
In Vitro and In Silico Analysis of the Inhibitory Activity of EGCG-Stearate against Herpes Simplex Virus-2


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. HSV-2 Maintenance
2.3. Preparation of Polyphenols
2.4. Cell Viability
2.5. Cell Proliferation
2.6. Antiviral Assay
2.7. Cell Morphology
2.8. Plaque Assay
2.9. Attachment Assay
2.10. Penetration Assay
2.11. Confocal Microscopy
2.12. Primer Design and Quantitative Polymerase Chain Reaction (qPCR)
2.13. Bioinformatic Analyses
2.14. Statistical Analyses
3. Results
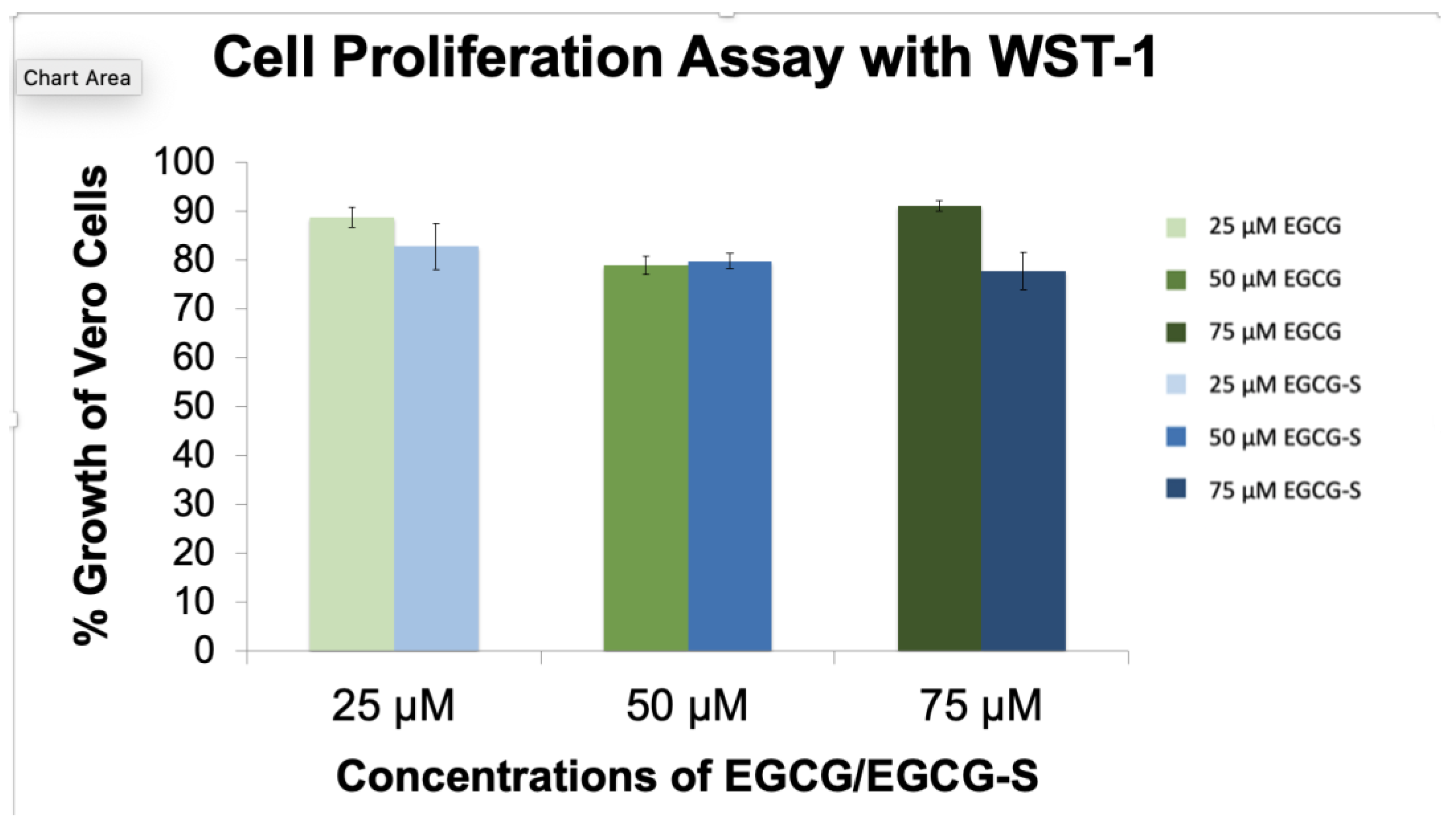
3.1. No Toxic Effect of EGCG and EGCG-S on Vero Cell Viability and Proliferation
3.2. Infection of Vero Cells with Treated HSV-2 Reduces Cytopathic Effects
3.3. EGCG and EGCG-S Inhibit HSV-2 Plaque Formation
3.4. EGCG and EGCG-S Affect the Attachment of HSV-2
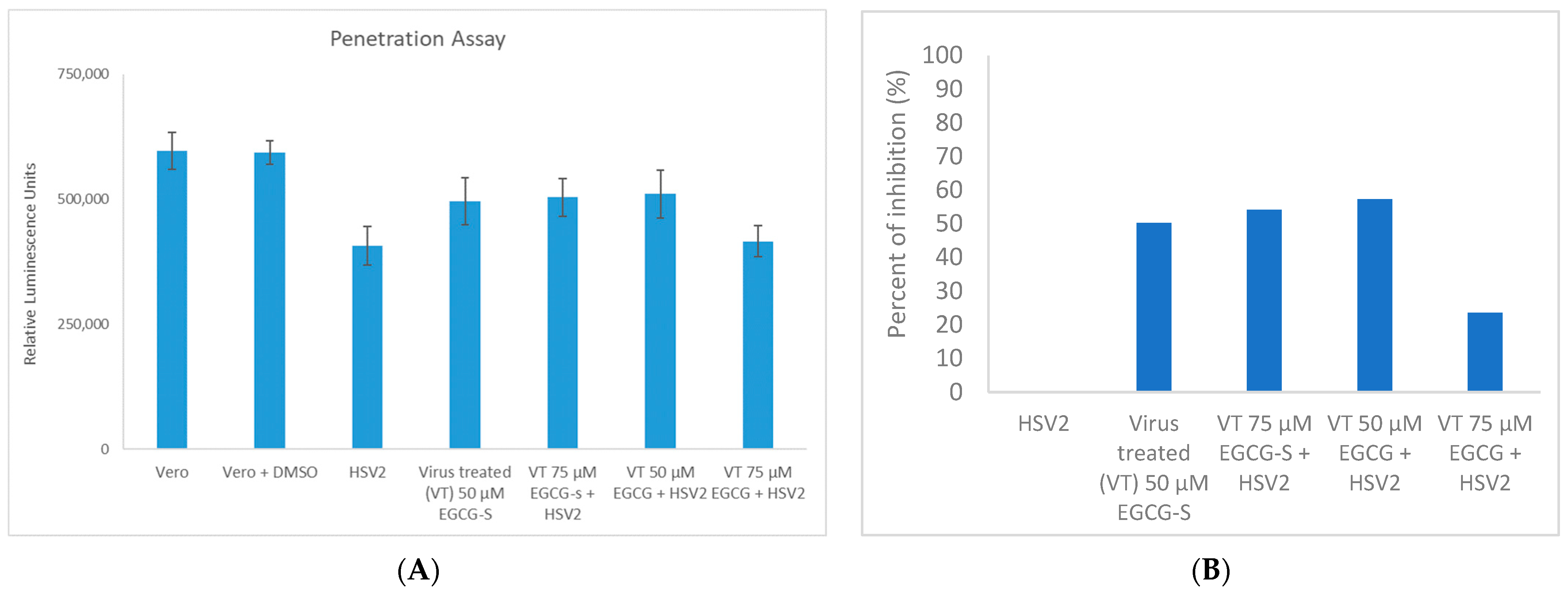
3.5. EGCG and EGCG-S Reduce HSV-2 Penetration in Vero Cells
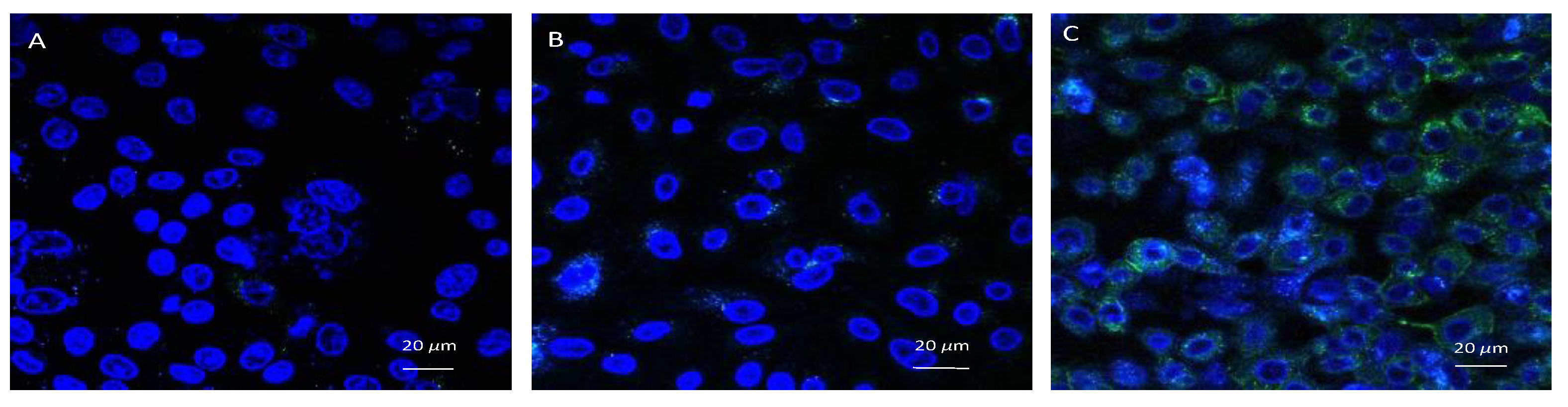
3.6. Confocal Microscopic Observation
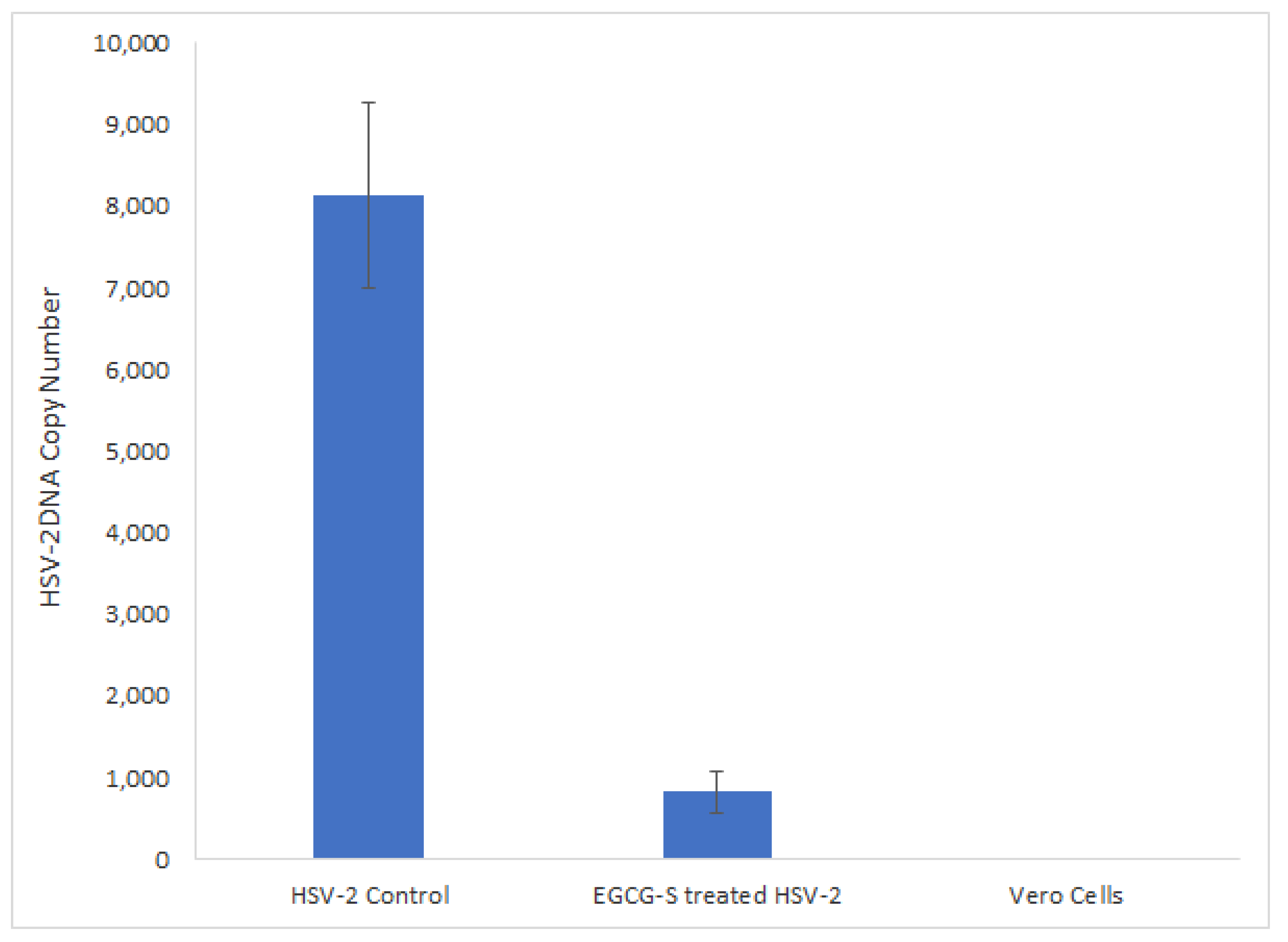
3.7. qPCR Quantitative Analysis of EGCG-S Treated or Non-Treated HSV-2
3.8. Bioinformatic Analysis of EGCG-S Binding to Glycoprotein D
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roizman, B. The Family Herpesviridae. In The Human Herpesviruses; Roizman, B., Whitley, R.J., Lopez, C., Eds.; Raven Press: New York, NY, USA, 1993. [Google Scholar]
- Roizman, B.; Pellett, P.E. The family Herpesviridae: A brief introduction. In Fields Virology, 4th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2001; Volume 2. [Google Scholar]
- Akhtar, J.; Shukla, D. Viral entry mechanisms: Cellular and viral mediators of herpes simplex virus entry. FEBS J. 2009, 276, 7228–7236. [Google Scholar] [CrossRef]
- Agelidis, A.M.; Shukla, D. Cell entry mechanisms of HSV: What we have learned in recent years. Future Virol. 2015, 10, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.J.; Akhtar, J.; Desai, P.; Shukla, D. A role for heparan sulfate in viral surfing. Biochem. Biophys. Res. Commun. 2010, 391, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Roizman, B.; Whitley, R.J. An inquiry into the molecular basis of HSV latency and reactivation. Annu. Rev. Microbiol. 2013, 67, 355–374. [Google Scholar] [CrossRef] [Green Version]
- Elion, G.B. Acyclovir: Discovery, mechanism of action and selectivity. J. Med. Virol. 1993, (Suppl. S1), 2–6. [Google Scholar] [CrossRef]
- Frobert, E.; Cortay, J.C.; Ooka, T.; Najioullah, F.; Thouvenot, D.; Lina, B.; Morfin, F. Genotypic detection of acyclovir-resistant HSV-1: Characterization of 67 ACV-sensitive and 14 ACV-resistant viruses. Antivir. Res. 2008, 79, 28–36. [Google Scholar] [CrossRef]
- Sergerie, Y.; Boivin, G. Thymidine kinase mutations conferring acyclovir resistance in herpes simplex type 1 recombinant viruses. Antimicrob. Agents Chemother. 2006, 50, 3889–3892. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Milacic, V.; Chen, M.S.; Wan, S.B.; Lam, W.H.; Huo, C.; Landis-Piwowar, K.R.; Cui, Q.C.; Wali, A.; Chan, T.H.; et al. Tea polyphenols, their biological effects and potential molecular targets. Histol. Histopathol. 2008, 23, 487–496. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea Polyphenols in Promotion of Human Health. Nutrients 2018, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, R.S.; Butt, M.S.; Huma, N.; Sultan, M.T.; Arshad, M.U.; Mushtaq, Z.; Saeed, F. Quantitative and qualitative portrait of green tea catechins (gtc) through hplc. Int. J. Food Prop. 2014, 17, 1626–1636. [Google Scholar] [CrossRef]
- Ohno, A.; Yu, H.; Dickinson, D.; Chu, T.-C.; Ogbureke, K.; Derossi, S.; Yamamoto, T.; Hsu, S. Epigallocatechin-3-gallate modulates antioxidant and DNA repair-related proteins in exocrine glands of a primary Sjogren’s syndrome mouse model prior to disease onset. Autoimmunity 2012, 45, 540–546. [Google Scholar] [CrossRef]
- Cavet, M.E.; Harrington, K.L.; Vollmer, T.R.; Ward, K.; Zhang, J.Z. Anti-inflammatory and anti-oxidative effects of the green tea polyphenol epigallocatechin gallate in human corneal epithelial cells. Mol. Vis. 2011, 17, 533–542. [Google Scholar]
- Tedeschi, E.; Suzuki, H.; Menegazzi, M. Anti-inflammatory action of EGCG, the main component of green tea, through STAT-1 inhibition. Ann. N. Y. Acad. Sci. 2002, 973, 435–437. [Google Scholar] [CrossRef]
- Wang, Y.C.; Bachrach, U. The specific anti-cancer activity of green tea (−)-epigallocatechin-3-gallate (EGCG). Amino Acids 2002, 22, 131–143. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, J.H.; Lee, C.K.; Oh, C.H.; Song, H.J. The antimicrobial activity of (−)-epigallocatehin-3-gallate and green tea extracts against Pseudomonas aeruginosa and Escherichia coli isolated from skin wounds. Ann. Dermatol. 2014, 26, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Ma, C.-M.; Shahidi, F. Antioxidant and antiviral activities of lipophilic epigallocatechin gallate (EGCG) derivatives. J. Funct. Foods 2012, 4, 87–93. [Google Scholar] [CrossRef]
- Parvez, A.K.; Saha, K.; Rahman, J.; Munmun, R.A.; Rahman, A.; Dey, S.K.; Rahman, S.; Islam, S.; Shariare, M.H. Antibacterial activities of green tea crude extracts and synergistic effects of epigallocatechin gallate (EGCG) with gentamicin against MDR pathogens. Heliyon 2019, 5, e02126. [Google Scholar] [CrossRef] [Green Version]
- Calland, N.; Sahuc, M.A.; Belouzard, S.; Pène, V.; Bonnafous, P.; Mesalam, A.A.; Deloison, G.; Descamps, V.; Sahpaz, S.; Wychowski, C.; et al. Polyphenols inhibit Hepatis C virus entry by a new mechanism of action. J. Virol. 2015, 89, 10053–10063. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, B.M.; Batista, M.N.; Braga, A.C.S.; Nogueira, M.L.; Rahal, P. The Green Tea Molecule EGCG Inhibits Zika Virus Entry. Virology 2016, 496, 215–218. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P. The Green Tea Polyphenol Epigallocatechin-3-Gallate, Inhibits Hepatitis C Virus Entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Jang, M.; Park, Y.I.; Cha, Y.E.; Park, R.; Namkoong, S.; Lee, J.I.; Park, J. Tea Polyphenols EGCG and Theaflavin Inhibit the Activity of SARS-CoV-2 3CL-Protease In Vitro. Evid.-Based Complement. Altern. Med. 2020, 2020, 5630838. [Google Scholar] [CrossRef]
- Jang, M.; Park, R.; Park, Y.; Cha, Y.; Yamamoto, A.; Lee, J.I.; Park, J. EGCG, a green tea polyphenol, inhibits human coronavirus replication in vitro. Biochem. Biophys. Res. Commun. 2021, 547, 23–28. [Google Scholar] [CrossRef]
- Henss, L.; Auste, A.; Schürmann, C.; Schmidt, C.; von Rhein, C.; Mühlebach, M.D.; Schnierle, B.S. The green tea catechin epigallocatechin gallate inhibits SARS-CoV-2 infection. J. Gen. Virol. 2021, 102, 001574. [Google Scholar] [CrossRef]
- Huang, H.C.; Tao, M.-H.; Hung, T.M.; Chen, J.C.; Lin, Z.J.; Huang, C. (−)-Epigallocatechin-3-Gallate Inhibits Entry of Hepatitis B Virus into Hepatocytes. Antivir. Res. 2014, 111, 100–111. [Google Scholar] [CrossRef]
- Isaacs, C.E.; Wen, G.Y.; Xu, W.; Jia, J.H.; Rohan, L.; Corbo, C.; di Maggio, V.; Jenkins, E.C., Jr.; Hillier, S. Epigallocatechin gallate inactivates clinical isolates of herpes simplex virus. Antimicrob. Agents Chemother. 2008, 52, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Shin, J.S.; Kim, P.; Jung, Y.S.; Jeong, H.S.; Hyun, J.K.; Lee, C.K. Inhibition of Influenza Virus Internalization by (−)-Epigallocatechin-3-Gallate. Antivir. Res. 2013, 100, 460–472. [Google Scholar] [CrossRef]
- Weber, J.M.; Ruzindana-Umunyana, A.; Imbeault, L.; Sircar, S. Inhibition of Adenovirus Infection and Adenein by Green Tea Catechins. Antivir. Res. 2003, 58, 167–173. [Google Scholar] [CrossRef]
- Xu, J.; Gu, W.Z.; Li, C.Y.; Xing, G.Z.; Song, Y.; Zheng, W.M. Epigallocatechin Gallate Inhibits Hepatitis B Virus via Farnesoid X Receptor Alpha. J. Nat. Med. 2016, 70, 584–591. [Google Scholar] [CrossRef]
- Wu, C.Y.; Yu, Z.Y.; Chen, Y.C.; Hung, S.L. Effects of epigallocatechin-3-gallate and acyclovir on herpes simplex virus type 1 infection in oral epithelial cells. J. Formos. Med. Assoc. 2021, 120, 2136–2143. [Google Scholar] [CrossRef]
- Chen, P.; Dickinson, D.; Hsu, S. Lipid-soluble green tea polyphenols: Stabilized for effective formulation. In Handbook of Green Tea and Health Research; McKinley, H., Jamieson, M., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2009; pp. 45–61. [Google Scholar]
- Xu, Y.-Q.; Yu, P.; Zhou, W. Combined effect of pH and temperature on the stability and antioxidant capacity of epigallocatechin gallate (EGCG) in aqueous system. J. Food Eng. 2019, 250, 46–54. [Google Scholar] [CrossRef]
- Zhu, Q.Y.; Zhang, A.; Tsang, D.; Huang, Y.; Chen, Z.Y. Stability of green tea catechins. J. Agric. Food Chem. 1997, 45, 4624–4628. [Google Scholar] [CrossRef]
- Sang, S.; Lambert, J.D.; Ho, C.-T.; Yang, C.S. The Chemistry and Biotransformation of Tea Constituents. Pharmacol. Res. 2011, 64, 87–99. [Google Scholar] [CrossRef]
- Ping, C.; Yao, T.; Dong, S.; Xiao-Ming, Z. A Novel Long-Chain Acyl-Derivative of Epigallocatechin-3-O-Gallate Prepared and Purified from Green Tea Polyphenols. J. Zhejiang Univ. Sci. 2003, 4, 714–718. [Google Scholar] [CrossRef]
- De Oliveira, A.; Adams, S.D.; Lee, L.H.; Murray, S.R.; Hsu, S.; Hammond, J.R.; Dickinson, D.; Chen, P.; Chu, T.-C. Inhibition of herpes simplex virus type 1 with the modified green tea polyphenol palmitoyl-epigallocatechin gallate. Food Chem. Toxicol. 2013, 52, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.N.; Adams, S.D.; Lee, L.H. Inhibition of Herpes Simplex Virus-1 by the Modified Green Tea Polyphenol EGCG-Stearate. Adv. Biosci. Biotechnol. 2018, 9, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Jiang, J.; Zheng, R.; Pearl, H.; Dickinson, D.; Fu, B.; Hsu, S. A Proprietary Topical Preparation Containing EGCG-Stearate and Glycerin with Inhibitory Effects on Herpes Simplex Virus: Case Study. Inflamm. Allergy-Drug Targets 2012, 11, 364–368. [Google Scholar] [CrossRef]
- Pieknik, J.R.; Bertke, A.S.; Tang, S.; Krause, P.R. A VP26-mNeonGreen Capsid Fusion HSV-2 Mutant Reactivates from Viral Latency in the Guinea Pig Genital Model with Normal Kinetics. Viruses 2018, 10, 246. [Google Scholar] [CrossRef] [Green Version]
- Cantatore, A.; Randall, S.D.; Traum, D.; Adams, S.D. Effect of black tea extract on herpes simplex virus-1 infection of cultured cells. BMC Complement. Altern. Med. 2013, 13, 139. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.D.; Tzeng, W.P.; Chen, M.H.; Frey, T.K. Analysis of intermolecular RNA-RNA recombination by rubella virus. Virology 2003, 309, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Harden, E.A.; Falshaw, R.; Carnachan, S.M.; Kern, E.R.; Prichard, M.N. Virucidal activity of polysaccharide extracts from four algal species against herpes simplex virus. Antivir. Res. 2009, 83, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Shen, W.; Zhang, X.; Li, M.; Wang, Y.; Zou, Y.; Li, Y.; Wang, H. Anti-HSV Activity of Kuwanon X from Mulberry Leaves with Genes Expression Inhibitory and HSV-1 Induced NF-κB Deactivated Properties. Biol. Pharm. Bull. 2016, 39, 1667–1674. [Google Scholar] [CrossRef] [Green Version]
- Kwofie, S.K.; Broni, E.; Asiedu, S.O.; Kwarko, G.B.; Dankwa, B.; Enninful, K.S.; Tiburu, E.K.; Wilson, M.D. Cheminformatics-Based Identification of Potential Novel Anti-SARS-CoV-2 Natural Compounds of African Origin. Molecules 2021, 26, 406. [Google Scholar] [CrossRef]
- Chang, M.W.; Lindstrom, W.; Olson, A.J.; Belew, R.K. Analysis of HIV Wild-Type and Mutant Structures via in Silico Docking against Diverse Ligand Libraries. J. Chem. Inf. Model. 2007, 47, 1258–1262. [Google Scholar] [CrossRef] [Green Version]
- Ringer, A.L.; Senenko, A.; Sherrill, C.D. Models of S/π interactions in protein structures: Comparison of the H2S-benzene complex with PDB data. Protein Sci. 2007, 16, 2216–2223. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Jia, J.; Wang, L.; Li, F.; Wang, Y.; Jiang, Y.; Song, X.; Qin, S.; Zheng, K.; Ye, J.; et al. Anti-HSV-1 activity of Aspergilli peptide D, a cyclic pentapeptide isolated from fungus Aspergillus sp. SCSIO 41501. Virol. J. 2020, 17, 41, Erratum in Virol. J. 2020, 17, 45. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.N.; Kesharwani, A.; Agarwal, A.; Polachira, S.K.; Nair, R.; Gupta, S.K. Herbal Gel Formulation Developed for Anti-Human Immunodeficiency Virus (HIV)-1 Activity Also Inhibits In Vitro HSV-2 Infection. Viruses 2018, 10, 580. [Google Scholar] [CrossRef] [Green Version]
- Spengler, G.; Kincses, A.; Mosolygó, T.; Marć, M.A.; Nové, M.; Gajdács, M.; Sanmartín, C.; McNeil, H.E.; Blair, J.M.A.; Domínguez-Álvarez, E. Antiviral, Antimicrobial and Antibiofilm Activity of Selenoesters and Selenoanhydrides. Molecules 2019, 24, 4264. [Google Scholar] [CrossRef] [Green Version]
- Mori, S.; Miyake, S.; Kobe, T.; Nakaya, T.; Fuller, S.D.; Kato, N.; Kaihatsu, K. Enhanced anti-influenza A virus activity of (−)-epigallocatechin-3-O-gallate fatty acid monoester derivatives: Effect of alkyl chain length. Bioorg. Med. Chem. Lett. 2008, 18, 4249–4252. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, S.; Li, C.; Yang, L.; Zu, Y. In vitro evaluation of the antiviral activity of the synthetic epigallocatechin gallate analog-epigallocatechin gallate (EGCG) palmitate against porcine reproductive and respiratory syndrome virus. Viruses 2014, 6, 938–950. [Google Scholar] [CrossRef]
- Koelle, D.M. Vaccines for herpes simplex virus infections. Curr. Opin. Investig. Drugs 2006, 7, 136–141. [Google Scholar]
- Paterson, I.; Anderson, E.A. The renaissance of natural products as drug candidates. Science 2005, 310, 451–453. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plaque Assay | Untreated HSV-2 (PFU/mL) | 75 μM EGCG (PFU/mL) | % of Inhibition | Mean & STDEV | 75 μM EGCG-S (PFU/mL) | % of Inhibition | Mean & STDEV |
|---|---|---|---|---|---|---|---|
| # 1 | 1.90 × 106 | 430 | 99.977 | 99.981 ± 0.0104 | 250 | 99.987 | 99.979 ± 0.008 |
| # 2 | 1.10 × 106 | 350 | 99.968 | 350 | 99.968 | ||
| # 3 | 1.40 × 106 | 120 | 99.991 | 280 | 99.980 | ||
| # 4 | 1.60 × 106 | 200 | 99.987 | 320 | 99.980 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stamos, J.D.; Lee, L.H.; Taylor, C.; Elias, T.; Adams, S.D. In Vitro and In Silico Analysis of the Inhibitory Activity of EGCG-Stearate against Herpes Simplex Virus-2. Microorganisms 2022, 10, 1462. https://doi.org/10.3390/microorganisms10071462
Stamos JD, Lee LH, Taylor C, Elias T, Adams SD. In Vitro and In Silico Analysis of the Inhibitory Activity of EGCG-Stearate against Herpes Simplex Virus-2. Microorganisms. 2022; 10(7):1462. https://doi.org/10.3390/microorganisms10071462
Chicago/Turabian StyleStamos, James D., Lee H. Lee, Calvin Taylor, Tony Elias, and Sandra D. Adams. 2022. "In Vitro and In Silico Analysis of the Inhibitory Activity of EGCG-Stearate against Herpes Simplex Virus-2" Microorganisms 10, no. 7: 1462. https://doi.org/10.3390/microorganisms10071462