
Influence of SARS-COV-2 Infection on Cytokine Production by Mitogen-Stimulated Peripheral Blood Mononuclear Cells and Neutrophils in COVID-19 Intensive Care Unit Patients
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Real-Time Reverse Transcriptase PCR Assays for Detection of SARS-CoV-2 RNA
2.3. Isolation of PBMCs
2.4. Isolation of Neutrophils
2.5. Mitogen-Induced Stimulation
2.6. Estimation of Cytokine Levels
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the Patients Studied
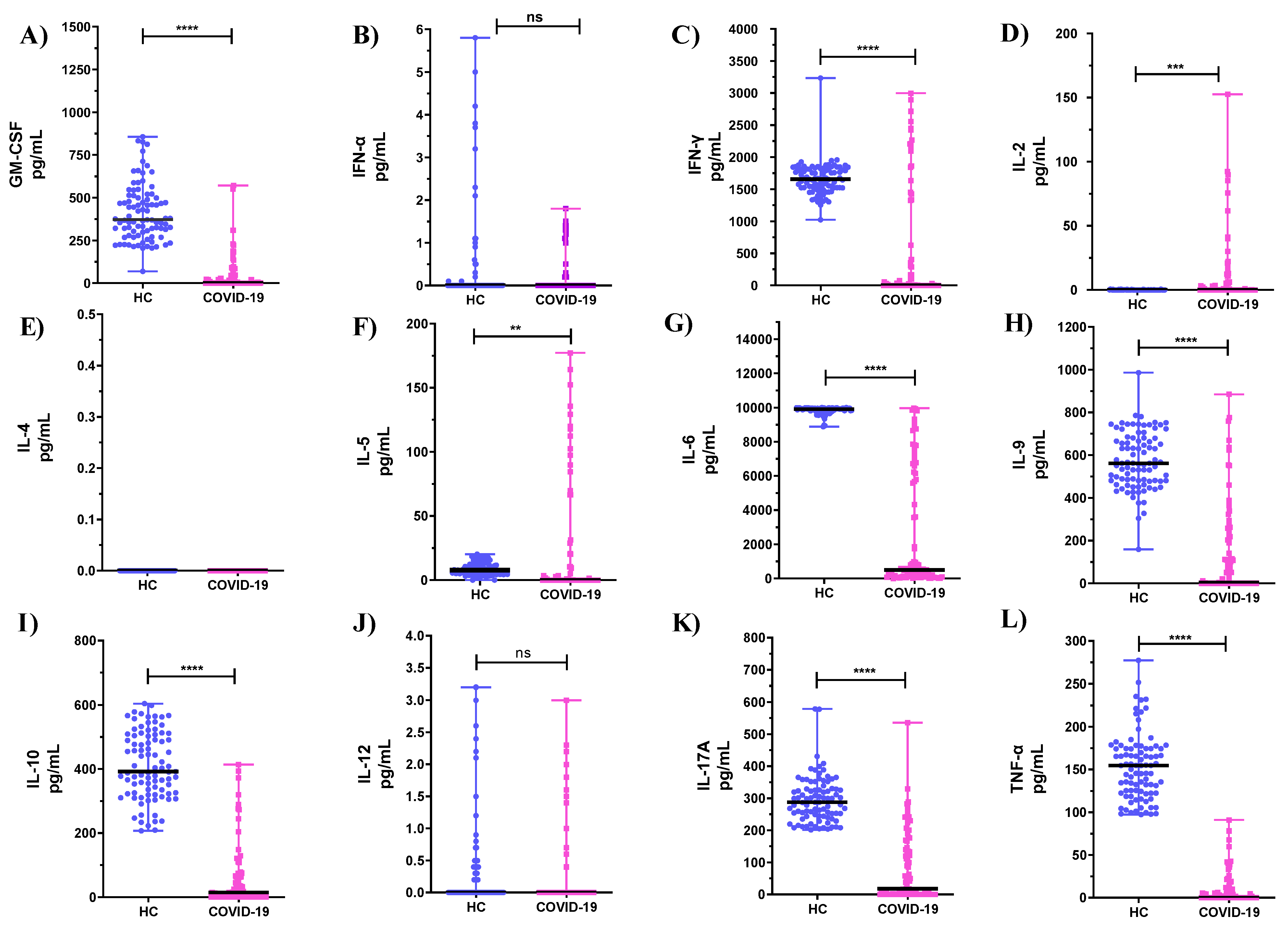
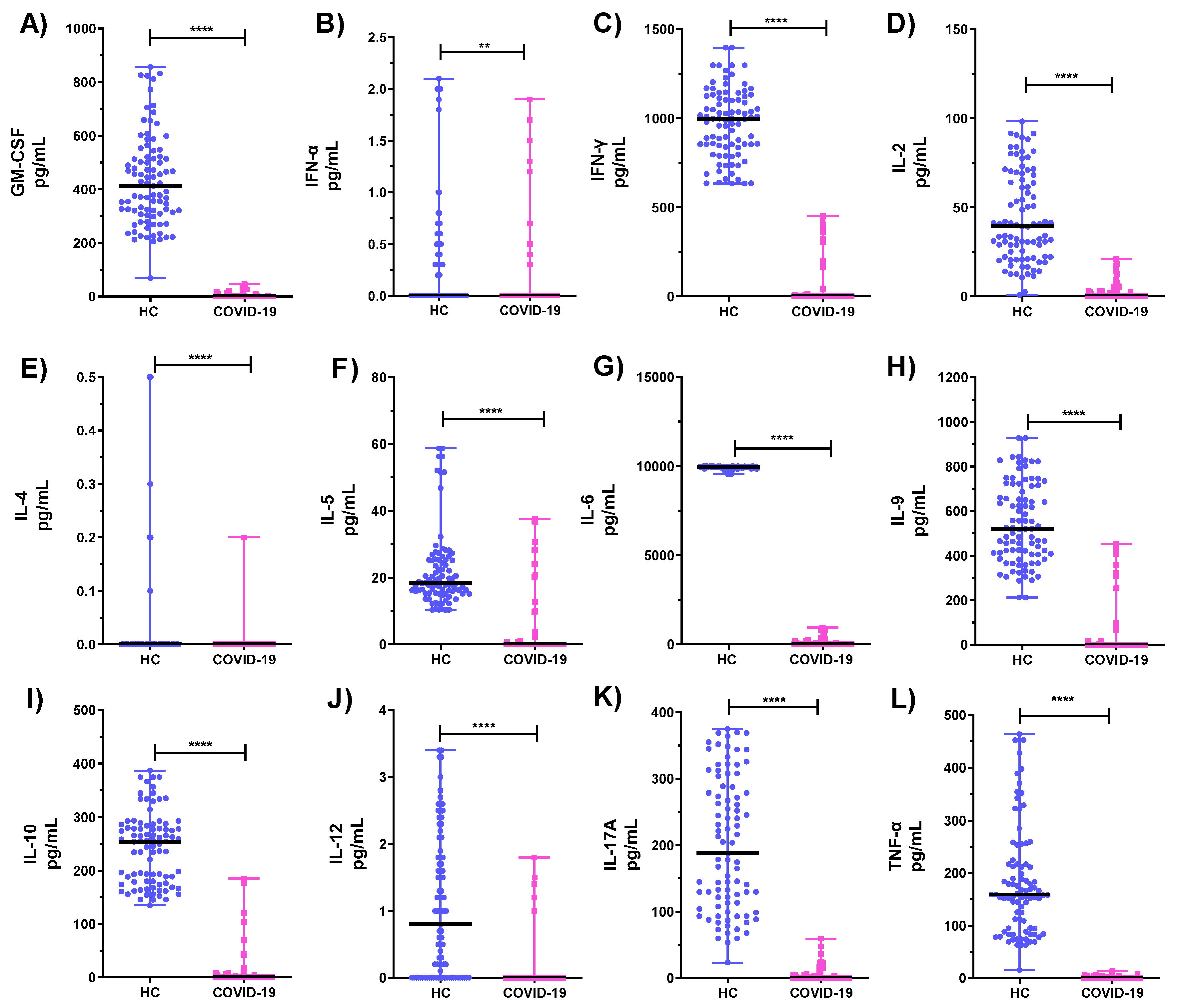
3.2. Comparison of Cytokine Levels Produced by PBMCs and Neutrophils from COVID-19 ICU Patients and Healthy Subjects (HC)
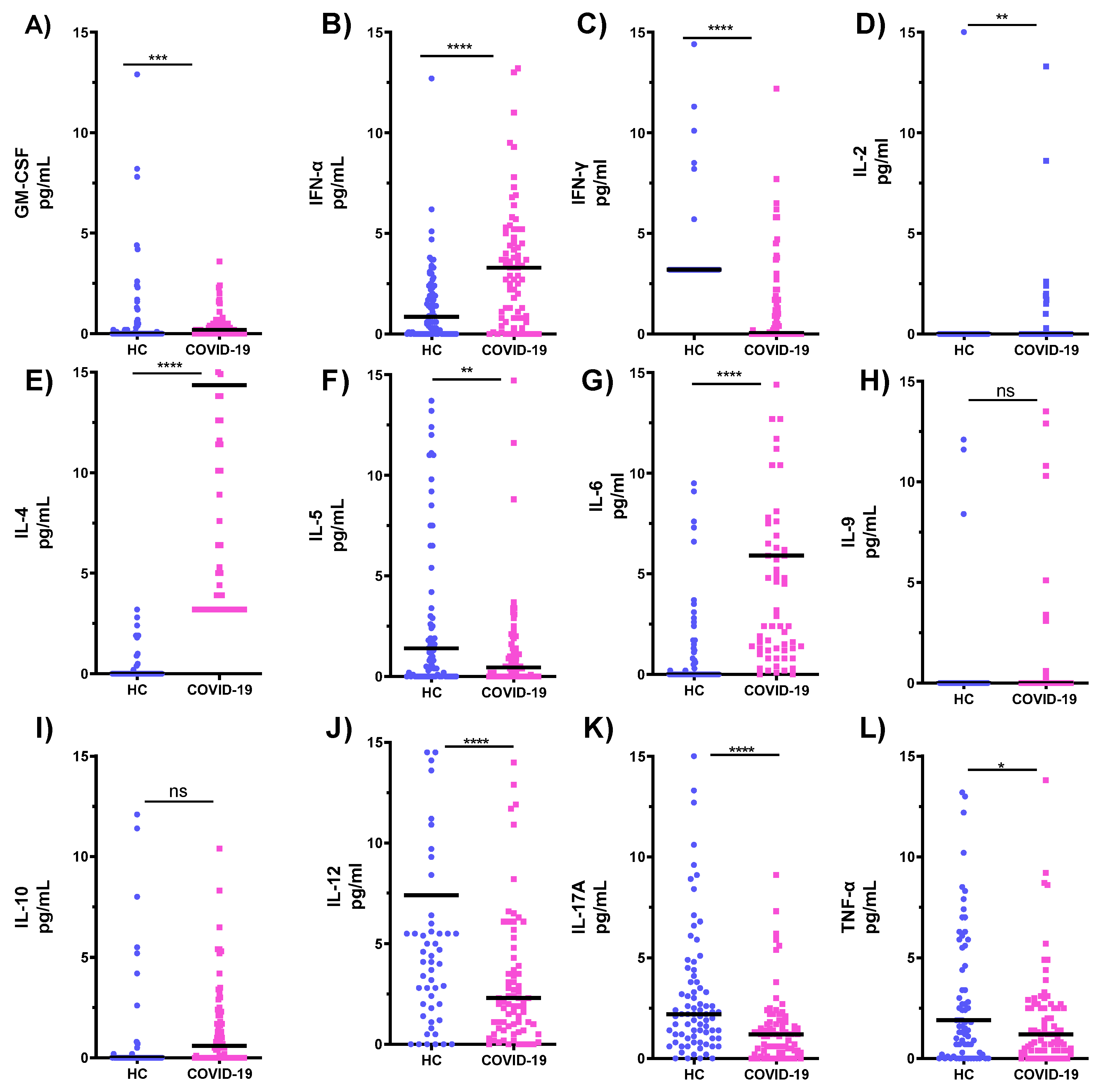
3.3. Comparison of Cytokines Levels in Serum Samples of COVID-19 ICU Patients and Healthy Subjects (HC)
3.4. Th1:Th2 Cytokine Ratios
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/events-as-they-happen (accessed on 31 July 2020).
- Singhal, T. Review of Coronavirus Disease-2019 (COVID-19). Indian J. Pediatr. 2020, 87, 281–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiese, A.; Frati, P.; Del Duca, F.; Santoro, P.; Manetti, A.C.; La Russa, R.; Di Paolo, M.; Turillazzi, E.; Fineschi, V. Myocardial Pathology in COVID-19-Associated Cardiac Injury: A Systematic Review. Diagnostics 2021, 11, 1647. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVI-19 or SARS-CoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 327–331. [Google Scholar] [PubMed]
- Maiese, A.; Baronti, A.; Manetti, A.C.; Di Paolo, M.; Turillazzi, E.; Frati, P.; Fineschi, V. Death after the Administration of COVID-19 Vaccines Approved by EMA: Has a Causal Relationship Been Demonstrated? Vaccines 2022, 10, 308. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current state of the science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X.; Huang, L.; Hoffmann, D.; Lu, M.; Qiu, Y. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11, 2063. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced host response to SARS-CoV-2 drives the development of COVID-19. Cell 2020, 181, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Ghazavi, A.; Ganji, A.; Keshavarzian, N.; Rabiemajd, S.; Mosayebi, G. Cytokine profile and disease severity in patients with COVID-19. Cytokine 2021, 137, 155323. [Google Scholar] [CrossRef] [PubMed]
- Bordon, J.; Aliberti, S.; Fernandez-Botran, R.; Uriarte, S.M.; Rane, M.J.; Duvvuri, P.; Peyrani, P.; Morlacchi, L.C.; Blasi, F.; Ramirez, J.A. Understanding the roles of cytokines, neutrophil activity, and neutrophil apoptosis in the protective versus harmful inflammatory response in pneumonia. Int. J. Infect. Dis. 2013, 17, e76–e83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galani, I.E.; Andreakos, E. Neutrophils in viral infections: Current concepts and caveats. J. Leukoc. Biol. 2015, 98, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Kuri-Cervantes, L.; Pampena, M.B.; Meng, W.; Rosenfeld, A.M.; Ittner, C.A.G.; Weisman, A.R.; Agyekum, R.S.; Mathew, D.; Baxter, A.E.; Vella, L.A.; et al. Comprehensive mapping of immune perturbations associated with severe COVID-19. Sci. Immunol. 2020, 5, eabd7114. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients with Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [Green Version]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Daßler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J. Exp. Med. 2020, 217, e20200652. [Google Scholar] [CrossRef]
- Golonka, R.M.; Saha, P.; Yeoh, B.S.; Chattopadhay, S.; Gewirtz, A.T.; Joe, B.; Vijay-Kumar, M. Harnessing innate immunity to eliminate SARS-CoV-2 and ameliorate COVID-19 disease. Physiol. Genom. 2020, 52, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Tomar, B.; Anders, H.J.; Desai, J.; Mulay, S.R. Neutrophils and Neutrophil Extracellular Traps Drive Necroinflammation in COVID-19. Cells 2020, 9, 1383. [Google Scholar] [CrossRef]
- Al-Herz, W.; Essa, S. Spectrum of Viral Infections Among Primary Immunodeficient Children: Report From a National Registry. Front. Immunol. 2019, 10, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, S.; Qin, H.; Meng, S.; Deng, X.; Lin, H.; Xin, X.; Liang, Y.; Chen, B.; Cui, Y.; et al. Establishment of a quantitative RT-PCR detection of SARS-CoV-2 virus. Eur. J. Med. Res. 2021, 26, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Essa, S.; Siddique, I.; Saad, M.; Raghupathy, R. Modulation of Production of Th1/Th2 Cytokines in Peripheral Blood Mononuclear Cells and Neutrophils by Hepatitis C Virus Infection in Chronically Infected Patients. Pathogens 2021, 10, 1519. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Péré, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Micallef, M.J.; Ohtsuki, T.; Kohno, K.; Tanabe, F.; Ushio, S.; Namba, M. Interferon-γ-inducing factor enhances T helper 1 cytokine production by stimulated human T cells: Synergism with interleukin-12 for interferon-γ production. Eur. J. Immunol. 1996, 26, 1647–1651. [Google Scholar] [CrossRef]
- Tang, M.; Tian, L.; Luo, G.; Yu, X. Interferon-gamma-Mediated osteoimmunology. Front. Immunol. 2018, 9, 1508. [Google Scholar] [CrossRef]
- Lees, J.R. Interferon gamma in autoimmunity: A complicated player on a complex stage. Cytokine 2015, 74, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Aleebrahim-Dehkordi, E.; Molavi, B.; Mokhtari, M.; Deravi, N.; Fathi, M.; Fazel, T.; Mohebalizadeh, M.; Koochaki, P.; Shobeiri, P.; Hasanpour-Dehkordi, A. T helper type (Th1/Th2) responses to SARS-CoV-2 and influenza A (H1N1) virus: From cytokines produced to immune responses. Transpl. Immunol. 2022, 70, 101495. [Google Scholar] [CrossRef]
- Mahallawi, W.H.; Khabour, O.F.; Zhang, Q.; Makhdoum, H.M.; Suliman, B.A. MERS-CoV infection in humans is associated with a pro-inflammatory Th1 and Th17 cytokine profile. Cytokine 2018, 104, 8–13. [Google Scholar] [CrossRef]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 infection: The role of cytokines in COVID-19 disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Li, H.; Lu, X.X.; Xiao, H.; Ren, J.; Zhang, F.R.; Liu, Z.S. Clinical features of severe pediatric patients with coronavirus disease 2019 in Wuhan: A single center’s observational study. World J. Pediatr. 2020, 16, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H. Clinical and immunologic features in severe and moderate Coronavirus Disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell Mol. Immol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas-Ramirez, N.; Woytschak, J.; Boyman, O. Interleukin-2: Biology, Design, and Application. Mod. Trends Immunol. 2015, 36, 763–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.; Lin, X.J.; Leonard, W.J. IL-2 family cytokines: New insights into the complex roles of IL-2 as a broad regulator of T helper cell differentiation. Curr. Opin. Immunol. 2011, 23, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spolski, R.; Li, P.; Leonard, W.J. Biology and Regulation of IL-2: From Molecular Mechanisms to human Therapy. Nat. Rev. Immunol. 2018, 18, 648–659. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Zhang, M.; Yang, C.X.; Zhang, N.; Wang, X.C.; Yang, X.P.; Dong, X.Q.; Zheng, Y.T. Elevated exhaustion levels and reduced functional diversity of T cells in peripheral blood may predict severe progression in COVID-19 patients. Cell Mol. Immunol. 2020, 17, 541–543. [Google Scholar] [CrossRef]
- Bot, A.; Holz, A.; Christen, U.; Wolfe, T.; Temann, A.; Flavell, R.; von Herrath, M. Local IL-4 expression in the lung reduces pulmonary influenza-virus-specific secondary cytotoxic T cell responses. Virology 2000, 269, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhang, X.R.; Ju, Z.Y.; He, W.F. Advances in the research of cytokine storm mechanism induced by Corona Virus Disease 2019 and the corresponding immunotherapies. Zhonghua Shao Shang Za Zhi 2020, 36, E005. [Google Scholar]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. IL-6 trans-signaling via the soluble IL-6 receptor: Importance for the pro-inflammatory activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, B.; Qu, Y.; Chen, Y.; Xiong, J.; Feng, Y.; Men, D.; Huang, Q.; Liu, Y.; Yang, B. Detectable serum SARS-CoV-2 viral load (RNAaemia) is closely associated with drastically elevated interleukin 6 (IL-6) level in critically ill COVID-19 patients. Clin. Infect. Dis. 2020, 71, 1937–1942. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. HLH across specialty collaboration, UK, COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 family of cytokines in health and disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef]
- Wan, S.; Yi, Q.; Fan, S.; Lv, J.; Zhang, X.; Guo, L.; Lang, C.; Xiao, Q.; Xiao, K.; Yi, Z. Characteristics of lymphocyte subsets and cytokines in peripheral blood of 123 hospitalized patients with 2019 novel coronavirus pneumonia (NCP). Br. J. Haematol. 2020, 189, 428–437. [Google Scholar] [CrossRef]
- Pasquereau, S.; Kumar, A.; Herbein, G. Targeting TNF and TNF Receptor Pathway in HIV-1 Infection: From Immune Activation to Viral Reservoirs. Viruses 2017, 9, 64. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Deceased | Transferred | OR (95% CI) | p-Value | |
|---|---|---|---|---|
| Study Sample | 10 (11.4%) | 78 (88.6%) | 0.97 (0.94–1.00) | 0.029 |
| Gender | 0.493 | |||
| F | 4 (40.0%) | 23 (29.5%) | ||
| M | 6 (60.0%) | 55 (70.5%) | 1.59 (0.41–6.18) | |
| Mean age, years | 60.9 | 52.8 | 0.95 (0.91–1.00) | 0.112 |
| Deceased | Transferred | OR (95% CI) | p-Value | |
|---|---|---|---|---|
| Fever: | 0.009 | |||
| No | 5 (50.0%) | 12 (15.4%) | ||
| Yes | 5 (50.0%) | 66 (84.6%) | 5.50 (1.38–21.95) | |
| Cough: | <0.0001 | |||
| No | 9 (90.0%) | 20 (25.6%) | ||
| Yes | 1 (10.0%) | 58 (74.4%) | 26.1 (3.11–219.21) | |
| Pneumonia: | 0.608 | |||
| No | 0 (0.00%) | 2 (2.56%) | ||
| Yes | 10 (100%) | 76 (97.4%) | 1.46 (0.06–32.50) | |
| Other Infections: | 0.0005 | |||
| No | 8 (80.0%) | 20 (25.6%) | ||
| Yes | 2 (20.0%) | 58 (74.4%) | 11.6 (2.27–59.27) |
| Deceased | Transferred | OR (95% CI) | p-Value | |
|---|---|---|---|---|
| HB | 11.6 (2.41) | 15.9 (19.1) | 1.12 (0.84–1.50) | 0.063 |
| WBC | 15.5 (8.26) | 12.1 (6.89) | 0.94 (0.86–1.02) | 0.231 |
| Lymphocytes | 1.06 (0.82) | 0.92 (0.57) | 0.73 (0.29–1.84) | 0.618 |
| PLT | 205 (113) | 259 (107) | 1.00 (1.00–1.01) | 0.180 |
| Lactate | 6.65 (9.25) | 2.24 (1.95) | 0.83 (0.72–0.97) | 0.191 |
| LDH | 1145 (1476) | 850 (1469) | 1.00 (1.00–1.00) | 0.584 |
| D-Dimer | 11521 (19281) | 1601 (2427) | 1.00 (1.00–1.00) | 0.161 |
| CRP | 122 (53.2) | 174 (96.5) | 1.01 (0.99–1.02) | 0.220 |
| Na | 139 (6.11) | 137 (6.34) | 0.95 (0.85–1.06) | 0.374 |
| K | 4.37 (0.65) | 4.17 (0.68) | 0.64 (0.25–1.69) | 0.369 |
| Acid/Base: | 0.316 | |||
| Abnormal | 7 (70.0%) | 38 (48.7%) | ─ | |
| Normal | 3 (30.0%) | 40 (51.3%) | 2.37 (0.59–12.3) | |
| Ferritin | 6272 (13632) | 1011 (772) | 1.00 (1.00–1.00) | 0.388 |
| Urea | 10.8 (4.11) | 8.62 (5.86) | 0.94 (0.85–1.04) | 0.154 |
| Creatinine | 149 (90.7) | 98.8 (70.8) | 0.99 (0.99–1.00) | 0.120 |
| ALT | 163 (379) | 99.7 (344) | 1.00 (1.00–1.00) | 0.627 |
| AST | 318 (763) | 197 (1057) | 1.00 (1.00–1.00) | 0.663 |
| TBill | 26.2 (38.7) | 13.2 (14.6) | 0.98 (0.96–1.00) | 0.318 |
| Trop | 34.2 (32.5) | 14.7 (50.6) | 0.99 (0.98–1.00) | 0.119 |
| BNP | 12909 (21201) | 1859 (3818) | 1.00 (1.00–1.00) | 0.462 |
| Th1/Th2 | PBMCs | Neutrophils | Serum | |||
|---|---|---|---|---|---|---|
| COVID-19 | HC | COVID-19 | HC | COVID-19 | HC | |
| IL-2/IL-4 | 48.23 | 12.56 | 67.565 | 11.011 | 0.0067 | 0.0007 |
| IL-2/IL-5 | 1.53 | 3.55 | 0.253 | 0.493 | 0.8571 | 0.0001 |
| IL-2/IL-9 | 0.475 | 0.02 | 0.009 | 0.05 | 0.8888 | 0.0002 |
| IL-2/IL-10 | 2.7602 | 0.04 | 0.081 | 0.33 | 0.48 | 0.0001 |
| TNF/IL-4 | 33.1 | 342.6 | 22.5 | 92.6 | 0.1179 | 0.4724 |
| TNF /IL-5 | 2.4 | 58.20 | 1.8 | 38.6 | 1.5 | 0.7894 |
| TNF/IL-9 | 0.326 | 0.773 | 0.2 | 0.301 | 1.5 | 1.7142 |
| TNF/IL-10 | 0.372 | 0.174 | 2.7 | 0.897 | 0.84 | 0.3208 |
| IL-6/IL-4 | 792.6 | 7,834.8 | 327.1 | 5,144.6 | 4.3 | 0.0866 |
| IL-6/IL-5 | 58.2 | 1143.4 | 122.6 | 451.20 | 54.7 | 0.1447 |
| IL-6/IL-9 | 7.80 | 17.844 | 23.6 | 18.317 | 53.9 | 0.3143 |
| IL-6/IL-10 | 13.92 | 45.452 | 239.3 | 40.865 | 30.64 | 0.0588 |
| IL-12/IL-4 | 0.283 | 0.00005 | 296.5 | 328.10 | 0.365 | 3.968 |
| IL-12 /IL-5 | 0.021 | 0.00001 | 0.084 | 0.0990 | 4.643 | 6.632 |
| IL-12/IL-9 | 0.003 | 0.000005 | 0.0003 | 0.0021 | 4.814 | 14.4 |
| IL-12/IL-10 | 0.016 | 0.000003 | 0.005 | 0.0067 | 2.6 | 2.695 |
| IFN-α/IL-4 | 0.347 | 0.8073 | 0.075 | 0.166 | 0.258 | 0.126 |
| IFN-α/IL-5 | 0.025 | 0.1096 | 0.0038 | 0.017 | 3.286 | 0.211 |
| IFN-α/IL-9 | 0.0003 | 0.0009 | 0.00005 | 0.001 | 3.407 | 0.457 |
| IFN-α/IL-10 | 0.0011 | 0.025 | 0.00056 | 0.001 | 1.84 | 0.085 |
| IFN-γ/IL-4 | 274.2 | 1177.3 | 443,600 | 533.6 | 0.124 | 3.772 |
| IFN- γ/IL-5 | 20.1 | 209.9 | 161.7 | 43.8 | 1.572 | 6.303 |
| IFN- γ/IL-9 | 2.7 | 3.0 | 1.6 | 1.8 | 1.629 | 13.69 |
| IFN- γ/IL-10 | 1.7 | 4.2 | 17.3 | 4.0 | 0.88 | 2.561 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Essa, S.; Shamsah, M.; Alsarraf, A.H.; Esmaeil, A.; Al-Shammasi, A.; Raghupathy, R. Influence of SARS-COV-2 Infection on Cytokine Production by Mitogen-Stimulated Peripheral Blood Mononuclear Cells and Neutrophils in COVID-19 Intensive Care Unit Patients. Microorganisms 2022, 10, 2194. https://doi.org/10.3390/microorganisms10112194
Essa S, Shamsah M, Alsarraf AH, Esmaeil A, Al-Shammasi A, Raghupathy R. Influence of SARS-COV-2 Infection on Cytokine Production by Mitogen-Stimulated Peripheral Blood Mononuclear Cells and Neutrophils in COVID-19 Intensive Care Unit Patients. Microorganisms. 2022; 10(11):2194. https://doi.org/10.3390/microorganisms10112194
Chicago/Turabian StyleEssa, Sahar, Mohammed Shamsah, Abdalaziz H. Alsarraf, Ali Esmaeil, Ahmed Al-Shammasi, and Raj Raghupathy. 2022. "Influence of SARS-COV-2 Infection on Cytokine Production by Mitogen-Stimulated Peripheral Blood Mononuclear Cells and Neutrophils in COVID-19 Intensive Care Unit Patients" Microorganisms 10, no. 11: 2194. https://doi.org/10.3390/microorganisms10112194