
microRNAs: Critical Players during Helminth Infections
 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. MiRNAs as Possible Diagnosis, Prognosis, Prevention, and Treatment Tools
3. Helminths and Neglected Tropical Diseases
4. Modulation of Host Immunity by Helminth Parasites
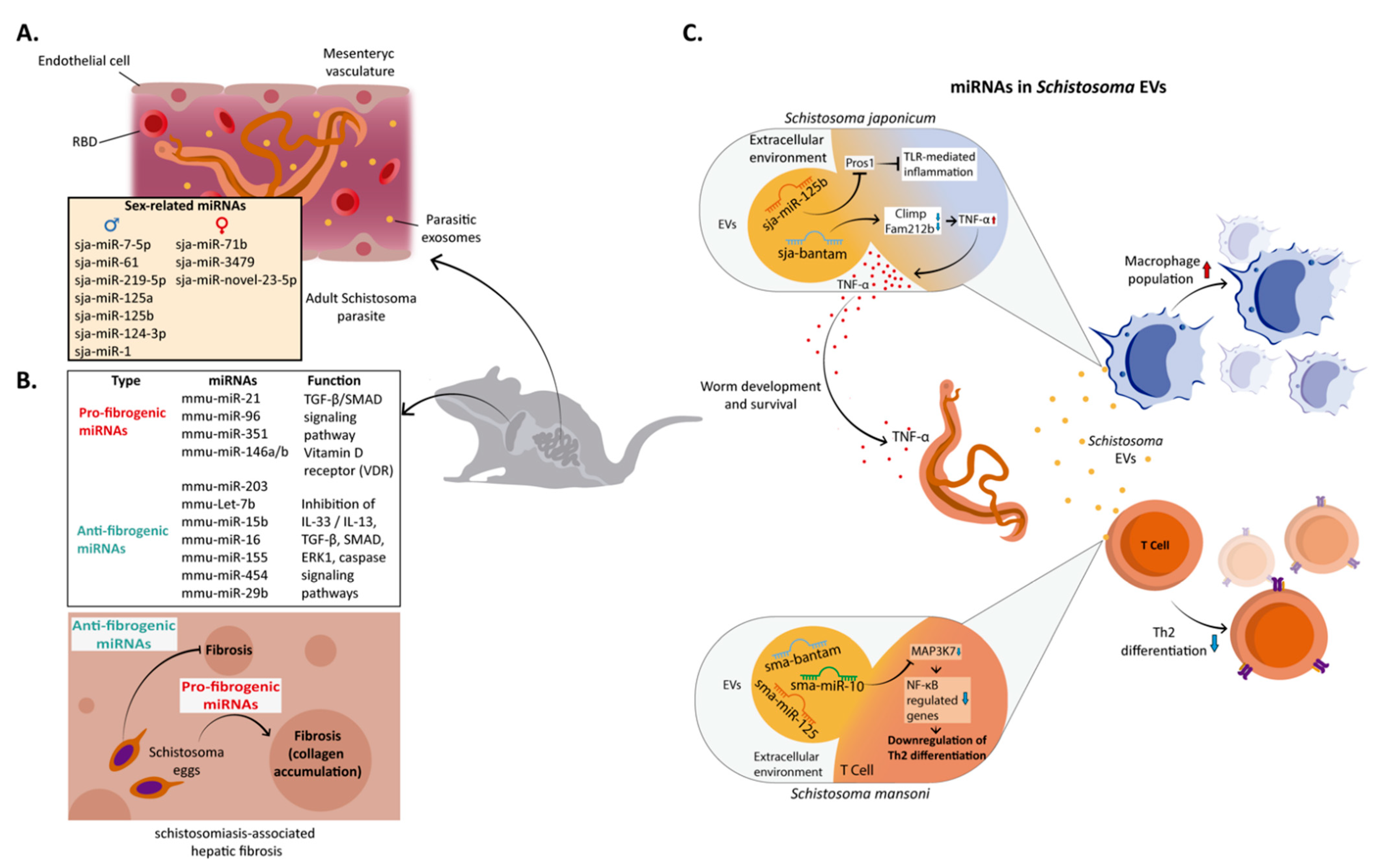
5. miRNA in Schistosoma–Host Interaction
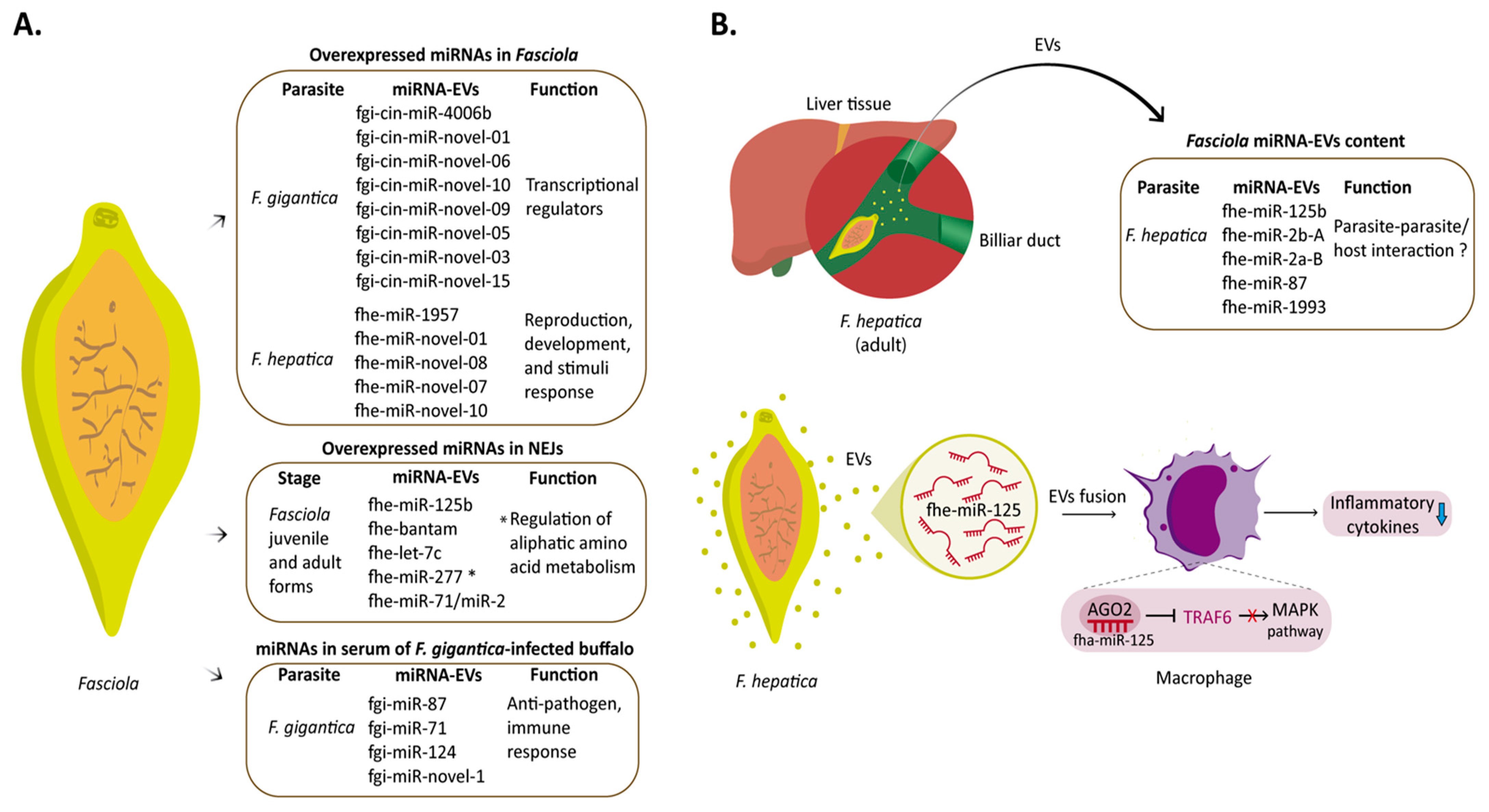
6. miRNA in Fasciola–Host Interaction
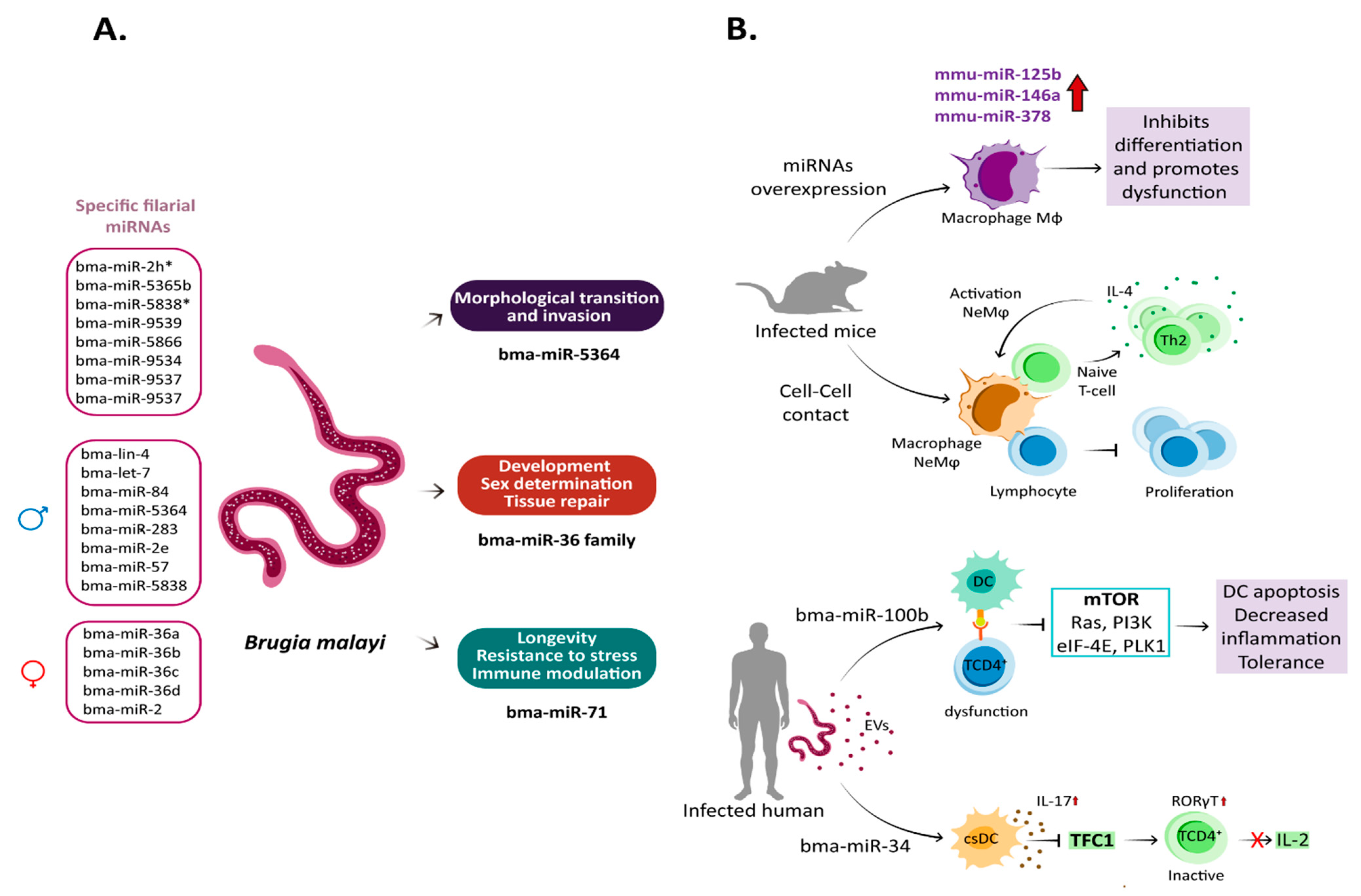
7. miRNA in Brugia malayi–Host Interaction
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carlson, C.J.; Dallas, T.A.; Alexander, L.W.; Phelan, A.L.; Phillips, A.J. What Would It Take to Describe the Global Diversity of Parasites? Proc. R. Soc. B Biol. Sci. 2020, 287, 20201841. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda, M.S.; Kinsella, J.M. Helminth Collection and Identification from Wildlife. J. Vis. Exp. 2013, 82, e51000. [Google Scholar] [CrossRef] [Green Version]
- Hotez, P.J.; Brindley, P.J.; Bethony, J.M.; King, C.H.; Pearce, E.J.; Jacobson, J. Helminth Infections: The Great Neglected Tropical Diseases. J. Clin. Investig. 2008, 118, 1311–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal-Anzardo, M.; Yagui Moscoso, M.; Beltrán Fabian, M.; Vidal-Anzardo, M.; Yagui Moscoso, M.; Beltrán Fabian, M. Parasitosis Intestinal: Helmintos. Prevalencia y Análisis de La Tendencia de Los Años 2010 a 2017 En El Perú. An. De La Fac. De Med. 2020, 81, 26–32. [Google Scholar] [CrossRef]
- Donohue, R.E.; Cross, Z.K.; Michael, E. The Extent, Nature, and Pathogenic Consequences of Helminth Polyparasitism in Humans: A Meta-Analysis. PLoS Negl. Trop. Dis. 2019, 13, e0007455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brindley, P.J.; Loukas, A. Helminth Infection–Induced Malignancy. PLoS Pathog. 2017, 13, e1006393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benesh, D.P.; Parker, G.; Chubb, J.C. Life-Cycle Complexity in Helminths: What Are the Benefits? Evolution 2021, 75, 1936–1952. [Google Scholar] [CrossRef]
- Drurey, C.; Maizels, R.M. Helminth Extracellular Vesicles: Interactions with the Host Immune System. Mol. Immunol. 2021, 137, 124–133. [Google Scholar] [CrossRef]
- McSorley, H.J.; Maizels, R.M. Helminth Infections and Host Immune Regulation. Clin. Microbiol. Rev. 2012, 25, 585–608. [Google Scholar] [CrossRef] [Green Version]
- Gazzinelli-Guimaraes, P.H.; Nutman, T.B. Helminth Parasites and Immune Regulation. F1000Research 2018, 7, 1685. [Google Scholar] [CrossRef]
- Mu, Y.; McManus, D.P.; Gordon, C.A.; Cai, P. Parasitic Helminth-Derived microRNAs and Extracellular Vesicle Cargos as Biomarkers for Helminthic Infections. Front. Cell. Infect. Microbiol. 2021, 11, 708952. [Google Scholar] [CrossRef] [PubMed]
- Sotillo, J.; Robinson, M.W.; Kimber, M.J.; Cucher, M.; Ancarola, M.E.; Nejsum, P.; Marcilla, A.; Eichenberger, R.M.; Tritten, L. The Protein and MicroRNA Cargo of Extracellular Vesicles from Parasitic Helminths–Current Status and Research Priorities. Int. J. Parasitol. 2020, 50, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; He, B.; Weiberg, A.; Buck, A.H.; Jin, H. Small RNAs and Extracellular Vesicles: New Mechanisms of Cross-Species Communication and Innovative Tools for Disease Control. PLoS Pathog. 2019, 15, e1008090. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas-Pirela, M.; Andrade-Alviárez, D.; Medina, L.; Castillo, C.; Liempi, A.; Guerrero-Muñoz, J.; Ortega, Y.; Maya, J.D.; Rojas, V.; Quiñones, W.; et al. microRNAs: Master Regulators in Host–Parasitic Protist Interactions. Open Biol. 2022, 12, 210395. [Google Scholar] [CrossRef]
- Paul, S.; Ruiz-Manriquez, L.M.; Serrano-Cano, F.I.; Estrada-Meza, C.; Solorio-Diaz, K.A.; Srivastava, A. Human microRNAs in Host-Parasite Interaction: A Review. 3 Biotech 2020, 10, 510. [Google Scholar] [CrossRef]
- Alizadeh, Z.; Mahami-Oskouei, M.; Spotin, A.; Ahmadpour, E.; Cai, P.; Sandoghchian Shotorbani, S.; Pashazadeh, F.; Ansari, F.; Mohammadi, H. microRNAs in Helminth Parasites: A Systematic Review. Curr. Mol. Med. 2022, 22, 779–808. [Google Scholar] [CrossRef]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-Cell Communication between Malaria-Infected Red Blood Cells via Exosome-like Vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Zhou, J.; Wang, H. Host microRNAs and Exosomes That Modulate Influenza Virus Infection. Virus Res. 2020, 279, 197885. [Google Scholar] [CrossRef] [PubMed]
- Mar-Aguilar, F.; Trevino, V.; Salinas-Hernández, J.E.; Taméz-Guerrero, M.M.; Barrón-González, M.P.; Morales-Rubio, E.; Treviño-Neávez, J.; Verduzco-Martínez, J.A.; Morales-Vallarta, M.R.; Reséndez-Pérez, D. Identification and Characterization of MicroRNAS from Entamoeba Histolytica HM1-IMSS. PLoS ONE 2013, 8, e68202. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.-J.; Ai, L.; Fu, J.-H.; Nisbet, A.J.; Liu, Q.-Y.; Chen, M.-X.; Zhou, D.-H.; Zhu, X.-Q. Comparative Characterization of microRNAs from the Liver Flukes Fasciola Gigantica and F. Hepatica. PLoS ONE 2012, 7, e53387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano-Román, R.; Siles-Lucas, M. microRNAs in Parasitic Diseases: Potential for Diagnosis and Targeting. Mol. Biochem. Parasitol. 2012, 186, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.; Wassmer, S.C. Harnessing the Potential of MiRNAs in Malaria Diagnostic and Prevention. Front. Cell. Infect. Microbiol. 2021, 11, 1196. [Google Scholar] [CrossRef] [PubMed]
- Sikka, R.; Bharti, P.K.; Gupta, H. microRNAs: An Opportunity to Overcome Significant Challenges in Malaria Detection and Control. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100115. [Google Scholar] [CrossRef]
- Ballinas-Verdugo, M.A.; Jiménez-Ortega, R.F.; Martínez-Martínez, E.; Rivas, N.; Contreras-López, E.A.; Carbó, R.; Sánchez, F.; Bojalil, R.; Márquez-Velasco, R.; Sánchez-Muñoz, F.; et al. Circulating MiR-146a as a Possible Candidate Biomarker in the Indeterminate Phase of Chagas Disease. Biol. Res. 2021, 54, 1–16. [Google Scholar] [CrossRef]
- Gómez-Ochoa, S.A.; Bautista-Niño, P.K.; Rojas, L.Z.; Hunziker, L.; Muka, T.; Echeverría, L.E. Circulating microRNAs and Myocardial Involvement Severity in Chronic Chagas Cardiomyopathy. Front. Cell. Infect. Microbiol. 2022, 12, 1145. [Google Scholar] [CrossRef]
- Saiyed, A.N.; Vasavada, A.R.; Johar, S.R.K. Recent Trends in MiRNA Therapeutics and the Application of Plant MiRNA for Prevention and Treatment of Human Diseases. Future J. Pharm. Sci. 2022, 8, 24. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.R.; Dias, F.; Teixeira, A.L.; Sousa, H.; Oliveira, J.; Medeiros, R. microRNAs as Potential Tools for Predicting Cancer Patients’ Susceptibility to SARS-CoV-2 Infection and Vaccination Response. Cells 2022, 11, 2279. [Google Scholar] [CrossRef]
- Ji, B.-Y.; You, Z.-H.; Cheng, L.; Zhou, J.-R.; Alghazzawi, D.; Li, L.-P. Predicting MiRNA-Disease Association from Heterogeneous Information Network with GraRep Embedding Model. Sci. Rep. 2020, 10, 6658. [Google Scholar] [CrossRef] [Green Version]
- Obad, S.; dos Santos, C.O.; Petri, A.; Heidenblad, M.; Broom, O.; Ruse, C.; Fu, C.; Lindow, M.; Stenvang, J.; Straarup, E.M.; et al. Silencing of MicroRNA Families by Seed-Targeting Tiny LNAs. Nat. Genet. 2011, 43, 371–378. [Google Scholar] [CrossRef]
- Sadovsky, Y.; Mouillet, J.-F.; Ouyang, Y.; Bayer, A.; Coyne, C.B. The Function of TrophomiRs and Other microRNAs in the Human Placenta. Cold Spring Harb. Perspect Med. 2015, 5, a023036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA Expression Profiles Classify Human Cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Anthiya, S.; Griveau, A.; Loussouarn, C.; Baril, P.; Garnett, M.; Issartel, J.-P.; Garcion, E. MicroRNA-Based Drugs for Brain Tumors. Trends Cancer 2018, 4, 222–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Ree, M.H.; van der Meer, A.J.; van Nuenen, A.C.; de Bruijne, J.; Ottosen, S.; Janssen, H.L.; Kootstra, N.A.; Reesink, H.W. Miravirsen Dosing in Chronic Hepatitis C Patients Results in Decreased MicroRNA-122 Levels without Affecting Other microRNAs in Plasma. Aliment Pharm. Ther. 2016, 43, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Medina, L.; Castillo, C.; Liempi, A.; Guerrero-Muñoz, J.; Rojas-Pirela, M.; Maya, J.D.; Prieto, H.; Kemmerling, U. Trypanosoma Cruzi and Toxoplasma Gondii Induce a Differential MicroRNA Profile in Human Placental Explants. Front. Immunol. 2020, 11, 595250. [Google Scholar] [CrossRef]
- Medina, L.; Guerrero-Muñoz, J.A.; Liempi, A.I.; Castillo, C.; Ortega, Y.; Sepúlveda, A.; Salomó, F.; Maya, J.D.; Kemmerling, U. Ex Vivo Infection of Human Placental Explants by Trypanosoma Cruzi Reveals a MicroRNA Profile Similar to That Seen in Trophoblast Differentiation. Pathogens 2022, 11, 361. [Google Scholar] [CrossRef]
- Li, H.; Ouyang, Y.; Sadovsky, E.; Parks, W.T.; Chu, T.; Sadovsky, Y. Unique MicroRNA Signals in Plasma Exosomes from Pregnancies Complicated by Preeclampsia. Hypertension 2020, 75, 762–771. [Google Scholar] [CrossRef]
- Motran, C.C.; Silvane, L.; Chiapello, L.S.; Theumer, M.G.; Ambrosio, L.F.; Volpini, X.; Celias, D.P.; Cervi, L. Helminth Infections: Recognition and Modulation of the Immune Response by Innate Immune Cells. Front. Immunol. 2018, 9, 664. [Google Scholar] [CrossRef]
- Ezenwa, V.O. Host Social Behavior and Parasitic Infection: A Multifactorial Approach. Behav. Ecol. 2004, 15, 446–454. [Google Scholar] [CrossRef]
- Charlier, J.; van der Voort, M.; Kenyon, F.; Skuce, P.; Vercruysse, J. Chasing Helminths and Their Economic Impact on Farmed Ruminants. Trends Parasitol. 2014, 30, 361–367. [Google Scholar] [CrossRef]
- DiNardo, A. Helminth Infections and Their Impact on Global Public Health. Clin. Infect. Dis. 2015, 60, 675. [Google Scholar] [CrossRef] [Green Version]
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.-N. Schistosomiasis. Nat. Rev. Dis. Prim. 2018, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Richter, J. The Impact of Chemotherapy on Morbidity Due to Schistosomiasis. Acta Trop. 2003, 86, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, D.; Perez-Saez, J.; Vounatsou, P.; Utzinger, J.; Arcand, J.-L. The Economic Impact of Schistosomiasis. Infect. Dis. Poverty 2021, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Schistosomiasis. Available online: https://www.who.int/news-room/fact-sheets/detail/schistosomiasis (accessed on 13 September 2022).
- Verjee, M.A. Schistosomiasis: Still a Cause of Significant Morbidity and Mortality. Res. Rep. Trop. Med. 2019, 10, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danso-Appiah, T. Schistosomiasis. In Neglected Tropical Diseases-Sub-Saharan Africa; Neglected Tropical Diseases; Gyapong, J., Boatin, B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 251–288. ISBN 978-3-319-25469-2. [Google Scholar]
- Saba, R.; Korkmaz, M.; Inan, D.; Mamikoğlu, L.; Turhan, Ö.; Günseren, F.; Çevikol, C.; Kabaalioğlu, A. Human Fascioliasis. Clin. Microbiol. Infect. 2004, 10, 385–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Sánchez, R.; Domenech Cañete, I.; Millán Marcelo, J.C.; Pino Santos, A. Fascioliasis, Revisión Clínico-Epidemiológica y Diagnóstico. Rev. Cuba. De Hig. Y Epidemiol. 2012, 50, 88–96. [Google Scholar]
- Nyindo, M.; Lukambagire, A.-H. Fascioliasis: An Ongoing Zoonotic Trematode Infection. Biomed. Res. Int. 2015, 2015, 786195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caravedo, M.A.; Cabada, M.M. Human Fascioliasis: Current Epidemiological Status and Strategies for Diagnosis, Treatment, and Control. Res. Rep. Trop. Med. 2020, 11, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Alba, A.; Vazquez, A.A.; Hurtrez-Boussès, S. Towards the Comprehension of Fasciolosis (Re-)Emergence: An Integrative Overview. Parasitology 2021, 148, 385–407. [Google Scholar] [CrossRef] [PubMed]
- Charlier, J.; Rinaldi, L.; Musella, V.; Ploeger, H.W.; Chartier, C.; Vineer, H.R.; Hinney, B.; von Samson-Himmelstjerna, G.; Băcescu, B.; Mickiewicz, M.; et al. Initial Assessment of the Economic Burden of Major Parasitic Helminth Infections to the Ruminant Livestock Industry in Europe. Prev. Vet. Med. 2020, 182, 105103. [Google Scholar] [CrossRef] [PubMed]
- Rebollo, M.P.; Bockarie, M.J. Can Lymphatic Filariasis Be Eliminated by 2020? Trends Parasitol. 2017, 33, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Melrose, W.; Warner, J.; Buttner, P.; Ward, L. Lymphatic Filariasis: A Method to Identify Subclinical Lower Limb Change in PNG Adolescents. PLoS Negl. Trop. Dis. 2011, 5, e1242. [Google Scholar] [CrossRef] [PubMed]
- Lourens, G.B.; Ferrell, D.K. Lymphatic Filariasis. Nurs. Clin. N. Am. 2019, 54, 181–192. [Google Scholar] [CrossRef]
- Medeiros, Z.M.; Vieira, A.V.B.; Xavier, A.T.; Bezerra, G.S.N.; Lopes, M.D.F.C.; Bonfim, C.V.; Aguiar-Santos, A.M. Lymphatic Filariasis: A Systematic Review on Morbidity and Its Repercussions in Countries in the Americas. Int. J. Environ. Res. Public Health 2021, 19, 316. [Google Scholar] [CrossRef]
- Dietrich, C.F.; Chaubal, N.; Hoerauf, A.; Kling, K.; Piontek, M.S.; Steffgen, L.; Mand, S.; Dong, Y. Review of Dancing Parasites in Lymphatic Filariasis. Ultrasound Int. Open 2019, 5, E65–E74. [Google Scholar] [CrossRef] [Green Version]
- Bizhani, N.; Hashemi Hafshejani, S.; Mohammadi, N.; Rezaei, M.; Rokni, M.B. Lymphatic Filariasis in Asia: A Systematic Review and Meta-Analysis. Parasitol. Res. 2021, 120, 411–422. [Google Scholar] [CrossRef]
- Mathew, C.G.; Bettis, A.A.; Chu, B.K.; English, M.; Ottesen, E.A.; Bradley, M.H.; Turner, H.C. The Health and Economic Burdens of Lymphatic Filariasis Prior to Mass Drug Administration Programs. Clin. Infect Dis. 2020, 70, 2561–2567. [Google Scholar] [CrossRef]
- Maizels, R.M.; McSorley, H.J. Regulation of the Host Immune System by Helminth Parasites. J. Allergy Clin. Immunol. 2016, 138, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Maizels, R.M.; Smits, H.H.; McSorley, H.J. Modulation of Host Immunity by Helminths: The Expanding Repertoire of Parasite Effector Molecules. Immunity 2018, 49, 801–818. [Google Scholar] [CrossRef] [Green Version]
- Regev-Rudzki, N.; Michaeli, S.; Torrecilhas, A.C. Editorial: Extracellular Vesicles in Infectious Diseases. Front. Cell. Infect. Microbiol. 2021, 11, 430. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, C.M.; Trelis, M.; Bernal, D.; Marcilla, A. Overview of the Interaction of Helminth Extracellular Vesicles with the Host and Their Potential Functions and Biological Applications. Mol. Immunol. 2021, 134, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gong, W.; Cao, S.; Yin, J.; Zhang, J.; Cao, J.; Shen, Y. Comprehensive Analysis of Non-Coding RNA Profiles of Exosome-Like Vesicles From the Protoscoleces and Hydatid Cyst Fluid of Echinococcus Granulosus. Front. Cell. Infect. Microbiol. 2020, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hao, P.; Chen, H.; Hu, W.; Yan, Q.; Liu, F.; Han, Z.-G. Genome-Wide Identification of Schistosoma Japonicum microRNAs Using a Deep-Sequencing Approach. PLoS ONE 2009, 4, e8206. [Google Scholar] [CrossRef]
- Huang, A.; Yang, Y.; Chen, S.; Xia, F.; Sun, D.; Fang, D.; Xiong, S.; Jin, L.; Zhang, J. MiR-34a Promotes DCs Development and Inhibits Their Function on T Cell Activation by Targeting WNT1. Oncotarget 2017, 8, 17191–17201. [Google Scholar] [CrossRef] [Green Version]
- Pérez, M.G.; Spiliotis, M.; Rego, N.; Macchiaroli, N.; Kamenetzky, L.; Holroyd, N.; Cucher, M.A.; Brehm, K.; Rosenzvit, M.C. Deciphering the Role of MiR-71 in Echinococcus Multilocularis Early Development in Vitro. PLoS Negl. Trop. Dis. 2019, 13, e0007932. [Google Scholar] [CrossRef] [Green Version]
- Ricafrente, A.; Nguyen, H.; Tran, N.; Donnelly, S. An Evaluation of the Fasciola Hepatica MiRnome Predicts a Targeted Regulation of Mammalian Innate Immune Responses. Front. Immunol. 2021, 11, 608686. [Google Scholar] [CrossRef]
- Ricafrente, A.; Cwiklinski, K.; Nguyen, H.; Dalton, J.P.; Tran, N.; Donnelly, S. Stage-Specific MiRNAs Regulate Gene Expression Associated with Growth, Development and Parasite-Host Interaction during the Intra-Mammalian Migration of the Zoonotic Helminth Parasite Fasciola Hepatica. BMC Genom. 2022, 23, 419. [Google Scholar] [CrossRef]
- Ancarola, M.E.; Lichtenstein, G.; Herbig, J.; Holroyd, N.; Mariconti, M.; Brunetti, E.; Berriman, M.; Albrecht, K.; Marcilla, A.; Rosenzvit, M.C.; et al. Extracellular Non-Coding RNA Signatures of the Metacestode Stage of Echinococcus Multilocularis. PLoS Negl. Trop. Dis. 2020, 14, e0008890. [Google Scholar] [CrossRef]
- Simões, M.C.; Lee, J.; Djikeng, A.; Cerqueira, G.C.; Zerlotini, A.; da Silva-Pereira, R.A.; Dalby, A.R.; LoVerde, P.; El-Sayed, N.M.; Oliveira, G. Identification of Schistosoma Mansoni microRNAs. BMC Genom. 2011, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Piao, X.; Liu, S.; Hou, N.; Wang, H.; Chen, Q. MicroRNA-Gene Expression Network in Murine Liver during Schistosoma Japonicum Infection. PLoS ONE 2013, 8, e67037. [Google Scholar] [CrossRef] [PubMed]
- Marco, A.; Kozomara, A.; Hui, J.H.L.; Emery, A.M.; Rollinson, D.; Griffiths-Jones, S.; Ronshaugen, M. Sex-Biased Expression of microRNAs in Schistosoma Mansoni. PLoS Negl. Trop. Dis. 2013, 7, e2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Xue, X.; Sun, J.; Luo, R.; Xu, X.; Jiang, Y.; Zhang, Q.; Pan, W. An “In-Depth” Description of the Small Non-Coding RNA Population of Schistosoma Japonicum Schistosomulum. PLoS Negl. Trop. Dis. 2010, 4, e596. [Google Scholar] [CrossRef] [PubMed]
- de Souza Gomes, M.; Muniyappa, M.K.; Carvalho, S.G.; Guerra-Sá, R.; Spillane, C. Genome-Wide Identification of Novel microRNAs and Their Target Genes in the Human Parasite Schistosoma Mansoni. Genomics 2011, 98, 96–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, P.; Hou, N.; Piao, X.; Liu, S.; Liu, H.; Yang, F.; Wang, J.; Jin, Q.; Wang, H.; Chen, Q. Profiles of Small Non-Coding RNAs in Schistosoma Japonicum during Development. PLoS Negl. Trop. Dis. 2011, 5, e1256. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Liu, S.; Piao, X.; Hou, N.; Gobert, G.N.; McManus, D.P.; Chen, Q. Comprehensive Transcriptome Analysis of Sex-Biased Expressed Genes Reveals Discrete Biological and Physiological Features of Male and Female Schistosoma Japonicum. PLoS Negl. Trop. Dis. 2016, 10, e0004684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, P.; Gobert, G.N.; McManus, D.P. microRNAs in Parasitic Helminthiases: Current Status and Future Perspectives. Trends Parasitol. 2016, 32, 71–86. [Google Scholar] [CrossRef]
- Xue, X.; Sun, J.; Zhang, Q.; Wang, Z.; Huang, Y.; Pan, W. Identification and Characterization of Novel microRNAs from Schistosoma Japonicum. PLoS ONE 2008, 3, e4034. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Piao, X.; Hao, L.; Liu, S.; Hou, N.; Wang, H.; Chen, Q. A Deep Analysis of the Small Non-Coding RNA Population in Schistosoma Japonicum Eggs. PLoS ONE 2013, 8, e64003. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhang, J.; Zheng, T.; Chen, H.; Nie, H.; Zheng, B.; Gong, Q. The Role of microRNAs in the Pathogenesis, Grading and Treatment of Hepatic Fibrosis in Schistosomiasis. Parasites Vectors 2019, 12, 611. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-J.; Fang, Z.-M.; Liu, W.-Q. NLRP3 Inflammasome Activation from Kupffer Cells Is Involved in Liver Fibrosis of Schistosoma Japonicum-Infected Mice via NF-ΚB. Parasites Vectors 2019, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Sun, Y.; Lei, N.; Fan, X.; Zhang, C.; Wang, Y.; Zheng, K.; Zhang, D.; Pan, W. MicroRNA-351 Promotes Schistosomiasis-Induced Hepatic Fibrosis by Targeting the Vitamin D Receptor. Proc. Natl. Acad. Sci. USA 2018, 115, 180–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Zhang, D.; Xie, J.; Su, Q.; He, X.; Bai, R.; Gao, G.; Pan, W. MicroRNA-96 Promotes Schistosomiasis Hepatic Fibrosis in Mice by Suppressing Smad7. Mol. Ther. Methods Clin. Dev. 2018, 11, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.-C. MiRNA-Mediated Macrophage Polarization and Its Potential Role in the Regulation of Inflammatory Response. Shock 2016, 46, 122–131. [Google Scholar] [CrossRef]
- Chen, X.; Tang, J.; Shuai, W.; Meng, J.; Feng, J.; Han, Z. Macrophage Polarization and Its Role in the Pathogenesis of Acute Lung Injury/Acute Respiratory Distress Syndrome. Inflamm. Res. 2020, 69, 883–895. [Google Scholar] [CrossRef]
- Kim, K.K.; Sheppard, D.; Chapman, H.A. TGF-Β1 Signaling and Tissue Fibrosis. Cold Spring Harb. Perspect Biol. 2018, 10, a022293. [Google Scholar] [CrossRef] [Green Version]
- Webster, H.C.; Gamino, V.; Andrusaite, A.T.; Ridgewell, O.J.; McCowan, J.; Shergold, A.L.; Heieis, G.A.; Milling, S.W.F.; Maizels, R.M.; Perona-Wright, G. Tissue-Based IL-10 Signalling in Helminth Infection Limits IFNγ Expression and Promotes the Intestinal Th2 Response. Mucosal. Immunol. 2022, 15, 1257–1269. [Google Scholar] [CrossRef]
- Dai, W.; Zhao, J.; Tang, N.; Zeng, X.; Wu, K.; Ye, C.; Shi, J.; Lu, C.; Ning, B.; Zhang, J.; et al. MicroRNA-155 Attenuates Activation of Hepatic Stellate Cell by Simultaneously Preventing EMT Process and ERK1 Signalling Pathway. Liver Int. 2015, 35, 1234–1243. [Google Scholar] [CrossRef]
- Wang, J.; Chu, E.S.H.; Chen, H.-Y.; Man, K.; Go, M.Y.Y.; Huang, X.R.; Lan, H.Y.; Sung, J.J.Y.; Yu, J. MicroRNA-29b Prevents Liver Fibrosis by Attenuating Hepatic Stellate Cell Activation and Inducing Apoptosis through Targeting PI3K/AKT Pathway. Oncotarget 2015, 6, 7325–7338. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.-P.; Ai, W.-B.; Wan, L.-Y.; Zhang, Y.-Q.; Wu, J.-F. The Roles of MicroRNA Families in Hepatic Fibrosis. Cell Biosci. 2017, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Tang, N.; Wu, Y.; Cao, W.; Liang, Y.; Gao, Y.; Hu, L.; Yang, Q.; Zhou, Y.; Tang, F.; Xiao, J. Lentivirus-Mediated over-Expression of Let-7b MicroRNA Suppresses Hepatic Fibrosis in the Mouse Infected with Schistosoma Japonicum. Exp. Parasitol. 2017, 182, 45–53. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xie, J.; Wang, Y.; Fan, X.; Su, Q.; Sun, Y.; Lei, N.; Zhang, D.; Gao, G.; Pan, W. Down-Regulation of MicroRNA-203-3p Initiates Type 2 Pathology during Schistosome Infection via Elevation of Interleukin-33. PLoS Pathog. 2018, 14, e1006957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Dang, Z.; Chong, S. Mmu-MiR-92a-2-5p Targets TLR2 to Relieve Schistosoma Japonicum-Induced Liver Fibrosis. Int. Immunopharmacol. 2019, 69, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Loverde, P.T.; Osman, A.; Hinck, A. Schistosoma Mansoni: TGF-Beta Signaling Pathways. Exp. Parasitol. 2007, 117, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.; Niles, E.G.; Verjovski-Almeida, S.; LoVerde, P.T. Schistosoma Mansoni TGF-Beta Receptor II: Role in Host Ligand-Induced Regulation of a Schistosome Target Gene. PLoS Pathog. 2006, 2, e54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Peng, J.; Hong, Y.; Zhang, M.; Han, Y.; Liu, D.; Fu, Z.; Shi, Y.; Xu, J.; Tao, J.; et al. MicroRNA Expression Profile in Different Tissues of BALB/c Mice in the Early Phase of Schistosoma Japonicum Infection. Mol. Biochem. Parasitol. 2013, 188, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kifle, D.W.; Chaiyadet, S.; Waardenberg, A.J.; Wise, I.; Cooper, M.; Becker, L.; Doolan, D.L.; Laha, T.; Sotillo, J.; Pearson, M.S.; et al. Uptake of Schistosoma Mansoni Extracellular Vesicles by Human Endothelial and Monocytic Cell Lines and Impact on Vascular Endothelial Cell Gene Expression. Int. J. Parasitol. 2020, 50, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Meningher, T.; Barsheshet, Y.; Ofir-Birin, Y.; Gold, D.; Brant, B.; Dekel, E.; Sidi, Y.; Schwartz, E.; Regev-Rudzki, N.; Avni, O.; et al. Schistosomal Extracellular Vesicle-Enclosed MiRNAs Modulate Host T Helper Cell Differentiation. EMBO Rep. 2020, 21, e47882. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, L.; Wang, J.; Qiu, L.; Chen, Y.; Davis, R.E.; Cheng, G. Schistosoma Japonicum Extracellular Vesicle MiRNA Cargo Regulates Host Macrophage Functions Facilitating Parasitism. PLoS Pathog. 2019, 15, e1007817. [Google Scholar] [CrossRef] [Green Version]
- Amiri, P.; Locksley, R.M.; Parslow, T.G.; Sadickt, M.; Rector, E.; Ritter, D.; McKerrow, J.H. Tumour Necrosis Factor α Restores Granulomas and Induces Parasite Egg-Laying in Schistosome-Infected SCID Mice. Nature 1992, 356, 604–607. [Google Scholar] [CrossRef]
- Witsell, A.L.; Schook, L.B. Tumor Necrosis Factor Alpha Is an Autocrine Growth Regulator during Macrophage Differentiation. Proc. Natl. Acad. Sci. USA 1992, 89, 4754–4758. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Junior, E.H.; Bertevello, C.R.; de Oliveira Silveira, G.; Guedes, C.B.; Rodrigues, G.D.; Ribeiro, V.S.; Amaral, M.S.; Kanamura, C.T.; Pinto, P.L.S.; Krüger, R.F.; et al. Human Tumor Necrosis Factor Alpha Affects the Egg-Laying Dynamics and Glucose Metabolism of Schistosoma Mansoni Adult Worms in Vitro. Parasites Vectors 2022, 15, 176. [Google Scholar] [CrossRef]
- Oliveira, K.C.; Carvalho, M.L.P.; Venancio, T.M.; Miyasato, P.A.; Kawano, T.; DeMarco, R.; Verjovski-Almeida, S. Identification of the Schistosoma Mansoni TNF-Alpha Receptor Gene and the Effect of Human TNF-Alpha on the Parasite Gene Expression Profile. PLoS Negl. Trop. Dis. 2009, 3, e556. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, K.C.; Carvalho, M.L.P.; Bonatto, J.M.C.; Schechtman, D.; Verjovski-Almeida, S. Human TNF-α Induces Differential Protein Phosphorylation in Schistosoma Mansoni Adult Male Worms. Parasitol. Res. 2016, 115, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; Hewitson, J.P.; Smith, K.A. Susceptibility and Immunity to Helminth Parasites. Curr. Opin. Immunol. 2012, 24, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.E.; Sutherland, T.E. Host Protective Roles of Type 2 Immunity: Parasite Killing and Tissue Repair, Flip Sides of the Same Coin. Semin. Immunol. 2014, 26, 329–340. [Google Scholar] [CrossRef]
- Hoy, A.M.; Lundie, R.J.; Ivens, A.; Quintana, J.F.; Nausch, N.; Forster, T.; Jones, F.; Kabatereine, N.B.; Dunne, D.W.; Mutapi, F.; et al. Parasite-Derived microRNAs in Host Serum As Novel Biomarkers of Helminth Infection. PLoS Negl. Trop. Dis. 2014, 8, e2701. [Google Scholar] [CrossRef] [Green Version]
- Mu, Y.; Cai, P.; Olveda, R.M.; Ross, A.G.; Olveda, D.U.; McManus, D.P. Parasite-Derived Circulating microRNAs as Biomarkers for the Detection of Human Schistosoma Japonicum Infection. Parasitology 2020, 147, 889–896. [Google Scholar] [CrossRef]
- Zhong, H.; Jin, Y. Multifunctional Roles of microRNAs in Schistosomiasis. Front. Microbiol. 2022, 13, 925386. [Google Scholar] [CrossRef]
- Cai, P.; Gobert, G.N.; You, H.; Duke, M.; McManus, D.P. Circulating MiRNAs: Potential Novel Biomarkers for Hepatopathology Progression and Diagnosis of Schistosomiasis Japonica in Two Murine Models. PLoS Negl. Trop. Dis. 2015, 9, e0003965. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Fu, Z.; Cao, X.; Lin, J. Changes in MicroRNA Expression in Response to Schistosoma Japonicum Infection. Parasite Immunol. 2017, 39, e12416. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Tripathi, S.; Singh, A.K.; Mondal, P.; Mishra, A.; Prasad, A. Micromanagement of Immune System: Role of MiRNAs in Helminthic Infections. Front. Microbiol. 2017, 8, 586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromm, B.; Ovchinnikov, V.; Høye, E.; Bernal, D.; Hackenberg, M.; Marcilla, A. On the Presence and Immunoregulatory Functions of Extracellular microRNAs in the Trematode Fasciola Hepatica. Parasite Immunol. 2017, 39, e12399. [Google Scholar] [CrossRef] [PubMed]
- Malatji, M.P.; Pfukenyi, D.M.; Mukaratirwa, S. Fasciola Species and Their Vertebrate and Snail Intermediate Hosts in East and Southern Africa: A Review. J. Helminthol. 2019, 94, e63. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Cwiklinski, K.; Hu, R.-S.; Zheng, W.-B.; Sheng, Z.-A.; Zhang, F.-K.; Elsheikha, H.M.; Dalton, J.P.; Zhu, X.-Q. Complex and Dynamic Transcriptional Changes Allow the Helminth Fasciola Gigantica to Adjust to Its Intermediate Snail and Definitive Mammalian Hosts. BMC Genom. 2019, 20, 729. [Google Scholar] [CrossRef] [Green Version]
- Gennarino, V.A.; Sardiello, M.; Avellino, R.; Meola, N.; Maselli, V.; Anand, S.; Cutillo, L.; Ballabio, A.; Banfi, S. MicroRNA Target Prediction by Expression Analysis of Host Genes. Genome Res. 2009, 19, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Fontenla, S.; Dell’Oca, N.; Smircich, P.; Tort, J.F.; Siles-Lucas, M. The MiRnome of Fasciola Hepatica Juveniles Endorses the Existence of a Reduced Set of Highly Divergent Micro RNAs in Parasitic Flatworms. Int. J. Parasitol. 2015, 45, 901–913. [Google Scholar] [CrossRef]
- Stark, A.; Brennecke, J.; Russell, R.B.; Cohen, S.M. Identification of Drosophila MicroRNA Targets. PLoS Biol. 2003, 1, e60. [Google Scholar] [CrossRef]
- Marcilla, A.; Trelis, M.; Cortés, A.; Sotillo, J.; Cantalapiedra, F.; Minguez, M.T.; Valero, M.L.; Sánchez del Pino, M.M.; Muñoz-Antoli, C.; Toledo, R.; et al. Extracellular Vesicles from Parasitic Helminths Contain Specific Excretory/Secretory Proteins and Are Internalized in Intestinal Host Cells. PLoS ONE 2012, 7, e45974. [Google Scholar] [CrossRef] [PubMed]
- Fromm, B.; Trelis, M.; Hackenberg, M.; Cantalapiedra, F.; Bernal, D.; Marcilla, A. The Revised MicroRNA Complement of Fasciola Hepatica Reveals a Plethora of Overlooked microRNAs and Evidence for Enrichment of Immuno-Regulatory microRNAs in Extracellular Vesicles. Int. J. Parasitol. 2015, 45, 697–702. [Google Scholar] [CrossRef]
- Tran, N.; Ricafrente, A.; To, J.; Lund, M.; Marques, T.M.; Gama-Carvalho, M.; Cwiklinski, K.; Dalton, J.P.; Donnelly, S. Fasciola Hepatica Hijacks Host Macrophage MiRNA Machinery to Modulate Early Innate Immune Responses. Sci. Rep. 2021, 11, 6712. [Google Scholar] [CrossRef] [PubMed]
- Montaner, S.; Galiano, A.; Trelis, M.; Martin-Jaular, L.; del Portillo, H.A.; Bernal, D.; Marcilla, A. The Role of Extracellular Vesicles in Modulating the Host Immune Response during Parasitic Infections. Front. Immunol. 2014, 5, 433. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Guo, A. Profiling Circulating microRNAs in Serum of Fasciola Gigantica-Infected Buffalo. Mol. Biochem. Parasitol. 2019, 232, 111201. [Google Scholar] [CrossRef] [PubMed]
- Morishita, A.; Oura, K.; Tadokoro, T.; Fujita, K.; Tani, J.; Masaki, T. MicroRNA Interference in Hepatic Host-Pathogen Interactions. Int. J. Mol. Sci. 2021, 22, 3554. [Google Scholar] [CrossRef] [PubMed]
- Bhoj, P.; Togre, N.; Khatri, V.; Goswami, K. Harnessing Immune Evasion Strategy of Lymphatic Filariae: A Therapeutic Approach against Inflammatory and Infective Pathology. Vaccines 2022, 10, 1235. [Google Scholar] [CrossRef]
- Poole, C.B.; Gu, W.; Kumar, S.; Jin, J.; Davis, P.J.; Bauche, D.; McReynolds, L.A. Diversity and Expression of microRNAs in the Filarial Parasite, Brugia Malayi. PLoS ONE 2014, 9, e96498. [Google Scholar] [CrossRef] [Green Version]
- Ricciardi, A.; Bennuru, S.; Tariq, S.; Kaur, S.; Wu, W.; Elkahloun, A.G.; Arakelyan, A.; Shaik, J.; Dorward, D.W.; Nutman, T.B.; et al. Extracellular Vesicles Released from the Filarial Parasite Brugia Malayi Downregulate the Host MTOR Pathway. PLoS Negl. Trop. Dis. 2021, 15, e0008884. [Google Scholar] [CrossRef]
- Abbott, A.L. Uncovering New Functions for microRNAs in Caenorhabditis Elegans. Curr. Biol. 2011, 21, R668–R671. [Google Scholar] [CrossRef] [Green Version]
- Favre, R.; Cermola, M.; Nunes, C.P.; Hermann, R.; Müller, M.; Bazzicalupo, P. Immuno-Cross-Reactivity of CUT-1 and Cuticlin Epitopes between Ascaris Lumbricoides, Caenorhabditis Elegans, and Heterorhabditis. J. Struct Biol. 1998, 123, 1–7. [Google Scholar] [CrossRef]
- Sapio, M.R.; Hilliard, M.A.; Cermola, M.; Favre, R.; Bazzicalupo, P. The Zona Pellucida Domain Containing Proteins, CUT-1, CUT-3 and CUT-5, Play Essential Roles in the Development of the Larval Alae in Caenorhabditis Elegans. Dev. Biol. 2005, 282, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Winter, A.D.; Gillan, V.; Maitland, K.; Emes, R.D.; Roberts, B.; McCormack, G.; Weir, W.; Protasio, A.V.; Holroyd, N.; Berriman, M.; et al. A Novel Member of the Let-7 MicroRNA Family Is Associated with Developmental Transitions in Filarial Nematode Parasites. BMC Genom. 2015, 16, 331. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Ghedin, E.; Berriman, M.; McQuillan, J.; Holroyd, N.; Mayhew, G.F.; Christensen, B.M.; Michalski, M.L. A Deep Sequencing Approach to Comparatively Analyze the Transcriptome of Lifecycle Stages of the Filarial Worm, Brugia Malayi. PLoS Negl. Trop. Dis. 2011, 5, e1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, N.C.; Lim, L.P.; Weinstein, E.G.; Bartel, D.P. An Abundant Class of Tiny RNAs with Probable Regulatory Roles in Caenorhabditis Elegans. Science 2001, 294, 858–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, M.; Artiles, K.; Pak, J.; Fire, A. Contributions of MRNA Abundance, Ribosome Loading, and Post- or Peri-Translational Effects to Temporal Repression of C. Elegans Heterochronic MiRNA Targets. Genome Res. 2012, 22, 2418–2426. [Google Scholar] [CrossRef] [Green Version]
- Lawson, H.; Vuong, E.; Miller, R.M.; Kiontke, K.; Fitch, D.H.; Portman, D.S. The Makorin Lep-2 and the LncRNA Lep-5 Regulate Lin-28 to Schedule Sexual Maturation of the C. Elegans Nervous System. Elife 2019, 8, e43660. [Google Scholar] [CrossRef]
- Zarkower, D.; Hodgkin, J. Zinc Fingers in Sex Determination: Only One of the Two C. Elegans Tra-1 Proteins Binds DNA in Vitro. Nucleic Acids Res. 1993, 21, 3691–3698. [Google Scholar] [CrossRef] [Green Version]
- Hart, A.H.; Reventar, R.; Bernstein, A. Genetic Analysis of ETS Genes in C. Elegans. Oncogene 2000, 19, 6400–6408. [Google Scholar] [CrossRef] [Green Version]
- Reeves, R. Nuclear Functions of the HMG Proteins. Biochim. Biophys. Acta 2010, 1799, 3–14. [Google Scholar] [CrossRef]
- Ngwa, C.J.; Farrukh, A.; Pradel, G. Zinc Finger Proteins of Plasmodium Falciparum. Cell. Microbiol. 2021, 23, e13387. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Czech, B.; Crunk, A.; Wallace, A.; Mitreva, M.; Hannon, G.J.; Davis, R.E. Deep Small RNA Sequencing from the Nematode Ascaris Reveals Conservation, Functional Diversification, and Novel Developmental Profiles. Genome Res. 2011, 21, 1462–1477. [Google Scholar] [CrossRef] [Green Version]
- Winter, A.D.; Weir, W.; Hunt, M.; Berriman, M.; Gilleard, J.S.; Devaney, E.; Britton, C. Diversity in Parasitic Nematode Genomes: The microRNAs of Brugia Pahangi and Haemonchus Contortus Are Largely Novel. BMC Genom. 2012, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Saavedra, E.; Horvitz, H.R. Many Families of C. Elegans microRNAs Are Not Essential for Development or Viability. Curr. Biol. 2010, 20, 367–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, C.; Winter, A.D.; Gillan, V.; Devaney, E. microRNAs of Parasitic Helminths–Identification, Characterization and Potential as Drug Targets. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palakodeti, D.; Smielewska, M.; Graveley, B.R. microRNAs from the Planarian Schmidtea Mediterranea: A Model System for Stem Cell Biology. RNA 2006, 12, 1640–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wit, E.; Linsen, S.E.V.; Cuppen, E.; Berezikov, E. Repertoire and Evolution of MiRNA Genes in Four Divergent Nematode Species. Genome Res. 2009, 19, 2064–2074. [Google Scholar] [CrossRef] [Green Version]
- Friedländer, M.R.; Adamidi, C.; Han, T.; Lebedeva, S.; Isenbarger, T.A.; Hirst, M.; Marra, M.; Nusbaum, C.; Lee, W.L.; Jenkin, J.C.; et al. High-Resolution Profiling and Discovery of Planarian Small RNAs. Proc. Natl. Acad. Sci. USA 2009, 106, 11546–11551. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-S.; Zhou, H.-C.; Wei, C.-Y.; Wang, Z.-H.; Hao, X.; Zhang, L.-H.; Li, J.-Z.; Wang, Z.-L.; Wang, H. Global Survey of MiRNAs and TRNA-Derived Small RNAs from the Human Parasitic Protist Trichomonas Vaginalis. Parasit. Vectors 2021, 14, 87. [Google Scholar] [CrossRef]
- González-Estévez, C.; Arseni, V.; Thambyrajah, R.S.; Felix, D.A.; Aboobaker, A.A. Diverse MiRNA Spatial Expression Patterns Suggest Important Roles in Homeostasis and Regeneration in Planarians. Int. J. Dev. Biol. 2009, 53, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Sasidharan, V.; Lu, Y.-C.; Bansal, D.; Dasari, P.; Poduval, D.; Seshasayee, A.; Resch, A.M.; Graveley, B.R.; Palakodeti, D. Identification of Neoblast- and Regeneration-Specific MiRNAs in the Planarian Schmidtea Mediterranea. RNA 2013, 19, 1394–1404. [Google Scholar] [CrossRef] [Green Version]
- de Lencastre, A.; Pincus, Z.; Zhou, K.; Kato, M.; Lee, S.S.; Slack, F.J. microRNAs Both Promote and Antagonize Longevity in C. Elegans. Curr. Biol. 2010, 20, 2159–2168. [Google Scholar] [CrossRef] [Green Version]
- Boulias, K.; Horvitz, H.R. The C. Elegans MicroRNA Mir-71 Acts in Neurons to Promote Germline-Mediated Longevity through Regulation of DAF-16/FOXO. Cell Metab. 2012, 15, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Kennedy, S.; Tolonen, A.C.; Ruvkun, G. DAF-16 Target Genes That Control C. Elegans Life-Span and Metabolism. Science 2003, 300, 644–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Voronin, D.; Poole, C.B.; Bachu, S.; Rogers, M.B.; Jin, J.; Ghedin, E.; Lustigman, S.; McReynolds, L.A.; Unnasch, T.R. Functional Analysis of MicroRNA Activity in Brugia Malayi. Int. J. Parasitol. 2015, 45, 579–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes Secreted by Nematode Parasites Transfer Small RNAs to Mammalian Cells and Modulate Innate Immunity. Nat. Commun. 2014, 5, 5488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Guo, X.; He, W.; Shao, Z.; Zhang, X.; Yang, J.; Shen, Y.; Luo, X.; Cao, J. Effects of Echinococcus Multilocularis MiR-71 Mimics on Murine Macrophage RAW264.7 Cells. Int. Immunopharmacol. 2016, 34, 259–262. [Google Scholar] [CrossRef]
- Anthony, R.M.; Urban, J.F.; Alem, F.; Hamed, H.A.; Rozo, C.T.; Boucher, J.-L.; Van Rooijen, N.; Gause, W.C. Memory TH2 Cells Induce Alternatively Activated Macrophages to Mediate Protection against Nematode Parasites. Nat. Med. 2006, 12, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Towatari, M.; May, G.E.; Marais, R.; Perkins, G.R.; Marshall, C.J.; Cowley, S.; Enver, T. Regulation of GATA-2 Phosphorylation by Mitogen-Activated Protein Kinase and Interleukin-3. J. Biol. Chem. 1995, 270, 4101–4107. [Google Scholar] [CrossRef] [Green Version]
- Rückerl, D.; Jenkins, S.J.; Laqtom, N.N.; Gallagher, I.J.; Sutherland, T.E.; Duncan, S.; Buck, A.H.; Allen, J.E. Induction of IL-4Rα-Dependent microRNAs Identifies PI3K/Akt Signaling as Essential for IL-4-Driven Murine Macrophage Proliferation in Vivo. Blood 2012, 120, 2307–2316. [Google Scholar] [CrossRef]
- Chaudhuri, A.A.; So, A.Y.-L.; Sinha, N.; Gibson, W.S.J.; Taganov, K.D.; O’Connell, R.M.; Baltimore, D. MicroRNA-125b Potentiates Macrophage Activation. J. Immunol. 2011, 187, 5062–5068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Huang, G.; Xu, Q.; Zhao, G.; Jiang, J.; Li, Y.; Guo, Z. MiR-146a-5p Attenuates Allergic Airway Inflammation by Inhibiting the NLRP3 Inflammasome Activation in Macrophages. IAA 2022, 183, 919–930. [Google Scholar] [CrossRef]
- Nam, S.; Kang, K.; Cha, J.S.; Kim, J.W.; Lee, H.G.; Kim, Y.; Yang, Y.; Lee, M.-S.; Lim, J.-S. Interferon Regulatory Factor 4 (IRF4) Controls Myeloid-Derived Suppressor Cell (MDSC) Differentiation and Function. J. Leukoc. Biol. 2016, 100, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.E.; Lawrence, R.A.; Maizels, R.M. APC from Mice Harbouring the Filarial Nematode, Brugia Malayi, Prevent Cellular Proliferation but Not Cytokine Production. Int. Immunol. 1996, 8, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loke, P.; MacDonald, A.S.; Robb, A.; Maizels, R.M.; Allen, J.E. Alternatively Activated Macrophages Induced by Nematode Infection Inhibit Proliferation via Cell-to-Cell Contact. Eur. J. Immunol. 2000, 30, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Loke, P.; MacDonald, A.S.; Allen, J.E. Antigen-Presenting Cells Recruited by Brugia Malayi Induce Th2 Differentiation of Naïve CD4(+) T Cells. Eur. J. Immunol. 2000, 30, 1127–1135. [Google Scholar] [CrossRef]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Babu, S.; Nutman, T.B. Immunopathogenesis of Lymphatic Filarial Disease. Semin. Immunopathol. 2012, 34, 847–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, M.; Fraser, L.M.; Agbedanu, P.N.; Harischandra, H.; Moorhead, A.R.; Day, T.A.; Bartholomay, L.C.; Kimber, M.J. Release of Small RNA-Containing Exosome-like Vesicles from the Human Filarial Parasite Brugia Malayi. PLoS Negl. Trop. Dis. 2015, 9, e0004069. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.D.; Pollizzi, K.N.; Heikamp, E.B.; Horton, M.R. Regulation of Immune Responses by MTOR. Annu. Rev. Immunol. 2012, 30, 39–68. [Google Scholar] [CrossRef] [Green Version]
- Deleyto-Seldas, N.; Efeyan, A. The MTOR–Autophagy Axis and the Control of Metabolism. Front. Cell Dev. Biol. 2021, 9, 1519. [Google Scholar] [CrossRef]
- Chen, P.; Zhao, X.; Ma, L. Downregulation of MicroRNA-100 Correlates with Tumor Progression and Poor Prognosis in Hepatocellular Carcinoma. Mol. Cell Biochem. 2013, 383, 49–58. [Google Scholar] [CrossRef]
- Negi, V.; Paul, D.; Das, S.; Bajpai, P.; Singh, S.; Mukhopadhyay, A.; Agrawal, A.; Ghosh, B. Altered Expression and Editing of MiRNA-100 Regulates ITreg Differentiation. Nucleic Acids Res. 2015, 43, 8057–8065. [Google Scholar] [CrossRef]
- Nihal, M.; Stutz, N.; Schmit, T.; Ahmad, N.; Wood, G.S. Polo-like Kinase 1 (Plk1) Is Expressed by Cutaneous T-Cell Lymphomas (CTCLs), and Its Downregulation Promotes Cell Cycle Arrest and Apoptosis. Cell Cycle 2011, 10, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, G.; Liu, X.; Zhou, L.; Jiang, M.; Yang, L. Polo-Like Kinase 1 (PLK1) Is Involved in Toll-like Receptor (TLR)-Mediated TNF-α Production in Monocytic THP-1 Cells. PLoS ONE 2013, 8, e78832. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.; Nickl, L.; Walch-Rueckheim, B.; Krammes, L.; Rheinheimer, S.; Diener, C.; Taenzer, T.; Kehl, T.; Sester, M.; Lenhof, H.-P.; et al. Wrinkle in the Plan: MiR-34a-5p Impacts Chemokine Signaling by Modulating CXCL10/CXCL11/CXCR3-Axis in CD4+, CD8+ T Cells, and M1 Macrophages. J. Immunother. Cancer 2020, 8, e001617. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-W.; Deftos, M.L.; Ojala, E.W.; Bevan, M.J. RORγt, a Novel Isoform of an Orphan Receptor, Negatively Regulates Fas Ligand Expression and IL-2 Production in T Cells. Immunity 1998, 9, 797–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar, G.; Mangani, D.; Anderson, A.C. T Cell Factor 1 (Tcf1): A Master Regulator of the T Cell Response in Disease. Sci. Immunol. 2020, 5, eabb9726. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas-Pirela, M.; Andrade-Alviárez, D.; Quiñones, W.; Rojas, M.V.; Castillo, C.; Liempi, A.; Medina, L.; Guerrero-Muñoz, J.; Fernández-Moya, A.; Ortega, Y.A.; et al. microRNAs: Critical Players during Helminth Infections. Microorganisms 2023, 11, 61. https://doi.org/10.3390/microorganisms11010061
Rojas-Pirela M, Andrade-Alviárez D, Quiñones W, Rojas MV, Castillo C, Liempi A, Medina L, Guerrero-Muñoz J, Fernández-Moya A, Ortega YA, et al. microRNAs: Critical Players during Helminth Infections. Microorganisms. 2023; 11(1):61. https://doi.org/10.3390/microorganisms11010061
Chicago/Turabian StyleRojas-Pirela, Maura, Diego Andrade-Alviárez, Wilfredo Quiñones, Maria Verónica Rojas, Christian Castillo, Ana Liempi, Lisvaneth Medina, Jesus Guerrero-Muñoz, Alejandro Fernández-Moya, Yessica Andreina Ortega, and et al. 2023. "microRNAs: Critical Players during Helminth Infections" Microorganisms 11, no. 1: 61. https://doi.org/10.3390/microorganisms11010061