
Ciliate Morpho-Taxonomy and Practical Considerations before Deploying Metabarcoding to Ciliate Community Diversity Surveys in Urban Receiving Waters

{kind=link}
Abstract
:1. Introduction
2. Ciliates Involved in Self-Purification in Treatment Plants and Urban Waterbodies
3. Traditional Ciliate Taxonomy and Its Investigation in Freshwater Systems
4. Phylogenetic Taxonomy and DNA Barcoding of Ciliates
5. High Throughput Sequencing and Metabarcoding
6. Factors That Might Sistort Ciliate Biodiversity Assessments Based on Metabarcoding
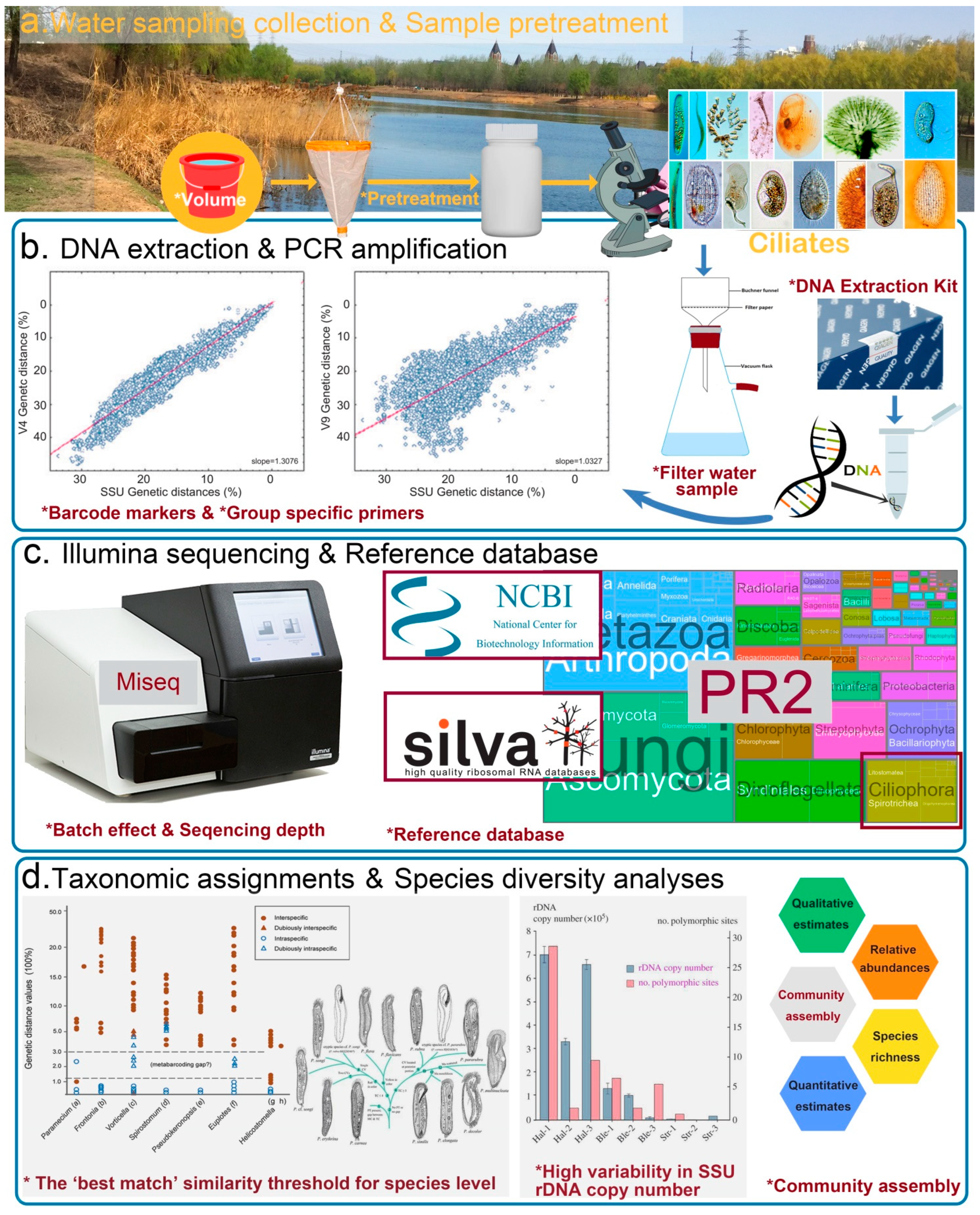
6.1. Field Sampling and Pretreatment
6.2. DNA Extraction
6.3. PCR Amplification and the Choice of Barcode
6.4. Amplicons Sequencing on NGS Platform
7. Bioinformatics
7.1. Data Filtering
7.2. Sequence Classification
8. Conclusion: Developing Universal and Standardized DNA-Based Methods for Ciliate Studies in Urban Waterbodies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sabater, S.; Guasch, H.; Romaní, A.; Muñoz, I. The effect of biological factors on the efficiency of river biofilms in improving water quality. Hydrobiologia 2002, 469, 149–156. [Google Scholar] [CrossRef]
- Postel, S.; Richter, B. Rivers for Life: Managing Water for People and Nature; Island Press: Washington, DC, USA, 2012; pp. 1–40. [Google Scholar]
- Pernet-Coudrier, B.; Qi, W.; Liu, H.; Müller, B.; Berg, M. Sources and pathways of nutrients in the Semi-Arid Region of Beijing–Tianjin, China. Environ. Sci. Technol. 2012, 46, 5294–5301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curds, C.; Cockburn, A. Protozoa in biological sewage-treatment processes—I. A survey of the protozoan fauna of British percolating filters and activated-sludge plants. Water Res. 1970, 4, 225–236. [Google Scholar] [CrossRef]
- Curds, C.; Cockburn, A. Protozoa in biological sewage-treatment processes—II. Protozoa as indicators in the activated-sludge process. Water Res. 1970, 4, 237–249. [Google Scholar] [CrossRef]
- Bick, H. Ciliated protozoa: An Illustrated Guide to the Species Used as Biological Indicators in Freshwater Biology; World Health Organization: Geneva, Switzerland, 1972; 198p. [Google Scholar]
- Foissner, W.; Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshw. Biol. 1996, 35, 375–482. [Google Scholar] [CrossRef]
- Song, W.; Warren, A.; Hu, X. Free-Living Ciliates in the Bohai and Yellow Seas; Science Press: Beijing, China, 2009. [Google Scholar]
- Chi, Y.; Chen, X.; Li, Y.; Wang, C.; Zhang, T.; Ayoub, A.; Warren, A.; Song, W.; Wang, Y. New contributions to the phylogeny of the ciliate class Heterotrichea (Protista, Ciliophora): Analyses at family-genus level and new evolutionary hypotheses. Sci. China Life Sci. 2021, 64, 606–620. [Google Scholar] [CrossRef]
- Moon-van der Staay, S.Y.; De Wachter, R.; Vaulot, D. Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 2001, 409, 607–610. [Google Scholar] [CrossRef]
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef] [Green Version]
- Massana, R.; Gobet, A.; Audic, S.; Bass, D.; Bittner, L.; Boutte, C.; Chambouvet, A.; Christen, R.; Claverie, J.-M.; Decelle, J.; et al. Marine protist diversity in European coastal waters and sediments as revealed by high-throughput sequencing. Environ. Microbiol. 2015, 17, 4035–4049. [Google Scholar] [CrossRef]
- Pawlowski, J.; Lejzerowicz, F.; Apotheloz-Perret-Gentil, L.; Visco, J.; Esling, P. Protist metabarcoding and environmental biomonitoring: Time for change. Eur. J. Protistol. 2016, 55, 12–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagnetti, R.; Miana, P.; Fabris, M.; Pavoni, B. Self-purification ability of a resurgence stream. Chemosphere 2003, 52, 1781–1795. [Google Scholar] [CrossRef] [PubMed]
- Foissner, W. Progress in taxonomy of planktonic freshwater ciliates. Mar. Microb. Food Webs 1994, 8, 9–35. [Google Scholar]
- Foissner, W. Protists as bioindicators in activated sludge: Identification, ecology and future needs. Eur. J. Protistol. 2016, 55, 75–94. [Google Scholar] [CrossRef]
- Shen, Y.F.; Zhang, Z.; Gong, X.J.; Gu, M.R.; Shi, X.Z.; Wei, Y.X. Modern Biomonitoring Techniques Using Freshwater Microbiota; China Building Industry Press: Beijing, China, 1900. [Google Scholar]
- Otterholt, E.; Charnock, C. Identification and phylogeny of the small eukaryote population of raw and drinking waters. Water Res. 2011, 45, 2527–2538. [Google Scholar] [CrossRef]
- Lin, W.; Yu, Z.; Zhang, H.; Thompson, I.P. Diversity and dynamics of microbial communities at each step of treatment plant for potable water generation. Water Res. 2014, 52, 218–230. [Google Scholar] [CrossRef]
- Stoeck, T.; Pan, H.; Dully, V.; Forster, D.; Jung, T. Towards an eDNA metabarcode-based performance indicator for full-scale municipal wastewater treatment plants. Water Res. 2018, 144, 322–331. [Google Scholar] [CrossRef]
- Sanford, R.A.; Lloyd, K.G.; Konstantinidis, K.T.; Löffler, F.E. Microbial taxonomy run amok. Trends Microbiol. 2021, 29, 394–404. [Google Scholar] [CrossRef]
- Warren, A.; Patterson, D.J.; Dunthorn, M.; Clamp, J.C.; Achilles-Day, U.E.M.; Aescht, E.; Al-Farraj, S.A.; Al-Quraishy, S.; Al-Rasheid, K.; Carr, M.; et al. Beyond the “Code”: A guide to the description and documentation of biodiversity in ciliated protists (Alveolata, Ciliophora). J. Eukaryot. Microbiol. 2017, 64, 539–554. [Google Scholar] [CrossRef] [Green Version]
- Dobell, C. Antony Van Leeuwenhoek and His Little Animals; Harcourt, Brace & Company: New York, NY, USA, 1932. [Google Scholar]
- Lane, N. The unseen world: Reflections on Leeuwenhoek (1677) ‘Concerning little animals’. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140344. [Google Scholar] [CrossRef]
- Fokin, S.I. A brief history of ciliate studies (late XVII the first third of the XX century). Protistology 2004, 3, 283–296. [Google Scholar]
- Foissner, W.; Chao, A.; Katz, L.A. Diversity and geographic distribution of ciliates (Protista: Ciliophora). Biodivers. Conserv. 2007, 17, 345–363. [Google Scholar] [CrossRef]
- Hu, X.; Lin, X.; Song, W. Ciliate Atlas: Species Found in the South China Sea; Science Press: Beijing, China, 2019. [Google Scholar]
- Sonneborn, T.M. Recent advances in the genetics of Paramecium and Euplotes. Adv. Genet. 1947, 1, 263–358. [Google Scholar] [PubMed]
- Blackburn, E.H.; Gall, J.G. A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J. Mol. Evol. 1978, 120, 33–53. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Godfray, H.C.J. Challenges for taxonomy. Nature 2002, 417, 17–19. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, C.; Lynch, M.; Gao, S. The compact macronuclear genome of the ciliate Halteria grandinella: A transcriptome-like genome with 23,000 nanochromosomes. mBio 2021, 12, e01964-20. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.G.; Pace, M.L.; Battey, J.F. Ciliate protozoans as links in freshwater planktonic food chains. Nature 1979, 277, 563–565. [Google Scholar] [CrossRef]
- Beaver, J.R.; Crisman, T.L. The trophic response of ciliated protozoans in freshwater lakes. Limnol. Oceanogr. 1982, 27, 246–253. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.; Meyer-Reil, L.; Thingstad, F. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Lima-Mendez, G.; Faust, K.; Henry, N.; Decelle, J.; Colin, S.; Carcillo, F.; Chaffron, S.; Ignacio-Espinosa, J.C.; Roux, S.; Vincent, F. Determinants of community structure in the global plankton interactome. Science 2015, 348, 1262073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Yi, Z.; Gentekaki, E.; Zhan, A.; Al-Farraj, S.A.; Song, W. Utility of combining morphological characters, nuclear and mitochondrial genes: An attempt to resolve the conflicts of species identification for ciliated protists. Mol. Phylogenet. Evol. 2016, 94, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yi, Z.; Warren, A.; Song, W.B. Species delimitation for the molecular taxonomy and ecology of the widely distributed microbial eukaryote genus Euplotes (Alveolata, Ciliophora). Proc. R. Soc. B 2018, 285, 20172159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobzhansky, T. Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Hennig, W. Phylogenetic systematics. University of Illinois Press Hu SK, Liu Z, Lie AAY, Countway PD, Kim DY, Jones AC, Gast RJ, Cary SC, Sherr EB, Sherr BF, Caron DA (2015)Estimating protistan diversity using high-throughput sequencing. J. Eukaryot. Microbiol. 1999, 62, 688–693. [Google Scholar]
- De Queiroz, K.; Gauthier, J. Phylogenetic taxonomy. Annu. Rev. Ecol. Syst. 1992, 23, 449–480. [Google Scholar] [CrossRef]
- Yang, Z.; Rannala, B. Molecular phylogenetics: Principles and practice. Nat. Rev. Genet. 2012, 13, 303–314. [Google Scholar] [CrossRef]
- Lee, M.S.; Palci, A. Morphological phylogenetics in the genomic age. Curr. Biol. 2015, 25, R922–R929. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.; Ratnasingham, S.; deWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B 2003, 270, S596–S599. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Lynn, D.H. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature; Springer: New York, NY, USA, 2008. [Google Scholar]
- Pawlowski, J.; Audic, S.; Adl, S.; Bass, D.; Belbahri, L.; Berney, C.; Bowser, S.S.; Cepicka, I.; Decelle, J.; Dunthorn, M. CBOL Protist Working Group: Barcoding Eukaryotic Richness beyond the Animal, Plant, and Fungal Kingdoms. PLoS Biol. 2012, 10, e1001419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chantangsi, C.; Lynn, D.H.; Brandl, M.T.; Cole, J.C.; Hetrick, N.; Ikonomi, P. Barcoding ciliates: A comprehensive study of 75 isolates of the genus Tetrahymena. Int. J. Syst. Evol. Microbiol. 2007, 57, 2412–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, D.; Tischer, K.; Berger, H.; Schlegel, M.; Berendonk, T.U. High mitochondrial haplotype diversity of Coleps sp. (Ciliophora: Prostomatida). Environ. Microbiol. 2008, 10, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Catania, F.; Wurmser, F.; Potekhin, A.A.; Przybos, E.; Lynch, M. Genetic diversity in the Paramecium aurelia species complex. Mol. Biol. Evol 2009, 26, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Strüder-Kypke, M.C.; Lynn, D.H. Comparative analysis of the mitochondrial cytochrome c oxidase subunit I (COI) gene in ciliates (Alveolata, Ciliophora) and evaluation of its suitability as a biodiversity marker. Syst. Biodivers. 2010, 8, 131–148. [Google Scholar] [CrossRef]
- Zhao, Y.; Gentekaki, E.; Yi, Z.; Lin, X. Genetic differentiation of the mitochondrial cytochrome oxidase c subunit I gene in genus Paramecium (Protista, Ciliophora). PLoS ONE 2013, 8, e77044. [Google Scholar] [CrossRef] [Green Version]
- Stoeck, T.; Breiner, H.W.; Filker, S.; Ostermaier, V.; Kammerlander, B.; Sonntag, B. A morphogenetic survey on ciliate plankton from a mountain lake pinpoints the necessity of lineage-specific barcode markers in microbial ecology. Environ. Microbiol. 2014, 16, 430–444. [Google Scholar] [CrossRef] [Green Version]
- Dunthorn, M.; Klier, J.; Bunge, J.; Stoeck, T. Comparing the hyper-variable V4 and V9 regions of the small subunit rDNA for assessment of ciliate environmental diversity. J. Eukaryot. Microbiol. 2012, 59, 185–187. [Google Scholar] [CrossRef]
- Choi, J.; Park, J.S. Comparative analyses of the V4 and V9 regions of 18S rDNA for the extant eukaryotic community using the Illumina platform. Sci. Rep. 2020, 10, 6519. [Google Scholar] [CrossRef] [Green Version]
- Stoeck, T.; Behnke, A.; Christen, R.; Amaral-Zettler, L.; Rodriguez-Mora, M.J.; Chistoserdov, A.; Orsi, W.; Edgcomb, V.P. Massively parallel tag sequencing reveals the complexity of anaerobic marine protistan communities. BMC Biol. 2009, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Weisse, T. Freshwater ciliates as ecophysiological model organisms-lessons from Daphnia, major achievements, and future perspectives. Arch. Hydrobiol. 2006, 167, 371–402. [Google Scholar] [CrossRef] [Green Version]
- Weisse, T.; Montagnes, D.J. Ecology of planktonic ciliates in a changing world: Concepts, methods, and challenges. J. Eukaryot. Microbiol. 2021, 69, e12879. [Google Scholar] [CrossRef] [PubMed]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; De Vargas, C.; Decelle, J. The Protist Ribosomal Reference database (PR2): A catalog of unicellular eukaryote small subunit rRNA sequences with curated taxonomy. Nucleic Acids Res. 2013, 41, D597–D604. [Google Scholar] [CrossRef] [PubMed]
- Boscaro, V.; Santoferrara, L.F.; Zhang, Q.; Gentekaki, E.; Syberg-Olsen, M.J.; del Campo, J.; Keeling, P.J. EukRef–Ciliophora: A manually curated, phylogeny-based database of small subunit rRNA gene sequences of ciliates. Environ. Microbiol. 2018, 20, 2218–2230. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.; Kolisko, M.; Boscaro, V.; Santoferrara, L.F.; Nenarokov, S.; Massana, R.; Guillou, L.; Simpson, A.; Berney, C.; de Vargas, C. EukRef: Phylogenetic curation of ribosomal RNA to enhance understanding of eukaryotic diversity and distribution. PLoS Biol. 2018, 16, e2005849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vd’ačný, P.; Foissner, W. Re-analysis of the 18S rRNA gene phylogeny of the ciliate class Colpodea. Eur. J. Protistol. 2019, 67, 89–105. [Google Scholar] [CrossRef]
- Mardis, E.R. Next-generation DNA sequencing methods. Annu. Rev. Genomics Hum. Genet. 2008, 9, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Bik, H.M.; Porazinska, D.L.; Creer, S.; Caporaso, J.G.; Knight, R.; Thomas, W.K. Sequencing our way towards understanding global eukaryotic biodiversity. Trends Ecol. Evol. 2012, 27, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- van Dijk, E.L.; Auger, H.; Jaszczyszyn, Y.; Thermes, C. Ten years of next-generation sequencing technology. Trends Genet. 2014, 30, 418–426. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [PubMed]
- Zinger, L.; Bonin, A.; Alsos, I.G.; Bálint, M.; Bik, H.; Boyer, F.; Chariton, A.A.; Creer, S.; Coissac, E.; Deagle, B.E. DNA metabarcoding—Need for robust experimental designs to draw sound ecological conclusions. Mol. Ecol. 2019, 28, 1857–1862. [Google Scholar] [PubMed] [Green Version]
- Zhan, Z.; Li, J.; Xu, K. Ciliate environmental diversity can be underestimated by the V4 region of SSU rDNA: Insights from species delimitation and multilocus phylogeny of Pseudokeronopsis (Protist, Ciliophora). Microorganisms 2019, 7, 493. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, T.; Wang, Y.; Katz, L.A.; Gao, F.; Song, W. Disentangling sources of variation in SSU rDNA sequences from single cell analyses of ciliates: Impact of copy number variation and experimental error. Proc. R. Soc. B 2017, 284, 20170425. [Google Scholar] [PubMed] [Green Version]
- Bálint, M.; Bahram, M.; Eren, A.M.; Faust, K.; Fuhrman, J.A.; Lindahl, B.; O’Hara, R.B.; Öpik, M.; Sogin, M.L.; Unterseher, M. Millions of reads, thousands of taxa: Microbial community structure and associations analyzed via marker genes. FEMS Microbiol. Rev. 2016, 40, 686–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šlapeta, J.; Moreira, D.; López-García, P. The extent of protist diversity: Insights from molecular ecology of freshwater eukaryotes. Proc. R. Soc. Lond. Ser. B Biol. 2005, 272, 2073–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.; Breiner, H.W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef]
- Campbell, R.G.; Ashjian, C.J.; Sherr, E.B.; Sherr, B.F.; Lomas, M.W.; Ross, C.; Alatalo, P.; Gelfman, C.; Van Keuren, D. Mesozooplankton grazing during spring sea-ice conditions in the eastern Bering Sea. Deep Sea Res Part II Top. Stud. Oceanogr. 2016, 134, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Gimmler, A.; Korn, R.; De Vargas, C.; Audic, S.; Stoeck, T. The Tara Oceans voyage reveals global diversity and distribution patterns of marine planktonic ciliates. Sci. Rep. 2016, 6, 33555. [Google Scholar] [CrossRef] [Green Version]
- Le Bescot, N.; Mahé, F.; Audic, S.; Dimier, C.; Garet, M.J.; Poulain, J.; Wincker, P.; de Vargas, C.; Siano, R. Global patterns of pelagic dinoflagellate diversity across protist size classes unveiled by metabarcoding. Environ. Microbiol. 2016, 18, 609–626. [Google Scholar] [CrossRef]
- Santoferrara, L.F.; Rubin, E.; Mcmanus, G.B. Global and local DNA (meta) barcoding reveal new biogeography patterns in tintinnid ciliates. J. Plankton Res. 2018, 40, 209–221. [Google Scholar] [CrossRef]
- Novotny, A.; Zamora-Terol, S.; Winder, M. DNA metabarcoding reveals trophic niche diversity of micro and mesozooplankton species. Proc. R. Soc. B 2021, 288, 20210908. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.M.; Sasso, T.; Valentini, A.; Dejean, T.; Martins, M.; Zamudio, K.R.; Haddad, C.F. eDNA metabarcoding: A promising method for anuran surveys in highly diverse tropical forests. Mol. Ecol. Resour. 2017, 17, 904–914. [Google Scholar] [CrossRef]
- Bessey, C.; Jarman, S.N.; Berry, O.; Olsen, Y.S.; Bunce, M.; Simpson, T.; Power, M.; McLaughlin, J.; Edgar, G.J.; Keesing, J. Maximizing fish detection with eDNA metabarcoding. Environ. DNA 2020, 2, 493–504. [Google Scholar] [CrossRef]
- Jeunen, G.J.; Knapp, M.; Spencer, H.G.; Taylor, H.R.; Lamare, M.D.; Stat, M.; Bunce, M.; Gemmell, N.J. Species-level biodiversity assessment using marine environmental DNA metabarcoding requires protocol optimization and standardization. Ecol. Evol. 2019, 9, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Loos, L.M.; Nijland, R. Biases in bulk: DNA metabarcoding of marine communities and the methodology involved. Mol. Ecol. 2021, 30, 3270–3288. [Google Scholar] [CrossRef] [PubMed]
- Eckert, K.A.; Kunkel, T.A. DNA polymerase fidelity and the polymerase chain reaction. Genome Res. 1991, 1, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, R.V.; Vollmers, C.; Newsom, L.A.; Wang, Y.; Heintzman, P.D.; Leighton, M.; Green, R.E.; Shapiro, B. Minimizing polymerase biases in metabarcoding. Mol. Ecol. Resour. 2018, 18, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Coissac, E.; Riaz, T.; Puillandre, N. Bioinformatic challenges for DNA metabarcoding of plants and animals. Mol. Ecol. 2012, 21, 1834–1847. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A method for studying protistan diversity using massively parallel sequencing of V9 hypervariable regions of small-subunit ribosomal RNA genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Pitsch, G.; Bruni, E.P.; Forster, D.; Qu, Z.; Sonntag, B.; Stoeck, T.; Posch, T. Seasonality of planktonic freshwater ciliates: Are analyses based on V9 regions of the 18S rRNA gene correlated with morphospecies counts? Front. Microbiol. 2019, 10, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamady, M.; Walker, J.J.; Harris, J.K.; Gold, N.J.; Knight, R. Error-correcting barcoded primers allow hundreds of samples to be pyrosequenced in multiplex. Nat. Methods 2008, 5, 235. [Google Scholar] [CrossRef]
- Leek, J.T.; Scharpf, R.B.; Bravo, H.C.; Simcha, D.; Langmead, B.; Johnson, W.E.; Geman, D.; Baggerly, K.; Irizarry, R.A. Tackling the widespread and critical impact of batch effects in high-throughput data. Nat. Rev. Genet. 2010, 11, 733–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoferrara, L.; Burki, F.; Filker, S.; Logares, R.; Dunthorn, M.; McManus, G.B. Perspectives from ten years of protist studies by high-throughput metabarcoding. J. Eukaryot. Microbiol. 2020, 67, 612–622. [Google Scholar] [CrossRef]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2, High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- Kolisko, M.; Flegontova, O.; Karnkowska, A.; Lax, G.; Maritz, J.M.; Pánek, T.; Táborský, P.; Carlton, J.M.; Čepička, I.; Horák, A.; et al. EukRef-excavates: Seven curated SSU ribosomal RNA gene databases. Database 2020, 2020, baaa080. [Google Scholar]
- Gong, J.; Dong, J.; Liu, X.; Massana, R. Extremely high copy numbers and polymorphisms of the rDNA operon estimated from single cell analysis of oligotrich and peritrich ciliates. Protist 2013, 164, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Paiva, T.d.S. Systematic redefinition of the Hypotricha (Alveolata, Ciliophora) based on combined analyses of morphological and molecular characters. Cladistics. Protist. 2020, 171, 125755. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J.; Willmott, K. Taxonomy: Renaissance or Tower of Babel? Trends Ecol. Evol. 2003, 18, 57–59. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. Lond. 2005, 85, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Pierce, R.W.; Turner, J.T. Ecology of planktonic ciliates in marine food webs. Rev. Aquatic Sci. 1992, 6, 139–181. [Google Scholar]
- Corliss, J.O. The Ciliated Protozoa: Characterization, Classification and Guide to the Literature; Pergamon Press: Oxford, UK, 1979. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Langlois, G.A. Ciliate Morpho-Taxonomy and Practical Considerations before Deploying Metabarcoding to Ciliate Community Diversity Surveys in Urban Receiving Waters. Microorganisms 2022, 10, 2512. https://doi.org/10.3390/microorganisms10122512
Zhao Y, Langlois GA. Ciliate Morpho-Taxonomy and Practical Considerations before Deploying Metabarcoding to Ciliate Community Diversity Surveys in Urban Receiving Waters. Microorganisms. 2022; 10(12):2512. https://doi.org/10.3390/microorganisms10122512
Chicago/Turabian StyleZhao, Yan, and Gaytha A. Langlois. 2022. "Ciliate Morpho-Taxonomy and Practical Considerations before Deploying Metabarcoding to Ciliate Community Diversity Surveys in Urban Receiving Waters" Microorganisms 10, no. 12: 2512. https://doi.org/10.3390/microorganisms10122512