
Long-Term Effects of Single-Dose Cephalosporin or Macrolide Use on the Prevalence of AmpC and Extended-Spectrum β-Lactamase Producing Escherichia coli in the Feces of Beef Cattle
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Bacterial Quantification and Confirmation
2.3. Detection of ESBL/AmpC Producing E. coli
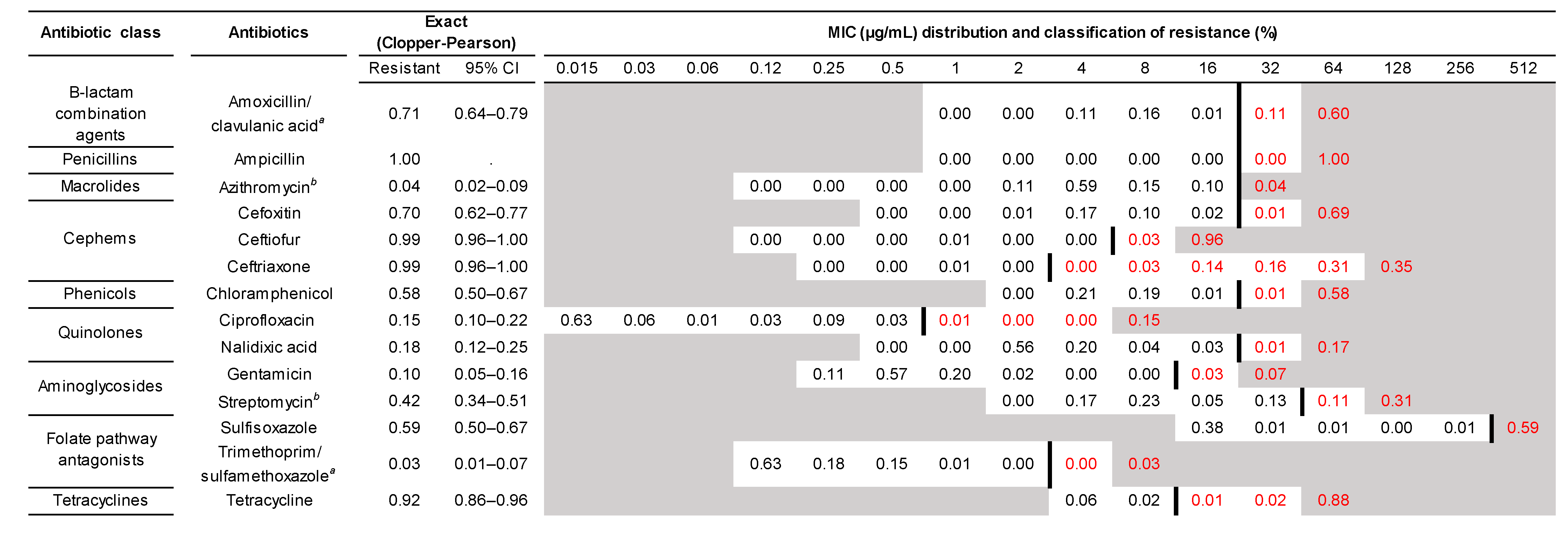
2.3.1. Phenotypic Antibiotic Susceptibility
2.3.2. Whole-Genome Sequencing and Bioinformatics
2.4. Statistical Analysis
3. Results
3.1. Count Data
3.2. ESBL/AmpC E. coli Populations
3.3. Genomic Characterization of AmpC/ESBL Producing E. coli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogliani, C. Restricting Antimicrobial Use in Food Animals: Lessons from Europe. Microbe jMag. 2011, 6, 274–279. [Google Scholar] [CrossRef]
- WHO. WHO Guidelines On Use of Medically Important Antimicrobials in Food-Producing Animals; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Anderson, A.D.; Nelson, J.M.; Rossiter, S.; Angulo, F.J. Public health consequences of use of antimicrobial agents in food animals in the United States. Microb. Drug. Resist. 2003, 9, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angulo, F.J.; Baker, N.L.; Olsen, S.J.; Anderson, A.; Barrett, T.J. Antimicrobial use in agriculture: Controlling the transfer of antimicrobial resistance to humans. Semin. Pediatr. Infect. Dis. 2004, 15, 78–85. [Google Scholar] [CrossRef]
- Angulo, F.J.; Mølbak, K. Human Health Consequences of Antimicrobial Drug—Resistant Salmonella and Other Foodborne Pathogens. Clin. Infect. Dis. 2005, 41, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L. Clinical practice. Bacterial diarrhea. N. Eng. J. Med. 2009, 361, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- CDC. Antibiotic Resistance Threats in the United States, 2013; Centers for Disease Control and Prevention, U.S. Department of Health and Human Services: Atlanta, GA, USA, 2013.
- CDC. Antibiotic Resistance Threats in the United States, 2019; Centers for Disease Control and Prevention, U.S. Department of Health and Human Services: Atlanta, GA, USA, 2019.
- Marshall, W.F.; Blair, J.E. The cephalosporins. Mayo. Clin. Proc. 1999, 74, 187–195. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th revision; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Carattoli, A. Animal reservoirs for extended spectrum beta-lactamase producers. Clin. Microbiol. Infect. 2008, 14 (Suppl. S1), 117–123. [Google Scholar] [CrossRef] [Green Version]
- Winokur, P.L.; Vonstein, D.L.; Hoffman, L.J.; Uhlenhopp, E.K.; Doern, G.V. Evidence for transfer of CMY-2 AmpC beta-lactamase plasmids between Escherichia coli and Salmonella isolates from food animals and humans. Antimicrob. Agents. Chemother. 2001, 45, 2716–2722. [Google Scholar] [CrossRef] [Green Version]
- Von Wintersdorff, C.J.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.; Wolffs, P.F. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Neuwirth, C.; Siebor, E.; Pechinot, A.; Duez, J.M.; Pruneaux, M.; Garel, F.; Kazmierczak, A.; Labia, R. Evidence of in vivo transfer of a plasmid encoding the extended-spectrum beta-lactamase TEM-24 and other resistance factors among different members of the family Enterobacteriaceae. J. Clin. Microbiol. 2001, 39, 1985–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; White, D.G.; McDermott, P.F.; Friedman, S.; English, L.; Ayers, S.; Meng, J.; Maurer, J.J.; Holland, R.; Walker, R.D. Identification and expression of cephamycinase bla(CMY) genes in Escherichia coli and Salmonella isolates from food animals and ground meat. Antimicrob. Agents. Chemother. 2001, 45, 3647–3650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum beta-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, D.A.; Li, C.; Mukherjee, S.; Hsu, C.H.; Bodeis Jones, S.; Gaines, S.A.; Kabera, C.; Loneragan, G.H.; Torrence, M.; Harhay, D.M.; et al. Whole-Genome Sequence Analysis of CTX-M Containing Escherichia coli Isolates from Retail Meats and Cattle in the United States. Microb. Drug. Resist. 2018, 24, 939–948. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Mehrotra, M.; Ghimire, S.; Adewoye, L. beta-Lactam resistance and beta-lactamases in bacteria of animal origin. Vet. Microbiol. 2007, 121, 197–214. [Google Scholar] [CrossRef]
- Mollenkopf, D.F.; Kleinhenz, K.E.; Funk, J.A.; Gebreyes, W.A.; Wittum, T.E. Salmonella enterica and Escherichia coli harboring blaCMY in retail beef and pork products. Foodborne Pathog. Dis. 2011, 8, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration; Department of Human and Health Services. New animal drugs; cephalosporin drugs; extralabel animal drug use; order of prohibition. In Fed. Regist.; 2012; 77, pp. 735–745. [Google Scholar]
- Food and Drug Administration. 2019 Summary Report on Antimicrobials Sold or Distributed for Use in Food-Producing Animals; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2020.
- USDA-APHIS-NAHMS. Feedlot 2011, Part IV: Health and SSS on U.S. Feedlots with a Capacity of 1000 or More Head; United States Department of Agriculture, Animal and Plant Health Inspection Service, National Animal Health Monitoring System: Fort Collins, CO, USA, 2013.
- Hibbard, B.; Robb, E.J.; Chester, S.T., Jr.; Dame, K.J.; Moseley, W.W.; Bryson, W.L. Dose determination and confirmation for ceftiofur crystalline-free acid administered in the posterior aspect of the ear for control and treatment of bovine respiratory disease. Vet. Ther. 2002, 3, 22–30. [Google Scholar]
- Freedom Of Information (FOI) Summary Original New Animal Drug Application (NADA) for EXCEDE® Ceftiofur Crystalline Free Acid Sterile Suspension (previosly known as NAXCEL XT®), NADA 141-209. Available online: https://animaldrugsatfda.fda.gov/adafda/app/search/public/document/downloadFoi/729 (accessed on 28 September 2022).
- Lowrance, T.C.; Loneragan, G.H.; Kunze, D.J.; Platt, T.M.; Ives, S.E.; Scott, H.M.; Norby, B.; Echeverry, A.; Brashears, M.M. Changes in antimicrobial susceptibility in a population of Escherichia coli isolated from feedlot cattle administered ceftiofur crystalline-free acid. Am. J. Vet. Res. 2007, 68, 501–507. [Google Scholar] [CrossRef]
- Kanwar, N.; Scott, H.M.; Norby, B.; Loneragan, G.H.; Vinasco, J.; Cottell, J.L.; Chalmers, G.; Chengappa, M.M.; Bai, J.; Boerlin, P. Impact of treatment strategies on cephalosporin and tetracycline resistance gene quantities in the bovine fecal metagenome. Sci. Rep. 2014, 4, 5100. [Google Scholar] [CrossRef] [Green Version]
- Kanwar, N.; Scott, H.M.; Norby, B.; Loneragan, G.H.; Vinasco, J.; McGowan, M.; Cottell, J.L.; Chengappa, M.M.; Bai, J.; Boerlin, P. Effects of ceftiofur and chlortetracycline treatment strategies on antimicrobial susceptibility and on tet(A), tet(B), and bla CMY-2 resistance genes among E. coli isolated from the feces of feedlot cattle. PLoS ONE 2013, 8, e80575. [Google Scholar] [CrossRef]
- Alali, W.Q.; Scott, H.M.; Norby, B.; Gebreyes, W.; Loneragan, G.H. Quantification of the bla(CMY-2) in feces from beef feedlot cattle administered three different doses of ceftiofur in a longitudinal controlled field trial. Foodborne Pathog. Dis. 2009, 6, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Weinroth, M.D.; Scott, H.M.; Norby, B.; Loneragan, G.H.; Noyes, N.R.; Rovira, P.; Doster, E.; Yang, X.; Woerner, D.R.; Morley, P.S.; et al. Effects of ceftiofur and chlortetracycline on the resistomes of feedlot cattle. Appl. Environ. Microbiol. 2018, 84, e00610-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aidara-Kane, A.; Angulo, F.J.; Conly, J.M.; Minato, Y.; Silbergeld, E.K.; McEwen, S.A.; Collignon, P.J.; Group, W.H.O.G.D. World Health Organization (WHO) guidelines on use of medically important antimicrobials in food-producing animals. Antimicrob. Resist. Infect. Control. 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, E.A.; Jordan, E.R.; Garcia, J.A.; Hagevoort, G.R.; Norman, K.N.; Lawhon, S.D.; Pineiro, J.M.; Scott, H.M. Effects of two-dose ceftiofur treatment for metritis on the temporal dynamics of antimicrobial resistance among fecal Escherichia coli in Holstein-Friesian dairy cows. PLoS ONE 2019, 14, e0220068. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Taylor, B.; Keane, J.A. Multilocus sequence typing by blast from de novo assemblies against PubMLST. J. Open Source Softw. 2016, 1, 118. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Allesoe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Arredondo-Alonso, S.; Rogers, M.R.C.; Braat, J.C.; Verschuuren, T.D.; Top, J.; Corander, J.; Willems, R.J.L.; Schurch, A.C. mlplasmids: A user-friendly tool to predict plasmid- and chromosome-derived sequences for single species. Microb. Genom. 2018, 4, e000224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kummerer, K. Resistance in the environment. J. Antimicrob. Chemother. 2004, 54, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Agga, G.E.; Arthur, T.M.; Durso, L.M.; Harhay, D.M.; Schmidt, J.W. Antimicrobial-Resistant Bacterial Populations and Antimicrobial Resistance Genes Obtained from Environments Impacted by Livestock and Municipal Waste. PLoS ONE 2015, 10, e0132586. [Google Scholar] [CrossRef]
- Cormier, A.C.; Chalmers, G.; McAllister, T.A.; Cook, S.; Zaheer, R.; Scott, H.M.; Booker, C.; Read, R.; Boerlin, P. Extended-Spectrum-Cephalosporin Resistance Genes in Escherichia coli from Beef Cattle. Antimicrob. Agents Chemother. 2016, 60, 1162–1163. [Google Scholar] [CrossRef] [Green Version]
- Subbiah, M.; Shah, D.H.; Besser, T.E.; Ullman, J.L.; Call, D.R. Urine from treated cattle drives selection for cephalosporin resistant Escherichia coli in soil. PLoS ONE 2012, 7, e48919. [Google Scholar] [CrossRef]
- Lee, S.; Teng, L.; DiLorenzo, N.; Weppelmann, T.A.; Jeong, K.C. Prevalence and Molecular Characteristics of Extended-Spectrum and AmpC beta-Lactamase Producing Escherichia coli in Grazing Beef Cattle. Front. Microbiol. 2019, 10, 3076. [Google Scholar] [CrossRef]
- Schmidt, J.W.; Griffin, D.; Kuehn, L.A.; Brichta-Harhay, D.M. Influence of therapeutic ceftiofur treatments of feedlot cattle on fecal and hide prevalences of commensal Escherichia coli resistant to expanded-spectrum cephalosporins, and molecular characterization of resistant isolates. Appl. Environ. Microbiol. 2013, 79, 2273–2283. [Google Scholar] [CrossRef] [Green Version]
- Levent, G.; Schlochtermeier, A.; Ives, S.E.; Norman, K.N.; Lawhon, S.D.; Loneragan, G.H.; Anderson, R.C.; Vinasco, J.; den Bakker, H.C.; Scott, H.M. High-Resolution Genomic Comparisons within Salmonella enterica Serotypes Derived from Beef Feedlot Cattle: Parsing the Roles of Cattle Source, Pen, Animal, Sample Type, and Production Period. Appl. Environ. Microbiol. 2021, 87, e0048521. [Google Scholar] [CrossRef] [PubMed]
- Alexander, T.W.; Yanke, L.J.; Topp, E.; Olson, M.E.; Read, R.R.; Morck, D.W.; McAllister, T.A. Effect of subtherapeutic administration of antibiotics on the prevalence of antibiotic-resistant Escherichia coli bacteria in feedlot cattle. Appl. Environ. Microbiol. 2008, 74, 4405–4416. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.A.; Ossa-Trujillo, C.; Vinasco, J.; Jordan, E.R.; Garcia Buitrago, J.A.; Hagevoort, R.; Norman, K.N.; Lawhon, S.D.; Pineiro, J.M.; Levent, G.; et al. Use of critically important antimicrobial classes early in life may adversely impact bacterial resistance profiles during adult years: Potential co-selection for plasmid-borne fluoroquinolone and macrolide resistance via extended-spectrum beta-lactam use in dairy cattle. Lett. Appl. Microbiol. 2021, 72, 220–224. [Google Scholar] [CrossRef]
- Platt, T.M.; Loneragan, G.H.; Scott, H.M.; Norby, B.; Thomson, D.U.; Brown, M.S.; Ives, S.E.; Brashears, M.M. Antimicrobial susceptibility of enteric bacteria recovered from feedlot cattle administered chlortetracycline in feed. Am. J. Vet. Res. 2008, 69, 988–996. [Google Scholar] [CrossRef]
- Poole, T.L.; Callaway, T.R.; Norman, K.N.; Scott, H.M.; Loneragan, G.H.; Ison, S.A.; Beier, R.C.; Harhay, D.M.; Norby, B.; Nisbet, D.J. Transferability of antimicrobial resistance from multidrug-resistant Escherichia coli isolated from cattle in the USA to E. coli and Salmonella Newport recipients. J. Glob. Antimicrob. Resist. 2017, 11, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Levent, G.; Schlochtermeier, A.; Ives, S.E.; Norman, K.N.; Lawhon, S.D.; Loneragan, G.H.; Anderson, R.C.; Vinasco, J.; Scott, H.M. Population dynamics of Salmonella enterica within beef cattle cohorts followed from single-dose metaphylactic antibiotic treatment until slaughter. Appl. Environ. Microbiol. 2019, 85, e01386-19. [Google Scholar] [CrossRef]
- Call, D.R.; Singer, R.S.; Meng, D.; Broschat, S.L.; Orfe, L.H.; Anderson, J.M.; Herndon, D.R.; Kappmeyer, L.S.; Daniels, J.B.; Besser, T.E. blaCMY-2-positive IncA/C plasmids from Escherichia coli and Salmonella enterica are a distinct component of a larger lineage of plasmids. Antimicrob. Agents Chemother. 2010, 54, 590–596. [Google Scholar] [CrossRef] [Green Version]
- Cottell, J.L.; Kanwar, N.; Castillo-Courtade, L.; Chalmers, G.; Scott, H.M.; Norby, B.; Loneragan, G.H.; Boerlin, P. blaCTX-M-32 on an IncN plasmid in Escherichia coli from beef cattle in the United States. Antimicrob. Agents Chemother. 2013, 57, 1096–1097. [Google Scholar] [CrossRef] [Green Version]
- Subbiah, M.; Top, E.M.; Shah, D.H.; Call, D.R. Selection pressure required for long-term persistence of blaCMY-2-positive IncA/C plasmids. Appl. Environ. Microbiol. 2011, 77, 4486–4493. [Google Scholar] [CrossRef] [Green Version]
- Webb, H.E.; Bugarel, M.; den Bakker, H.C.; Nightingale, K.K.; Granier, S.A.; Scott, H.M.; Loneragan, G.H. Carbapenem-Resistant Bacteria Recovered from Faeces of Dairy Cattle in the High Plains Region of the USA. PLoS ONE 2016, 11, e0147363. [Google Scholar] [CrossRef] [Green Version]
- Markland, S.; Weppelmann, T.A.; Ma, Z.; Lee, S.; Mir, R.A.; Teng, L.; Ginn, A.; Lee, C.; Ukhanova, M.; Galindo, S.; et al. High Prevalence of Cefotaxime Resistant Bacteria in Grazing Beef Cattle: A Cross Sectional Study. Front. Microbiol. 2019, 10, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunrinu, O.J.; Norman, K.N.; Vinasco, J.; Levent, G.; Lawhon, S.D.; Fajt, V.R.; Volkova, V.V.; Gaire, T.; Poole, T.L.; Genovese, K.J.; et al. Can the use of older-generation beta-lactam antibiotics in livestock production over-select for beta-lactamases of greatest consequence for human medicine? An in vitro experimental model. PLoS ONE 2020, 15, e0242195. [Google Scholar] [CrossRef] [PubMed]
- Hope, K.J.; Apley, M.D.; Schrag, N.F.D.; Lubbers, B.V.; Singer, R.S. Antimicrobial use in 22 U.S. beef feedyards: 2016–2017. Zoonoses Public Health 2020, 67 (Suppl. S1), 94–110. [Google Scholar] [CrossRef] [PubMed]
- De Donder, K.D.; Apley, M.D. A review of the expected effects of antimicrobials in bovine respiratory disease treatment and control using outcomes from published randomized clinical trials with negative controls. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 97–111. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
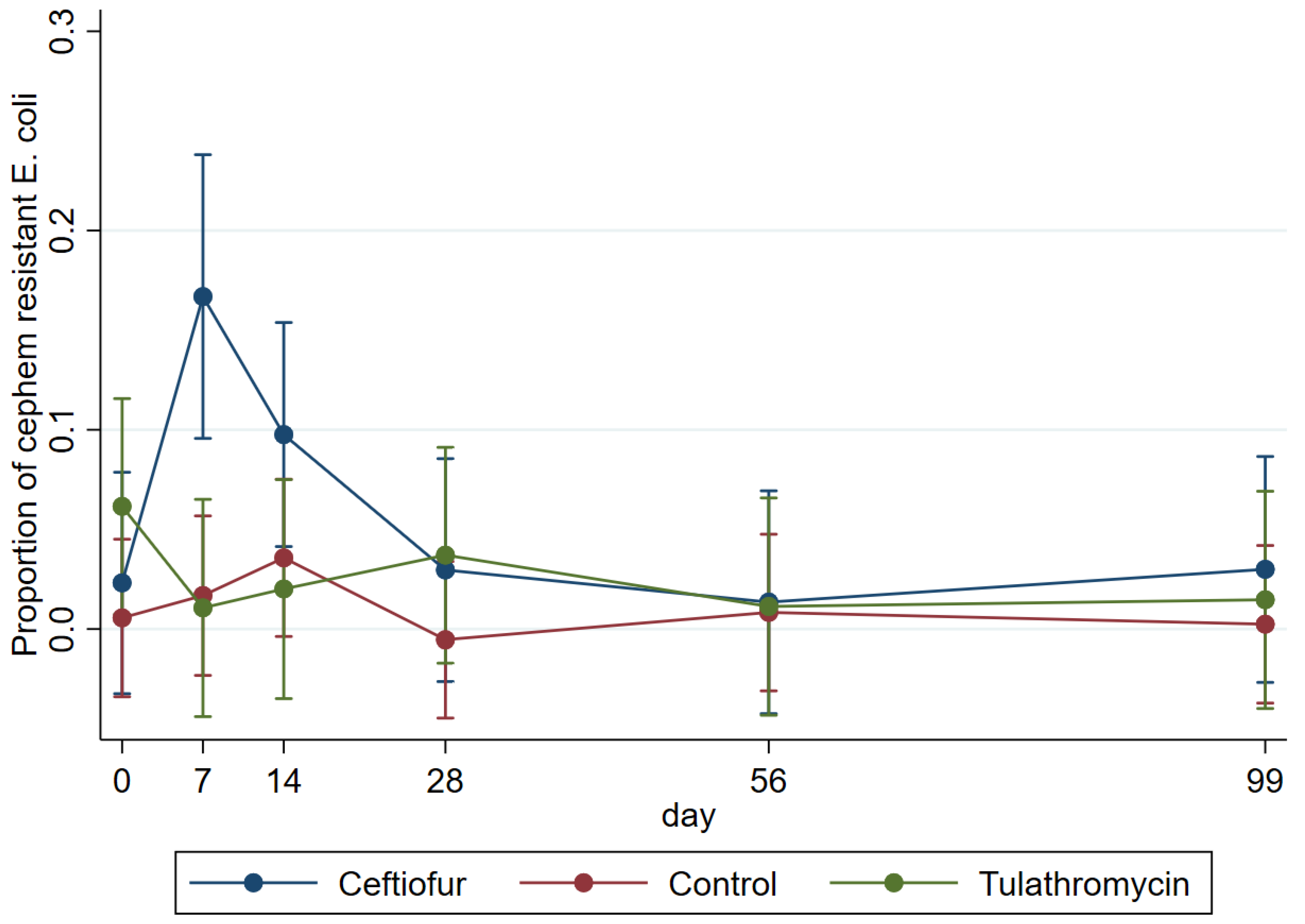
| Day 0 | Day 7 | Day 14 | Day 28 | Day 56 | Day 99 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AmpC | ESBL | AmpC | ESBL | AmpC | ESBL | AmpC | ESBL | AmpC | ESBL | AmpC | ESBL | |
| Ceftiofur | 0.10 (4/40) | 0.00 (0/40) | 0.13 (5/40) | 0.15 (6/40) | 0.13 (5/40) | 0.05 (2/40) | 0.08 (3/40) | 0.05 (2/40) | 0.21 (8/39) | 0.15 (6/39) | 0.18 (7/39) | 0.00 (0/39) |
| ESBL/AmpC | 0.10 (4/40) | 0.28 (11/40) | 0.18 (7/40) | 0.13 (5/40) | 0.36 (14/39) | 0.18 (7/39) | ||||||
| Control | 0.04 (2/53) | 0.09 (5/53) | 0.02 (1/54) | 0.04 (2/54) | 0.13 (7/54) | 0.02 (1/54) | 0.11 (6/54) | 0.00 (0/54) | 0.19 (10/54) | 0.02 (1/54) | 0.16 (9/54) | 0.02 (1/54) |
| ESBL/AmpC | 0.13 (7/53) | 0.06 (3/54) | 0.15 (8/54) | 0.11 (6/54) | 0.22 (12/54) * | 0.19 (10/54) | ||||||
| Tulathromycin | 0.20 (8/40) | 0.08 (3/40) | 0.13 (5/40) | 0.03 (1/40) | 0.23 (9/40) | 0.03 (1/40) | 0.13 (5/40) | 0.13 (5/40) | 0.00 (0/39) | 0.08 (3/39) | 0.18 (7/39) | 0.03 (1/39) |
| ESBL/AmpC | 0.28 (11/40) | 0.15 (6/40) | 0.25 (10/40) | 0.25 (10/40) | 0.08 (3/39) | 0.21 (8/39) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levent, G.; Schlochtermeier, A.; Vinasco, J.; Jennings, J.; Richeson, J.; Ives, S.E.; Norman, K.N.; Lawhon, S.D.; Loneragan, G.H.; Scott, H.M. Long-Term Effects of Single-Dose Cephalosporin or Macrolide Use on the Prevalence of AmpC and Extended-Spectrum β-Lactamase Producing Escherichia coli in the Feces of Beef Cattle. Microorganisms 2022, 10, 2071. https://doi.org/10.3390/microorganisms10102071
Levent G, Schlochtermeier A, Vinasco J, Jennings J, Richeson J, Ives SE, Norman KN, Lawhon SD, Loneragan GH, Scott HM. Long-Term Effects of Single-Dose Cephalosporin or Macrolide Use on the Prevalence of AmpC and Extended-Spectrum β-Lactamase Producing Escherichia coli in the Feces of Beef Cattle. Microorganisms. 2022; 10(10):2071. https://doi.org/10.3390/microorganisms10102071
Chicago/Turabian StyleLevent, Gizem, Ashlynn Schlochtermeier, Javier Vinasco, Jenny Jennings, John Richeson, Samuel E. Ives, Keri N. Norman, Sara D. Lawhon, Guy H. Loneragan, and H. Morgan Scott. 2022. "Long-Term Effects of Single-Dose Cephalosporin or Macrolide Use on the Prevalence of AmpC and Extended-Spectrum β-Lactamase Producing Escherichia coli in the Feces of Beef Cattle" Microorganisms 10, no. 10: 2071. https://doi.org/10.3390/microorganisms10102071