
Mycobacterium abscessus: It’s Complex
by
 , , ,
, , ,
Hazem F. M. Abdelaal
1 ,
,
Edward D. Chan
2,3,
Lisa Young
4,
Susan L. Baldwin
1 and
Rhea N. Coler
1,5,6,* 1
Center for Global Infectious Disease Research, Seattle Children’s Research Institute, Seattle, WA 98145, USA
2
Department of Academic Affairs and Medicine, National Jewish Health, Denver, CO 80206, USA
3
Pulmonary Section, Rocky Mountain Regional Veterans Affairs Medical Center, Aurora, CO 80045, USA
4
Department of Medicine, Johns Hopkins School of Medicine, Baltimore, MD 21205, USA
5
Department of Pediatrics, University of Washington School of Medicine, Seattle, WA 98195, USA
6
Department of Global Health, University of Washington, Seattle, WA 98195, USA
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(7), 1454; https://doi.org/10.3390/microorganisms10071454
Submission received: 1 June 2022
/
Revised: 12 July 2022
/
Accepted: 16 July 2022
/
Published: 19 July 2022
(This article belongs to the Special Issue Novel Models Targetting Vaccines and Drugs for M. tuberculosis, M. avium and M. absessus Infection)
Abstract
:Mycobacterium abscessus (M. abscessus) is an opportunistic pathogen usually colonizing abnormal lung airways and is often seen in patients with cystic fibrosis. Currently, there is no vaccine available for M. abscessus in clinical development. The treatment of M. abscessus-related pulmonary diseases is peculiar due to intrinsic resistance to several commonly used antibiotics. The development of either prophylactic or therapeutic interventions for M. abscessus pulmonary infections is hindered by the absence of an adequate experimental animal model. In this review, we outline the critical elements related to M. abscessus virulence mechanisms, host–pathogen interactions, and treatment challenges associated with M. abscessus pulmonary infections. The challenges of effectively combating this pathogen include developing appropriate preclinical animal models of infection, developing proper diagnostics, and designing novel strategies for treating drug-resistant M. abscessus.
1. Introduction
Non-tuberculous mycobacteria (NTM) are environmental bacteria that are found ubiquitously in the soil and in natural and potable water, and biofilms. While human exposure to NTM is widespread, diseases caused by them are relatively uncommon because of the low pathogenicity of most NTM. However, due to NTM prevalence in natural and human-made environmental niches in combination with host risk factors, NTM infections are becoming an emerging global health concern in several countries. Clinical manifestations range from cutaneous infections to chronic lung disease to disseminated disease. Of the approximately 200 species of NTM identified, most infections are caused by Mycobacterium avium (M. avium) complex (MAC, comprised of several species, with the most common being M. avium, M. intracellulare, and M. chimaera, among several others), M. abscessus complex (with subspecies abscessus, subsp. massiliense, and subsp. bolletii), and M. kansasii [1]. However, depending on the region of the world, some other “less common” NTM species may be relatively prevalent such as M. malmoense in Northern Europe and M. xenopi in Canada and certain regions of Europe (central, southern, and the United Kingdom). Lung disease due to NTM is estimated to be increasing by approximately 8% per year in the United States in 2.3 million Medicare beneficiaries that were over 65 years of age, and in whom 58% were female subjects [2]. Other regions of the world have also noted an increase prevalence of NTM lung disease (NTM-LD). For example, NTM cases also increased from 0.9 to 2.9 per 100,000, respectively, from 1995 to 2006 in England, Wales, and Northern Ireland [3] and are also increasing in Canada [4]. The two main categories of NTM include the rapidly growing mycobacteria and the slowly growing mycobacteria. The rapid growers include the Mycobacterium chelonae-abscessus complex, including M. abscessus subsp. abscessus, M. abscessus subsp. bolletti, M. abscessus subsp. massiliense, M. chelonae, and M. fortuitum [5]. The opportunistic slow growers include the M. avium complex (MAC), with several different species including M. avium, M. intracellulare subsp. intracellulare, and subsp. chimaera, among others.
As an opportunistic pathogen, M. abscessus often colonizes the lung airways in patients with cystic fibrosis (CF), chronic obstructive pulmonary disease (COPD), or bronchiectasis [6], whereas humans (and mice) with normal lung airways are typically resistant to lung infection. Much more remains to be discovered about the host–pathogen interactions associated with a progressive pulmonary parenchymal infection.
The intrinsic drug resistance in M. abscessus has limited its therapeutic options, with only a few active antibiotics available that are effective. Severe M. abscessus lung infections are often treated with a cocktail of oral and parenteral antibiotics as well as surgical lung resection in those with localized but recalcitrant lung disease.
An evaluation of the in vivo susceptibility and efficacy of these drugs against acute M. abscessus infection has been performed using both immunocompromised mice and zebrafish embryos. Imipenem-cilastatin and clarithromycin conferred protection against M. abscessus as demonstrated by restricted mycobacterial growth, decreased pathologic manifestations such as brain abscesses, and increased embryo survival [7]. However, antibiotic use has been limited due to non-standardized in vivo antibiotic susceptibility testing, limited cellular/animal testing models, non-uniform susceptibility among different clinical isolates of M. abscessus, and incongruent clinical response, often resulting in long-term failure [8]. Worldwide, it is plausible that increased outbreaks and nosocomial transmission of M. abscessus complex are rising, reflecting an emerging antibiotic resistance crisis that threatens public health [9,10]. Further research is needed to identify optimal in vivo conditions to test the drug efficacy and optimize therapeutic regimens for M. abscessus infections.
2. Critical Elements of M. abscessus Virulence and Pathogenicity
Classifying M. abscessus as an opportunistic pathogen occurring in immunocompromised individuals has recently been challenged based on the observation that the M. abscessus clade, with numerous virulence factors, may actually fit the criteria of a true pathogen [11]. The ability of M. abscessus to thrive in various environmental niches [12] helps explain its ability to also colonize human-made sources and materials, such as potable water and plumbing systems, as well as medical and surgical instruments/devices resulting in nosocomial NTM infections [13,14,15].
The increasing incidence of M. abscessus infections in cystic fibrosis (CF) and non-CF populations reported globally [16] provides further evidence that M. abscessus possesses a diverse repertoire of virulence factors that are representative of a successful human pathogen [11]. M. abscessus pathogenic evolution may be forced by the competing requirement to maintain transmission fitness [17]. Perhaps the most prominent finding in these evolution-based studies are the results that identified strong evolutionary pressure on M. abscessus genes promoting survival within the macrophage, which seem to come with a fitness cost in favor of enhanced virulence of M. abscessus isolates.
Due to the ability of M. abscessus to survive and replicate within free-living amoeba, an essential role of the ESX-4 M. abscessus type VII secretion system (T7SS) was discovered, further substantiating that a genetic factor may have naturally selected for intracellular survival of M. abscessus [18]. ESX-4 in M. abscessus is known to block phagosomal acidification and disrupt phagosomes, similar to the role of ESX-1 in Mycobacterium tuberculosis (M. tuberculosis) [18]. In humans and other animal species, the transcriptomic changes of M. abscessus during intracellular growth in macrophages have demonstrated upregulation of genes such as heat shock and oxidative stress (e.g., GroEL-ES and hsp) to cope with intracellular stresses [19]. Those factors are associated with a switch to a slower growth phenotype, and the utilization of fatty acids as an energy source, which displays the range of mechanisms M. abscessus uses to persist in the host. An additional important virulence factor includes the complex lipids of M. abscessus, including glycopeptidolipids (GPLs), involved in adherence of M. abscessus. The loss of GPL is associated with the transition from a smooth (S) to a rough (R) morphotype in NTM [20,21], resulting in increased virulence in part by forming large extracellular serpentine cording, leading to the prevention of phagocytosis, increased inflammation, and the characteristic formation of abscesses [22,23]. Interactions between M. abscessus and host myeloid cells revealed the utilization of this extracellular cording to subvert the host innate immune response [7]. Cord-deficient M. abscessus mutants exhibit altered mycolic acid composition as well as extremely diminished phenotypes (i.e., lack of granuloma formation and replication in macrophages) in wild-type and immunocompromised zebrafish embryos [24]. GPLs also inhibits apoptosis in M. abscessus-infected macrophages by interacting with the mitochondrial cyclophilin D [25]. During persistent infection, the transition to this unique R colony morphotype is critical for invading and destroying a variety of cell types, including macrophages in monolayer culture, as seen in the zebrafish embryo infection model [5]. However, the cellular triggers for this transition between morphotypes remain unclear.
Another role for GPL is triggering a strong humoral immune response [23]. Anti-GPL response may explain the conversion to the more virulent R morphotype in order to escape such response [23,26].
The involvement of complex lipids of M. abscessus in virulence has also shed light on the importance and role of lipid-specific transporters. For example, the Mycobacterial membrane protein Large (MmpL) permeases family, including MmpL4a and MmpL4b, are involved in GPL transport [27]. Another MmpL discovered to be associated with M. abscessus virulence is MmpL8 [28]. The loss of function (LOS) of MmpL8 results in reduced glycosyl diacylated nonadecyl diol (GDND) production and decreased intracellular survival and virulence.
The growing incidence of M. abscessus can also be linked to the global rise in individuals who are immunocompromised or have pre-existing lung conditions [29]. M. abscessus can also cause extrapulmonary infections in the skin, soft tissue, bones, joints, lymph nodes, and internal organs [30]. Like its fellow pathogenic mycobacteria, M. abscessus can form distinctive granulomas by evading phagosomal defense mechanisms (macrophages and neutrophils), inducing the production of inflammatory cytokines, such as tumor necrosis factor (TNF), and recruiting B and T lymphocytes to the site of infection.
3. NTM Diseases
3.1. NTM Lung Disease in Subjects with Known Risk Factors
Non-tuberculous mycobacterial lung disease (NTM-LD) occurs primarily in three broad groups of patients (Figure 1): (i) Acquired: Those with acquired anatomic lung or immune abnormalities with no identifiable genetic basis; e.g., localized bronchiectasis from prior unrelated infections, smoking-related emphysema, pneumoconiosis such as silicosis [31,32,33] as well as the use of either inhaled glucocorticoids or tumor necrosis factor (TNF) antagonists [34], (ii) Genetic/Hereditary: Those with genetic disorders that predispose to bronchiectasis and/or lung infections; e.g., cystic fibrosis (CF), primary ciliary dyskinesia, alpha-1-antitrypsin (AAT) deficiency, Williams–Campbell syndrome, Mounier-Kuhn syndrome, Sjogren’s syndrome, pulmonary alveolar proteinosis (PAP), and common variable immunodeficiency [35,36], and (iii) No Obvious Predisposing Factors: Those with no known prior lung or immunological abnormalities [37,38,39]. The pathophysiologic mechanisms by which primary immunologic and/or genetic disorders predispose to isolated NTM-LD are listed in Table 1. One of the acquired risk factors includes chronic aspiration. Chronic aspiration due to gastroesophageal reflux disease (GERD) has been reported to be more common in patients with lung disease associated with MAC and other NTM [40,41,42]. MAC disease can also occur in patients that aspirate due to other reasons. Swallowing disorders can also lead to the risk of aspiration (Figure 1) [40].
Another acquired risk factor listed in Figure 1 is calcified chest adenopathy. In a retrospective study including 79 patients with pulmonary MAC disease, calcified chest adenopathy was present in one-third of the patients, and furthermore, those with calcified lymphadenopathy in the chest lived in a Histoplasma capsulatum endemic area, whereas those living outside this endemic region had no adenopathy [43]. This led to a hypothesis that fungal infection could predispose the lungs of some patients to MAC infection by either airway distortion or parenchymal damage [43].
Smoking is known to be a risk factor for TB, which is not surprising based on the damaging effects that smoking has on the lungs and the immune system (for a recent review, see Quan et al. [44]). Smoking-related emphysema is an acquired risk factor for NTM lung disease as well (Figure 1). PAP as a risk factor is supported by the finding that mice with disruption of the granulocyte-monocyte colony stimulating factor-1 are more vulnerable to M. abscessus [45]. We discuss below in greater detail two conditions well established to be associated with NTM-LD: CF and AAT deficiency.
3.1.1. CFTR Anomalies and Susceptibility to NTM
CF is an autosomal recessive disorder caused by mutation of the CYSTIC FIBROSIS TRANSMEMBRANE CONDUCTANCE REGULATOR (CFTR) gene. The prevalence of a CFTR gene mutation in the Caucasian population is estimated to be 1 in 20 individuals, resulting in occurrences of CF in 1 in 2000–2500 live births [52]. CF patients are particularly susceptible to recurrent and chronic bacterial and mycotic infections, including those due to Staphylococcus aureus, Pseudomonas aeruginosa, Burkholderia cepacia complex, and NTM, among others [53,54,55]. The mechanisms for susceptibility in CF patients are complicated by thickened mucus and inability to clear bacteria, secondary ciliary dysfunction, and reduced human beta-defensin-2 function and level [56]. Macrophage dysfunction due to the CFTR mutation has also been demonstrated and resulting in impaired phagocytosis and reduced efferocytosis (with reduced clearance of apoptotic neutrophils), and excessive production of inflammatory mediators directed towards microbial stimuli [34,57]. Although the topic of whether the M1 (“pro-inflammatory”) or the M2 (“anti-inflammatory”) macrophage phenotype predominates in the CF lung is controversial, there is evidence that CF macrophages are defective in switching between these two phenotypes [58].
3.1.2. AAT Anomalies and Susceptibility to NTM
AAT deficiency predisposes to NTM-LD by two main mechanisms. One is that AAT deficiency is a risk factor for both chronic obstructive pulmonary disease and bronchiectasis, well-known underlying lung conditions for subsequent NTM-LD [34,56,57,61].
Another mechanism is that AAT itself has host-defense properties against NTM through induction of autophagy in macrophages [62]. This finding is supported epidemiologically by studies showing that the presence of heterozygous AAT anomalies—which are not known to cause bronchiectasis on their own—were more common in patients with NTM-LD compared to the general population in the U.S. [63,64]. Thus, susceptibility of AAT-deficient individuals to NTM-LD may occur because of impaired innate immunity as well as alterations in lung architecture (bronchiectasis and COPD).
3.2. NTM-LD in Patients without a Known Underlying Cause
In individuals with NTM-LD without any known predisposing conditions, the bronchiectasis, bronchiolitis, and the sequelae of atelectasis and cavities are presumed to be caused by the NTM infection and chronic airway inflammation. It has been observed that a significant number of NTM-LD subjects without an identifiable predisposing factor possess a life-long slender body habitus (Figure 1) [38,46,65,66,67] and reduced visceral fat with the caveat that weight loss may also occur post-NTM infection [68]. Low body weight itself has been hypothesized as a risk factor for NTM-LD and tuberculosis [35,69,70,71]. Furthermore, low BMI (<18.5 kg/m2) is associated with a greater number of diseased lung segments and NTM-LD-specific mortality [69,72]. Interestingly, NTM-LD has been reported in younger women (ages 20–53 years old) diagnosed with anorexia nervosa [73,74,75,76]. In general, since NTM-LD is much less common in younger individuals than in the elderly, this lends credence to the possibility that a thin body habitus is a risk factor for NTM-LD. Elderly Caucasian, post-menopausal females with slender body morphotypes are disproportionately predisposed to NTM-LD compared to males [47,54]. Hormonal changes and immune-senescence (changes in the immune system as a result of aging) likely contribute to risk factors for NTM-LD, in which an accumulation of damaged DNA and other factors are thought to result in a low-grade inflammatory phenotype termed ‘inflammaging’, which can alter immune function [47,49].
A possible mechanism by which slender individuals with low body fat content may be predisposed to NTM infections is the relative deficiency of leptin, a satiety hormone [77]. Leptin has a number of immunomodulatory functions that can potentially enhance host immunity against NTM, including the differentiation of uncommitted T0 cells toward the TH1 interferon-gamma (IFNγ)-producing phenotype [77]. Indeed, mice deficient in leptin are more susceptible to experimental M. abscessus lung infection [65,77,78]. Reduced levels of leptin in the sera of pulmonary NTM patients have also been observed [79].
Some NTM-LD patients have a greater than expected preponderance of abnormalities within the thoracic cage region, such as pectus excavatum and scoliosis [46,65,66,67,75,79,80]. We and others have postulated that thoracic cage abnormalities may be a marker for an underlying and yet-to-be identified genetic predisposition, perhaps related to a minor variant of Marfan syndrome (due to mutations of fibrillin-1) or ciliary dysfunction (due to mutations of different genes that encode for ciliary proteins) [65,66], [75,80,81,82]. Pectus excavatum and scoliosis have also been described in other connective tissue disorders, such as Loeys–Dietz syndrome (LDS, due to gain-of-function mutation of transforming growth factor-beta receptors 1/2—TGFβR1/2) and Shprintzen–Goldberg Syndrome (SGS, due to mutation of the Sloan Kettering Institute (SKI) protein, a downstream inhibitor of TGFβ signaling) [83]. While these disorders are due to monogenic mutations of different genes, each result in increased signaling of TGFβ, a cytokine known to predispose to NTM infection [84,85].
In light of this, the whole blood of NTM patients was found to produce more TGFβ, and lower levels of IFNγ upon ex vivo stimulation with various Toll-like receptor agonists or with M. intracellulare as compared to similarly stimulated whole blood from uninfected controls [65]. Daniels et al. analyzed for the presence of dural ectasia—an enlarged dural sac seen in MFS, LDS, and SGS—in patients with idiopathic bronchiectasis, CF subjects, MFS, and controls and found that the L1–L5 dural sac diameter was significantly greater in patients with idiopathic bronchiectasis as compared to controls and to CF subjects, suggesting the possibility of an underlying connective tissue disorder in those with idiopathic bronchiectasis [82]. They also found a strong correlation between dural sac size and NTM-LD, as well as dural sac size and long fingers [82]. NTM-LD was also reported in a patient with congenital contractural arachnodactyly, a genetic disorder due to FIBRILLIN-2 gene mutation and which shares many clinical features with MFS [86].
Fowler et al. described reduced ciliary beat frequency in the nasal epithelium and reduced nasal nitric oxide (NO) in NTM-LD patients compared to controls; the ciliary beat frequency was increased by NO donors or compounds that increased the concentration of cyclic guanosine monophosphate, a downstream mediator of NO [87]. Subsequent whole exome sequencing of NTM-LD subjects showed, compared to control data from the 1000G Project, increased variants in immune, CFTR, ciliary, and/or connective tissue genes, implicating a multigenic disorder for some patients with NTM-LD [81].
Because the variants of immune genes were significantly more common in NTM-LD patients than in unaffected family members, immune gene variants may be the discriminating genetic factor for the development of NTM-LD [81]. Furthermore, the number of CFTR variants per person was actually greater in both control groups (family members not infected with NTM and in the 1000G Project cohort) than in NTM-LD subjects [81]—in contrast to the other three non-CFTR gene categories in which the number of variants was least in the control 1000G Project cohort—it favors the possibility that the risk for NTM-LD in CF patients is perhaps due to the presence of bronchiectasis and not to the CFTR mutation per se. Becker and colleagues performed whole exome sequencing on 11 NTM-LD subjects with slender body habitus, pectus excavatum, and scoliosis and found one with mutation of the Fibrillin-1 gene and four (two being sisters) with heterozygous mutations of the Macrophage-Stimulating 1 Receptor (MST1R) gene and in none of 29 NTM-LD patients without pectus excavatum or scoliosis [88]. While these investigators showed that MST1R may function to increase IFNγ production, MST1R was previously reported to be a tyrosine kinase receptor found on the apical epithelial surfaces of fallopian tubes and airways and upon binding to its ligand (macrophage stimulating protein), enhanced ciliary beat frequency [89,90].
3.3. Disseminated NTM Disease
Patients with extrapulmonary visceral organ or disseminated NTM disease are almost always frankly immunocompromised, such as those receiving tumor necrosis factor (TNF) antagonist therapy, organ transplantation, and having untreated AIDS (Figure 1 and Figure 2) [91,92,93]. Figure 2 includes several host-defense pathways used against M. abscessus in addition to mechanisms that can interfere with host-defense and lead to NTM disease. The use of immunosuppressive drugs, such as inhaled corticosteroids, can increase the risk of NTM disease [50,51] (Figure 1 and Figure 2). Mutations in GATA2 (guanine-adenine-thymine-adenine-2), a transcription factor, can lead to monocytopenia and mycobacterial (MAC) infection (called monoMAC syndrome), causing disseminated NTM with decreases in monocytes, DC’s, B cells and NK cells [94,95] (Figure 2). Individuals with other certain rare inherited disorders—particularly those with defects of the interleukin 12 (IL-12)/interferon-gamma (IFNγ) cytokine axis, and that fall under the rubric of Mendelian Susceptibility to Mycobacterial Diseases (MSMD)—are predisposed to an extrapulmonary visceral organ or disseminated NTM infections (Figure 2) [96,97,98,99,100,101,102,103,104,105,106,107,108]. Several of the MSMD-causing mutations have been identified in seven different autosomal genes and are described in detail in the review by Bustamante et al. [100]. Some of the proteins encoded by these genes are included in Figure 2 and represent the importance of each in the host’s defense against mycobacterial infections, including the IL-12 receptors (encoded by IL12 B (p40 subunit) and IL12B1 (b1 chain of the IL-12 receptor), IFNγ receptors (encoded by IFNGR1 and IFNGR2), transcriptional factor induced by IFNγ (IRF8), and signal transducer and activator of transcription 1 (STAT1); IFNγ-inducible factor (ISG15) (one of the seven autosomal genes with MSMD-causing mutations, not shown in Figure 2). MSMD-causing mutations in the X-linked gene include the nuclear factor-kappa B (NF-kB) essential modulator (NEMO), shown in Figure 2. The major component of the NADPH oxidase complex (CYBB) is another X-linked gene with MSMD-causing mutations [100]. Susceptibility to disseminated NTM in such individuals is corroborated experimentally by the increased vulnerability to M. abscessus in the IFNγ-knockout mice (Ordway et al., 2008). Individuals with acquired autoantibodies to IFNγ have more recently been described to be also more vulnerable to extrapulmonary visceral organ and disseminated NTM disease [109]. TNF-α inhibitors (including anti-TNF-α monoclonal antibodies and soluble TNF receptor fusion proteins (TNFR) used to suppress the immune response in patients with chronic inflammatory diseases, such as rheumatoid arthritis (RA), have been reported to increase the rate of mycobacterial disease, including NTM, compared to untreated patients and the general population [93,110,111]. The mechanism of anti-TNF monoclonal antibodies and soluble TNFR in host-defense impairment have been described elsewhere [112].
Many mycobacteria, including M. abscessus and fungi, are recognized by Toll-like receptor 2 (TLR2) and the beta-glucan receptor Dectin-1 [113]. Dectin-1 signaling leads to caspase-1 and IL-1β activation through the nucleotide-binding domain (NOD)-like receptor protein 3 (NLRP3)/ASC inflammasome, leading to host defense responses against M. abscessus [114]. Both Dectin-1 and TLR2 are necessary for M. abscessus-induced expression of innate antimicrobial responses, including interleukin-1 beta (IL-1β) and LL-37 [115]. TLR2-deficient mice are extremely susceptible to rough variants of M. abscessus due to failure of TH1-induced immunity [116]. Interleukin-8 (IL-8, also called CXCL8) is a chemokine-induced by infection and produced by macrophages and other cells and is a neutrophil chemotactic factor. Early neutrophil responses may help control infection with NTM, as shown with M. fortuitum [117]. These “experiments of nature” provide great insights into which elements of the immune system provide host-induced protection against mycobacteria.
4. Treatment against M. abscessus Related Infections
4.1. Antibiotics Used for Treating M. abscessus
Despite its low virulence, treatment of M. abscessus is particularly difficult because of its intrinsic resistance to several commonly used antibiotics (Figure 3). Recommendations from The American Thoracic Society/Infectious Diseases Society of America include macrolides (typically Azithromycin favored over Clarithromycin) [118], Aminoglycosides (Amikacin), Carbapenems (Imipenem), and Cephamycins (Cefoxitin) [118,119,120].
Macrolides target bacterial 23S rRNA, inhibiting bacterial protein synthesis. M. abscessus possess two major forms of macrolide resistance, and both involve the bacterial 23S rRNA by different mechanisms. The first is “genetic” macrolide resistance and is due to single point mutation in position 2058 or 2059 of the bacterial 23S rRNA gene (also known as the rrl gene) [121]. The second form is known as “inducible” macrolide resistance, wherein a functional ERM41 gene encodes a methylase that occupies a site on 23S rRNA preventing macrolides from binding [122,123] (Figure 4). Among the M. abscessus organisms, the majority of subsp. abscessus and subsp. bolletii strains possess a functional ERM41 gene, which confers an inducible resistance to macrolides.
Conversely, a minority (15–20%) of subsp. abscessus isolates possess a T28C mutation of the ERM41 gene, resulting in a non-functional methylase with preserved macrolide susceptibility. Similarly, all subsp. massiliense strains contain a partially deleted, non-functional ERM41 gene and thus also have preserved macrolide susceptibility. Thus, in the absence of rrl gene mutation, NTM-LD patients infected with M. abscessus with a non-functional ERM41 gene and hence preserved macrolide susceptibility (a minority of subsp. abscessus strains and all strains of subsp. massiliense) have better clinical outcomes than those infected with M. abscessus isolates with a functional ERM41 gene and consequently inducible macrolide resistance (most subsp. abscessus and essentially all of subsp. bolletii).
The enzymatic modification of antibiotics by N-acetyltransferases confers aminoglycoside resistance. These specific enzymes add chemical groups to the 2′ amino groups of aminoglycosides, thus blocking the antibiotic from binding to its target protein [124]. In recent years, poor outcomes in patients infected with susceptible strains (approximately 90% of M. abscessus clinical isolates) stem from the development of resistance to amikacin, a key drug. In particular, patients with amikacin-resistant M. abscessus-LD (frequently involving rrs mutations) have shown unsatisfactory treatment outcomes, which is problematic given amikacin’s important role in long-term treatment [125]. M. abscessus can acquire fluoroquinolone resistance through cumulative mutations in a highly conserved region in the quinolone resistance-determining region (QRDR) of a DNA gyrase gene [126].
Unique structural and pathological traits contribute to drug resistance in clinical isolates of M. abscessus, including a capacity to form biofilms that prevents drug penetration [127]. A major pathogenic trait is an indolent progression; the rapid, silent growth can go undetected, eventually causing a severe deterioration in the human host [128]. Patients with chronic structural lung diseases such as CF and emphysema are at exceptionally high risk of pulmonary disease [129]. Early signs include sudden, progressive lung dysfunction, often accompanied by caseous lesions and alveolar granulomas [130,131]. While technically serving as a host defense mechanism, the granuloma also enables latent NTM infection and drug evasion by blocking drug penetration. Thus, early diagnosis and detection are vital but limited by poor understanding of its pathogenesis and the inability to adequately differentiate its symptoms from TB. Given the lack of standardized diagnostic criteria, misdiagnosis and treatment with anti-Mycobacterium tuberculosis medications are frequent but inappropriate considering the distinct treatment needs [126].
4.2. Strategies for Treating Drug-Resistant M. abscessus
Worldwide, outbreaks and nosocomial transmission of M. abscessus complex are rising, reflecting an emerging drug resistance crisis and a critical public health problem [9]. In recent years, poor treatment outcomes in patients infected with susceptible strains, which comprise approximately 90% of M. abscessus clinical isolates, have been spurred by the development of resistance to amikacin, a key drug against this pathogen. Given the current suboptimal outcome in patients with M. abscessus infection, more effective antimicrobials are needed not only for killing efficacy but also for a shorter time of treatment.
Currently, macrolides (Clarithromycin or Azithromycin) are the most used antibiotics against M. abscessus. Thus, it is not surprising whether a M. abscessus isolate is susceptible or resistant to the macrolides is a key decision point in both the initial choice of antibiotic regimen and clinical outcome (Figure 3). While the absence or presence of macrolide resistance is also a key decision point for treatment and outcome of MAC-LD, the option for oral antibiotics for M. abscessus is much more limited than for MAC.
Several antibiotic alternatives to treat multi-drug resistant (MDR) M. abscessus include natural plant-derived products with antimicrobial effects, antimicrobial nanoparticles, antimicrobial peptides, antibiotic combinations, structurally modified antibiotics, pathogen-specific monoclonal antibodies, drug-induced changes in small regulatory RNAs (sRNAs), and therapeutic bacteriophages [132,133]. Of these, the use of pathogen-specific bacteriophages, known as phage therapy (PT), has shown exciting results. Abundant in nature and prolific, phages can either actively replicate (the lytic cycle) or lie dormant (the lysogenic cycle) in their hosts. Genetic engineering can be used to enhance the killing properties and host range of phages [132]. The first successful use of PT to treat a severe M. abscessus subsp. massiliense infection occurred in 2019 in a 15-year-old lung-transplant patient [134]. No adverse effects were observed following a cocktail regimen with three phages (one natural, two engineered). However, limitations to PT include a lack of lytic phages with an extensive host range and a possibility of emerging phage resistance. Bacteriophage cocktail therapy and CRISPR-Cas genomic technology are being strongly considered to increase mycobacteriophages’ host range and therapeutic potential against MDR M. abscessus.
Recently, compassionate use with phage therapy was included in a pilot study in patients with various mycobacterial infections, untreatable with antibiotics, which showed positive clinical responses in 11 out of 20 patients [135]. Favorable or partial responses were observed in two patients with M. abscessus subsp. massiliense, six patients with M. abscessus subsp. abscessus, one patient with M. chelonae, one patient with Mav complex, and one patient with disseminated BCG [135]. The development of phage treatments could provide a crucial tool for physicians when no other options are available.
5. Novel Therapeutic Strategies
Treatment of M. abscessus infection is becoming more challenging with increased resistance to many of the current drugs and the lack of a sufficient pipeline of new drug candidates.
There are several novel drug approaches, however, that are currently being investigated. Guo et al. have recently shown in vitro efficacy of Cotezolid (MRX-I), (S)-5-([isoxazol-3-ylamino]methyl)-3-(2,3,5-trifluoro-4-[4-oxo-3,4-dihydropyridin-1(2H)-yl]phenyl)oxazolidin-2-one, which is an oxazolidinone, against M. abscessus [136]. Linezolid, also an oxazolidinone, is recommended for use against M. abscessus; however, Cotezolid may have an advantage by inducing fewer side effects; oral MRX-I administration was found to be well tolerated in humans in a Phase 1 study where adverse events were shown to be mild to moderate [137]. In the study by Guo et al., both Cotezolid (MRX-I) and linezolid are effective against M. abscessus but not M. avium or M. intracellulare [136]. Furthermore, Contezolid (MRX-I) was compatible with other M. abscessus drugs, including Azithromycin, Clarithromycin, Cefoxitin, Imipenum, Tigecycline, Bedaquiline, Amikacin, and Amoxifloxacin [136].
An analog of Linezolid, called Sutezolid, exhibits lower in vitro minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC) against M. abscessus compared to Linezolid (Dae Hun Kim, AAC, 2021, PMID:33903101) and may have fewer in vivo toxicities than linezolid as shown in studies testing the use of these drugs in healthy volunteers for intended use against Mycobacterium tuberculosis [138,139].
Ganapathy et al. have recently shown that a novel mycobacterial DNA gyrase inhibitor (MGI), an advanced M. tb drug candidate, EC/11716, has in vitro bactericidal activity against both Mav and M. abscessus and importantly has activity against M. abscessus biofilms [140]. EC/11716 was also shown to have in vivo efficacy in a preclinical M. abscessus NOD SCID mouse model [140]
Another promising drug candidate for use against M. abscessus is T405, which is a novel b-lactam of the penem subclass and was recently shown to have in vitro synergy in combination with other antibiotics, including imipenem cefditoren or avibactam [141]. Furthermore, T405 combined with probenecid exhibited bactericidal efficacy in the C3HeB/FeJ in vivo mouse model against the well-characterized ATCC29977 reference strain (Rimal B. et al., AAC, 2022, PMID:35638855). Beta-lactam antibiotics are known to interfere with bacterial cell wall peptidoglycan biosynthesis (for the mechanism of activity of b-lactams, see a recent review by Turner et al.) [142].
Diazabicyclooctanes (DBOs), including Durlobactam (DUR), are included within a class of novel b-lactamase inhibitors that inhibit peptidoglycan transpeptidases which, when combined with dual b-lactams, could potentially improve clinical efficacy and reduce the toxicity of Mab treatment regimens [143,144].
Unlike M. tuberculosis, there is currently no vaccine available for M. abscessus, and there are no vaccines in clinical development. Therapeutic vaccination as an adjunct to drug treatment against M. abscessus and other NTM could shorten drug treatment regimens and decrease the side effects associated with the current repertoire of available drugs used against NTM.
Recently, two relatively new additions to the anti-NTM drugs, developed originally to treat tuberculosis or leprosy, were used against M. abscessus-PD; Bedaquiline (BDQ) and Clofazimine (CFZ), respectively [145,146]. BDQ, an ATPase inhibitor, is the first drug approved to treat MDR-TB by the FDA in 40 years [147]. In a recent study by Sarathy et al., 3,5-dialkoxypyridine analogues of BDQ showed promising in vitro and in vivo activities against M. abscessus, similar to its BDQ parent [148]. Given that analogues of BDQ are less lipophilic, have higher clearance, and display lower cardiotoxicity, they are promising drug candidates to be co-administered with currently used drugs. On the other hand, CFZ is an approved drug for leprosy being repurposed for TB treatment [149]. CFZ is considered to be one of few candidates that are being tested for monotherapy against M. abscessus-PD [5]. In a recent trial, after one year of CFZ-containing regimes, treatment of M. abscessus-PD patients showed conversion to culture negative (CCN) [150]. Both BDQ and CFZ drugs have shown efficacy against M. abscessus alone and in combination [145,146].
Notably, Amikacin is known to induce systematic toxicity, including hearing loss, loss of balance, or both, especially when given by the intravenous route [151]. To reduce these adverse effects and to increase drug concentrations in endobronchial tissues, amikacin by aerosolization has been increasingly used [152]. In previous trials, inhaled amikacin demonstrated increased efficacy in terms of increased chance of CCN [152,153]. Amikacin liposome inhalation suspension (ALIS), in which amikacin is encapsulated in liposomes and delivered into the lungs via aerosol nebulization, has shown increased efficacy against M. avium refractory lung disease [154]. Compared with intravenous administration of non-liposomal amikacin, ALIS increased amikacin concentration by 42-fold in lung tissues, 69-fold in airways and 274-fold in macrophages [155]. ALIS is currently in a phase II trial for treating M. abscessus-LD [5]. Recently, a compassionate use study using ALIS in patients with M. abscessus pulmonary disease previously treated with various treatment regimens was described. This study included 41 patients, 61% of which had a ‘good outcome’ defined as outcomes cure, microbiologic cure, and clinical cure [156].
Another novel approach for treating M. abscessus-PD is using the apoptotic bodies to target host immune responses targeted to the pathogen, rather than directly targeting the pathogen. The principal of using apoptotic bodies is to improve phagocytosis, phagolysosomal maturation, and intracellular mycobacterial killing by sending in a second lipid messenger (bioactive lipids) known for promoting phagosomal maturation through recognition of specific lipid-binding domains [157,158]. Apoptotic bioactive-lipids (ABL) loaded with different bioactive lipids have been evaluated in case of bacterial interference of phagolysosome biogenesis and genetically impaired phagolysosome-dependent antimicrobial response, i.e., CF [159]. In both conditions, ABLs demonstrated a significant increase in intraphagosomal acidification and induction of reactive oxygen species (ROS) production and ultimately promoted intracellular mycobacterial killing in macrophages [159]. Recently, in a study by Poerio et al., ABLs loaded with phosphatidylinositol 5-phosphate (ABL/PI5P), alone or in combination with amikacin, have been evaluated for the treatment of M. abscessus-PD [160]. The combination treatment of ABL/PI5P and amikacin showed a significant reduction of pulmonary mycobacterial burden.
BCG, the only approved vaccine used for the prevention of serious forms of TB in children and adolescents, showed cross-protective immunity against M. avium and M. abscessus-related infection [161]. This fact was confirmed through the epidemiological evidence suggesting that BCG vaccination decreases the risk of developing NTM-PD [162]. It was suggested that BCG vaccination can be used as either a therapeutic or prophylactic vaccine against M. abscessus-PD [161,163]. BCG, as a live-attenuated vaccine, induces T-cell expansion important for intracellular pathogens like M. abscessus [161,164]. However, BCG is contraindicated for immune-compromised individuals such as HIV/AIDS patients [165,166]. Additionally, BCG does not reduce M. avium infection in the mice model in case of prior exposure to NTM [167,168]. The use of BCG as an intervention to prevent or treat M. abscessus-PD is hindered by the complex mechanism of NTM exposure.
Due to the complexities and difficulties in the treatment of M. abscessus infection, there is an urgent need for a therapeutic vaccine to overcome the lengthy treatment time and required toxic concentrations of antibiotics [169]. A therapeutic vaccine could also help with the acquired drug resistance to antibiotics used with M. abscessus infections. The design of a therapeutic vaccine can be acquired from the knowledge gained in the M. tb field. The most prominent example is the use of the Phase 2a clinical trial ID93 + glucopyranosyl lipid adjuvant (GLA) formulated in an oil-in-water stable nanoemulsion (SE) as a therapeutic vaccine against M. tb [170,171,172]. This vaccine design highlights the importance of selecting both mycobacterial antigens and a potent immune-stimulating adjuvant. The use of protein/adjuvant immunotherapy combined with a drug treatment strategy is commonly used for other vaccine studies for infectious diseases [170,173,174,175,176,177]. However, the lack of vaccine development for M. abscessus infection (either prophylactic or therapeutic) calls for the need to invest in these strategies to overcome the complexities involved with the treatment of M. abscessus infections.
6. Preclinical Models for M. abscessus
The biggest challenge in discovering novel host-directed therapeutic interventions for M. abscessus infections is the absence of an adequate experimental animal model. A summary of the preclinical M. abscessus models is shown in Table 2. M. abscessus are generally less virulent than Mycobacterium tuberculosis complex members, shown by the decreased capacity to induce a sustained progressive infection in an immunocompetent mouse model [178]. Therefore, there is an urgent need therefore for the development of a M. abscessus challenge model for the development of host-directed therapies and other host interventions such as therapeutic vaccines. Experimental animal models have been of great benefit for developing prophylactic and therapeutic vaccine strategies for treating M. tb, which has been the focus of our laboratory for several years [168,169,179,180], and we have now begun applying our expertise to vaccines against NTM infections [168,169]. Others have also focused on this challenging quest for therapeutic solutions against NTM [160,181,182,183,184,185,186]. Many preclinical models have been proposed to study NTM Infection for early drug discovery and vaccine research [30].
Regarding M. abscessus, many nonmammalian models are also used, such as Amoebas (Dictyostelium discoideum) [187], Drosophila melanogaster [188], Galleria mellonella larvae [189], Silkworm [190], and zebrafish [191,192,193]. Nonmammalian models are valuable models for screening anti-mycobacterial drugs and imaging host–pathogen interactions at a cellular level due to their relative transparency combined with the development of recombinant bacterial strains that express fluorescent proteins. Drawbacks of those models, however, include their inability to mimic chronic infection that can only be modeled in a mammalian host.
The mouse infection model, developed for several infectious disease pathogens, has been more extensively utilized than any other preclinical model for drug discovery and vaccine research. As previously mentioned, the use of an immunocompetent mouse model in M. abscessus infection is not considered an adequate model due to the rapid clearance of the M. abscessus [194]. The existence of several immunocompromised mouse models, including severe combined immunodeficiency (SCID) mice, granulocyte monocyte-colony stimulating factor knockout mice (GM-CSF−/−), and NOD.CB17-Prkdcscid/NCrCrl mice with compromised B cells, T cells, and natural killer cells resulted in M. abscessus progressive infection, similar to that seen with human M. abscessus-LD [194]. Recently, a protective role for type 1 IFN (IFNβ) has been shown, where M. abscessus clearance in macrophages was facilitated through the production of NO in a NO-dependent fashion [195]. The same authors also showed that NOD2-mediated activation of p38 and JNK, ultimately leading to NO production, can effectively clear M. abscessus in macrophages. Whereas individual immune factors may be implicated as risk factors for M. abscessus infections, the use of transgenic mice with single-gene deletion for NOS, TNF, IFNγ, or MyD88 may be compensated for with a different mechanism of the immune system [195,196,197,198]. Additionally, the route of infection greatly influences the host-immune response against M. abscessus lung infection, and so infection results [78,199]. The challenge dose of M. abscessus also requires optimization for establishing pulmonary lung infection. An aerosol infection with M. abscessus was shown to require 1 × 105–109 CFUs to enable a progressive infection in an immunocompromised mouse model [78].
The use of nude and IFNγ knock-out (GKO) mice present two models in which antibiotic therapy studies can be performed [178]. These animal models, however, are not conducive to studying the efficacy of either prophylactic or therapeutic vaccines against NTM, including M. abscessus, for which other mouse models are required. One lesser studied mouse strain for M. abscessus infection is the Beige mouse, a model for Chédiak–Higashi syndrome [200], an immune disorder characterized by impaired phagocytosis due to a mutation of a lysosomal trafficking regulator protein [201]. This mouse strain also has defective polymorphonuclear cells, monocytes, and NK cells, with delayed chemotaxis and microbicidal capacity [202]. The Beige model is considered the standard model for much slow-growing NTM, such as M. avium, as extreme susceptibility to MAC infection, has been demonstrated by us and others [168,203,204,205]. Previous studies in the Beige mouse model showed a dominant Th2 immunity that allows for MAC growth [206]. Even so, infection of Beige mice with M. abscessus does not lead to a persistent infection, which limits its utility as a model for vaccine development [194]. In a previous study on seven mouse strains, including Beige, BALB/c, Nude, GKO, A/J, Swiss, and C57BL/6 mice, most immunocompetent mice were able to rapidly clear the infection with M. abscessus (by 30 days in the lungs and 60 days in the spleen) [178]. Notably, those infections were done via the intravenous route, decreasing the chances for progressive infection. Additionally, the laboratory M. abscessus ATCC 19977 strain is often utilized in published studies. Future considerations could aim to establish progressive infection through using the aerosol route and through the use of a clinical isolate, which would likely show a higher degree of virulence. Furthermore, one could compare infection with the S and R variants of M. abscessus in the Beige mice to further shed light on the mechanism(s) by which the R variant is more virulent than the S variant in an in vivo model.
Author Contributions
Conceptualization, S.L.B. and H.F.M.A.; writing—original draft preparation, H.F.M.A., E.D.C. and L.Y.; writing—review and editing, H.F.M.A., E.D.C., L.Y., S.L.B. and R.N.C.; supervision, R.N.C.; funding acquisition, R.N.C. and S.L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This review was supported by the National Institute of Allergy and Infectious Diseases (NIAID) of the National Institute of Health (NIH) under award numbers R01AI125160, R21AI142267, and 75N93021C000029.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to express their gratitude to Seattle Children’s Research Institute Leadership for their guidance and mentorship. The authors would also like to thank Brittany Williams (SCRI) for her help in creating the figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tortoli, E.; Brown-Elliott, B.A.; Chalmers, J.D.; Cirillo, D.M.; Daley, C.L.; Emler, S.; Floto, R.A.; Garcia, M.J.; Hoefsloot, W.; Koh, W.-J.; et al. Same meat, different gravy: Ignore the new names of mycobacteria. Eur. Respir. J. 2019, 54, 1900795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adjemian, J.; Olivier, K.N.; Seitz, A.E.; Holland, S.M.; Prevots, D.R. Prevalence of Nontuberculous Mycobacterial Lung Disease in U.S. Medicare Beneficiaries. Am. J. Respir. Crit. Care Med. 2012, 185, 881–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.; E Kruijshaar, M.; Ormerod, L.P.; Drobniewski, F.; Abubakar, I. Increasing reports of non-tuberculous mycobacteria in England, Wales and Northern Ireland, 1995–2006. BMC Public Health 2010, 10, 612. [Google Scholar] [CrossRef] [PubMed]
- Brode, S.K.; Marchand-Austin, A.; Jamieson, F.B.; Marras, T.K. Pulmonary versus Nonpulmonary Nontuberculous Mycobacteria, Ontario, Canada. Emerg. Infect. Dis. 2017, 23, 1898–1901. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.D.; Herrmann, J.-L.; Kremer, L. Non-tuberculous mycobacteria and the rise of Mycobacterium abscessus. Nat. Rev. Microbiol. 2020, 18, 392–407. [Google Scholar] [CrossRef]
- Griffith, D.E.; Brown-Elliott, B.A.; Benwill, J.L.; Wallace, R.J. Mycobacterium abscessus. “Pleased to Meet You, Hope You Guess My Name…”. Ann. Am. Thorac. Soc. 2015, 12, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Bernut, A.; Herrmann, J.-L.; Kissa, K.; Dubremetz, J.-F.; Gaillard, J.-L.; Lutfalla, G.; Kremer, L. Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc. Natl. Acad. Sci. USA 2014, 111, E943–E952. [Google Scholar] [CrossRef] [Green Version]
- Jeon, K.; Kwon, O.J.; Lee, N.Y.; Kim, B.-J.; Kook, Y.-H.; Lee, S.-H.; Kil Park, Y.; Kim, C.K.; Koh, W.-J. Antibiotic Treatment of Mycobacterium abscessus Lung Disease. Am. J. Respir. Crit. Care Med. 2009, 180, 896–902. [Google Scholar] [CrossRef]
- Kothavade, R.J.; Dhurat, R.S.; Mishra, S.N.; Kothavade, U.R. Clinical and laboratory aspects of the diagnosis and management of cutaneous and subcutaneous infections caused by rapidly growing mycobacteria. Eur. J. Clin. Microbiol. 2013, 32, 161–188. [Google Scholar] [CrossRef]
- Cândido, P.H.C.; Nunes, L.D.S.; Marques, E.A.; Folescu, T.W.; Coelho, F.S.; de Moura, V.C.N.; da Silva, M.G.; Gomes, K.M.; Lourenço, M.C.D.S.; Aguiar, F.S.; et al. Multidrug-Resistant Nontuberculous Mycobacteria Isolated from Cystic Fibrosis Patients. J. Clin. Microbiol. 2014, 52, 2990–2997. [Google Scholar] [CrossRef] [Green Version]
- Ferrell, K.C.; Johansen, M.D.; Triccas, J.A.; Counoupas, C. Virulence Mechanisms of Mycobacterium abscessus: Current Knowledge and Implications for Vaccine Design. Front. Microbiol. 2022, 13, 842017. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O., III. Surrounded by mycobacteria: Nontuberculous mycobacteria in the human environment. J. Appl. Microbiol. 2009, 107, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.; Smith, B.; Brogan, O.; Rayner, A.; Harris, G.; Watt, B. Non-tuberculous mycobacteria in a hospital’s piped water supply. J. Hosp. Infect. 1992, 21, 152–154. [Google Scholar] [CrossRef]
- Le Dantec, C.; Duguet, J.-P.; Montiel, A.; Dumoutier, N.; Dubrou, S.; Vincent, V. Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems. Appl. Environ. Microbiol. 2002, 68, 5318–5325. [Google Scholar] [CrossRef] [Green Version]
- Thomson, R.; Tolson, C.; Sidjabat, H.; Huygens, F.; Hargreaves, M. Mycobacterium abscessus isolated from municipal water—A potential source of human infection. BMC Infect. Dis. 2013, 13, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierre-Audigier, C.; Ferroni, A.; Sermet-Gaudelus, I.; Le Bourgeois, M.; Offredo, C.; Vu-Thien, H.; Fauroux, B.; Mariani, P.; Munck, A.; Bingen, E.; et al. Age-Related Prevalence and Distribution of Nontuberculous Mycobacterial Species among Patients with Cystic Fibrosis. J. Clin. Microbiol. 2005, 43, 3467–3470. [Google Scholar] [CrossRef] [Green Version]
- Bryant, J.M.; Brown, K.P.; Burbaud, S.; Everall, I.; Belardinelli, J.M.; Rodriguez-Rincon, D.; Grogono, D.M.; Peterson, C.M.; Verma, D.; Evans, I.E.; et al. Stepwise pathogenic evolution of Mycobacterium abscessus. Science 2021, 372, abb8699. [Google Scholar] [CrossRef]
- Laencina, L.; Dubois, V.; Le Moigne, V.; Viljoen, A.; Majlessi, L.; Pritchard, J.; Bernut, A.; Piel, L.; Roux, A.-L.; Gaillard, J.-L.; et al. Identification of genes required for Mycobacterium abscessus growth in vivo with a prominent role of the ESX-4 locus. Proc. Natl. Acad. Sci. USA 2018, 115, E1002–E1011. [Google Scholar] [CrossRef] [Green Version]
- Dubois, V.; Pawlik, A.; Bories, A.; Le Moigne, V.; Sismeiro, O.; Legendre, R.; Varet, H.; Rodríguez-Ordóñez, M.D.P.; Gaillard, J.-L.; Coppée, J.-Y.; et al. Mycobacterium abscessus virulence traits unraveled by transcriptomic profiling in amoeba and macrophages. PLoS Pathog. 2019, 15, e1008069. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, A.; Garnier, G.; Orgeur, M.; Tong, P.; Lohan, A.; Le Chevalier, F.; Sapriel, G.; Roux, A.-L.; Conlon, K.; Honoré, N.; et al. Identification and characterization of the genetic changes responsible for the characteristic smooth-to-rough morphotype alterations of clinically persistent Mycobacterium abscessus. Mol. Microbiol. 2013, 90, 612–629. [Google Scholar] [CrossRef]
- Sondén, B.; Kocíncová, D.; Deshayes, C.; Euphrasie, D.; Rhayat, L.; Laval, F.; Frehel, C.; Daffé, M.; Etienne, G.; Reyrat, J.-M. Gap, a mycobacterial specific integral membrane protein, is required for glycolipid transport to the cell surface. Mol. Microbiol. 2005, 58, 426–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoades, E.R.; Archambault, A.S.; Greendyke, R.; Hsu, F.-F.; Streeter, C.; Byrd, T.F. Mycobacterium abscessus Glycopeptidolipids Mask Underlying Cell Wall Phosphatidyl-myo-Inositol Mannosides Blocking Induction of Human Macrophage TNF-α by Preventing Interaction with TLR2. J. Immunol. 2009, 183, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, A.V.; Viljoen, A.; Ghigo, E.; Herrmann, J.-L.; Kremer, L. Glycopeptidolipids, a Double-Edged Sword of the Mycobacterium abscessus Complex. Front. Microbiol. 2018, 9, 1145. [Google Scholar] [CrossRef] [PubMed]
- Halloum, I.; Carrère-Kremer, S.; Blaise, M.; Viljoen, A.; Bernut, A.; Le Moigne, V.; Vilchèze, C.; Guérardel, Y.; Lutfalla, G.; Herrmann, J.-L.; et al. Deletion of a Dehydratase Important for Intracellular Growth and Cording Renders Rough Mycobacterium abscessus Avirulent. Proc. Natl. Acad. Sci. USA 2016, 113, E4228–E4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whang, J.; Back, Y.W.; Lee, K.-I.; Fujiwara, N.; Paik, S.; Choi, C.H.; Park, J.-K.; Kim, H.-J. Mycobacterium abscessus glycopeptidolipids inhibit macrophage apoptosis and bacterial spreading by targeting mitochondrial cyclophilin D. Cell Death Dis. 2017, 8, e3012. [Google Scholar] [CrossRef] [PubMed]
- Kam, J.Y.; Hortle, E.; Krogman, E.; Warner, S.E.; Wright, K.; Luo, K.; Cheng, T.; Cholan, P.M.; Kikuchi, K.; Triccas, J.A.; et al. Rough and smooth variants of Mycobacterium abscessus are differentially controlled by host immunity during chronic infection of adult zebrafish. Nat. Commun. 2022, 13, 952. [Google Scholar] [CrossRef]
- Bernut, A.; Viljoen, A.; Dupont, C.; Sapriel, G.; Blaise, M.; Bouchier, C.; Brosch, R.; de Chastellier, C.; Herrmann, J.-L.; Kremer, L. Insights into the smooth-to-rough transitioning in Mycobacterium bolletii unravels a functional Tyr residue conserved in all mycobacterial MmpL family members. Mol. Microbiol. 2016, 99, 866–883. [Google Scholar] [CrossRef] [Green Version]
- Dubois, V.; Viljoen, A.; Laencina, L.; Le Moigne, V.; Bernut, A.; Dubar, F.; Blaise, M.; Gaillard, J.-L.; Guérardel, Y.; Kremer, L.; et al. MmpL8 MAB controls Mycobacterium abscessus virulence and production of a previously unknown glycolipid family. Proc. Natl. Acad. Sci. USA 2018, 115, E10147–E10156. [Google Scholar] [CrossRef] [Green Version]
- Winthrop, K.L.; Marras, T.K.; Adjemian, J.; Zhang, H.; Wang, P.; Zhang, Q. Incidence and Prevalence of Nontuberculous Mycobacterial Lung Disease in a Large U.S. Managed Care Health Plan, 2008–2015. Ann. Am. Thorac. Soc. 2020, 17, 178–185. [Google Scholar] [CrossRef]
- Rampacci, E.; Stefanetti, V.; Passamonti, F.; Henao-Tamayo, M. Preclinical Models of Nontuberculous Mycobacteria Infection for Early Drug Discovery and Vaccine Research. Pathogens 2020, 9, 641. [Google Scholar] [CrossRef]
- Kim, Y.M.; Kim, M.; Kim, S.K.; Park, K.; Jin, S.-H.; Lee, U.S.; Kim, Y.; Chae, G.T.; Lee, S.-B. Mycobacterial infections in coal workers’ pneumoconiosis patients in South Korea. Scand. J. Infect. Dis. 2009, 41, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, D.Y. Pulmonary Mycobacterial Infections due to Mycobacterium intracellulare-avium Complex. Chest 1979, 75, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, P.; Murray, J.; Glynn, J.; Thomas, R.G.; Godfrey-Faussett, P.; Shearer, S. Risk factors for pulmonary disease due to culture-positive M. tuberculosis or nontuberculous mycobacteria in South African gold miners. Eur. Respir. J. 2000, 15, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andréjak, C.; Nielsen, R.; Thomsen, V.; Duhaut, P.; Sørensen, H.T.; Thomsen, R.W. Chronic respiratory disease, inhaled corticosteroids and risk of non-tuberculous mycobacteriosis. Thorax 2013, 68, 256–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, E.D.; Kaminska, A.M.; Gill, W.; Chmura, K.; Feldman, N.E.; Bai, X.; Floyd, C.M.; Fulton, K.E.; Huitt, G.A.; Strand, M.; et al. Alpha-1-antitrypsin (AAT) anomalies are associated with lung disease due to rapidly growing mycobacteria and AAT inhibits Mycobacterium abscessus infection of macrophages. Scand. J. Infect. Dis. 2007, 39, 690–696. [Google Scholar] [CrossRef]
- Noone, P.G.; Leigh, M.W.; Sannuti, A.; Minnix, S.L.; Carson, J.L.; Hazucha, M.; Zariwala, M.A.; Knowles, M.R. Primary Ciliary Dyskinesia. Am. J. Respir. Crit. Care Med. 2004, 169, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Griffith, D.E.; Girard, W.M.; Wallace, R.J. Clinical Features of Pulmonary Disease Caused by Rapidly Growing Mycobacteria: An Analysis of 154 Patients. Am. Rev. Respir. Dis. 1993, 147, 1271–1278. [Google Scholar] [CrossRef]
- Okumura, M.; Iwai, K.; Ogata, H.; Ueyama, M.; Kubota, M.; Aoki, M.; Kokuto, H.; Tadokoro, E.; Uchiyama, T.; Saotome, M.; et al. Clinical Factors on Cavitary and Nodular Bronchiectatic Types in Pulmonary Mycobacterium avium Complex Disease. Intern. Med. 2008, 47, 1465–1472. [Google Scholar] [CrossRef] [Green Version]
- Prince, D.S.; Peterson, D.D.; Steiner, R.M.; Gottlieb, J.E.; Scott, R.; Israel, H.L.; Figueroa, W.G.; Fish, J.E. Infection with Mycobacterium avium Complex in Patients without Predisposing Conditions. N. Engl. J. Med. 1989, 321, 863–868. [Google Scholar] [CrossRef]
- Thomson, R.M.; Armstrong, J.G.; Looke, D.F. Gastroesophageal Reflux Disease, Acid Suppression, and Mycobacterium avium Complex Pulmonary Disease. Chest 2007, 131, 1166–1172. [Google Scholar] [CrossRef] [Green Version]
- Dawrs, S.N.; Kautz, M.; Chan, E.D.; Honda, J.R. Mycobacterium abscessus and Gastroesophageal Reflux: An In Vitro Study. Am. J. Respir. Crit. Care Med. 2020, 202, 466–469. [Google Scholar] [CrossRef]
- Kim, T.; Yoon, J.H.; Yang, B.; Ryu, J.; Yoon, C.K.; Kim, Y.; Sohn, J.W.; Lee, H.; Choi, H. Healthcare Utilization and Medical Cost of Gastrointestinal Reflux Disease in Non-tuberculous Mycobacterial Pulmonary Disease: A Population-Based Study, South Korea, 2009–2017. Front. Med. 2022, 9, 793453. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.D.; Iseman, M.D. Potential association between calcified thoracic lymphadenopathy due to previous Histoplasma capsulatum infection and pulmonary Mycobacterium avium complex disease. South. Med. J. 1999, 92, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Quan, D.H.; Kwong, A.J.; Hansbro, P.M.; Britton, W.J. No smoke without fire: The impact of cigarette smoking on the immune control of tuberculosis. Eur. Respir. Rev. 2022, 31, 210252. [Google Scholar] [CrossRef] [PubMed]
- De Groote, M.A.; Johnson, L.; Podell, B.; Brooks, E.; Basaraba, R.; Gonzalez-Juarrero, M. GM-CSF knockout mice for preclinical testing of agents with antimicrobial activity against Mycobacterium abscessus. J. Antimicrob. Chemother. 2014, 69, 1057–1064. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.D.; Iseman, M.D. Slender, Older Women Appear to Be More Susceptible to Nontuberculous Mycobacterial Lung Disease. Gend. Med. 2010, 7, 5–18. [Google Scholar] [CrossRef]
- E Palmer, C.; Jablon, S.; Edwards, P.Q. Tuberculosis morbidity of young men in relation to tuberculin sensitivity and body build. Am. Rev. Tuberc. 1957, 76, 517–539. [Google Scholar] [CrossRef]
- Tverdal, A. Body mass index and incidence of tuberculosis. Eur. J. Respir. Dis. 1986, 69, 355–362. Available online: https://europepmc.org/article/med/3792471 (accessed on 15 June 2022).
- Park, J.; Cho, J.; Lee, C.-H.; Han, S.K.; Yim, J.-J. Progression and Treatment Outcomes of Lung Disease Caused by Mycobacterium abscessus and Mycobacterium massiliense. Clin. Infect. Dis. 2017, 64, 301–308. [Google Scholar] [CrossRef]
- Martiniano, S.L.; Nick, J.A.; Daley, C.L. Nontuberculous Mycobacterial Infections in Cystic Fibrosis. Clin. Chest Med. 2016, 37, 83–96. [Google Scholar] [CrossRef]
- Mojica, J.E.; Richards, C.J.; Husseini, J.S.; Hariri, L.P. Case 40-2018: A 47-Year-Old Woman with Recurrent Sinusitis, Cough, and Bronchiectasis. N. Engl. J. Med. 2018, 379, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, M.H.; van Wengen, A.; de Visser, A.W.; van Kralingen, K.W.; van Dissel, J.T.; van de Vosse, E. Pulmonary Mycobacterium abscessus: A canary in the cystic fibrosis coalmine. J. Infect. 2012, 64, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Fifor, A.; Krukowski, K.; Honda, J.R. Sex, ancestry, senescence, and aging (SAnSA) are stark drivers of nontuberculous mycobacterial pulmonary disease. J. Clin. Tuberc. Other Mycobact. Dis. 2022, 26, 100297. [Google Scholar] [CrossRef] [PubMed]
- Prevots, D.R.; Shaw, P.A.; Strickland, D.; Jackson, L.A.; Raebel, M.A.; Blosky, M.A.; Montes De Oca, R.; Shea, Y.R.; Seitz, A.E.; Holland, S.M.; et al. Nontuberculous Mycobacterial Lung Disease Prevalence at Four Integrated Health Care Delivery Systems. Am. J. Respir. Crit. Care Med. 2010, 182, 970–976. [Google Scholar] [CrossRef] [Green Version]
- Fields, L.; Reeve, K.F.; Adams, B.J.; Verhave, T. Stimulus Generalization and Equivalence Classes: A Model for Natural Categories. J. Exp. Anal. Behav. 1991, 55, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Henkle, E.; Winthrop, K.L. Nontuberculous Mycobacteria Infections in Immunosuppressed Hosts. Clin. Chest Med. 2015, 36, 91–99. [Google Scholar] [CrossRef] [Green Version]
- National Institutes of Health Consensus Development Conference Statement on Genetic Testing for Cystic Fibrosis. Genetic Testing for Cystic Fibrosis. Arch. Intern. Med. 1999, 159, 1529–1539. [Google Scholar] [CrossRef]
- Dalcin, D.; Ulanova, M. The Role of Human Beta-Defensin-2 in Pseudomonas aeruginosa Pulmonary Infection in Cystic Fibrosis Patients. Infect. Dis. Ther. 2013, 2, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Shrestha, C.L.; Kopp, B.T. Cystic fibrosis transmembrane conductance regulator (CFTR) modulators have differential effects on cystic fibrosis macrophage function. Sci. Rep. 2018, 8, 17066. [Google Scholar] [CrossRef] [Green Version]
- Assani, K.; Shrestha, C.L.; Rinehardt, H.; Zhang, S.; Robledo-Avila, F.; Wellmerling, J.; Partida-Sanchez, S.; Cormet-Boyaka, E.; Reynolds, S.D.; Schlesinger, L.S.; et al. AR-13 reduces antibiotic-resistant bacterial burden in cystic fibrosis phagocytes and improves cystic fibrosis transmembrane conductance regulator function. J. Cyst. Fibros. 2019, 18, 622–629. [Google Scholar] [CrossRef]
- Roesch, E.A.; Nichols, D.P.; Chmiel, J.F. Inflammation in cystic fibrosis: An update. Pediatr. Pulmonol. 2018, 53, S30–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziedalski, T.M.; Kao, P.N.; Henig, N.R.; Jacobs, S.S.; Ruoss, S.J. Prospective Analysis of Cystic Fibrosis Transmembrane Regulator Mutations in Adults with Bronchiectasis or Pulmonary Nontuberculous Mycobacterial Infection. Chest 2006, 130, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Ravnholt, C.; Qvist, T.; Pressler, T.; Kolpen, M.; Høiby, N. Antibody testing for Mycobacterium avium complex infection in cystic fibrosis patients. Eur. Respir. J. 2018, 52, OA510. [Google Scholar] [CrossRef]
- Chen, Y.; Armstrong, D.A.; Salas, L.A.; Hazlett, H.F.; Nymon, A.B.; Dessaint, J.A.; Aridgides, D.S.; Mellinger, D.L.; Liu, X.; Christensen, B.C.; et al. Genome-wide DNA methylation profiling shows a distinct epigenetic signature associated with lung macrophages in cystic fibrosis. Clin. Epigenet. 2018, 10, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartalija, M.; Ovrutsky, A.R.; Bryan, C.L.; Pott, G.B.; Fantuzzi, G.; Thomas, J.; Strand, M.J.; Bai, X.; Ramamoorthy, P.; Rothman, M.S.; et al. Patients with Nontuberculous Mycobacterial Lung Disease Exhibit Unique Body and Immune Phenotypes. Am. J. Respir. Crit. Care Med. 2013, 187, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, R.D.; Greenberg, D.E.; Ehrmantraut, M.E.; Guide, S.V.; Ding, L.; Shea, Y.; Brown, M.R.; Chernick, M.; Steagall, W.K.; Glasgow, C.G.; et al. Pulmonary Nontuberculous Mycobacterial Disease. Am. J. Respir. Crit. Care Med. 2008, 178, 1066–1074. [Google Scholar] [CrossRef] [Green Version]
- Tasaka, S.; Hasegawa, N.; Nishimura, T.; Yamasawa, W.; Kamata, H.; Shinoda, H.; Kimizuka, Y.; Fujiwara, H.; Hirose, H.; Ishizaka, A. Elevated Serum Adiponectin Level in Patients with Mycobacterium avium-intracellulare Complex Pulmonary Disease. Respiration 2010, 79, 383–387. [Google Scholar] [CrossRef]
- Silwal, P.; Kim, I.S.; Jo, E.-K. Autophagy and Host Defense in Nontuberculous Mycobacterial Infection. Front. Immunol. 2021, 12, 728742. [Google Scholar] [CrossRef]
- Lee, S.J.; Ryu, Y.J.; Lee, J.H.; Chang, J.H.; Shim, S.S. The Impact of Low Subcutaneous Fat in Patients with Nontuberculous Mycobacterial Lung Disease. Lung 2014, 192, 395–401. [Google Scholar] [CrossRef]
- Shin, M.S.; Ho, K.-J. Bronchiectasis in Patients With α1-Antitrypsin Deficiency. Chest 1993, 104, 1384–1386. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, K.; Nagata, N.; Maki, S.; Omori, H.; Kumazoe, H.; Ueno, K.; Matsunaga, Y.; Hara, M.; Takakura, K.; Fukumoto, N.; et al. Patients with MAC Lung Disease Have a Low Visceral Fat Area and Low Nutrient Intake. Pulm. Med. 2015, 2015, 218253. [Google Scholar] [CrossRef] [PubMed]
- Edwards, L.B.; Livesay, V.T.; Acquaviva, F.A.; Palmer, C.E. Height, Weight, Tuberculous Infection, and Tuberculous Disease. Arch. Environ. Health Int. J. 1971, 22, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Cosson, M.-A.; Bertrand, J.-B.; Martin, C.; Veziris, N.; Picard, C.; Goulvestre, C.; Coignard, S.; Benoit, J.-P.; Silvera, S.; Moro, M.-R.; et al. Temporal interferon-gamma release response to Mycobacterium kansasii infection in an anorexia nervosa patient. J. Med. Microbiol. 2012, 61, 1617–1620. [Google Scholar] [CrossRef] [PubMed]
- Portillo, K.; Morera, J. Nutritional status and eating disorders: Neglected risks factor for nontuberculous mycobacterial lung disease? Med. Hypotheses 2012, 78, 39–41. [Google Scholar] [CrossRef]
- Iseman, M.D. Mycobacterium avium and slender women: An unrequited affair. Trans. Am. Clin. Climatol. Assoc. 1998, 109, 199–204. [Google Scholar]
- Grayeb, D.E.; Chan, E.D.; Swanson, L.M.; Gibson, D.G.; Mehler, P.S. Nontuberculous mycobacterial lung infections in patients with eating disorders: Plausible mechanistic links in a case series. AME Case Rep. 2021, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Lord, G.; Matarese, G.; Howard, J.K.; Baker, R.J.; Bloom, S.R.; Lechler, R.I. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature 1998, 394, 897–901. [Google Scholar] [CrossRef]
- Ordway, D.; Henao-Tamayo, M.; Smith, E.; Shanley, C.; Harton, M.; Troudt, J.; Bai, X.; Basaraba, R.J.; Orme, I.M.; Chan, E.D. Animal model of Mycobacterium abscessus lung infection. J. Leukoc. Biol. 2008, 83, 1502–1511. [Google Scholar] [CrossRef]
- Guide, S.V.; Holland, S.M. Host susceptibility factors in mycobacterial infection: Genetics and body morphotype. Infect. Dis. Clin. N. Am. 2002, 16, 163–186. [Google Scholar] [CrossRef]
- Iseman, M.D.; Buschman, D.L.; Ackerson, L.M. Pectus Excavatum and Scoliosis: Thoracic Anomalies Associated with Pulmonary Disease Caused by Mycobacterium avium Complex. Am. Rev. Respir. Dis. 1991, 144, 914–916. [Google Scholar] [CrossRef]
- Szymanski, E.P.; Leung, J.M.; Fowler, C.J.; Haney, C.; Hsu, A.P.; Chen, F.; Duggal, P.; Oler, A.J.; McCormack, R.; Podack, E.; et al. Pulmonary Nontuberculous Mycobacterial Infection. A Multisystem, Multigenic Disease. Am. J. Respir. Crit. Care Med. 2015, 192, 618–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, M.L.A.; Birchard, K.R.; Lowe, J.R.; Patrone, M.V.; Noone, P.G.; Knowles, M.R. Enlarged Dural Sac in Idiopathic Bronchiectasis Implicates Heritable Connective Tissue Gene Variants. Ann. Am. Thorac. Soc. 2016, 13, 1712–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbert, J.; Chan, E.D. The association between body shape and nontuberculous mycobacterial lung disease. Expert Rev. Respir. Med. 2013, 7, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Champsi, J.; Young, L.S.; Bermudez, L.E. Production of TNF-alpha, IL-6 and TGF-beta, and expression of receptors for TNF-alpha and IL-6, during murine Mycobacterium avium infection. Immunology 1995, 84, 549. [Google Scholar] [PubMed]
- Denis, M.; Ghadirian, E. Transforming growth factor beta (TGF-b1) plays a detrimental role in the progression of experimental Mycobacterium avium infection; in vivo and in vitro evidence. Microb. Pathog. 1991, 11, 367–372. [Google Scholar] [CrossRef]
- Paulson, M.L.; Olivier, K.N.; Holland, S.M. Pulmonary non-tuberculous mycobacterial infection in congenital contractural arachnodactyly. Int. J. Tuberc. Lung Dis. 2012, 16, 561–563. [Google Scholar] [CrossRef] [Green Version]
- Fowler, C.J.; Olivier, K.N.; Leung, J.M.; Smith, C.C.; Huth, A.G.; Root, H.; Kuhns, D.B.; Logun, C.; Zelazny, A.; Frein, C.A.; et al. Abnormal Nasal Nitric Oxide Production, Ciliary Beat Frequency, and Toll-like Receptor Response in Pulmonary Nontuberculous Mycobacterial Disease Epithelium. Am. J. Respir. Crit. Care Med. 2013, 187, 1374–1381. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Szymanski, E.P.; Olivier, K.N.; Liu, X.; Tettelin, H.; Holland, S.M.; Duggal, P. Whole-Exome Sequencing Identifies the 6q12-q16 Linkage Region and a Candidate Gene, TTK, for Pulmonary Nontuberculous Mycobacterial Disease. Am. J. Respir. Crit. Care Med. 2017, 196, 1599–1604. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, O.; Iwama, A.; Amitani, R.; Takehara, T.; Yamaguchi, N.; Yamamoto, T.; Masuyama, K.; Yamanaka, T.; Ando, M.; Suda, T. Role of macrophage-stimulating protein and its receptor, RON tyrosine kinase, in ciliary motility. J. Clin. Investig. 1997, 99, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Takano, Y.; Sakamoto, O.; Suga, M.; Suda, T.; Ando, M. Elevated levels of macrophage-stimulating protein in induced sputum of patients with bronchiectasis. Respir. Med. 2000, 94, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Doucette, K.; Fishman, J.A. Nontuberculous Mycobacterial Infection in Hematopoietic Stem Cell and Solid Organ Transplant Recipients. Clin. Infect. Dis. 2004, 38, 1428–1439. [Google Scholar] [CrossRef] [PubMed]
- French, A.L.; Benator, D.A.; Gordin, F.M. Nontuberculous Mycobacterial Infections. Med. Clin. N. Am. 1997, 81, 361–379. [Google Scholar] [CrossRef]
- Winthrop, K.L.; Baxter, R.; Liu, L.; Varley, C.D.; Curtis, J.R.; Baddley, J.W.; McFarland, B.; Austin, D.; Radcliffe, L.; Suhler, E.; et al. Mycobacterial diseases and antitumour necrosis factor therapy in USA. Ann. Rheum. Dis. 2013, 72, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, M.; Harada, N.; Watanabe, J.; Yoshikawa, H.; Shirai, Y.; Komura, M.; Koyama, M.; Ito, J.; Tsukune, Y.; Horimoto, Y.; et al. Disseminated nontuberculous mycobacteriosis and fungemia after second delivery in a patient with MonoMAC syndrome/GATA2 mutation: A case report. BMC Infect. Dis. 2021, 21, 502. [Google Scholar] [CrossRef]
- Hsu, A.; Sampaio, E.P.; Khan, J.; Calvo, K.; Lemieux, J.E.; Patel, S.; Frucht, D.M.; Vinh, D.; Auth, R.D.; Freeman, A.F.; et al. Mutations in GATA2 are associated with the autosomal dominant and sporadic monocytopenia and mycobacterial infection (MonoMAC) syndrome. Blood 2011, 118, 2653–2655. [Google Scholar] [CrossRef]
- Bogunovic, D.; Byun, M.; Durfee, L.A.; Abhyankar, A.; Sanal, O.; Mansouri, D.; Salem, S.; Radovanovic, I.; Grant, A.V.; Adimi, P.; et al. Mycobacterial Disease and Impaired IFN-γ Immunity in Humans with Inherited ISG15 Deficiency. Science 2012, 337, 1684–1688. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, J.; Arias, A.A.; Vogt, G.; Picard, C.; Galicia, L.B.; Prando, C.; Grant, A.V.; Marchal, C.C.; Hubeau, M.; Chapgier, A.; et al. Germline CYBB mutations that selectively affect macrophages in kindreds with X-linked predisposition to tuberculous mycobacterial disease. Nat. Immunol. 2011, 12, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Okada, S.; Markle, J.G.; Deenick, E.K.; Mele, F.; Averbuch, D.; Lagos, M.; Alzahrani, M.; Al-Muhsen, S.; Halwani, R.; Ma, C.S.; et al. Impairment of immunity to Candida and Mycobacterium in humans with bi-allelic RORC mutations. Science 2015, 349, 606–613. [Google Scholar] [CrossRef] [Green Version]
- Prando, C.; Samarina, A.; Bustamante, J.; Boisson-Dupuis, S.; Cobat, A.; Picard, C.; AlSum, Z.; Al-Jumaah, S.; Al-Hajjar, S.; Frayha, H.; et al. Inherited IL-12p40 Deficiency. Medicine 2013, 92, 109–122. [Google Scholar] [CrossRef]
- Bustamante, J.; Boisson-Dupuis, S.; Abel, L.; Casanova, J.-L. Mendelian susceptibility to mycobacterial disease: Genetic, immunological, and clinical features of inborn errors of IFN-γ immunity. Semin. Immunol. 2014, 26, 454–470. [Google Scholar] [CrossRef] [Green Version]
- Döffinger, R.; Jouanguy, E.; Dupuis, S.; Fondanèche, M.; Stephan, J.; Emile, J.; Lamhamedi-Cherradi, S.; Altare, F.; Pallier, A.; Barcenas-Morales, G.; et al. Partial Interferon-γ Receptor Signaling Chain Deficiency in a Patient with Bacille Calmette-Guérin and Mycobacterium abscessus Infection. J. Infect. Dis. 2000, 181, 379–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E Dorman, S.; Picard, C.; Lammas, D.; Heyne, K.; van Dissel, J.T.; Baretto, R.; Rosenzweig, S.D.; Newport, M.; Levin, M.; Roesler, J.; et al. Clinical features of dominant and recessive interferon γ receptor 1 deficiencies. Lancet 2004, 364, 2113–2121. [Google Scholar] [CrossRef]
- Dupuis, S.; Dargemont, C.; Fieschi, C.; Thomassin, N.; Rosenzweig, S.; Harris, J.; Holland, S.M.; Schreiber, R.D.; Casanova, J.-L. Impairment of Mycobacterial but Not Viral Immunity by a Germline Human STAT1 Mutation. Science 2001, 293, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Filipe-Santos, O.; Bustamante, J.; Haverkamp, M.H.; Vinolo, E.; Ku, C.-L.; Puel, A.; Frucht, D.M.; Christel, K.; Von Bernuth, H.; Jouanguy, E.; et al. X-linked susceptibility to mycobacteria is caused by mutations in NEMO impairing CD40-dependent IL-12 production. J. Exp. Med. 2006, 203, 1745–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hambleton, S.; Salem, S.; Bustamante, J.; Bigley, V.; Boisson-Dupuis, S.; Azevedo, J.; Fortin, A.; Haniffa, M.; Ceron-Gutierrez, L.; Bacon, C.M.; et al. IRF8Mutations and Human Dendritic-Cell Immunodeficiency. N. Engl. J. Med. 2011, 365, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, M.H.; van de Vosse, E.; van Dissel, J.T. Nontuberculous mycobacterial infections in children with inborn errors of the immune system. J. Infect. 2014, 68, S134–S150. [Google Scholar] [CrossRef]
- Hsu, A.; Johnson, K.D.; Falcone, E.L.; Sanalkumar, R.; Sanchez, L.; Hickstein, D.D.; Cuellar-Rodriguez, J.; Lemieux, J.E.; Zerbe, C.S.; Bresnick, E.H.; et al. GATA2 haploinsufficiency caused by mutations in a conserved intronic element leads to MonoMAC syndrome. Blood 2013, 121, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Minegishi, Y.; Saito, M.; Morio, T.; Watanabe, K.; Agematsu, K.; Tsuchiya, S.; Takada, H.; Hara, T.; Kawamura, N.; Ariga, T.; et al. Human Tyrosine Kinase 2 Deficiency Reveals Its Requisite Roles in Multiple Cytokine Signals Involved in Innate and Acquired Immunity. Immunity 2006, 25, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Chi, C.-Y.; Chu, C.-C.; Liu, J.-P.; Lin, C.-H.; Ho, M.-W.; Lo, W.-J.; Lin, P.-C.; Chen, H.-J.; Chou, C.-H.; Feng, J.-Y.; et al. Anti–IFN-γ autoantibodies in adults with disseminated nontuberculous mycobacterial infections are associated with HLA-DRB1*16:02 and HLA-DQB1*05:02 and the reactivation of latent varicella-zoster virus infection. Blood 2013, 121, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.-W.; Jo, K.-W.; Kang, B.-H.; Kim, M.Y.; Yoo, B.; Lee, C.-K.; Kim, Y.-G.; Yang, S.-K.; Byeon, J.-S.; Kim, K.-J.; et al. Mycobacterial diseases developed during anti-tumour necrosis factor-α therapy. Eur. Respir. J. 2014, 44, 1289–1295. [Google Scholar] [CrossRef]
- Mufti, A.H.; Toye, B.W.; Mckendry, R.R.; Angel, J.B. Mycobacterium abscessus infection after use of tumor necrosis factor α inhibitor therapy: Case report and review of infectious complications associated with tumor necrosis factor α inhibitor use. Diagn. Microbiol. Infect. Dis. 2005, 53, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Differences between anti-tumor necrosis factor-α monoclonal antibodies and soluble TNF receptors in host defense impairment. J. Rheumatol. 2005, 32, 40–47. [Google Scholar]
- Yadav, M.; Schorey, J.S. The β-glucan receptor dectin-1 functions together with TLR2 to mediate macrophage activation by mycobacteria. Blood 2006, 108, 3168–3175. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Yuk, J.; Kim, K.; Jang, J.; Kang, G.; Park, J.B.; Son, J.; Jo, E. Mycobacterium abscessus activates the NLRP3 inflammasome via Dectin-1–Syk and p62/SQSTM1. Immunol. Cell Biol. 2012, 90, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.-M.; Yang, C.-S.; Yuk, J.-M.; Lee, J.-Y.; Kim, K.H.; Shin, S.J.; Takahara, K.; Lee, S.J.; Jo, E.-K. Mycobacterium abscessus activates the macrophage innate immune response via a physical and functional interaction between TLR2 and dectin-1. Cell. Microbiol. 2008, 10, 1608–1621. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Kang, M.-J.; Kim, W.S.; Han, S.J.; Kim, H.M.; Kim, H.W.; Kwon, K.W.; Kim, S.J.; Bin Cha, S.; Eum, S.-Y.; et al. Essential Engagement of Toll-like Receptor 2 in Initiation of Early Protective Th1 Response against Rough Variants of Mycobacterium abscessus. Infect. Immun. 2015, 83, 1556–1567. [Google Scholar] [CrossRef] [Green Version]
- Seiler, P.; Aichele, P.; Raupach, B.; Odermatt, B.; Steinhoff, U.; Kaufmann, S.H.E. Rapid Neutrophil Response Controls Fast-Replicating Intracellular Bacteria but Not Slow-Replicating Mycobacterium tuberculosis. J. Infect. Dis. 2000, 181, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Lange, C.; Böttger, E.C.; Cambau, E.; E Griffith, D.; Guglielmetti, L.; van Ingen, J.; Knight, S.L.; Marras, T.K.; Olivier, K.N.; Santin, M.; et al. Consensus management recommendations for less common non-tuberculous mycobacterial pulmonary diseases. Lancet Infect. Dis. 2022, 22, e178–e190. [Google Scholar] [CrossRef]
- Lee, M.-C.; Sun, P.-L.; Wu, T.-L.; Wang, L.-H.; Yang, C.-H.; Hung, S.-I.; Kuo, A.-J.; Liu, T.-P.; Lu, J.-J.; Chiu, C.-H.; et al. Antimicrobial resistance in Mycobacterium abscessus complex isolated from patients with skin and soft tissue infections at a tertiary teaching hospital in Taiwan. J. Antimicrob. Chemother. 2017, 72, 2782–2786. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-R.; Sheng, W.-H.; Hung, C.-C.; Yu, C.-J.; Lee, L.-N.; Hsueh, P.-R. Mycobacterium abscessus Complex Infections in Humans. Emerg. Infect. Dis. 2015, 21, 1638–1646. [Google Scholar] [CrossRef]
- Carneiro, M.D.S.; Nunes, L.D.S.; De David, S.M.M.; Barth, A.L. Lack of association between rrl and erm(41) mutations and clarithromycin resistance in Mycobacterium abscessus complex. Mem. Inst. Oswaldo Cruz 2017, 112, 775–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, K.A.; Zhang, Y.; Brown-Elliott, B.A.; Wallace, R.J. Molecular basis of intrinsic macrolide resistance in clinical isolates of Mycobacterium fortuitum. J. Antimicrob. Chemother. 2005, 55, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.-E.; Shin, S.J.; Won, C.-J.; Min, K.-N.; Oh, T.; Hahn, M.-Y.; Lee, K.; Lee, S.H.; Daley, C.L.; Kim, S.; et al. Macrolide Treatment for Mycobacterium abscessus and Mycobacterium massiliense Infection and Inducible Resistance. Am. J. Respir. Crit. Care Med. 2012, 186, 917–925. [Google Scholar] [CrossRef]
- van Ingen, J.; Egelund, E.F.; Levin, A.; Totten, S.E.; Boeree, M.J.; Mouton, J.W.; Aarnoutse, R.E.; Heifets, L.B.; Peloquin, C.A.; Daley, C.L. The Pharmacokinetics and Pharmacodynamics of Pulmonary Mycobacterium avium Complex Disease Treatment. Am. J. Respir. Crit. Care Med. 2012, 186, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, C.-K.; Bae, I.K.; Jeong, S.H.; Yim, J.-J.; Jung, J.Y.; Park, M.S.; Kim, Y.S.; Kim, S.K.; Chang, J.; et al. The drug susceptibility profile and inducible resistance to macrolides of Mycobacterium abscessus and Mycobacterium massiliense in Korea. Diagn. Microbiol. Infect. Dis. 2015, 81, 107–111. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Vasireddy, S.; Vasireddy, R.; Iakhiaeva, E.; Howard, S.T.; Nash, K.; Parodi, N.; Strong, A.; Gee, M.; Smith, T.; et al. Utility of Sequencing the erm (41) Gene in Isolates of Mycobacterium abscessus subsp. abscessus with Low and Intermediate Clarithromycin MICs. J. Clin. Microbiol. 2015, 53, 1211–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dokic, A.; Peterson, E.; Arrieta-Ortiz, M.L.; Pan, M.; Di Maio, A.; Baliga, N.; Bhatt, A. Mycobacterium abscessus biofilms produce an extracellular matrix and have a distinct mycolic acid profile. Cell Surf. 2021, 7, 100051. [Google Scholar] [CrossRef]
- Cullen, A.R.; Cannon, C.L.; Mark, E.J.; Colin, A.A. Mycobacterium abscessus Infection in Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2000, 161, 641–645. [Google Scholar] [CrossRef]
- Broda, A.; Jebbari, H.; Beaton, K.; Mitchell, S.; Drobniewski, F. Comparative Drug Resistance of Mycobacterium abscessus and M. chelonae Isolates from Patients with and without Cystic Fibrosis in the United Kingdom. J. Clin. Microbiol. 2013, 51, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Tomashefski Jr, J.F.; Stern, R.C.; Demko, C.A.; Doershuk, C.F. Nontuberculous mycobacteria in cystic fibrosis. An autopsy study. Am. J. Respir. Crit. Care Med. 2012, 154, 523–528. [Google Scholar] [CrossRef]
- Saxena, S.; Spaink, H.P.; Forn-Cuní, G. Drug Resistance in Nontuberculous Mycobacteria: Mechanisms and Models. Biology 2021, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Shahraki, A.H.; Mirsaeidi, M. Phage Therapy for Mycobacterium abscessus and Strategies to Improve Outcomes. Microorganisms 2021, 9, 596. [Google Scholar] [CrossRef]
- Senhaji-Kacha, A.; Esteban, J.; Garcia-Quintanilla, M. Considerations for Phage Therapy Against Mycobacterium abscessus. Front. Microbiol. 2021, 11, 609017. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.M.; Smith, B.E.; Cristinziano, M.; Freeman, K.G.; Jacobs-Sera, D.; Belessis, Y.; Whitney Brown, A.; Cohen, K.A.; Davidson, R.M.; van Duin, D.; et al. Phage Therapy of Mycobacterium Infections: Compassionate-use of Phages in Twenty Patients with Drug-Resistant Mycobacterial Disease. Clin. Infect. Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Xu, L.; Tan, F.; Zhang, Y.; Fan, J.; Wang, X.; Zhang, Z.; Li, B.; Chu, H. A Novel Oxazolidinone, Contezolid (MRX-I), Expresses Anti-Mycobacterium abscessus Activity In Vitro. Antimicrob. Agents Chemother. 2021, 65, e00889-21. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Ge, Y.; Hafkin, B. Single- and Multiple-Dose Study to Determine the Safety, Tolerability, Pharmacokinetics, and Food Effect of Oral MRX-I versus Linezolid in Healthy Adult Subjects. Antimicrob. Agents Chemother. 2017, 61, e02181-16. [Google Scholar] [CrossRef] [Green Version]
- Wallis, R.S.; Jakubiec, W.; Kumar, V.; Bedarida, G.; Silvia, A.; Paige, D.; Zhu, T.; Mitton-Fry, M.; Ladutko, L.; Campbell, S.; et al. Biomarker-Assisted Dose Selection for Safety and Efficacy in Early Development of PNU-100480 for Tuberculosis. Antimicrob. Agents Chemother. 2011, 55, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Wallis, R.S.; Jakubiec, W.M.; Kumar, V.; Silvia, A.M.; Paige, D.; Dimitrova, D.; Li, X.; Ladutko, L.; Campbell, S.; Friedland, G.; et al. Pharmacokinetics and Whole-Blood Bactericidal Activity against Mycobacterium tuberculosis of Single Doses of PNU-100480 in Healthy Volunteers. J. Infect. Dis. 2010, 202, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Ganapathy, U.S.; del Río, R.G.; Cacho-Izquierdo, M.; Ortega, F.; Lelièvre, J.; Barros-Aguirre, D.; Aragaw, W.W.; Zimmerman, M.D.; Lindman, M.; Dartois, V.; et al. A Mycobacterium tuberculosis NBTI DNA Gyrase Inhibitor Is Active against Mycobacterium abscessus. Antimicrob. Agents Chemother. 2021, 65, e01514-21. [Google Scholar] [CrossRef]
- Rimal, B.; Batchelder, H.R.; Story-Roller, E.; Panthi, C.M.; Tabor, C.; Nuermberger, E.L.; Townsend, C.A.; Lamichhane, G. T405, a New Penem, Exhibits In Vivo Efficacy against M. abscessus and Synergy with β-Lactams Imipenem and Cefditoren. Antimicrob. Agents Chemother. 2022, 66, e00536-22. [Google Scholar] [CrossRef]
- Turner, J.; Muraoka, A.; Bedenbaugh, M.; Childress, B.; Pernot, L.; Wiencek, M.; Peterson, Y.K. The Chemical Relationship Among Beta-Lactam Antibiotics and Potential Impacts on Reactivity and Decomposition. Front. Microbiol. 2022, 13, 807955. [Google Scholar] [CrossRef] [PubMed]
- Dousa, K.M.; Nguyen, D.C.; Kurz, S.G.; Taracila, M.A.; Bethel, C.R.; Schinabeck, W.; Kreiswirth, B.N.; Brown, S.T.; Boom, W.H.; Hotchkiss, R.S.; et al. Inhibiting Mycobacterium abscessus Cell Wall Synthesis: Using a Novel Diazabicyclooctane β-Lactamase Inhibitor to Augment β-Lactam Action. mBio 2022, 13, e03529-21. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.C.; Dousa, K.M.; Kurz, S.G.; Brown, S.T.; Drusano, G.; Holland, S.M.; Kreiswirth, B.N.; Boom, W.H.; Daley, C.L.; A Bonomo, R. “One-Two Punch”: Synergistic ß-Lactam Combinations for Mycobacterium abscessus and Target Redundancy in the Inhibition of Peptidoglycan Synthesis Enzymes. Clin. Infect. Dis. 2021, 73, 1532–1536. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Jhun, B.W.; Moon, S.M.; Kim, S.-Y.; Jeon, K.; Kwon, O.J.; Huh, H.J.; Lee, N.Y.; Shin, S.J.; Daley, C.L.; et al. In Vitro Activity of Bedaquiline and Delamanid against Nontuberculous Mycobacteria, Including Macrolide-Resistant Clinical Isolates. Antimicrob. Agents Chemother. 2019, 63, e00665-19. [Google Scholar] [CrossRef] [Green Version]
- Ruth, M.M.; Sangen, J.; Remmers, K.; Pennings, L.; Svensson, E.; E Aarnoutse, R.; Zweijpfenning, S.M.H.; Hoefsloot, W.; Kuipers, S.; Magis-Escurra, C.; et al. A bedaquiline/clofazimine combination regimen might add activity to the treatment of clinically relevant non-tuberculous mycobacteria. J. Antimicrob. Chemother. 2019, 74, 935–943. [Google Scholar] [CrossRef]
- Mahajan, R. Bedaquiline: First FDA-approved tuberculosis drug in 40 years. Int. J. Appl. Basic Med. Res. 2013, 3, 1–2. [Google Scholar] [CrossRef]
- Sarathy, J.P.; Ganapathy, U.S.; Zimmerman, M.D.; Dartois, V.; Gengenbacher, M.; Dick, T. TBAJ-876, a 3,5-Dialkoxypyridine Analogue of Bedaquiline, Is Active against Mycobacterium abscessus. Antimicrob. Agents Chemother. 2020, 64, e02404-19. [Google Scholar] [CrossRef] [Green Version]
- Yano, T.; Kassovska-Bratinova, S.; Teh, J.S.; Winkler, J.; Sullivan, K.; Isaacs, A.; Schechter, N.M.; Rubin, H. Reduction of Clofazimine by Mycobacterial Type 2 NADH: Quinone Oxidoreductase. J. Biol. Chem. 2011, 286, 10276–10287. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Jhun, B.W.; Moon, S.M.; Lee, H.; Park, H.Y.; Jeon, K.; Kim, D.H.; Kim, S.-Y.; Shin, S.J.; Daley, C.L.; et al. Clofazimine-Containing Regimen for the Treatment of Mycobacterium abscessus Lung Disease. Antimicrob. Agents Chemother. 2017, 61, e02052-16. [Google Scholar] [CrossRef] [Green Version]
- Khan, O.; Chaudary, N. The Use of Amikacin Liposome Inhalation Suspension (Arikayce) in the Treatment of Refractory Nontuberculous Mycobacterial Lung Disease in Adults. Drug Des. Dev. Ther. 2020, 14, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Olivier, K.N.; Shaw, P.A.; Glaser, T.S.; Bhattacharyya, D.; Fleshner, M.; Brewer, C.C.; Zalewski, C.K.; Folio, L.R.; Siegelman, J.R.; Shallom, S.; et al. Inhaled Amikacin for Treatment of Refractory Pulmonary Nontuberculous Mycobacterial Disease. Ann. Am. Thorac. Soc. 2014, 11, 30–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, K.; Ishii, M.; Namkoong, H.; Asami, T.; Iketani, O.; Asakura, T.; Suzuki, S.; Sugiura, H.; Yamada, Y.; Nishimura, T.; et al. The efficacy, safety, and feasibility of inhaled amikacin for the treatment of difficult-to-treat non-tuberculous mycobacterial lung diseases. BMC Infect. Dis. 2017, 17, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, D.E.; Eagle, G.; Thomson, R.; Aksamit, T.R.; Hasegawa, N.; Morimoto, K.; Addrizzo-Harris, D.J.; O’Donnell, A.E.; Marras, T.K.; Flume, P.A.; et al. Amikacin liposome inhalation suspension for treatment-refractory lung disease caused by Mycobacterium avium complex (CONVERT) a prospective, open-label, randomized study. Am. J. Respir. Crit. Care Med. 2018, 198, 1559–1569. [Google Scholar] [CrossRef]
- Zhang, J.; Leifer, F.; Rose, S.; Chun, D.Y.; Thaisz, J.; Herr, T.; Nashed, M.; Joseph, J.; Perkins, W.R.; DiPetrillo, K. Amikacin Liposome Inhalation Suspension (ALIS) Penetrates Non-tuberculous Mycobacterial Biofilms and Enhances Amikacin Uptake into Macrophages. Front. Microbiol. 2018, 9, 915. [Google Scholar] [CrossRef] [Green Version]
- Zweijpfenning, S.M.H.; Chiron, R.; Essink, S.; Schildkraut, J.; Akkerman, O.W.; Aliberti, S.; Altenburg, J.; Arets, B.; van Braeckel, E.; Delaere, B.; et al. Safety and Outcomes of Amikacin Liposome Inhalation Suspension for Mycobacterium abscessus Pulmonary Disease. Chest 2022, 162, 76–81. [Google Scholar] [CrossRef]
- Steinberg, B.E.; Grinstein, S. Pathogen destruction versus intracellular survival: The role of lipids as phagosomal fate determinants. J. Clin. Investig. 2008, 118, 2002–2011. [Google Scholar] [CrossRef] [Green Version]
- Auricchio, G.; Garg, S.K.; Martino, A.; Volpe, E.; Ciaramella, A.; De Vito, P.; Baldini, P.M.; Colizzi, V.; Fraziano, M. Role of macrophage phospholipase D in natural and CpG-induced antimycobacterial activity. Cell. Microbiol. 2003, 5, 913–920. [Google Scholar] [CrossRef]
- Poerio, N.; Bugli, F.; Taus, F.; Santucci, M.B.; Rodolfo, C.; Cecconi, F.; Torelli, R.; Varone, F.; Inchingolo, R.; Majo, F.; et al. Liposomes loaded with bioactive lipids enhance antibacterial innate immunity irrespective of drug resistance. Sci. Rep. 2017, 7, 45120. [Google Scholar] [CrossRef] [Green Version]
- Poerio, N.; Riva, C.; Olimpieri, T.; Rossi, M.; Lorè, N.I.; De Santis, F.; De Angelis, L.H.; Ciciriello, F.; D’Andrea, M.M.; Lucidi, V.; et al. Combined Host- and Pathogen-Directed Therapy for the Control of Mycobacterium abscessus Infection. Microbiol. Spectr. 2022, 10, e02546-21. [Google Scholar] [CrossRef]
- Abate, G.; Hamzabegovic, F.; Eickhoff, C.S.; Hoft, D.F. BCG Vaccination Induces M. avium and M. abscessus Cross-Protective Immunity. Front. Immunol. 2019, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trnka, L.; Daňková, D.; Švandová, E. Six years’ experience with the discontinuation of BCG vaccination: 4. Protective effect of BCG vaccination against the Mycobacterium avium intracellulare complex. Tuber. Lung Dis. 1994, 75, 348–352. [Google Scholar] [CrossRef]
- Orujyan, D.; Narinyan, W.; Rangarajan, S.; Rangchaikul, P.; Prasad, C.; Saviola, B.; Venketaraman, V. Protective Efficacy of BCG Vaccine against Mycobacterium leprae and Non-Tuberculous Mycobacterial Infections. Vaccines 2022, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Fujita, J.; Ohtsuki, Y.; Shigeto, E.; Suemitsu, I.; Yamadori, I.; Bandoh, S.; Shiode, M.; Nishimura, K.; Hirayama, T.; Matsushima, T.; et al. Pathological findings of bronchiectases caused by Mycobacterium avium intracellulare complex. Respir. Med. 2003, 97, 933–938. [Google Scholar] [CrossRef] [Green Version]
- Hesseling, A.C.; Schaaf, H.S.; Hanekom, W.A.; Beyers, N.; Cotton, M.F.; Gie, R.P.; Marais, B.J.; Van Helden, P.; Warren, R.M. Danish Bacille Calmette-Guerin Vaccine-Induced Disease in Human Immunodeficiency Virus-Infected Children. Clin. Infect. Dis. 2003, 37, 1226–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudes, P. Disseminated Mycobacterium bovis Infection from BCG Vaccination and HIV Infection. JAMA 1989, 262, 2386. [Google Scholar] [CrossRef]
- Orme, I.; Roberts, A.; Collins, F. Lack of evidence for a reduction in the efficacy of subcutaneous BCG vaccination in mice infected with nontuberculous mycobacteria. Tubercle 1986, 67, 41–46. [Google Scholar] [CrossRef]
- Larsen, S.E.; Reese, V.A.; Pecor, T.; Berube, B.J.; Cooper, S.K.; Brewer, G.; Ordway, D.; Henao-Tamayo, M.; Podell, B.K.; Baldwin, S.L.; et al. Subunit vaccine protects against a clinical isolate of Mycobacterium avium in wild type and immunocompromised mouse models. Sci. Rep. 2021, 11, 9040. [Google Scholar] [CrossRef]
- Baldwin, S.L.; Larsen, S.E.; Ordway, D.; Cassell, G.; Coler, R.N. The complexities and challenges of preventing and treating nontuberculous mycobacterial diseases. PLoS Negl. Trop. Dis. 2019, 13, e0007083. [Google Scholar] [CrossRef]
- Coler, R.N.; Bertholet, S.; Moutaftsi, M.; Guderian, J.A.; Windish, H.P.; Baldwin, S.L.; Laughlin, E.M.; Duthie, M.; Fox, C.; Carter, D.; et al. Development and Characterization of Synthetic Glucopyranosyl Lipid Adjuvant System as a Vaccine Adjuvant. PLoS ONE 2011, 6, e16333. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.L.; Bertholet, S.; Reese, V.A.; Ching, L.K.; Reed, S.G.; Coler, R.N. The Importance of Adjuvant Formulation in the Development of a Tuberculosis Vaccine. J. Immunol. 2012, 188, 2189–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, T.A.; Penn-Nicholson, A.; Luabeya, A.K.K.; Fiore-Gartland, A.; Du Plessis, N.; Loxton, A.G.; Vergara, J.; Rolf, T.A.; Reid, T.D.; Toefy, A.; et al. Safety and immunogenicity of the adjunct therapeutic vaccine ID93 + GLA-SE in adults who have completed treatment for tuberculosis: A randomised, double-blind, placebo-controlled, phase 2a trial. Lancet Respir. Med. 2021, 9, 373–386. [Google Scholar] [CrossRef]
- Baldwin, S.L.; Reese, V.A.; Huang, P.-W.D.; Beebe, E.A.; Podell, B.K.; Reed, S.G.; Coler, R.N. Protection and Long-Lived Immunity Induced by the ID93/GLA-SE Vaccine Candidate against a Clinical Mycobacterium tuberculosis Isolate. Clin. Vaccine Immunol. 2016, 23, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, S.E.; Baldwin, S.L.; Orr, M.T.; Reese, V.A.; Pecor, T.; Granger, B.; Cauwelaert, N.D.; Podell, B.K.; Coler, R.N. Enhanced Anti-Mycobacterium tuberculosis Immunity over Time with Combined Drug and Immunotherapy Treatment. Vaccines 2018, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Schmidt, C. The benefits of immunotherapy combinations. Nature 2017, 552, S67–S69. [Google Scholar] [CrossRef] [Green Version]
- Ramamurthy, D.; Nundalall, T.; Cingo, S.; Mungra, N.; Karaan, M.; Naran, K.; Barth, S. Recent advances in immunotherapies against infectious diseases. Immunother. Adv. 2020, 1, ltaa007. [Google Scholar] [CrossRef]
- Lerat, I.; Cambau, E.; Roth Dit Bettoni, R.; Gaillard, J.-L.; Jarlier, V.; Truffot, C.; Veziris, N. In Vivo Evaluation of Antibiotic Activity Against Mycobacterium abscessus. J. Infect. Dis. 2014, 209, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Veziris, N.; Ibrahim, M.; Lounis, N.; Andries, K.; Jarlier, V. Sterilizing Activity of Second-Line Regimens Containing TMC207 in a Murine Model of Tuberculosis. PLoS ONE 2011, 6, e17556. [Google Scholar] [CrossRef]
- Bertholet, S.; Horne, D.J.; Laughlin, E.M.; Savlov, M.; Tucakovic, I.; Coler, R.N.; Narita, M.; Reed, S.G. Effect of Chemotherapy on Whole-Blood Cytokine Responses to Mycobacterium tuberculosis Antigens in a Small Cohort of Patients with Pulmonary Tuberculosis. Clin. Vaccine Immunol. 2011, 18, 1378–1386. [Google Scholar] [CrossRef] [Green Version]
- Ji, B.; Chauffour, A.; Robert, J.; Lefrançois, S.; Jarlier, V. Orally Administered Combined Regimens for Treatment of Mycobacterium ulcerans Infection in Mice. Antimicrob. Agents Chemother. 2007, 51, 3737–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valerga, M.; Viola, C.; Thwaites, A.; Bases, O.; Ambroggi, M.; Poggi, S.; Marino, R. Mycobacterium bovis tuberculosis in a female patient with AIDS. Rev. Argent. Microbiol. 2005, 37, 96–98. [Google Scholar] [PubMed]
- Bermudez, L.E.; Nash, K.A.; Petrofsky, M.; Young, L.S.; Inderlied, C.B. Effect of Ethambutol on Emergence of Clarithromycin-Resistant Mycobacterium avium Complex in the Beige Mouse Model. J. Infect. Dis. 1996, 174, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.-E.; Ahn, J.-H.; Park, E.-K.; Jeong, H.; Lee, H.-J.; Jung, Y.-J.; Shin, S.J.; Jeong, H.-S.; Yoo, J.S.; Shin, E.; et al. B Cell-Based Vaccine Transduced with ESAT6-Expressing Vaccinia Virus and Presenting α-Galactosylceramide Is a Novel Vaccine Candidate Against ESAT6-Expressing Mycobacterial Diseases. Front. Immunol. 2019, 10, 2542. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, L.E.; Martinelli, J.; Petrofsky, M.; Kolonoski, P.; Young, L.S. Recombinant Granulocyte-Macrophage Colony-Stimulating Factor Enhances the Effects of Antibiotics against Mycobacterium avium Complex Infection in the Beige Mouse Model. J. Infect. Dis. 1994, 169, 575–580. [Google Scholar] [CrossRef]
- Gomez-Flores, R.; Tucker, S.D.; Kansal, R.; Tamez-Guerra, R.; Mehta, R.T. Enhancement of antibacterial activity of clofazimine against Mycobacterium avium-Mycobacterium intracellulare complex infection induced by IFN-gamma is mediated by TNF-alpha. J. Antimicrob. Chemother. 1997, 39, 189–197. [Google Scholar] [CrossRef]
- Richter, A.; Shapira, T.; Av-Gay, Y. THP-1 and Dictyostelium Infection Models for Screening and Characterization of Anti-Mycobacterium abscessus Hit Compounds. Antimicrob. Agents Chemother. 2020, 64, e01601-19. [Google Scholar] [CrossRef]
- Oh, C.-T.; Moon, C.; Jeong, M.S.; Kwon, S.-H.; Jang, J. Drosophila melanogaster model for Mycobacterium abscessus infection. Microbes Infect. 2013, 15, 788–795. [Google Scholar] [CrossRef]
- Meir, M.; Grosfeld, T.; Barkan, D. Establishment and Validation of Galleria mellonella as a Novel Model Organism to Study Mycobacterium abscessus Infection, Pathogenesis, and Treatment. Antimicrob. Agents Chemother. 2018, 62, e02539-17. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, K.; Koyama, N.; Hamamoto, H.; Yagi, A.; Uchida, R.; Kanamoto, A.; Tomoda, H. Evaluation of Anti-Mycobacterial Compounds in a Silkworm Infection Model with Mycobacteroides abscessus. Molecules 2020, 25, 4971. [Google Scholar] [CrossRef]
- Bernut, A.; Le Moigne, V.; Lesne, T.; Lutfalla, G.; Herrmann, J.-L.; Kremer, L. In Vivo Assessment of Drug Efficacy against Mycobacterium abscessus Using the Embryonic Zebrafish Test System. Antimicrob. Agents Chemother. 2014, 58, 4054–4063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubée, V.; Bernut, A.; Cortes, M.; Lesne, T.; Dorchene, D.; Lefebvre, A.-L.; Hugonnet, J.-E.; Gutmann, L.; Mainardi, J.-L.; Herrmann, J.-L.; et al. β-Lactamase inhibition by avibactam in Mycobacterium abscessus. J. Antimicrob. Chemother. 2014, 70, 1051–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, A.-L.; Le Moigne, V.; Bernut, A.; Veckerlé, C.; Compain, F.; Herrmann, J.-L.; Kremer, L.; Arthur, M.; Mainardi, J.-L. Inhibition of the β-Lactamase Bla Mab by Avibactam Improves the In Vitro and In Vivo Efficacy of Imipenem against Mycobacterium abscessus. Antimicrob. Agents Chemother. 2017, 61, e02440-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obregón-Henao, A.; Arnett, K.A.; Henao-Tamayo, M.; Massoudi, L.; Creissen, E.; Andries, K.; Lenaerts, A.J.; Ordway, D.J. Susceptibility of Mycobacterium abscessus to Antimycobacterial Drugs in Preclinical Models. Antimicrob. Agents Chemother. 2015, 59, 6904–6912. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.-H.; Park, J.-Y.; Kim, D.-Y.; Lee, T.-S.; Jung, D.-H.; Kim, Y.-J.; Lee, Y.-J.; Lee, Y.-J.; Seo, I.-S.; Song, E.-J.; et al. Type I Interferons Are Involved in the Intracellular Growth Control of Mycobacterium abscessus by Mediating NOD2-Induced Production of Nitric Oxide in Macrophages. Front. Immunol. 2021, 12, 738070. [Google Scholar] [CrossRef]
- Rottman, M.; Catherinot, E.; Hochedez, P.; Emile, J.-F.; Casanova, J.-L.; Gaillard, J.-L.; Soudais, C. Importance of T Cells, Gamma Interferon, and Tumor Necrosis Factor in Immune Control of the Rapid Grower Mycobacterium abscessus in C57BL/6 Mice. Infect. Immun. 2007, 75, 5898–5907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dick, T.; Shin, S.J.; Koh, W.-J.; Dartois, V.; Gengenbacher, M. Rifabutin Is Active against Mycobacterium abscessus in Mice. Antimicrob. Agents Chemother. 2020, 64, e01943-19. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Lee, M.-S.; Kim, D.-J.; Yang, S.-J.; Lee, S.-J.; Noh, E.-J.; Shin, S.J.; Park, J.-H. Nucleotide-Binding Oligomerization Domain 2 Contributes to Limiting Growth of Mycobacterium abscessus in the Lung of Mice by Regulating Cytokines and Nitric Oxide Production. Front. Immunol. 2017, 8, 1477. [Google Scholar] [CrossRef] [Green Version]
- Ordway, D.J.; Orme, I.M. Animal Models of Mycobacteria Infection. Curr. Protoc. Immunol. 2011, 94, 19.5.1–19.5.50. [Google Scholar] [CrossRef] [Green Version]
- Saunders, B.M.; Cheers, C. Intranasal infection of beige mice with Mycobacterium avium complex: Role of neutrophils and natural killer cells. Infect. Immun. 1996, 64, 4236–4241. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.; De Domenico, I.; Ward, D.M. Chediak-Higashi syndrome. Curr. Opin. Hematol. 2008, 15, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Rutledge, B.J.; Gu, L.; Fiorillo, J.; Lukacs, N.W.; Kunkel, S.L.; North, R.; Gerard, C.; Rollins, B.J. Abnormalities in Monocyte Recruitment and Cytokine Expression in Monocyte Chemoattractant Protein 1–deficient Mice. J. Exp. Med. 1998, 187, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Gangadharam, P.R. Beige mouse model for Mycobacterium avium complex disease. Antimicrob. Agents Chemother. 1995, 39, 1647–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelberg, R.; Castro, A.G.; Gomes, S.; Pedrosa, J.; Silva, M.T. Susceptibility of beige mice to Mycobacterium avium: Role of neutrophils. Infect. Immun. 1995, 63, 3381–3387. [Google Scholar] [CrossRef] [Green Version]
- Gangadharam, P.; Perumal, V.; Farhi, D.; LaBrecque, J. The beige mouse model for mycobacterium avium complex (MAC) disease: Optimal conditions for the host and parasite. Tubercle 1989, 70, 257–271. [Google Scholar] [CrossRef]
- Bermudez, L.E.; Parker, A.; Petrofsky, M. Apoptosis of Mycobacterium avium-infected macrophages is mediated by both tumour necrosis factor (TNF) and Fas, and involves the activation of caspases. Clin. Exp. Immunol. 1999, 116, 94–99. [Google Scholar] [CrossRef]
Figure 1.
Risk factors for NTM lung disease [6,16,31,32,33,35,36,38,40,41,42,43,44,45,46,47,48,49,50,51].

Figure 2.
Genetic and acquired causes of disseminated NTM disease.

Figure 3.
A schematic representation of M. abscessus current treatment protocol.

Figure 4.
Mechanism of inducible resistance in M. abscessus. In M. abscessus sensu stricto and M. bolletii, macrolide binds to 23S rRNA and inhibits bacterial protein synthesis. With induction of methylase production by clarithromycin, the methylase prevents the binding of macrolide, creating an inducible resistance. Since clarithromycin induction of the ERM41 gene to produce methylase is much greater than by azithromycin, clarithromycin is much more likely to induce macrolide resistance than azithromycin.
Figure 4.
Mechanism of inducible resistance in M. abscessus. In M. abscessus sensu stricto and M. bolletii, macrolide binds to 23S rRNA and inhibits bacterial protein synthesis. With induction of methylase production by clarithromycin, the methylase prevents the binding of macrolide, creating an inducible resistance. Since clarithromycin induction of the ERM41 gene to produce methylase is much greater than by azithromycin, clarithromycin is much more likely to induce macrolide resistance than azithromycin.


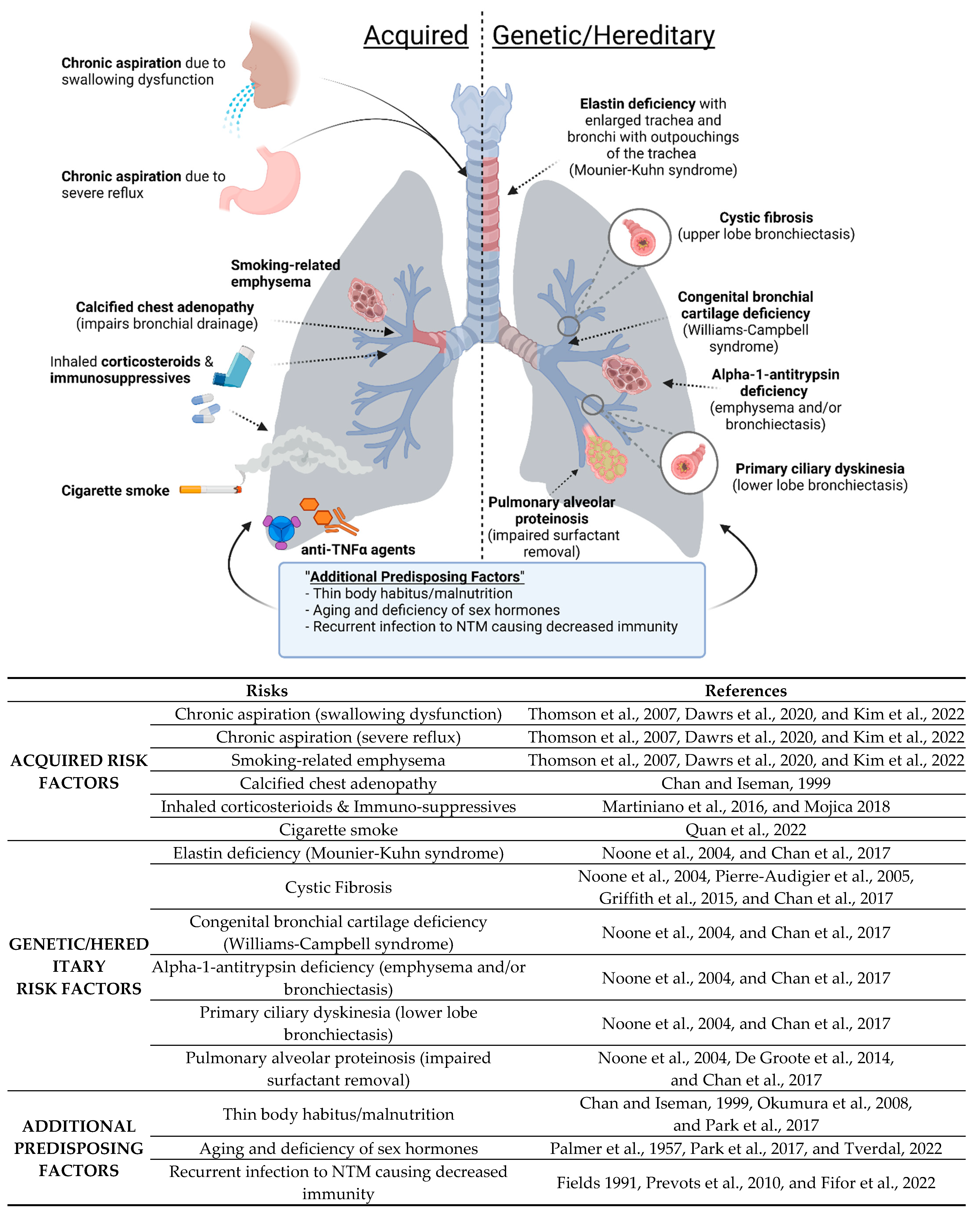{kind=link}
{kind=link}
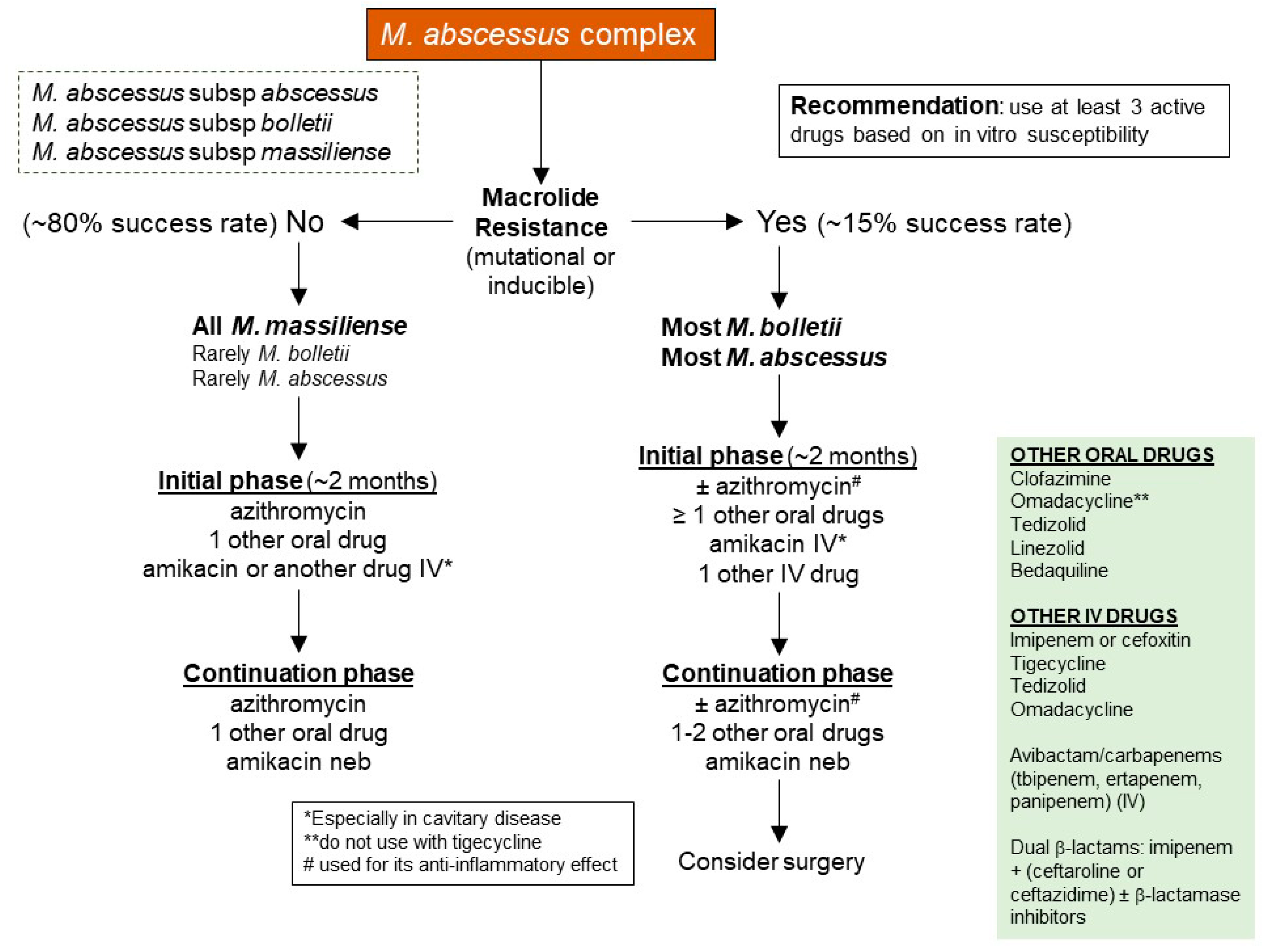{kind=link}
{kind=link}
Table 1.
Major known mechanisms that predispose to NTM-LD.
| Mechanism for Predisposition | Associated Conditions |
|---|---|
| Structural lung disease | Emphysema, bronchiectasis of any cause, including CF, alpha-1-antitrypsin deficiency (AAT), and Sjogren’s syndrome |
| Primary or ciliary dysfunction | Primary ciliary dyskinesia, bronchiectasis of any cause, MST1R dysfunction (?) |
| Thickened secretions | CF |
| Macrophage dysfunction | Alpha-1-antitrypsin (AAT) deficiency or anomaly, silica exposure, pulmonary alveolar proteinosis |
| Deficiency of specific immune molecules | Anti-TNF therapy, common variable immunodeficiency, CF (human beta-defensin?), inhaled glucocorticoid |
| Cartilage deficiency in airways | Williams–Campbell syndrome |
| Elastin deficiency in airways | Mounier-Kuhn syndrome |
MST1R = macrophage-stimulating 1 receptor.
Table 2.
Summary of different models used for M. abscessus infection.
| Type of Model | Model | Nature | Advantages | Drawbacks |
|---|---|---|---|---|
| Nonmammalian models | Amoebas (Dictyostelium discoideum) |
|
|
|
| Drosophila melanogaster |
|
|
| |
| Galleria mellonella larvae |
|
|
| |
| Zebrafish |
|
|
| |
| Silk worm |
|
|
| |
| Mammalian models | Nude Mice |
|
|
|
| GKO Mice |
|
|
| |
| Beige Mice |
|
|
| |
| C57BL/6 Mice |
|
|
| |
| BALB/c |
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abdelaal, H.F.M.; Chan, E.D.; Young, L.; Baldwin, S.L.; Coler, R.N. Mycobacterium abscessus: It’s Complex. Microorganisms 2022, 10, 1454. https://doi.org/10.3390/microorganisms10071454
AMA Style
Abdelaal HFM, Chan ED, Young L, Baldwin SL, Coler RN. Mycobacterium abscessus: It’s Complex. Microorganisms. 2022; 10(7):1454. https://doi.org/10.3390/microorganisms10071454
Chicago/Turabian StyleAbdelaal, Hazem F. M., Edward D. Chan, Lisa Young, Susan L. Baldwin, and Rhea N. Coler. 2022. "Mycobacterium abscessus: It’s Complex" Microorganisms 10, no. 7: 1454. https://doi.org/10.3390/microorganisms10071454
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.