
Co-Occurrence Relationship and Stochastic Processes Affect Sedimentary Archaeal and Bacterial Community Assembly in Estuarine–Coastal Margins

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
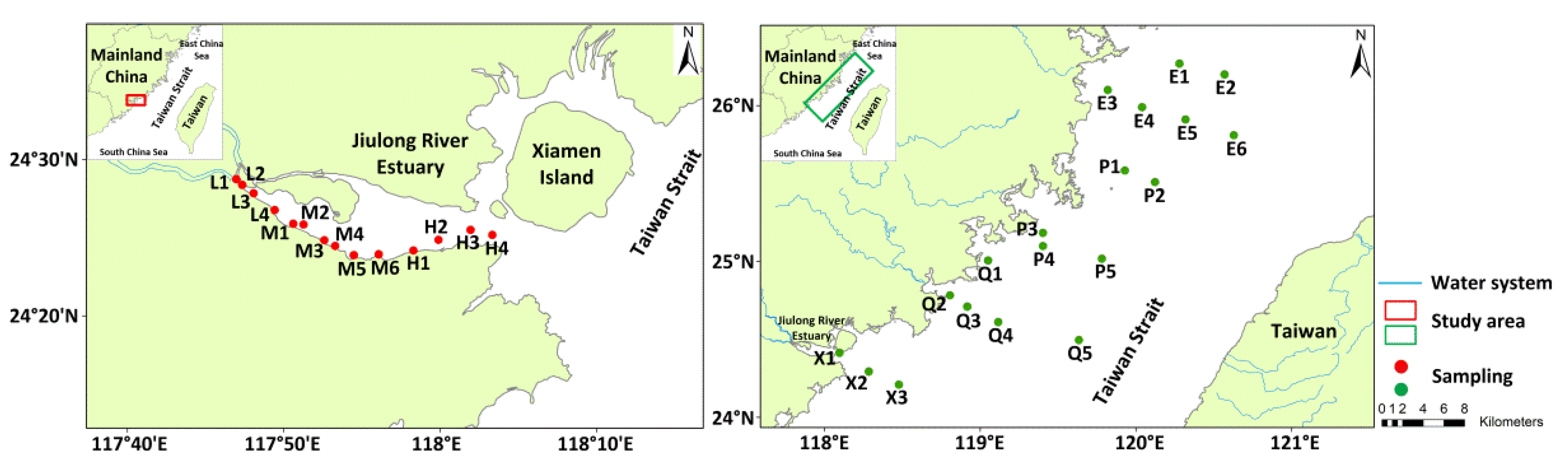
2.1. Description of Study Area
2.2. Samples Collection
2.3. Physicochemical Analysis
2.4. DNA Extraction, Polymerase Chain Reaction (PCR), and Sequencing
2.5. Quantitative PCR (qPCR)
2.6. Sequence Denoising, Operational Taxonomic Units (OTU) Clustering, and Diversity Analysis
2.7. Variation Partitioning Analysis (VPA), Null Model and Network Analysis
3. Results
3.1. Biogeochemical Parameters in the Sediments
3.2. Abundance and Diversity of Bacterial and Archaeal Community
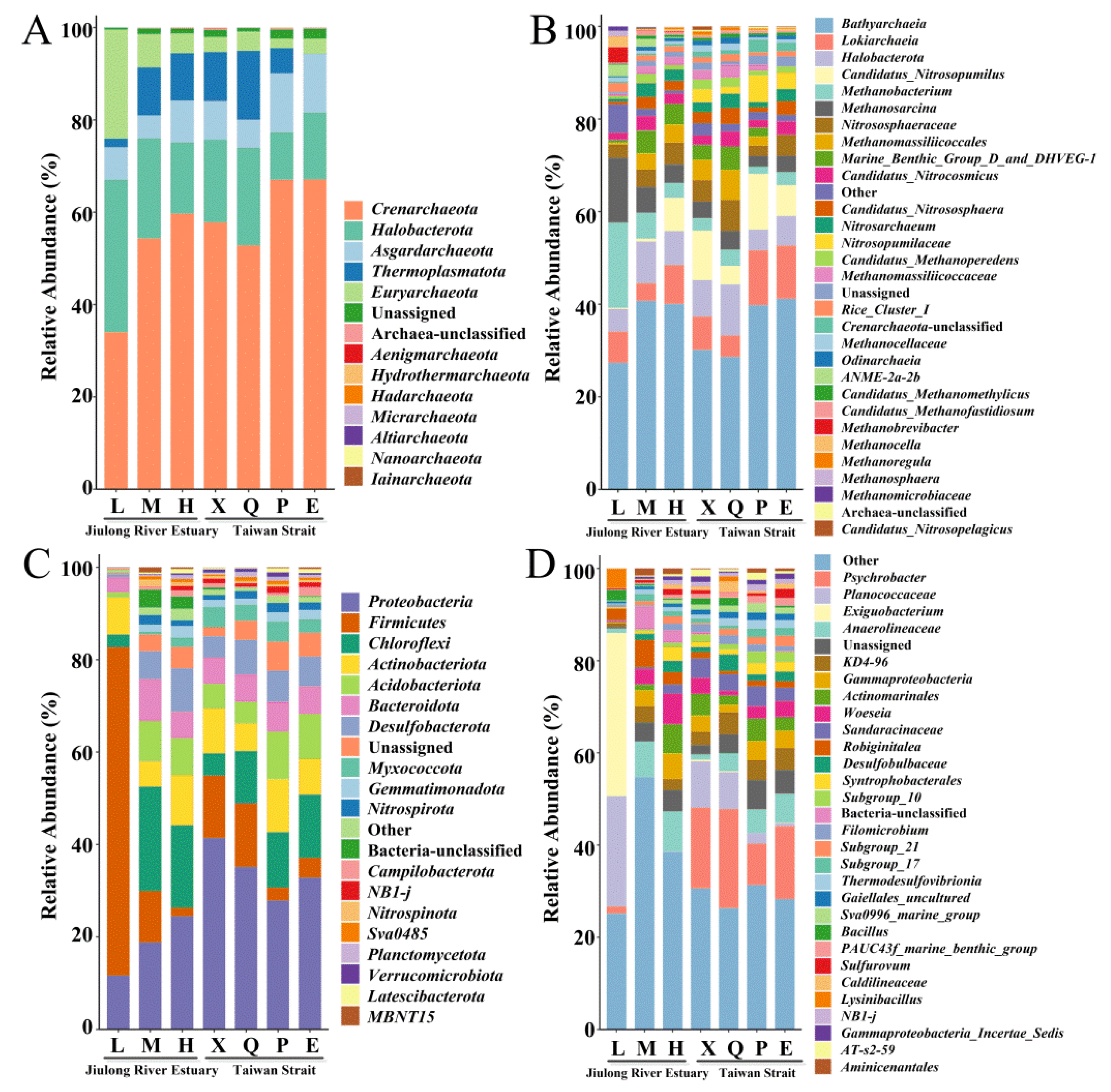
3.3. Taxonomy and Composition of Bacterial and Archaeal Community
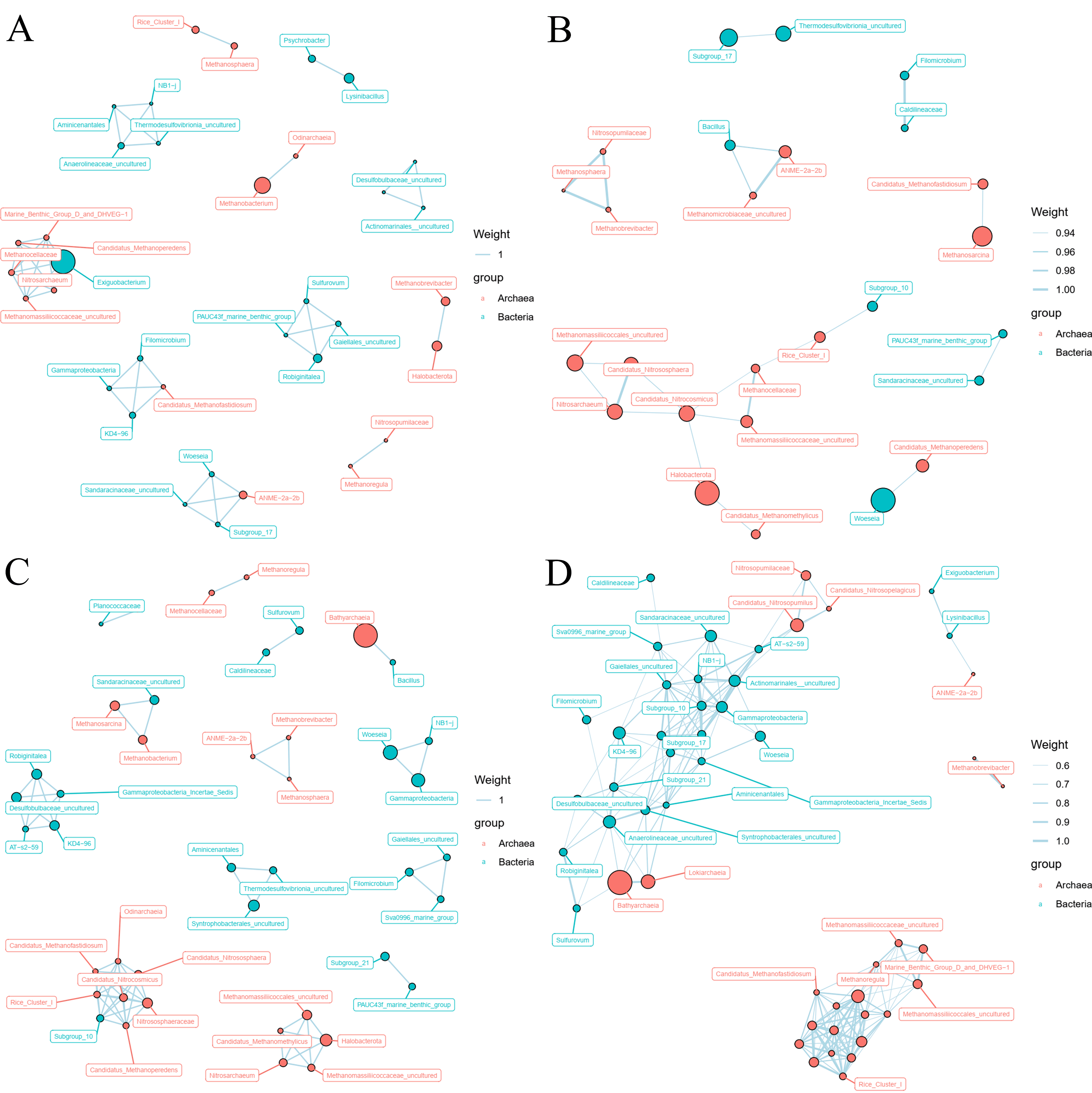
3.4. Archaeal and Bacterial Co-Occurrence Network Analysis
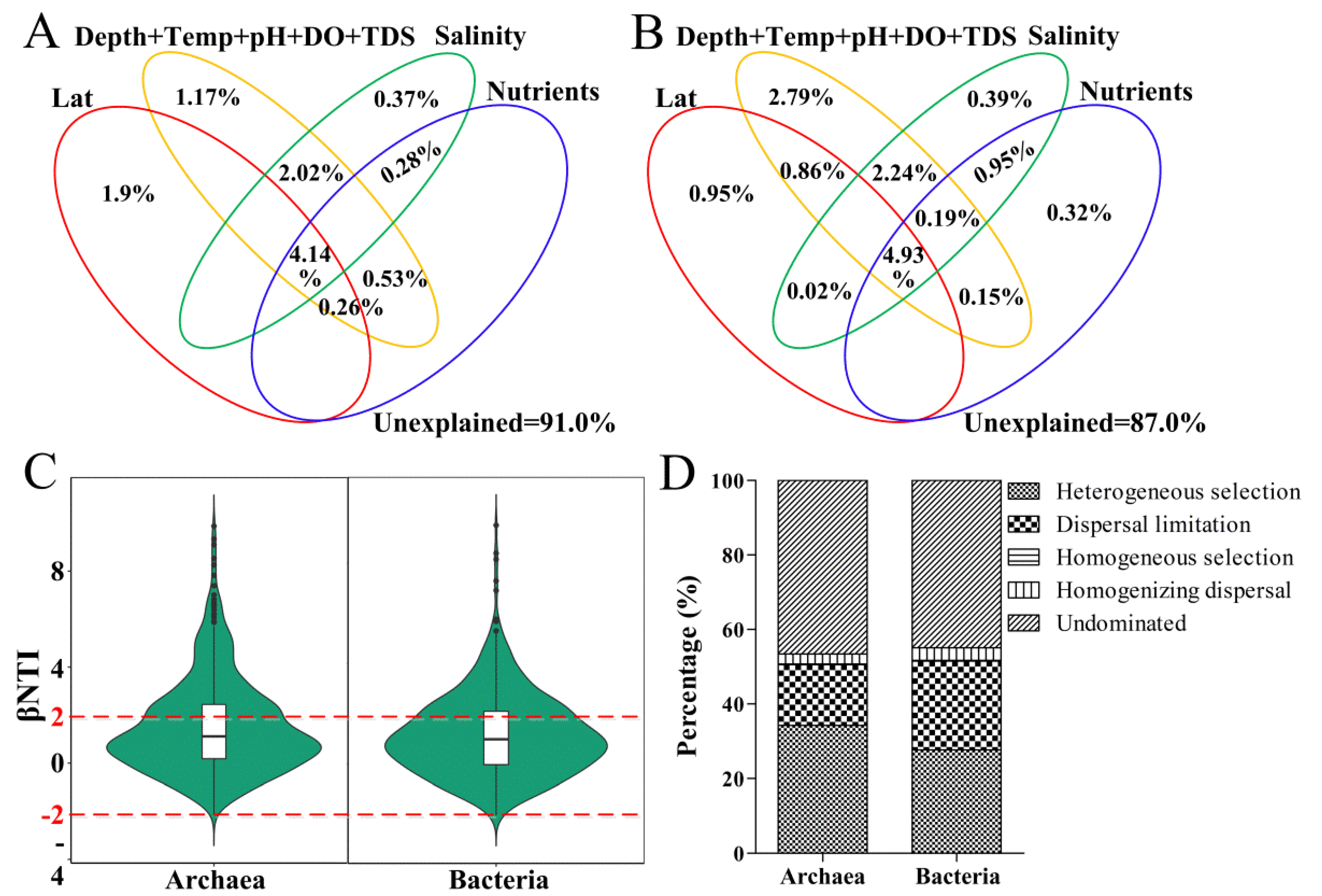
3.5. Ecological Processes Governing Community Structure
4. Discussion
4.1. Characteristics of Microbial Community in the Sediments of Estuarine–Coastal Margins
4.2. Mechanisms Underlying the Microbial Assembly in the Sediments of Estuarine–Coastal Margins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bianchi, T.S.; Cui, X.; Blair, N.E.; Burdige, D.J.; Eglinton, T.I.; Galy, V. Centers of organic carbon burial and oxidation at the land-ocean interface. Org. Geochem. 2018, 115, 138–155. [Google Scholar] [CrossRef]
- Yuan, K.; Wang, X.; Lin, L.; Zou, S.; Li, Y.; Yang, Q.; Luan, T. Characterizing the parent and alkyl polycyclic aromatic hydrocarbons in the Pearl River Estuary, Daya Bay and northern South China Sea: Influence of riverine input. Environ. Pollut. 2015, 199, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Abril, G.; Borges, A.V. Carbon dioxide and methane emissions from estuaries. In Greenhouse Gas Emissions—Fluxes and Processes; Tremblay, A., Varfalvy, L., Roehm, C., Garneau, M., Eds.; Environmental Science; Springer: Berlin/Heidelberg, Germany, 2005; pp. 187–207. [Google Scholar]
- Herlemann, D.P.R.; Lundin, D.; Andersson, A.F.; Labrenz, M.; Jürgens, K. Phylogenetic signals of salinity and season in bacterial community composition across the salinity gradient of the Baltic Sea. Front. Microbiol. 2016, 7, 1883. [Google Scholar] [CrossRef] [PubMed]
- Filker, S.; Kühner, S.; Heckwolf, M.; Dierking, J.; Stoeck, T. A fundamental difference between macrobiota and microbial eukaryotes: Protistan plankton has a species maximum in the freshwater-marine transition zone of the Baltic Sea. Environ. Microbiol. 2019, 21, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhan, L.; Chen, L.; Zhang, J.; Wu, M.; Liu, J. Spatial and temporal patterns of methane and its influencing factors in the Jiulong River estuary, southeastern China. Mar. Chem. 2021, 228, 103909. [Google Scholar] [CrossRef]
- An, S.; Joye, S.B. Enhancement of coupled nitrification-denitrification by benthic photosynthesis in shallow estuarine sediments. Limnol. Oceanogr. 2001, 46, 62–74. [Google Scholar] [CrossRef]
- Andersson, A.; Brugel, S.; Paczkowska, J.; Rowe, O.F.; Figueroa, D.; Kratzer, S.; Legrand, C. Influence of allochthonous dissolved organic matter on pelagic basal production in a northerly estuary. Estuar. Coast. Shelf Sci. 2018, 204, 225–235. [Google Scholar] [CrossRef]
- Eyre, B.D.; Ferguson, A.J.P.; Webb, A.; Maher, D.; Oakes, J.M. Metabolism of different benthic habitats and their contribution to the carbon budget of a shallow oligotrophic sub-tropical coastal system (southern Moreton Bay, Australia). Biogeochemistry 2011, 102, 87–110. [Google Scholar] [CrossRef]
- Herbert, R.A. Nitrogen cycling in coastal marine ecosystems. FEMS Microbiol. Rev. 1999, 23, 563–590. [Google Scholar] [CrossRef]
- Wang, K.; Razzano, M.; Mou, X. Cyanobacterial blooms alter the relative importance of neutral and selective processes in assembling freshwater bacterioplankton community. Sci. Total Environ. 2020, 706, 135724. [Google Scholar] [CrossRef]
- Gardolinski, P.C.F.C.; Worsfold, P.J.; McKelvie, I.D. Seawater induced release and transformation of organic and inorganic phosphorus from river sediments. Water Res. 2004, 38, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.J.; Lazar, C.S.; Teske, A.P.; Dick, G.J. Genomic resolution of linkages in carbon, nitrogen, and sulfur cycling among widespread estuary sediment bacteria. Microbiome 2015, 3, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouskill, N.J.; Eveillard, D.; Chien, D.; Jayakumar, A.; Ward, B.B. Environmental factors determining ammonia-oxidizing organism distribution and diversity in marine environments. Environ. Microbiol. 2012, 14, 714–729. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.H.; Eisen, J.A.; Penn, K.; Allison, S.D.; Horner-Devine, M.C. Drivers of bacterial beta-diversity depend on spatial scale. Proc. Natl. Acad. Sci. USA 2011, 108, 7850–7854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunato, C.S.; Crump, B.C. Microbial gene abundance and expression patterns across a river to ocean salinity gradient. PLoS ONE 2015, 10, e0140578. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, X.; Wang, M.; Qiao, Y.; Zheng, Y.; Zhang, X. Bacterial and archaeal communities in sediments of the North Chinese Marginal Seas. Microb. Ecol. 2015, 70, 105–117. [Google Scholar] [CrossRef]
- Amend, A.S.; Oliver, T.A.; Amaral-Zettler, L.A.; Boetius, A.; Fuhrman, J.A.; Horner-Devine, M.C.; Huse, S.M.; Welch, D.B.M.; Martiny, A.C.; Ramette, A.; et al. Macroecological patterns of marine bacteria on a global scale. J. Biogeogr. 2013, 40, 800–811. [Google Scholar] [CrossRef]
- Liu, X.; Pan, J.; Liu, Y.; Li, M.; Gu, J.D. Diversity and distribution of Archaea in global estuarine ecosystems. Sci. Total Environ. 2018, 637, 349–358. [Google Scholar] [CrossRef]
- Webster, G.; O’Sullivan, L.A.; Meng, Y.; Williams, A.S.; Sass, A.M.; Watkins, A.J.; Parkes, R.J.; Weightman, A.J. Archaeal community diversity and abundance changes along a natural salinity gradient in estuarine sediments. FEMS Microbiol. Ecol. 2015, 91, 1–18. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, S.; Liu, X.; Yao, P.; Ge, T.; Zhang, X. Spatiotemporal dynamics of the archaeal community in coastal sediments: Assembly process and co-occurrence relationship. ISME J. 2020, 14, 1463–1478. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Liu, N.; Yang, M.; Wang, L.; Liang, X.; Liu, C. Co-occurrence of planktonic bacteria and archaea affects their biogeographic patterns in China’s coastal wetlands. Environ. Microbiome 2021, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Zhang, W.; Yang, J.; Lin, Y.; Yu, Z.; Lin, S. Biogeographic patterns of abundant and rare bacterioplankton in three subtropical bays resulting from selective and neutral processes. ISME J. 2018, 12, 2198–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Hu, H.; Yan, H.; Hou, D.; Wang, Y.; Dong, P.; Zhang, D. Archaeal biogeography and interactions with microbial community across complex subtropical coastal waters. Mol. Ecol. 2019, 28, 3101–3118. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Lin, C.; Yan, Y.; Hu, G.; Huang, H.; Wang, X. Distribution and provenance implication of rare earth elements and Sr-Nd isotopes in surface sediments of Jiulong River, Southeast China. J. Soil Sediment 2019, 19, 1499–1510. [Google Scholar] [CrossRef]
- Chen, C.F.; Ju, Y.R.; Lim, Y.C.; Hsu, N.H.; Lu, K.T.; Hsieh, S.L.; Dong, C.D.; Chen, C.W. Microplastics and their affiliated PAHs in the sea surface connected to the southwest coast of Taiwan. Chemosphere 2020, 254, 126818. [Google Scholar] [CrossRef]
- Lv, M.; Luan, X.; Liao, C.; Wang, D.; Liu, D.; Zhang, G.; Jiang, G.; Chen, L. Human impacts on polycyclic aromatic hydrocarbon distribution in Chinese intertidal zones. Nat. Sustain. 2020, 3, 878–884. [Google Scholar] [CrossRef]
- Zeng, X.; Xu, L.; Hu, Q.; Liu, Y.; Hu, J.; Liao, W.; Yu, Z. Occurrence and distribution of organophosphorus flame retardants/plasticizers in coastal sediments from the Taiwan Strait in China. Mar. Pollut. Bull. 2020, 151, 110843. [Google Scholar] [CrossRef]
- Xu, H.; Pittock, J. Limiting the effects of hydropower dams on freshwater biodiversity: Options on the Lancang River, China. Mar. Freshwater Res. 2018, 70, 169–194. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.; Si, D.; Chen, Q. Evolution of microbial community along with increasing solid concentration during highsolids anaerobic digestion of sewage sludge. Bioresour. Technol. 2016, 216, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Kemnitz, D.; Kolb, S.; Conrad, R. Phenotypic characterization of Rice Cluster III archaea without prior isolation by applying quantitative polymerase chain reaction to an enrichment culture. Environ. Microbiol. 2005, 7, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; Dewaal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microb. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, S.; Yang, Y.; Xu, Y.; Zhang, J.; Lu, Y. Balance between community assembly processes mediates species coexistence in agricultural soil microbiomes across eastern China. ISME J. 2020, 14, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Xun, W.; Li, W.; Xiong, W.; Ren, Y.; Liu, Y.; Miao, Y.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Diversity-triggered deterministic bacterial assembly constrains community functions. Nat. Commun. 2019, 10, 3833. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Pi, Y.; Song, Y.; Jiang, Y.; Zhou, L.; Liu, W.; Zhu, G. Hotspot of dissimilatory nitrate reduction to ammonium (DNRA) process in freshwater sediments of riparian zones. Water Res. 2020, 173, 115539. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, Y.; Hu, Y.; Cai, J.; Liu, X.; Bai, C.; Tang, X.; Zhang, Y.; Jang, K.S.; Spencer, R.G.; et al. Microbial production and consumption of dissolved organic matter in glacial ecosystems on the Tibetan plateau. Water Res. 2019, 160, 18–28. [Google Scholar] [CrossRef]
- Ghosh, A.; Dey, N.; Bera, A.; Tiwari, A.; Sathyaniranjan, K.B.; Chakrabarti, K.; Chattopadhyay, D. Culture independent molecular analysis of bacterial communities in the mangrove sediment of Sundarban, India. Aquat. Biosyst. 2010, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.M.; Abell, G.; Philippot, L.; Prosser, J.; et al. Microbes as engines of ecosystem function: When does community structure enhance predictions of ecosystem processes. Front. Microbiol. 2016, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Nayfach, S.; Roux, S.; Seshadri, R.; Udwary, D.; Varghese, N.; Schulz, F.; Wu, D.; Paez-Espino, D.; Chen, I.M.; Huntemann, M.; et al. A genomic catalog of Earth’s microbiomes. Nat. Biotechnol. 2021, 39, 499–509. [Google Scholar] [CrossRef]
- Bange, H.W. Nitrous oxide and methane in European coastal waters. Estuar. Coast. Shelf Sci. 2006, 70, 361–374. [Google Scholar] [CrossRef]
- Bernhard, A.E.; Bollmann, A. Estuarine nitrifiers: New players, patterns and processes. Estuar. Coast. Shelf Sci. 2010, 88, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chidthaisong, A.; Conrad, R. Turnover of glucose and acetate coupled to reduction of nitrate, ferric iron and sulfate and to methanogenesis in anoxic rice field soil. FEMS Microbiol. Ecol. 2020, 31, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ding, W.; Jia, Z.; Cai, Z. The impact of dissolved organic carbon on the spatial variability of methanogenic archaea communities in natural wetland ecosystems across China. Appl. Microbiol. Biot. 2012, 96, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Parkes, R.J.; Brock, F.; Banning, N.; Hornibrook, E.R.; Roussel, E.G.; Weightman, A.J.; Fry, J.C. Changes in methanogenic substrate utilization and communities with depth in a salt-marsh, creek sediment in southern England. Estuar. Coast. Shelf Sci. 2012, 96, 170–178. [Google Scholar] [CrossRef]
- Middelburg, J.J.; Nieuwenhuize, J.; Iversen, N.; Høgh, N.; De Wilde, H.; Helder, W.; Seifert, R.; Christof, O. Methane distribution in European tidal estuaries. Biogeochemistry 2002, 59, 95–119. [Google Scholar] [CrossRef]
- Martens, C.S.; Berner, R.A. Methane production in the interstitial waters of sulfate-depleted marine sediments. Science 1974, 185, 1167–1169. [Google Scholar] [CrossRef]
- Tee, H.S.; Waite, D.; Lear, G.; Handley, K.M. Microbial river-to-sea continuum: Gradients in benthic and planktonic diversity, osmoregulation and nutrient cycling. Microbiome 2021, 9, 1–8. [Google Scholar] [CrossRef]
- Zou, D.; Wan, R.; Han, L.; Xu, M.; Li, M. Genomic characteristics of a novel species of ammonia-oxidizing archaea from the Jiulong River Estuary. Appl. Environ. Microb. 2020, 86, e00736-20. [Google Scholar] [CrossRef]
- Dupont, C.L.; Larsson, J.; Yooseph, S.; Ininbergs, K.; Goll, J.; Asplund-Samuelsson, J.; McCrow, J.P.; Celepli, N.; Allen, L.Z.; Ekman, M.; et al. Functional Tradeoffs Underpin Salinity-Driven Divergence in Microbial Community Composition. PLoS ONE 2014, 9, e89549. [Google Scholar] [CrossRef]
- Eiler, A.; Zaremba-Niedzwiedzka, K.; Martínez-García, M.; McMahon, K.D.; Stepanauskas, R.; Andersson, S.G.; Bertilsson, S. Productivity and salinity structuring of the microplankton revealed by comparative freshwater metagenomics. Environ. Microbiol. 2014, 16, 2682–2698. [Google Scholar] [CrossRef] [Green Version]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Liebermeister, W.; Milo, R. Glycolytic strategy as a tradeoff between energy yield and protein cost. Proc. Natl. Acad. Sci. USA 2013, 110, 10039–10044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrichs, M.E.; Mori, C.; Dlugosch, L. Complex interactions between aquatic organisms and their chemical environment elucidated from different perspectives. In Youmares 9-The Oceans: Our Research, Our Future; Jungblut, S., Liebich, V., Bode-Dalby, M., Eds.; Springer: Champaign, IL, USA, 2020; pp. 279–297. [Google Scholar]
- Cloern, J.E.; Jassby, A.D.; Schraga, T.S.; Nejad, E.; Martin, C. Ecosystem variability along the estuarine salinity gradient: Examples from long-term study of San Francisco Bay. Limnol. Oceanogr. 2017, 62, 272–291. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Knight, R. Global patterns in bacterial diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, A.N.; Wang, J.; Liu, S.; Jiang, X.; Dang, C.; Ma, T.; Liu, S.; Chen, Q.; Xie, S.; et al. Integrated biogeography of planktonic and sedimentary bacterial communities in the Yangtze River. Microbiome 2018, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Wang, Y.; Warren, A.; Huang, B.; Sun, P. Diversity distribution and assembly mechanisms of planktonic and benthic microeukaryote communities in intertidal zones of Southeast Fujian, China. Front. Microbiol. 2019, 10, 2640. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Mansfeldt, C.; Achermann, S.; Men, Y.; Walser, J.C.; Villez, K.; Joss, A.; Johnson, D.R.; Fenner, K. Microbial residence time is a controlling parameter of the taxonomic composition and functional profile of microbial communities. ISME J. 2019, 13, 1589–1601. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshwater Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Ma, B.; Wang, H.; Dsouza, M.; Lou, J.; He, Y.; Dai, Z.; Brookes, P.C.; Xu, J.; Gilbert, J.A. Geographic patterns of co-occurrence network topological features for soil microbiota at continental scale in eastern China. ISME J. 2016, 10, 1891–1901. [Google Scholar] [CrossRef]
- Logares, R.; Lindström, E.S.; Langenheder, S.; Logue, J.B.; Paterson, H.; Laybourn-Parry, J.; Rengefors, K.; Tranvik, L.; Bertilsson, S. Biogeography of bacterial communities exposed to progressive long-term environmental change. ISME J. 2013, 7, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Ranjith, E.; Lidita, K. Influence of salinity stress on bacterial community composition and beta-glucosidase activity in a tropical estuary: Elucidation through microcosm experiments. Mar. Environ. Res. 2020, 159, 104997. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Classifi-Cation | Depth (m) | Temp (°C) | Sal (‰) | pH | Chla (μg/L) | DO (mg/L) | BGA-PE (μg/L) | TDS (mg/L) | NH3-N (mg/kg) | TN (%) | TOC (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jiulong River Estuary | L | 8.93 ± 1.58 c | 17.7 ± 0.319 b | 5.63 ± 3.02 d | 7.72 ± 0.0572 c | 3.79 ± 0.541 a | 6.45 ± 0.379 c | 9 ± 2.03 a | 6404 ± 3225 d | 108 ± 9.6 a | 0.133 ± 0.0126 a | 1.19 ± 0.0755 a |
| M | 6.95 ± 1.49 c | 17.2 ± 0.193 b | 14.6 ± 1.76 c | 7.8 ± 0.0921 c | 2.85 ± 0.273 b | 7.47 ± 0.25 b | 7.47 ± 0.669 b | 15,575 ± 1715 cd | 52.5 ± 37.3 b | 0.0733 ± 0.0484 b | 0.855 ± 0.383 ab | |
| H | 9.3 ± 1.46 c | 16.9 ± 0.164 b | 24.1 ± 2.51 b | 8.03 ± 0.0265 b | 2.37 ± 1.06 bc | 8.08 ± 0.0606 a | 5.74 ± 0.256 c | 24,639 ± 2332 bc | 32 ± 21.1 bc | 0.083 ± 0.0208 b | 0.88 ± 0.0321 b | |
| Taiwan Strait | X | 26.5 ± 15.21 bc | 21.42 ± 0.68 a | 31.11 ± 1.1 a | NA | NA | NA | NA | NA | 21.62 ± 7.99 c | 0.06 ± 0.05 b | 0.51 ± 0.32 b |
| Q | 37.42 ± 15.02 ab | 22.4 ± 1.54 a | 32.38 ± 1.53 a | 8.13 ± 0.05 a | 1.67 ± 0.32 c | 7.36 ± 0.11 b | 4.21 ± 0.66 d | 31,274.5 ± 1050.75 ab | 33.02 ± 14.05 bc | 0.06 ± 0.04 b | 0.47 ± 0.31 b | |
| P | 45.6 ± 19.76 ab | 21.43 ± 1.66 a | 31.95 ± 1.9 a | NA | NA | NA | NA | NA | 20.28 ± 5.98 c | 0.08 ± 0.03 ab | 0.6 ± 0.25 b | |
| E | 52.53 ± 18.23 a | 22.31 ± 1.2 a | 32.16 ± 1.88 a | 8.13 ± 0.04 a | 1.54 ± 0.3 c | 7.25 ± 0.34 b | 4.28 ± 0.59 d | 38,532.25 ± 14,846.99 a | 19.9 ± 4.17 c | 0.11 ± 0.03 ab | 0.76 ± 0.16 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, Y.; Tang, Y.; Cai, L.; Yang, Z.; Chen, X.; Ouyang, Y.; Dai, J.; Yang, M. Co-Occurrence Relationship and Stochastic Processes Affect Sedimentary Archaeal and Bacterial Community Assembly in Estuarine–Coastal Margins. Microorganisms 2022, 10, 1339. https://doi.org/10.3390/microorganisms10071339
Yue Y, Tang Y, Cai L, Yang Z, Chen X, Ouyang Y, Dai J, Yang M. Co-Occurrence Relationship and Stochastic Processes Affect Sedimentary Archaeal and Bacterial Community Assembly in Estuarine–Coastal Margins. Microorganisms. 2022; 10(7):1339. https://doi.org/10.3390/microorganisms10071339
Chicago/Turabian StyleYue, Yihong, Yi Tang, Ling Cai, Zhihong Yang, Xueping Chen, Yurong Ouyang, Juanjuan Dai, and Ming Yang. 2022. "Co-Occurrence Relationship and Stochastic Processes Affect Sedimentary Archaeal and Bacterial Community Assembly in Estuarine–Coastal Margins" Microorganisms 10, no. 7: 1339. https://doi.org/10.3390/microorganisms10071339