cinnamon essential oil nanocapsules (CEO-NPs) | ---------- | ------------ | Retard the microbial growth and decreased lipid oxidation | Increased storage of beef slices for 15 days | [82] |
| Peptide nisin | Dissolution of 3 mg/mL nisin in acetic acid solution at pH 4.0 to obtain a stock solution of nisin | Nanocarrier based on polysaccharide with curcumin | Ultrafiltration tubes, Magnetic stirring | Stirring for 30 min at 25 °C with final 4.0 pH | 14.00 mm on B. subtilis and 12.97 mm on L. monocytogenes | Nanocarriers were fabricated to provide multifunctional potential in the food and show powerful antimicrobial activity | [83] |
| Nisin | 1 mg/mL nisin form Film-forming solutions (FFS) at 600 rpm by stirring at room temperature for 40 min | Nano-rhamnosomes | Field Emission Scanning Electron Microscope (FE-SEM) | 10 kV accelerated voltage | Inhibit the E. coli and L. monocytogenes growth | Prolonged bioactive preservation of food by broad-spectrum antimicrobial activity to combat Gram-negative and Gram-positive foodborne pathogens | [84] |
| Nisin | Digestion of protein with trypsin 20 μg and incubation at 37 °C, dried for 16 h, and storage temp is −20 °C | Phosphatidylcholine liposomes | Sonication and Thin-film hydration method | Dried through thin film for 24 h in a desiccator and nisin solution in phosphate buffer (10 mm) addition at 100 μg/mL and sonicated for 3 min at 55 kHz | Liposome-encapsulated nisin decreases stresses and lowers the occurrence of L. monocytogenes | Liposome encapsulation might be an effective approach to prevent nisin resistance | [85] |
| Nisin | Nisin was dissolved in the water phase with soy oil (20 mL) and gelatin (1%) | Polyacrylate Sodium (PAAS) and polyvinyl alcohol (PVA) | Sonication, Response Surface Methodology, Electrospinning | Ultra-sonication time (15 min) and 15%, centrifugation (6000 rpm) at 4 °C for 5 min after freeze-drying for 48 h, at −50 °C | Staphylococcus aureus and Escherichia coli growth was prevented for 16 days | Nanofiber can potentially retard food microorganisms’ activity in food and prolong the strawberry’s shelf life | [86] |
| Plantaricin | Sodium sulfate method | Silver nanoparticles | ------------ | ------------ | Showed inhibitory activity towards Listeria monocytogenes | The stability period got increased from 5 days to 60 days | [87] |
| Bacteriocin | ----------- | Au-zein-based nanomats | Electrospinning method | Samples were stored at (4 ± 1 °C) | ~1 log CFU/g reduction of bacteria | Reduced the growth of mesophilic aerobic bacteria in skinless fish fillets | [88] |
| Nisin | ---------- | Nanofibers (NP) with polyethylene (PE) packs | Electrospinning method | 1.2 mL/h, 8 cm, and 20 kV | Total mesophilic bacteria from 5.03 to 3.52 log CFU/g, Lactic acid bacteria from 3.22 to 2.02 log CFU/g | Prevention of off-odor and reduction of microbial growth in rainbow trout fillets | [89] |
4. Advantage of Antimicrobial Peptides at the Food Industry Scale
5. Conclusions
6. Future Recommendations
Funding
Acknowledgments
Conflicts of Interest
References
- García, P.; Rodríguez, L.; Rodríguez, A.; Martínez, B. Food biopreservation: Promising strategies using bacteriocins, bacteriophages and endolysins. Trends Food Sci. Technol. 2010, 21, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Chowdhuri, A.; Iqbal, A.; Giasuddin, M.; Bhuiyan, A.A. Study on Isolation and Identification of Salmonella and Escherichia coli from Different Poultry Feeds of Savar Region of Dhaka, Bangladesh. J. Sci. Res. 2011, 3, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Li, P.; Zhao, F.; Zhang, T.; Shan, Y.; Wang, X.; Liu, B.; Chen, Y.; Zhao, X.; Lü, X. Current status and potentiality of class II bacteriocins from lactic acid bacteria: Structure, mode of action and applications in the food industry. Trends Food Sci. Technol. 2022, 120, 387–401. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing Innate Immunity For Food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Fidan, H.; Esatbeyoglu, T.; Simat, V.; Trif, M.; Tabanelli, G.; Kostka, T.; Montanari, C.; Ibrahim, S.A.; Özogul, F. Recent developments of lactic acid bacteria and their metabolites on foodborne pathogens and spoilage bacteria: Facts and gaps. Food Biosci. 2022, 47, 101741. [Google Scholar] [CrossRef]
- Papagianni, M. Ribosomally synthesized peptides with antimicrobial properties: Biosynthesis, structure, function, and applications. Biotechnol. Adv. 2003, 21, 465–499. [Google Scholar] [CrossRef]
- De Vuyst, L.; Leroy, F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- Cui, Y.; Luo, L.; Yi, X.; Lu, Y.; Yanglei, Y.; Liu, B.; Zhou, Y.; Lü, X. Mining, heterologous expression, purification, antibactericidal mechanism, and application of bacteriocins: A review. Compr. Rev. Food Sci. Food Saf. 2020, 20, 863–899. [Google Scholar] [CrossRef] [PubMed]
- Al-holy, M.A.; Al-nabulsi, A.; Osaili, T.M.; Ayyash, M.M.; Shaker, R.R. Inactivation of Listeria innocua in brined white cheese by a combination of nisin and heat. Food Control 2012, 23, 48–53. [Google Scholar] [CrossRef]
- Pimentel-filho, N.D.J.; Mantovani, C.; De Carvalho, F.; Dias, R.S.; Vanetti, M.C.D. Original article Efficacy of bovicin HC5 and nisin combination against Listeria monocytogenes and Staphylococcus aureus in fresh cheese. Int. J. Food Sci. Technol. 2014, 49, 416–422. [Google Scholar] [CrossRef]
- Pei, J.; Yue, T.; Yuan, Y.; Dai, L. Activity of paracin C from lactic acid bacteria against Alicyclobacillus in apple juice: Application of a novelty bacteriocin. J. Food Safety 2017, 37, e12350. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Ross, R.P.; Stanton, C. Characterization and Application of Antilisterial Enterocins on Model Fresh Cheese. J. Food Prot. 2017, 80, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Woraprayote, W.; Pumpuang, L.; Tosukhowong, A.; Roytrakul, S.; Perez, R.H.; Zendo, T.; Sonomoto, K.; Benjakul, S.; Visessanguan, W. Two putatively novel bacteriocins active against Gram-negative food borne pathogens produced by Weissella hellenica BCC 7293. Food Control 2015, 55, 176–184. [Google Scholar] [CrossRef]
- Lu, Y.; Aizhan, R.; Yan, H.; Li, X.; Wang, X.; Yi, Y.; Shan, Y. Characterization, modes of action, and application of a novel broad-spectrum bacteriocin BM1300 produced by Lactobacillus crustorum MN047. Braz. J. Food Microbiol. 2020, 51, 2023–2048. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Fact. 2014, 13, S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, N.A.; Brandelli, A. Nanostructures for delivery of natural antimicrobials in food. Crit. Rev. Food Sci. Nutr. 2018, 58, 2202–2212. [Google Scholar] [CrossRef]
- Chandrakasan, G.; Rodríguez-Hernández, A.I.; del Rocío López-Cuellar, M.; Palma-Rodríguez, H.M.; Chavarría-Hernández, N. Bacteriocin encapsulation for food and pharmaceutical applications: Advances in the past 20 years. Biotechnol. Lett. 2019, 41, 453–469. [Google Scholar] [CrossRef]
- Fahim, H.A.; Khairalla, A.S.; El-Gendy, A.O. Nanotechnology: A valuable strategy to improve bacteriocin formulations. Front. Microbiol. 2016, 7, 1385. [Google Scholar] [CrossRef] [Green Version]
- Ge, J.; Kang, J.; Ping, W. Effect of Acetic Acid on Bacteriocin Production by Gram-Positive. J. Microbiol. Biotechnol. 2019, 29, 1341–1348. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M.T. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Abdulkarim, I.H.; Mohammed, S.S.D.; Orukotan, A.A. Gene Identification for Bacteriocin Production by Lactic Acid Bacteria Isolated from Selected Fermented Foods. Asian J. Biochem. Mol. Biol. 2020, 3, 1–12. [Google Scholar] [CrossRef]
- Brandelli, A. Nanostructures as Promising Tools for Delivery of Antimicrobial Peptides. Mini-Rev. Med. Chem. 2012, 12, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Naskar, A.; Kim, K. Potential Novel Food-Related and Biomedical Applications of Nanomaterials Combined with Bacteriocins. Pharmaceutics 2021, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Lelis, C.A.; De Carvalho, A.P.A.; Conte Junior, C.A. Systematic Review on Nanoencapsulation Natural Antimicrobials in Foods: In Vitro versus In Situ Evaluation, Mechanisms of Action and Implications on Physical-Chemical Quality. Int. J. Mol. Sci. 2021, 22, 12055. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Narmani, A.; Jafari, S.M. Dendrimers as efficient nanocarriers for the protection and delivery of bioactive phytochemicals. Adv. Colloid Interface Sci. 2020, 78, 102125. [Google Scholar] [CrossRef]
- Cintas, L.M.; Casaus, M.P.; Herranz, C.; Nes, I.F.; Hernández, P.E. Review: Bacteriocins of Lactic Acid Bacteria. Food Sci. Technol. Int. 2001, 7, 281–305. [Google Scholar] [CrossRef]
- Savadogo, A.; Ouattara, A.C.; Bassole, H.I.; Traore, S.A. Bacteriocins and lactic acid bacteria—A minireview. Afr. J. Biotechnol. 2006, 5, 678–683. [Google Scholar]
- Todorov, S.D.; Chikindas, M.L. Lactic Acid Bacteria Bacteriocins and their Impact on Human Health. In Lactic Acid Bacteria; CRC Press: Boca Raton, FL, USA, 2020; ISBN 9780429422591. [Google Scholar]
- Es, B.; Bhat, S.G. Marine Bacteriocins: A Review Marine Bacteriocins: A Review. J. Bacteriol. Mycol. 2016, 2, 00040. [Google Scholar] [CrossRef]
- Muñoz, M.; Jaramillo, D.; Melendez, P.; Alméciga-diaz, C.J. Native and Heterologous Production of Bacteriocins from Gram-Positive Microorganisms. Recent Pat. Biotechnol. 2011, 5, 199–211. [Google Scholar] [CrossRef]
- Atanaskovic, I.; Kleanthous, C. Tools and Approaches for Dissecting Protein Bacteriocin Import in Gram-Negative Bacteria. Front. Microbiol. 2019, 10, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebuffat, S. Bacteriocins from Gram-Negative Bacteria: A Classification? In Prokaryotic Antimicrobial Peptides; Springer: New York, NY, USA, 2011; pp. 55–72. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duche, D.; Kleanthous, C.; Lloube, R.; Postle, K.; Riley, M.; Slatin, S. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [Green Version]
- Sylvie, D.; Beeckman, A.; Vanrompay, D.C.G. Bacterial Secretion Systems with an Emphasis on the Chlamydial Type III Secretion System. Curr. Issues Mol. Biol. 2005, 12, 17–42. [Google Scholar]
- Gillor, O.; Kirkup, B.C.; Riley, M.A. Colicins and Microcins: The Next Generation Antimicrobials. Adv. Appl. Microbiol. 2004, 54, 129–146. [Google Scholar] [CrossRef]
- Yang, S.; Lin, C.; Sung, C.T.; Fang, J. Antibacterial activities of bacteriocins: Application in foods and pharmaceuticals. Front. Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Bharti, V.; Singh, S.; Jain, N.; Ahirwal, L. Bacteriocin: A novel approach for preservation of food. Int. J. Pharm. Pharm. Sci. 2015, 7, 20–29. [Google Scholar]
- Hildeliza, Q.B.; Chanona-pe, J.; Jose, L.S.M.; Gutie, G.F.; Jimene, A. Nanoencapsulation: A New Trend in Food Engineering Processing. Food Eng. Rev. 2010, 2, 39–50. [Google Scholar] [CrossRef]
- Divya, K.; Jisha, M.S. Chitosan nanoparticles preparation and applications. Environ. Chem. Lett. 2018, 16, 101–112. [Google Scholar] [CrossRef]
- Kailasapathy, K. Microencapsulation of Probiotic Bacteria: Technology and Potential Applications Further Reading. Curr. Issues Intest. Microbiol. 2002, 3, 39–48. [Google Scholar] [PubMed]
- Namasivayam, S.K.R.; Samrat, K.; Debnath, S.; Jayaprakash, C. Biocompatible chitosan nanoparticles incorporated bacteriocin (CSNps-B) preparation for the controlled release and improved anti-bacterial activity against food borne pathogenic bacteria Listeria monocytogenes. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 625–631. [Google Scholar]
- Juncioni, L.; Arauz, D.; Faustino, A.; Vessoni, T.C. Nisin biotechnological production and application: A review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Hsu, S.D.; Breukink, E.; Tischenko, E.; Lutters, M.A.G.; De Kruijff, B.; Kaptein, R.; Bonvin, A.M.J.J.; Van Nuland, N.A.J. The nisin—Lipid II complex reveals a pyrophosphate cage that provides a blueprint for novel antibiotics. Nat. Struct. Mol. Biol. 2004, 11, 963–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, N. Niosomes as nanocarrier systems. In Nanomaterials and Nanosystems for Biomedical Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007; ISBN 9781402062896. [Google Scholar]
- A Eldaly, E.; Mahmoud, F.A.; Abobakr, H.M. Preservative effect of chitosan coating on shelf life and sensory properties of chicken fillets during chilled storage. J. Nutr. Food Secur. 2018, 3, 139–148. [Google Scholar]
- da Silva Malheiros, P.; Daroit, D.J.; Brandelli, A. Food applications of liposome-encapsulated antimicrobial peptides. Trends Food Sci. Technol. 2010, 21, 284–292. [Google Scholar] [CrossRef]
- Bahrami, A.; Delshadi, R.; Jafari, S.M.; Williams, L. Nanoencapsulated nisin: An engineered natural antimicrobial system for the food industry. Trends Food Sci. Technol. 2019, 94, 20–31. [Google Scholar] [CrossRef]
- Riesute, R.; Salomskiene, J.; Moreno, D.S.; Gustiene, S. Effect of yeasts on food quality and safety and possibilities of their inhibition. Trends Food Sci. Technol. 2021, 108, 1–10. [Google Scholar] [CrossRef]
- Aditya, N.P.; Espinosa, Y.G.; Norton, I.T. Encapsulation systems for the delivery of hydrophilic nutraceuticals: Food application. Biotechnol. Adv. 2017, 35, 450–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, C.A.; Castro, M.P.; Rivas, F.P.; Schelegueda, L.I. Bacteriocins in food: Evaluation of the factors affecting their effectiveness. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex Research Center: Badajoz, Spain, 2013. [Google Scholar]
- Kemperman, R.; Kuipers, A.; Karsens, H.; Nauta, A.; Kuipers, O.; Kok, J. Identification and characterization of two novel clostridial bacteriocins, circularin A and closticin 574. Appl. Environ. Microbiol. 2003, 69, 1589–1597. [Google Scholar] [CrossRef] [Green Version]
- Laridi, R.; Kheadr, E.E.; Benech, R.O.; Vuillemard, J.C.; Lacroix, C.; Fliss, I. Liposome encapsulated nisin Z: Optimization, stability and release during milk fermentation. Int. Dairy J. 2003, 13, 325–336. [Google Scholar] [CrossRef]
- de Mello, M.B.; da Silva Malheiros, P.; Brandelli, A.; Pesce da Silveira, N.; Jantzen, M.M.; de Souza da Motta, A. Characterization and antilisterial effect of phosphatidylcholine nanovesicles containing the antimicrobial peptide pediocin. Probiotics Antimicrob. Proteins 2013, 5, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Bouaraba, L.; Maherania, B.; Kheirolomoomb, A.; Hasana, M.; Aliakbarianc, B.; Lindera, M.; Arab-Tehrany, E. Influence of lecithin—Lipid composition on physico-chemical properties of nanoliposomes loaded with a hydrophobic molecule. Colloids Surf. B Biointerfaces 2014, 115, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Narsaiah, K.; Robin, S.N.J.; Whilson, A.; Mandge, H.; Manikantan, M.R.; Malik, R.K.; Vij, S. Pediocin-Loaded Nanoliposomes and Hybrid Alginate—Nanoliposome Delivery Systems for Slow Release of Pediocin. J. Biosci. 2012, 3, 37–42. [Google Scholar] [CrossRef]
- Acedo, J.Z.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria Jeella. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-sieiro, P.; Montalbán-lópez, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, A.; Parshad, V.R. Novel trends to revolutionize preservation and packaging of fruits/fruit products: Microbiological and nanotechnological perspectives. Crit. Rev. Food Sci. Nutr. 2013, 55, 159–182, ISBN 9116124030. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Smith, K.; Brown, C.L.; Smith, K.; Mccaughey, L.; Walker, D. Colicin-like bacteriocins as novel therapeutic agents for the treatment of chronic biofilm-mediated infection. Prog. Chall. Bacteriocins Res. 2012, 40, 1549–1552. [Google Scholar] [CrossRef] [Green Version]
- Chavan, M.A.; Riley, M.A. Molecular Evolution of Bacteriocins in Gram-Negative Bacteria; Springer: Berlin/Heidelberg, Germany, 2007; pp. 19–43. [Google Scholar]
- Saha, S. A Tale of Two Tails: Characterization of R-Type and F-Type Pyocins of Pseudomonas aeruginosa. Ph.D. Thesis, University of Toronto, Toronto, ON, Canada, 2016. [Google Scholar]
- Duquesne, S.; Destoumieux-Garzón, D.; Peduzzi, J.; Rebuffat, S. Microcins, gene-encoded antibacterial peptides from enterobacteria. Nat. Prod. Rep. 2007, 24, 708–734. [Google Scholar] [CrossRef] [PubMed]
- Ponnachan, B.; Dash, N.; Jaybhay, P.; Tiwari, R. Reviewing isolation and production of probiotics. Int. J. Med. Biomed. Stud. 2021, 5. [Google Scholar] [CrossRef]
- Mousavi Khaneghah, A.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.A.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between probiotics and pathogenic microorganisms in hosts and foods: A review. Trends Food Sci. Technol. 2020, 95, 205–218. [Google Scholar] [CrossRef]
- Feng, L.; Mumper, R.J. A critical review of lipid-based nanoparticles for taxane delivery. Cancer Lett. 2012, 334, 157–175. [Google Scholar] [CrossRef] [Green Version]
- Micenková, L.; Bosák, J.; Kucera, J.; Hrala, M.; Dolejšová, T. Colicin Z, a structurally and functionally novel colicin type that selectively kills enteroinvasive Escherichia coli and Shigella strains. Nat. Res. 2019, 9, 11127. [Google Scholar] [CrossRef] [Green Version]
- Crone, W.J.K.; Vior, N.M.; Santos-aberturas, J.; Schmitz, L.G.; Leeper, F.J.; Truman, A.W. Zuschriften Dissecting Bottromycin Biosynthesis Using Comparative Untargeted Metabolomics Zuschriften. Angew. Chem. 2016, 128, 9791–9795. [Google Scholar] [CrossRef]
- Gomaa, A.I.; Martinent, C.; Hammami, R.; Fliss, I. Dual Coating of Liposomes as Encapsulating Matrix of Antimicrobial Peptides: Development and Characterization. Front. Chem. 2017, 5, 103. [Google Scholar] [CrossRef] [Green Version]
- Satchwell, K.L. Application of Microcin N and Tridecaptin A 1 to Control Bacterial Pathogens. Master’s Thesis, University of Alberta, Edmonton, AB, USA, 2016. [Google Scholar]
- Martin-serrano, Á.; Rafael, G.; Ortega, P.; De Mata, F.J. Nanosystems as Vehicles for the Delivery of Antimicrobial Peptides (AMPs). Pharmaceutics 2019, 11, 448. [Google Scholar] [CrossRef] [Green Version]
- Heng, N.C.; Wescombe, P.A.; Burton, J.P.; Jack, R.W.; Tagg, J.R. The diversity of bacteriocins in Gram-positive bacteria. In Bacteriocins; Springer: Berlin/Heidelberg, Germany, 2007; ISBN 9783540366034. [Google Scholar]
- Ansari, A. Bacteriocin from LAB for Medical and Health Applications. In Beneficial Microorganisms in Medical and Health Applications; Springer: Cham, Swizerland, 2015; pp. 199–221. [Google Scholar] [CrossRef]
- Wencewicz, T.A.; Miller, M.J. Sideromycins as Pathogen-Targeted Antibiotics. In Antibacterials; Springer: Berlin/Heidelberg, Germany, 2017; pp. 151–183. [Google Scholar] [CrossRef]
- Ekblad, B.; Kyriakou, P.K.; Oppega, C.; Nissen-meyer, J.; Kaznessis, Y.N. Structure—Function Analysis of the Two-Peptide Bacteriocin Plantaricin EF. Biochemistry 2016, 55, 5106–5116. [Google Scholar] [CrossRef] [PubMed]
- Dias, N.; Augusta, I.M. A Review “Clean Labeling”: Applications of Natural Ingredients in Bakery Products. J. Food Nutr. Res. 2018, 6, 285–294. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kristiansen, P.E.; Straume, D.; Jensen, M.S.; Aleksandrzak-piekarczyk, T.; Nes, I.F.; Diep, D.B.; Vederas, J.C. The Leaderless Bacteriocin Enterocin K1 Is Highly Potent against Enterococcus faecium: A Study on Structure, Target Spectrum and Receptor. Front. Microbiol. 2017, 8, 774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.C.; Kumari, A.; Yadav, R. Development of peptide and protein nanotherapeutics by nanoencapsulation and nanobioconjugation. Peptides 2011, 32, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. MDPI Antibiot. 2020, 9, 553. [Google Scholar] [CrossRef] [PubMed]
- Cabuk, B.; Okulklu, B.; Stanciuc, N.; Harsa, S.T. Nanoencapsulation of Biologically Active Peptides from Whey Proteins. J. Nutr. Health Food Sci. 2017, 2, 1–4. [Google Scholar]
- Mosquera, M.; Giménez, B.; Mallmann, I.; Ferreira, J.; Montero, P.; Gómez-guillén, M.C.; Brandelli, A. Nanoencapsulation of an active peptidic fraction from sea bream scales collagen. Food Chem. 2014, 156, 144–150. [Google Scholar] [CrossRef]
- Khalid, A.S.; Niaz, T.; Zarif, B.; Shabbir, S.; Noor, T.; Shahid, R.; Imran, M. Milk phospholipids-based nanostructures functionalized with rhamnolipids and bacteriocin: Intrinsic and synergistic antimicrobial activity for cheese preservation. Food Biosci. 2022, 47, 101442. [Google Scholar] [CrossRef]
- Zhang, M.; Luo, W.; Yang, K.; Li, C. Effects of sodium alginate edible coating with cinnamon essential oil nanocapsules and Nisin on quality and shelf life of beef slices during refrigeration. J. Food Prot. 2022, 85, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wu, Y.; Liu, C.; Zou, Y.; Huang, L.; Liang, Y.; Ren, J.; Liu, Y.; Lin, Q. Elaboration and characterization of curcumin-loaded soy soluble polysaccharide (SSPS)-based nanocarriers mediated by antimicrobial peptide nisin. Food Chem. 2021, 336, 127669. [Google Scholar] [CrossRef] [PubMed]
- Niaz, T.; Imran, M. Diffusion kinetics of nisin from composite coatings reinforced with nano-rhamnosomes. J. Food Eng. 2021, 288, 110143. [Google Scholar] [CrossRef]
- Pinilla, C.M.B.; Stincone, P.; Brandelli, A. Proteomic analysis reveals differential responses of Listeria monocytogenes to free and nanoencapsulated nisin. Int. J. Food Microbiol. 2021, 346, 109170. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ma, D.; Ji, T.; Sameen, D.E.; Ahmed, S.; Li, S. Long-Term Antibacterial E ff ect of Electrospun Polyvinyl Alcohol / Polyacrylate Sodium Nanofiber Containing Nisin-Loaded Nanoparticles. Nanomaterials 2020, 10, 1803. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.A.; Abushady, H.M.; Refay, R.M.; Mailam, M.A. Enhancement of the antibacterial potential of plantaricin by incorporation into silver nanoparticles. J. Genet. Eng. Biotechnol. 2021, 19, 13. [Google Scholar] [CrossRef]
- Çetinkaya, T.; Ceylan, Z.; Meral, R.; Kılıçer, A.; Altay, F. A novel strategy for Au in food science: Nanoformulation in dielectric, sensory properties, and microbiological quality of fish meat. Food Biosci. 2021, 41, 101024. [Google Scholar] [CrossRef]
- Oner, B.; Meral, R.; Ceylan, Z. Determination of some quality indices of rainbow trout fillets treated with nisin-loaded polyvinylalcohol-based nanofiber and packed with polyethylene package. LWT 2021, 149, 111854. [Google Scholar] [CrossRef]
- Akbar, A.; Sadiq, M.B.; Ali, I.; Anwar, M.; Muhammad, J.; Shafee, M.; Ullah, S.; Qasim, S.; Ahmad, S.; Anal, A.K.; et al. Lactococcus lactis subsp. lactis isolated from fermented milk products and its antimicrobial potential. CyTA-J. Food 2019, 17, 214–220. [Google Scholar] [CrossRef] [Green Version]
- José, M.; Burgos, G.; Pulido, R.P.; López, C. The Cyclic Antibacterial Peptide Enterocin AS-48: Isolation, Mode of Action, and Possible Food Applications. Int. J. Mol. Sci. 2014, 15, 22706–22727. [Google Scholar] [CrossRef] [Green Version]
- Aktypis, A.; Kalantzopoulos, G. Purification and characterization of thermophilin ST-1, a novel bacteriocin produced by Streptococcus thermophilus ACA-DC 0001. Le Lait 2003, 83, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Kotenkova, E.; Ushakova, N.; Fedosov, E. The effect of lactating and some gastrointenstinal symbiotic bacteria on the growth of young rabbits in prepubertal period. Muz. Olten. Craiova Oltenia. Stud. Şi Comunicãri. Ştiinţele Nat. 2010, 26, 1091–1096. [Google Scholar]
- Andryukov, B.; Mikhailov, V.; Besednova, N. The Biotechnological Potential of Secondary Metabolites from Marine Bacteria. J. Mar. Sci. Eng. 2019, 7, 176. [Google Scholar] [CrossRef]
- Balciunas, E.M.; Martinez, F.A.C.; Todorov, S.D.; de Melo Franco, B.D.G.; Converti, A.; de Souza Oliveira, R.P. Novel biotechnological applications of bacteriocins: A review. Food Control 2013, 32, 134–142. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafique, B.; Ranjha, M.M.A.N.; Murtaza, M.A.; Walayat, N.; Nawaz, A.; Khalid, W.; Mahmood, S.; Nadeem, M.; Manzoor, M.F.; Ameer, K.; et al. Recent Trends and Applications of Nanoencapsulated Bacteriocins against Microbes in Food Quality and Safety. Microorganisms 2023, 11, 85. https://doi.org/10.3390/microorganisms11010085
Shafique B, Ranjha MMAN, Murtaza MA, Walayat N, Nawaz A, Khalid W, Mahmood S, Nadeem M, Manzoor MF, Ameer K, et al. Recent Trends and Applications of Nanoencapsulated Bacteriocins against Microbes in Food Quality and Safety. Microorganisms. 2023; 11(1):85. https://doi.org/10.3390/microorganisms11010085
Chicago/Turabian StyleShafique, Bakhtawar, Muhammad Modassar Ali Nawaz Ranjha, Mian Anjum Murtaza, Noman Walayat, Asad Nawaz, Waseem Khalid, Shahid Mahmood, Muhammad Nadeem, Muhammad Faisal Manzoor, Kashif Ameer, and et al. 2023. "Recent Trends and Applications of Nanoencapsulated Bacteriocins against Microbes in Food Quality and Safety" Microorganisms 11, no. 1: 85. https://doi.org/10.3390/microorganisms11010085