
Enhancing Red Yeast Biomass Yield and Lipid Biosynthesis by Using Waste Nitrogen Source by Glucose Fed-Batch at Low Temperature

 , ,
, ,  ,
,
Abstract
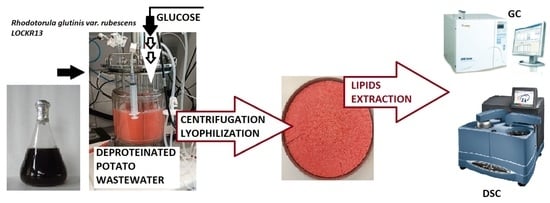
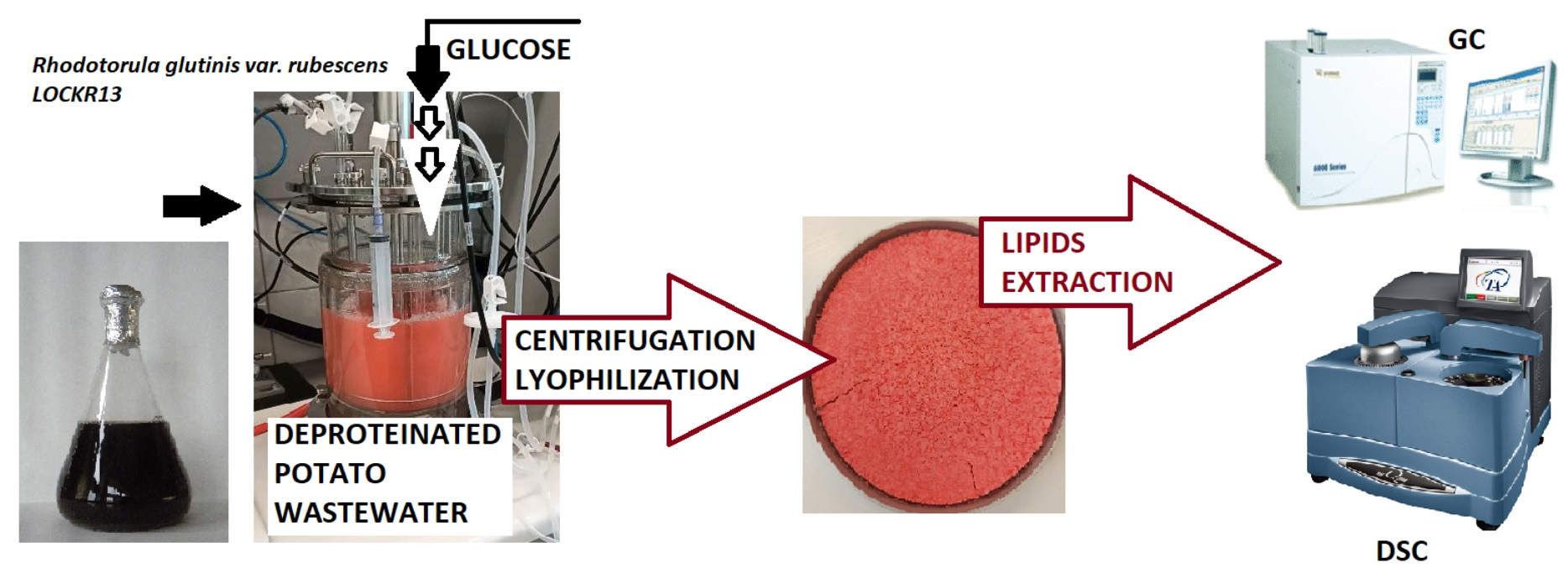
:
1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Industrial Waste Used as Component of the Medium
2.3. Inoculation, Batch Culture, and Fed-Batch Culture Conditions
2.4. DPW Chemical Characteristics
2.5. Biomass Yield and Lipids Content
2.6. Calculation of Bioprocess Efficiency Parameters
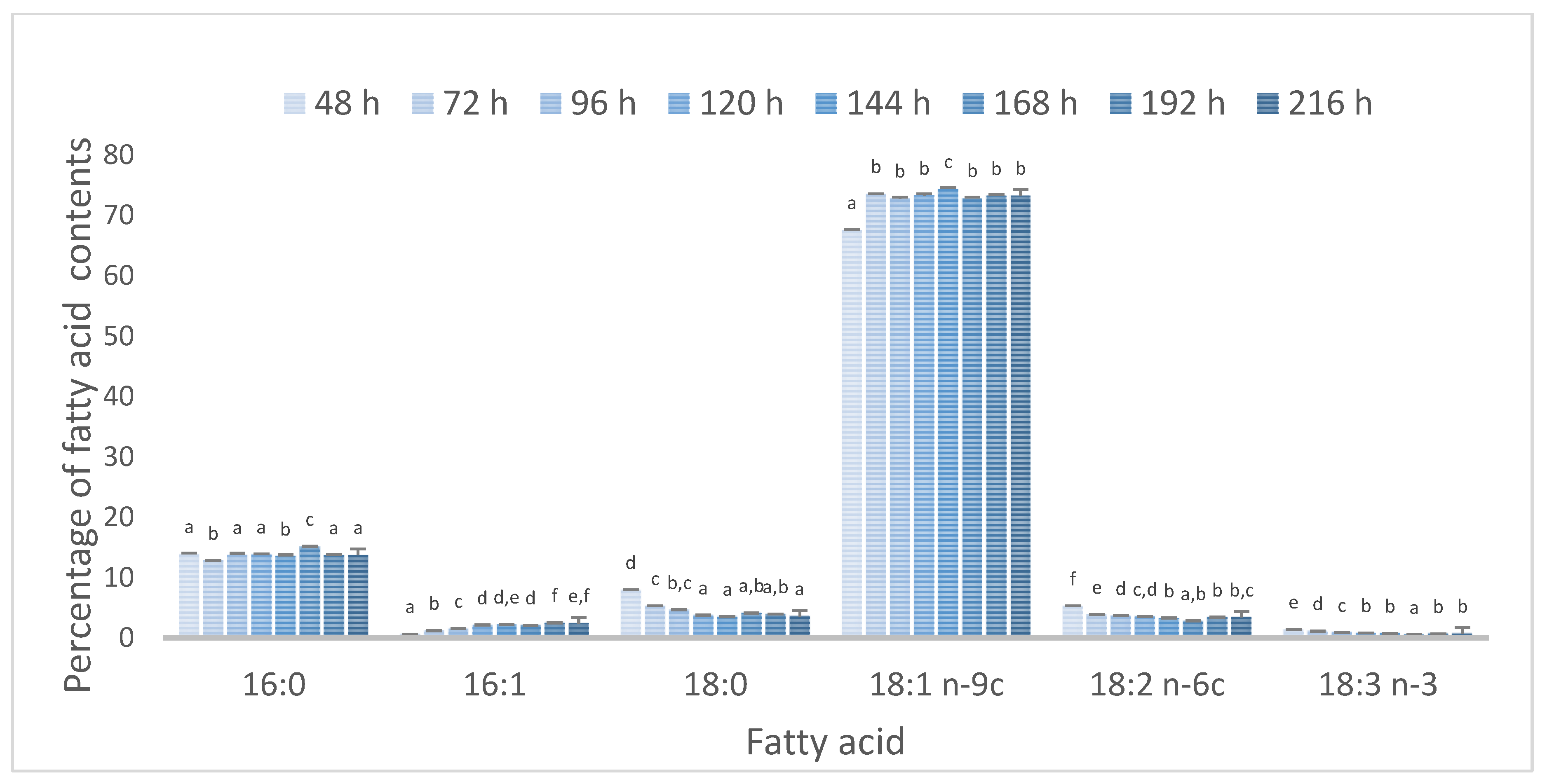
2.7. Lipids Analysis-Relative Composition of Fatty Acids
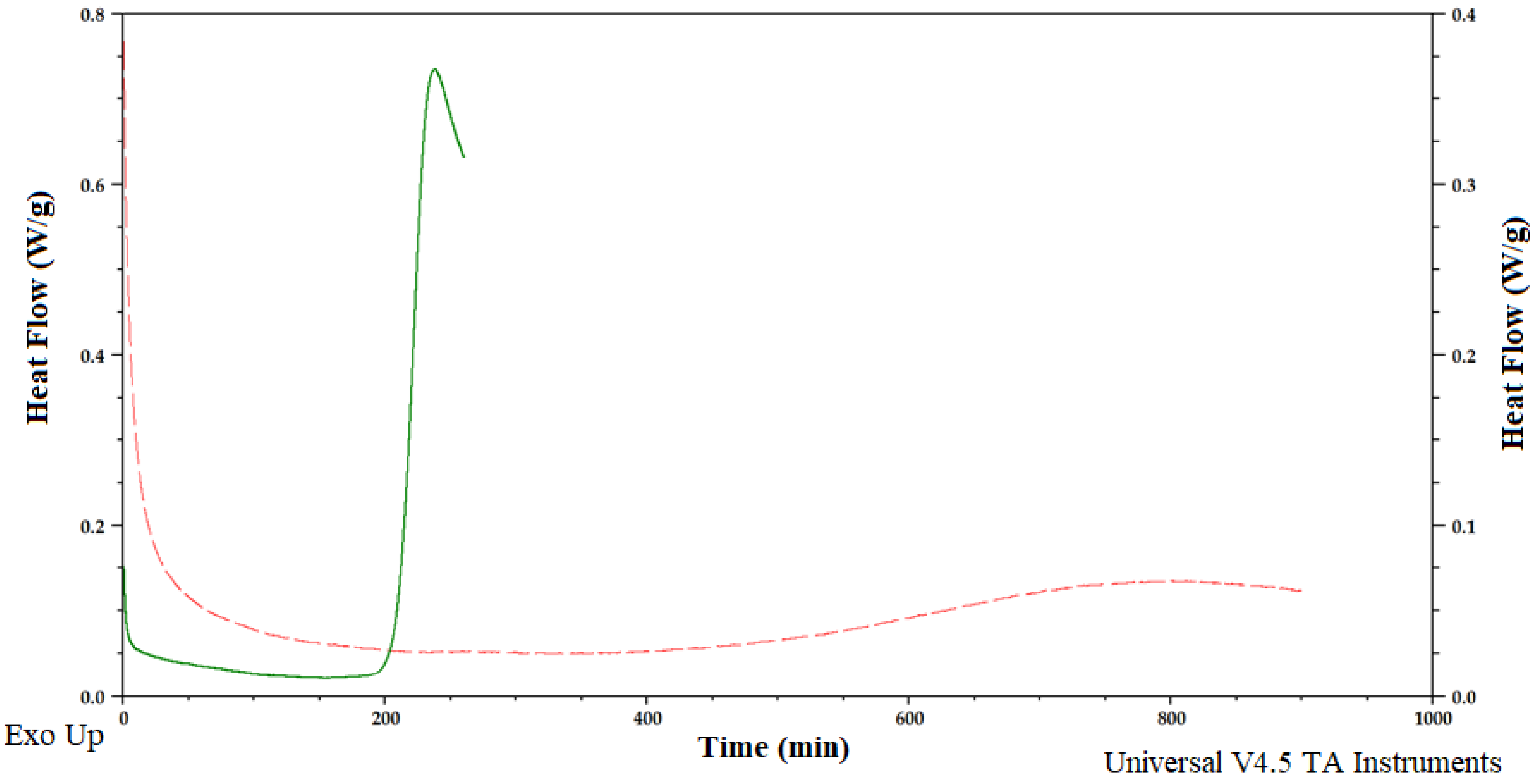
2.8. Thermal Analysis of Yeast Lipids
2.9. Statistical Analysis
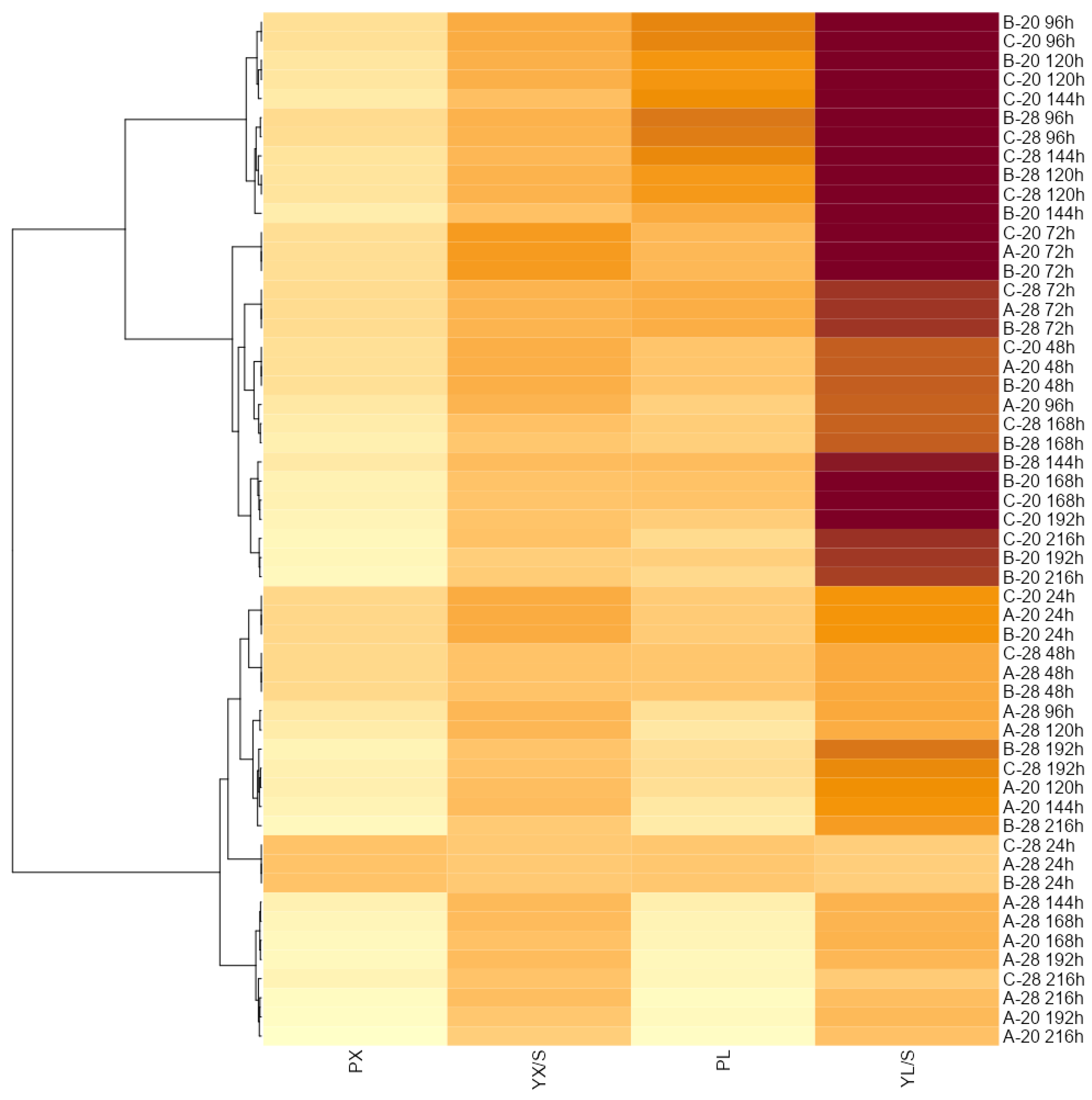
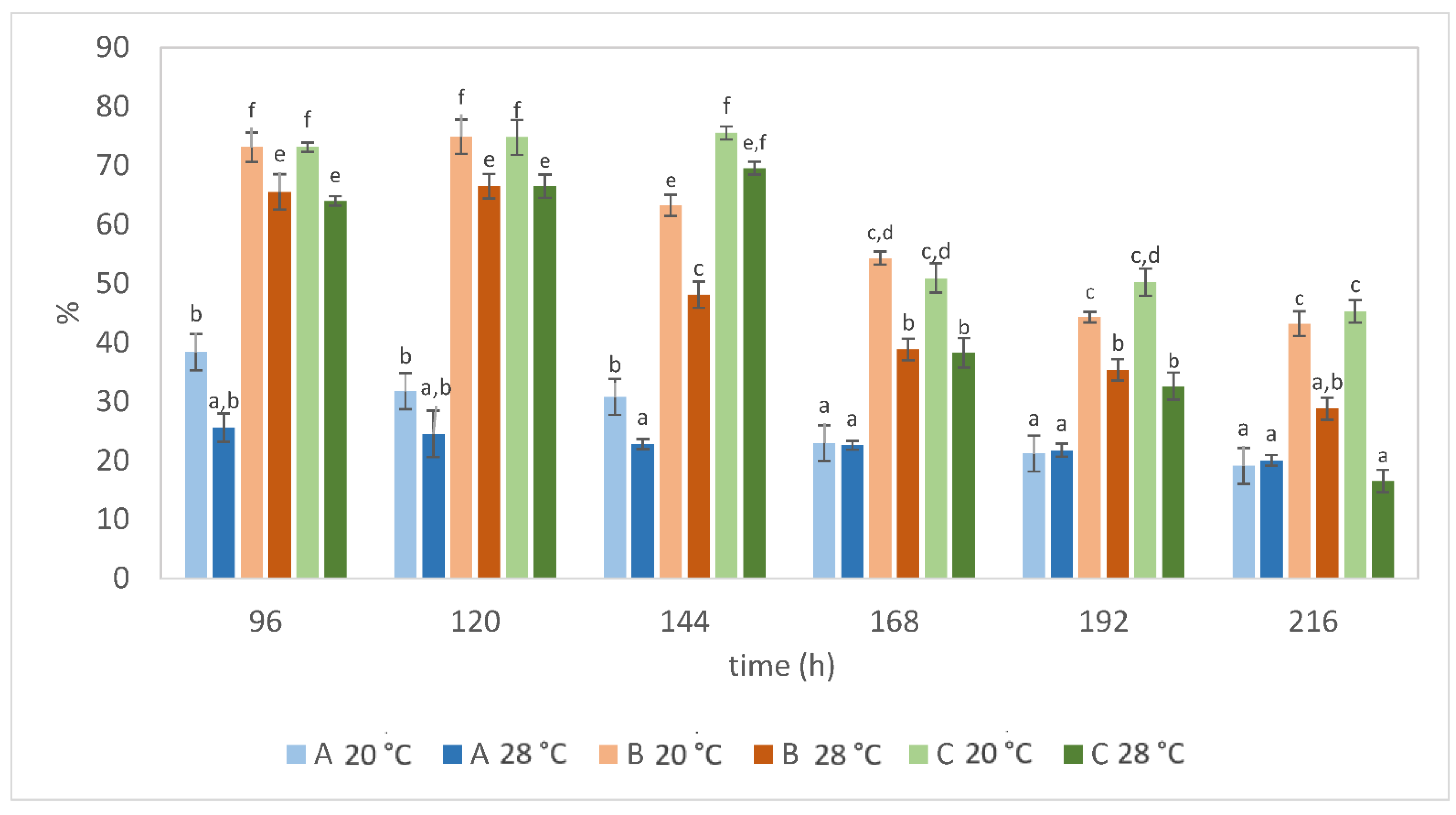
3. Results and Discussion
3.1. The Characteristic of the Potato Wastewater Used as a Medium
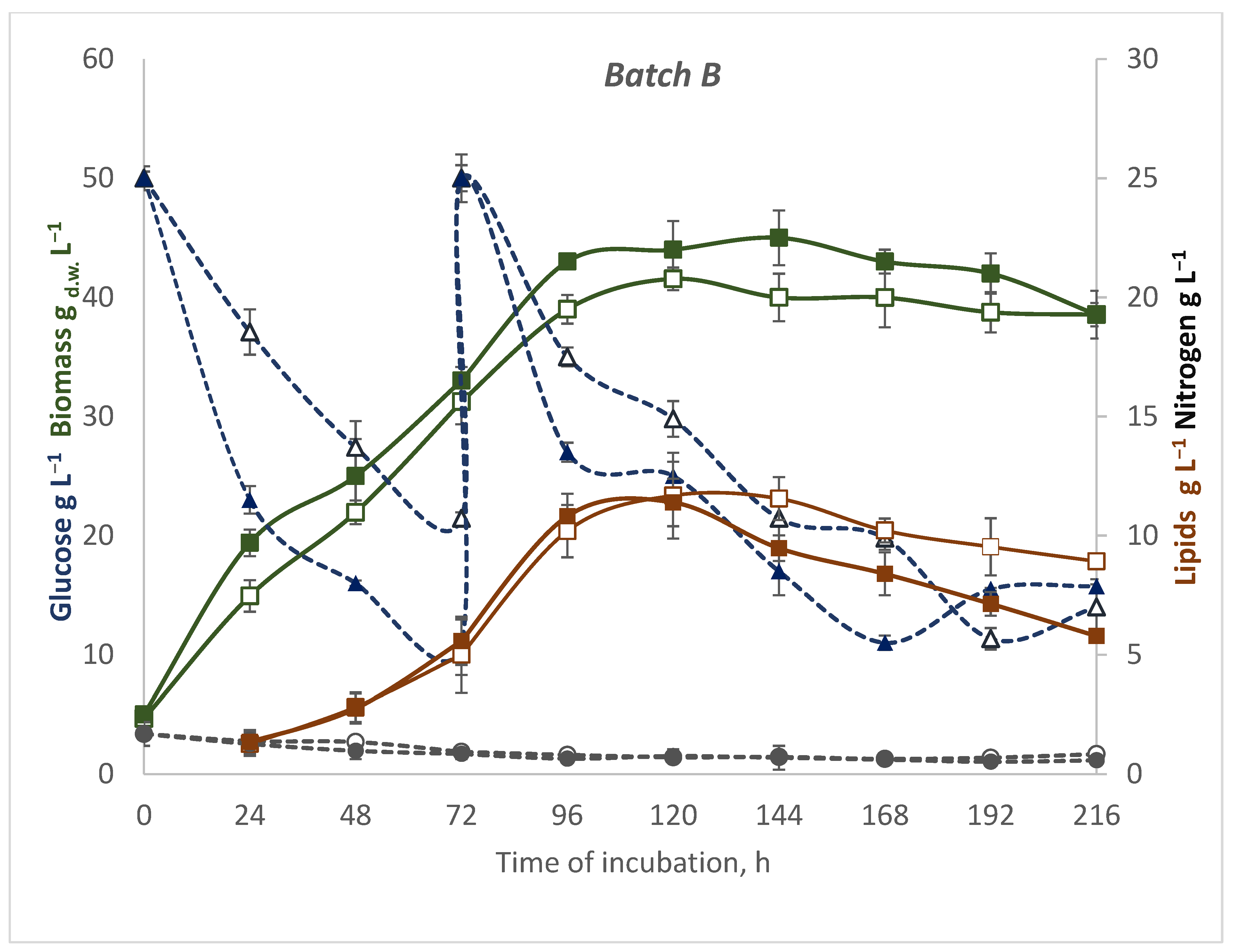
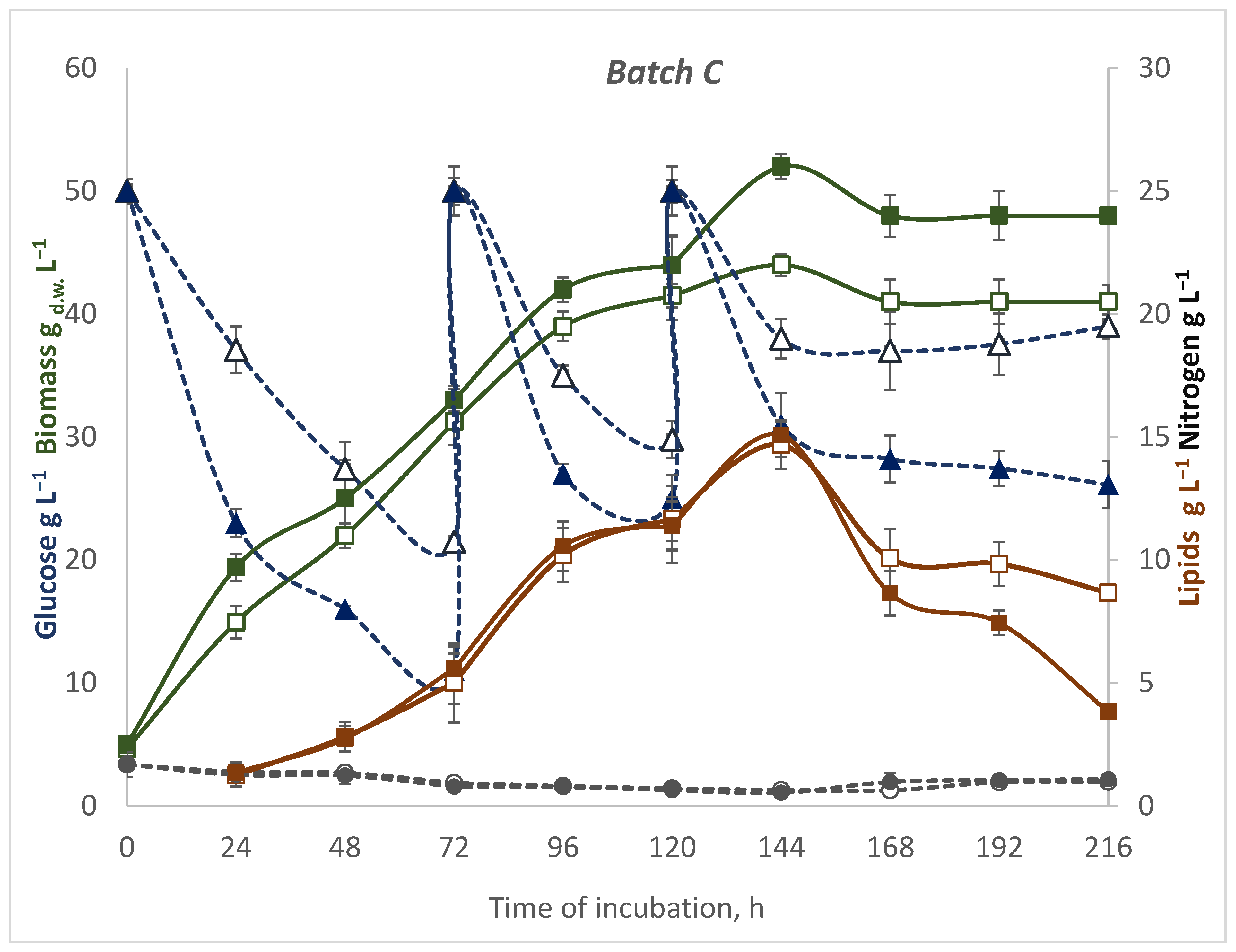
3.2. Effect of Batch and Fed-Batch Strategy on Yeast Growth and Lipid Accumulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula Glutinis—Potential Source of Lipids, Carotenoids, and Enzymes for Use in Industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily Yeasts as Oleaginous Cell Factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipids of Oleaginous Yeasts. Part II: Technology and Potential Applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Němcová, A.; Szotkowski, M.; Samek, O.; Cagáňová, L.; Sipiczki, M.; Márová, I. Use of Waste Substrates for the Lipid Production by Yeasts of the Genus Metschnikowia—Screening Study. Microorganisms 2021, 9, 2295. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and Torularhodin: “New” Fungal Carotenoids for Industry? Microb. Cell Fact. 2018, 17, 49. [Google Scholar] [CrossRef] [Green Version]
- Kot, A.M.; Błażejak, S.; Kieliszek, M.; Gientka, I.; Bryś, J. Simultaneous Production of Lipids and Carotenoids by the Red Yeast Rhodotorula from Waste Glycerol Fraction and Potato Wastewater. Appl. Biochem. Biotechnol. 2019, 189, 589–607. [Google Scholar] [CrossRef]
- Byrtusová, D.; Szotkowski, M.; Kurowska, K.; Shapaval, V.; Márová, I. Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites. Microorganisms 2021, 9, 1280. [Google Scholar] [CrossRef] [PubMed]
- Gientka, I.; Aleksandrzak-Piekarczyk, T.; Bzducha-Wróbel, A.; Synowiec, A.; Błażejak, S. Deproteinated Potato Wastewater as a Sustainable Nitrogen Source in Trichosporon Domesticum Yeast Lipids Biosynthesis—A Concept of Valorization of Wastewater from Starch Industry. Potato Res. 2019, 62, 221–237. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Yang, Z.; Hu, B.; Ji, X.; Wei, Y.; Lin, L.; Zhang, Q. Correlation of Polyunsaturated Fatty Acids with the Cold Adaptation of Rhodotorula glutinis. Yeast 2015, 32, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.-J.; Lee, J.K. Effect of Changes in the Composition of Cellular Fatty Acids on Membrane Fluidity of Rhodobacter Ssphaeroides. J. Microbiol. Biotechnol. 2015, 25, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Chintalapati, S.; Kiran, M.D.; Shivaji, S. Role of membrane lipid fatty acids in cold adaptation. Cell. Mol. Biol. 2004, 50, 631–642. [Google Scholar] [PubMed]
- Ratledge, C.; Wynn, J.P. The Biochemistry and Molecular Biology of Lipid Accumulation in Oleaginous Microorganisms. In Advances in Applied Microbiology; Laskin, A.I., Bennett, J.W., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2002; Volume 51, pp. 1–52. [Google Scholar] [CrossRef]
- Buzzini, P. Batch and fed-batch carotenoid production by Rhodotorula glutinis-Debaryomyces castellii co-cultures in corn syrup. J. Appl. Microbiol. 2001, 90, 843–847. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liu, G.-L.; Chi, Z.; Chi, Z.-M. Single cell oil production from hydrolysate of cassava starch by marine-derived yeast Rhodotorula mucilaginosa TJY15a. Biomass Bioenergy 2010, 34, 101–107. [Google Scholar] [CrossRef]
- Vamvakaki, A.-N.; Kandarakis, I.; Kaminarides, S.; Komaitis, M.; Papanikolaou, S. Cheese whey as a renewable substrate for microbial lipid and biomass production by Zygomycetes. Eng. Life Sci. 2010, 10, 348–360. [Google Scholar] [CrossRef]
- Xue, F.; Miao, J.; Zhang, X.; Luo, H.; Tan, T. Studies on lipid production by Rhodotorula glutinis fermentation using monosodium glutamate wastewater as culture medium. Bioresour. Technol. 2008, 99, 5923–5927. [Google Scholar] [CrossRef] [PubMed]
- Tchakouteu, S.S.; Kopsahelis, N.; Chatzifragkou, A.; Kalantzi, O.; Stoforos, N.G.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Rhodosporidium toruloides cultivated in NaCl-enriched glucose-based media: Adaptation dynamics and lipid production. Eng. Life Sci. 2017, 17, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Aggelis, G. Modeling Lipid Accumulation and Degradation in Yarrowia lipolytica Cultivated on Industrial Fats. Curr. Microbiol. 2003, 46, 0398–0402. [Google Scholar] [CrossRef] [PubMed]
- Slaný, O.; Klempová, T.; Shapaval, V.; Zimmermann, B.; Kohler, A.; Čertík, M. Animal Fat as a Substrate for Production of n-6 Fatty Acids by Fungal Solid-State Fermentation. Microorganisms 2021, 9, 170. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I.K. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Source Agricultural Produce from Cooperatives. Available online: https://www.selinawamucii.com/ (accessed on 20 May 2020).
- Karamerou, E.E.; Parsons, S.; McManus, M.C.; Chuck, C.J. Using techno-economic modelling to determine the minimum cost possible for a microbial palm oil substitute. Biotechnol. Biofuels 2021, 14, 57. [Google Scholar] [CrossRef]
- Gientka, I.; Duda, M.; Bzducha-Wróbel, A.; Błażejak, S. Deproteinated Potato Wastewater as a Low-Cost Nitrogen Substrate for Very High Yeast Biomass Quantities: Starting Point for Scaled-up Applications. Eur. Food Res. Technol. 2019, 245, 919–928. [Google Scholar] [CrossRef] [Green Version]
- Minihane, B.J.; Brown, D.E. Fed-Batch Culture Technology. Biotechnol. Adv. 1986, 4, 207–218. [Google Scholar] [CrossRef]
- Poontawee, R.; Limtong, S. Feeding Strategies of Two-Stage Fed-Batch Cultivation Processes for Microbial Lipid Production from Sugarcane Top Hydrolysate and Crude Glycerol by the Oleaginous Red Yeast Rhodosporidiobolus Fluvialis. Microorganisms 2020, 8, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiebe, M.G.; Koivuranta, K.; Penttilä, M.; Ruohonen, L. Lipid Production in Batch and Fed-Batch Cultures of Rhodosporidium toruloidesfrom 5 and 6 Carbon Carbohydrates. BMC Biotechnol. 2012, 12, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendicho, C.; Lavilla, I. Water Analysis Sewage. In Encyclopedia of Analytical Science, 3rd ed.; Worsfold, P., Poole, C., Townshend, A., Miró, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 371–381. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Pobiega, K.; Błażejak, S.; Kieliszek, M. The scale-up cultivation of Candida utilis in waste potato juice water with glycerol affects biomass and β(1,3)/(1,6)-glucan characteristic and yield. Appl. Microbiol. Biotechnol. 2018, 102, 9131–9145. [Google Scholar] [CrossRef] [Green Version]
- Assmann, C.; Scott, A.; Biller, D. Online total organic carbon (TOC) monitoring for water and wastewater treatment plants processes and operations optimization. Drink. Water Eng. Sci. 2017, 10, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kirk, P.L. Kjeldahl Method for Total Nitrogen. Anal. Chem. 1950, 22, 354–358. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- N’Guessan, F.K.; Coulibaly, H.W.; Alloue-Boraud, M.W.A.; Cot, M.; Djè, K.M. Production of freeze-dried yeast culture for the brewing of traditional sorghum beer, tchapalo. Food Sci. Nutr. 2015, 4, 34–41. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J.A. Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Zhang, G.C.; French, W.T.; Hernandez, R.; Alley, E.; Paraschivescu, M. Effects of Furfural and Acetic Acid on Growth and Lipid Production from Glucose and Xylose by Rhodotorula Glutinis. Biomass Bioenergy 2011, 35, 734–740. [Google Scholar] [CrossRef]
- Zhang, G.; French, W.T.; Hernandez, R.; Alley, E.; Paraschivescu, M. Comparison of simple and rapid cell wall disruption methods for improving lipid extraction from yeast cells. J. Microbiol. Methods 2019, 176, 105999. [Google Scholar] [CrossRef]
- Polish Norm: PN-EN ISO: 5509:2001: Oil and Vegetable and Animal Fats. In Preparation of Methyl Ester of Fatty Acids; Polish Committee for Standardization: Warsaw, Poland, 2001.
- Górska, A.; Brzezińska, R.; Wirkowska-Wojdyła, M.; Bryś, J.; Domian, E.; Ostrowska-Ligęza, E. Application of Thermal Methods to Analyze the Properties of Coffee Silverskin and Oil Extracted from the Studied Roasting By-Product. Appl. Sci. 2020, 10, 8790. [Google Scholar] [CrossRef]
- Ostrowska-Ligęza, E.; Dolatowska-Żebrowska, K.; Wirkowska-Wojdyła, M.; Bryś, J.; Górska, A. Comparison of Thermal Characteristics and Fatty Acids Composition in Raw and Roasted Cocoa Beans from Peru (Criollo) and Ecuador (Forastero). Appl. Sci. 2021, 11, 2698. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Błażejak, S.; Molenda, M.; Reczek, L. Biosynthesis of β(1,3)/(1,6)-glucans of cell wall of the yeast Candida utilis ATCC 9950 strains in the culture media supplemented with deproteinated potato juice water and glycerol. Eur. Food Res. Technol. 2015, 240, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Bryś, J.; Gientka, I.; Bzducha-Wróbel, A.; Maliszewska, M.; Reczek, L. Effect of initial pH of medium with potato wastewater and glycerol on protein, lipid and carotenoid biosynthesis by Rhodotorula glutinis. Electr. J. Biotechnol. 2017, 27, 25–31. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Koczoń, P.; Błażejak, S.; Kozera, J.; Kieliszek, M. Valorization of Deproteinated Potato Juice Water into β-Glucan Preparation of C. utilis Origin: Comparative Study of Preparations Obtained by Two Isolation Methods. Waste Biomass Valor. 2020, 11, 3257–3271. [Google Scholar] [CrossRef] [Green Version]
- Kot, A.; Błażejak, S.; Konarski, K.; Bryś, J.; Gientka, I.; Kurcz, A.; Piwowarek, K. Próba zastosowania glicerolu i ziemniaczanej wody sokowej do produkcji karotenoidów przez drożdże Rhodotorula Gracilis. Zesz. Probl. Postępów Nauk Rol. 2017, 589, 49–57. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Tan, T. Lipid and carotenoid production by Rhodotorula glutinis under irradiation/high-temperature and dark/low-temperature cultivation. Bioresour. Technol. 2014, 157, 149–153. [Google Scholar] [CrossRef]
- Vu, V.H. High-Cell-Density Fed-Batch Culture of Saccharomyces cerevisiae KV-25 Using Molasses and Corn Steep Liquor. J. Microbiol. Biotechnol. 2009, 19, 1603–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.G.; Kwak, M.Y.; Rhee, J.S. High Density Cell Culture of Rhodotorulaglutinis Using Oxygen-Enriched Air. Biotechnol. Lett. 1986, 8, 715–718. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Z.; Bai, F. High-Density Cultivation of Oleaginous Yeast Rhodosporidium Toruloides Y4 in Fed-Batch Culture. Enzym. Microb. Technol. 2007, 41, 312–317. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Kieliszek, M.; Gientka, I.; Piwowarek, K.; Brzezińska, R. Production of lipids and carotenoids by Rhodotorula gracilis ATCC 10788 yeast in a bioreactor using low-cost wastes. Biocatal. Agric. Biotechnol. 2020, 26, 101634. [Google Scholar] [CrossRef]
- Fell, J.W.; Statzell-Tallman, A. “Rhodotorula” Harison. In The Yeast, a Taxonomic Study; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; pp. 800–827. [Google Scholar]
- Schnierda, T.; Bauer, F.F.; Divol, B.; van Resenburg, E.; Görgens, J.F. Optimization of carbon and nitrogen medium components for biomass production using non-Saccharomyces wine yeasts. Lett. Appl. Microbiol. 2014, 58, 478–785. [Google Scholar] [CrossRef]
- Christen, S.; Uwe Sauer, U. Intracellular characterization of aerobic glucose metabolism in seven yeast species by 13C flux analysis and metabolomics. FEMS Yeast Res. 2011, 11, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Elfeky, N.; Elmahmoudy, M.; Bao, Y. Manipulation of Culture Conditions: Tool for Correlating/Improving Lipid and Carotenoid Production by Rhodotorula glutinis. Processes 2020, 8, 140. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Single Cell Oil Production by Yarrowia Lipolytica Growing on an Industrial Derivative of Animal Fat in Batch Cultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312. [Google Scholar] [CrossRef]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of Cultivation Conditions for Biotechnological Production of Lipid by Rhodotorula Kratochvilovae (Syn, Rhodosporidium Kratochvilovae) SY89 for Biodiesel Preparation. 3 Biotech 2017, 7, 145. [Google Scholar] [CrossRef] [Green Version]
- Touchette, D.; Altshuler, I.; Gostinčar, C.; Zalar, P.; Raymond-Bouchard, I.; Zajc, J.; McKay, C.P.; Gunde-Cimerman, N.; Whyte, L.G. Novel Antarctic Yeast Adapts to Cold by Switching Energy Metabolism and Increasing Small RNA Synthesis. ISME J. 2022, 16, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Lee, M.-C.; Puthumana, J.; Park, J.C.; Kang, S.; Han, J.; Shin, K.-H.; Park, H.G.; Om, A.-S.; Lee, J.-S. Effects of Temperature on Growth and Fatty Acid Synthesis in the Cyclopoid Copepod Paracyclopina Nana. Fish. Sci. 2017, 83, 725–734. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; Huijberts, G.N.M.; Eggink, G. High-Cell-Density Cultivation of the Lipid Accumulating Yeast Cryptococcus Curvatus Using Glycerol as a Carbon Source. Appl. Microbiol. Biotechnol. 1996, 45, 575–579. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Wu, D.; Li, J.; Dayal Tyagi, R.; Surampalli, R.Y. Economical lipid production from Trichosporon oleaginosus via dissolved oxygen adjustment and crude glycerol addition. Bioresour. Technol. 2018, 773, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.A.; Rayo, J.; Giralda, J.M. Improving industrial full-scale production of baker’s yeast by optimizing aeration control. J. AOAC Int. 2008, 91, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, S.; Kampisopoulou, E.; Blanchard, F.; Rondags, E.; Gardeli, C.; Koutinas, A.A.; Chevalot, I.; Aggelis, G. Production of secondary metabolites through glycerol fermentation under carbon-excess conditions by the yeasts Yarrowia lipolytica and Rhodosporidium toruloides. Eur. J. Lipid Sci. Technol. 2017, 119, 1600507. [Google Scholar] [CrossRef]
- Magdouli, S.; Brar, S.K.; Blais, J.F. Morphology and rheological behavior ofYarrowia lipolytica: Impact of dissolved oxygen level on cell growth and lipid composition. Proc. Biochem. 2018, 65, 1–10. [Google Scholar] [CrossRef]
- Capusoni, C.; Rodighiero, V.; Cucchetti, D.; Galafassi, S.; Bianchi, D.; Franzosi, G.; Compagno, C. Characterization of lipid accumulation and lipidome analysis in the oleaginous yeasts Rhodosporidium azoricum and Trichosporon oleaginosus. Bioresour. Technol. 2017, 238, 281–289. [Google Scholar] [CrossRef]
- Yen, H.W.; Zhang, Z. Effects of dissolved oxygen level on cell growth and total lipid accumulation in the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2011, 112, 71–74. [Google Scholar] [CrossRef]
- Najjar, A.; Robert, S.; Guérin, C.; Violet-Asther, M.; Carrière, F. Quantitative Study of Lipase Secretion, Extracellular Lipolysis, and Lipid Storage in the Yeast Yarrowia Lipolytica Grown in the Presence of Olive Oil: Analogies with Lipolysis in Humans. Appl. Microbiol. Biotechnol. 2011, 89, 1947–1962. [Google Scholar] [CrossRef]
- Serio, M.; Di Giacinto, L.; Di Loreto, G.; Giansante, L.; Pellegrino, M.; Vito, R.; Perri, E. Chemical and Sensory Characteristics of Italian Virgin Olive Oils from Grossa Di Gerace Cv: Grossa Di Gerace’ Characterization. Eur. J. Lipid Sci. Technol. 2015, 118, 288–298. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Mechanisms and Factors for Edible Oil Oxidation. Compr. Rev. Food Sci. Food Saf. 2006, 5, 169–186. [Google Scholar] [CrossRef]
- Saldaña, M.D.A.; Martínez-Monteagudo, S.I. Oxidative Stability of Fats and Oils Measured by Differential Scanning Calorimetry for Food and Industrial Applications; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Di Mascio, P.; Murphy, M.E.; Sies, H. Antioxidant Defense Systems: The Role of Carotenoids, Tocopherols, and Thiols. Am. J. Clin. Nutr. 1991, 53 (Suppl. S1), 194S–200S. [Google Scholar] [CrossRef] [PubMed]
- Gentili, A.; Caretti, F.; Bellante, S.; Ventura, S.; Canepari, S.; Curini, R. Comprehensive profiling of carotenoids and fat-soluble vitamins in milk from different animal species by LC-DAD-MS/MS hyphenation. J. Agric. Food Chem. 2013, 61, 1628–1639. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Carotenoid Extraction Methods: A Review of Recent Developments. Food Chem. 2017, 240, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-Bermudez, S.P.; Aguilar-Hernandez, I.; Cardenas-Chavez, D.L.; Ornelas-Soto, N.; Romero-Ogawa, M.A.; Parra-Saldivar, R. Extraction and Purification of High-Value Metabolites from Microalgae: Essential Lipids, Astaxanthin and Phycobiliproteins. Microb. Biotechnol. 2015, 8, 190–209. [Google Scholar] [CrossRef]
- Jaeschke, D.P.; Menegol, T.; Rech, R.; Mercali, G.D.; Marczak, L.D.F. Carotenoid and Lipid Extraction from Heterochlorella Luteoviridis Using Moderate Electric Field and Ethanol. Process Biochem. 2016, 51, 1636–1643. [Google Scholar] [CrossRef]
- Liu, Z.; van den Berg, C.; Weusthuis, R.A.; Dragone, G.; Mussatto, S.I. Strategies for an Improved Extraction and Separation of Lipids and Carotenoids from Oleaginous Yeast. Sep. Purif. Technol. 2021, 257, 117946. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COD | Dry Substance | TOC | Total Sugars | Directly Reducing Sugars | Total Nitrogen | Protein | P | K | Na | Ca | Mg | S | References | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kiejdahl | Lowry | |||||||||||||
| gO2/L | gd.w./L | g/L | g/L | g/L | g/L | g/L | g/L | g/L | g/L | g/L | g/L | g/L | g/L | |
| 30.0 ± 0.5 | 37.6 ± 0.0 | 14.7 ± 0.0 | 7.9 ± 0.1 | 7.0 ± 0.1 | 2.6 ± 0.0 | 16.0 ± 0.0 | n.d. | 0.42 ± 0.02 | 3.93 ± 0.25 | 0.58 ± 0.06 | 0.56 ± 0.04 | 0.310 ± 0.015 | 5.88 ± 0.93 | [40] |
| 35.12 ± 3.7 | 33.5 ± 0.5 | n.d. | n.d. | 13.8 ± 0.2 | n.d. | 12.9 ± 0.5 | 4.3 ± 0.4 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | [41] |
| n.d. | 33.26 ± 0.43 | n.d. | n.d. | 4.40 ± 0.03 | 1.62 ± 0.06 | n.d. | n.d. | 0.33 ± 0.02 | 4.14 ± 0.32 | n.d. | 0.10 ± 0.01 | 0.24 ± 0.03 | n.d. | [23] |
| 30.82 ± 0.5 | 39.7 ± 0.5 | n.d. | n.d. | 7.3 ± 0.5 | 2.32 ± 0.07 | 14.5 ± 0.05 | n.d. | 0.27 ± 0.02 | 4.31 ± 0.25 | 0.06 ± 0.00 | 0.16 ± 0.01 | 0.24 ± 0.01 | n.d. | [6] |
| n.d. | 51.9 ± 0.3 | 22.9 ± 0.8 | 17.0 ± 0.1 | 16.4 ± 0.1 | 2.0 ± 0.1 | 12.5 ± 0.1 | n.d. | 0.29 ± 0.01 | 4.1 ± 0.4 | 0.73 ± 0.04 | 1.27 ± 0.2 | 0.27 ± 0.02 | 12.7 ± 2.1 | [42] |
| 28.9 ± 1.9 | 38.19 ± 7.4 | 11.72 ± 0.75 | n.d. | 4.49 ± 0.44 | 1.669 ± 0.047 | 10.43 ± 0.5 | 3.48 ± 0.27 | 0.66 ± 0.01 | 5.29 ± 0.90 | n.d. | 0.014 ± 0.002 | 0.261 ± 0.039 | n.d. | This study |
| Fatty Acid | Batch A | 20 °C Batch B | Batch C | Batch A | 28 °C Batch B | Batch C |
|---|---|---|---|---|---|---|
| C16:0 | 13.11 ± 0.59 | 13.52 ± 0.47 | 12.98 ± 0.39 | 17.83 ± 0.22 | 17.24 ± 0.17 | 17.56 ± 0.79 |
| Δ9C16:1 | 2.41 ± 0.34 | 2.21 ± 0.14 | 2.5 ± 0.16 | 4.15 ± 0.19 | 3.85 ± 0.38 | 3.98 ± 0.21 |
| C17:0 | 0.15 ± 0.03 | 0.11 ± 0.09 | 0.03 ± 0.00 | 2.80 ± 0.22 | 2.79 ± 0.15 | 2.81 ± 0.20 |
| C18:0 | 4.26 ± 0.49 | 3.51 ± 0.52 | 5.02 ± 0.39 | 7.51 ± 0.34 | 7.49 ± 0.33 | 7.53 ± 0.22 |
| Δ9C18:1 | 72.71 ± 1.66 | 74.37 ± 1.13 | 74.05 ± 1.59 | 50.25 ± 1.09 | 51.03 ± 0.98 | 51.52 ± 1.53 |
| Δ9,12C18:2 | 3.69 ± 0.34 | 3.26 ± 0.39 | 3.53 ± 0.22 | 4.11 ± 0.45 | 4.17 ± 0.33 | 4.21 ± 0.59 |
| Δ9,12,15C18:3 | 0.61 ± 0.16 | 0.74 ± 0.09 | 1.14 ± 0.55 | 1.32 ± 0.14 | 1.27 ± 0.32 | 1.24 ± 0.09 |
| C22:0 | n.d. | n.d. | n.d. | 6.05 ± 0.39 | 5.85 ± 0.51 | 5.57 ± 0.09 |
| Sum of others | 1.44 ± 0.19 | 2.03 ± 0.29 | 1.57 ± 0.33 | 3.24 ± 0.24 | 3.14 ± 0.28 | 3.21 ± 0.21 |
| ∑SFA | 18.43 b | 18.19 b | 18.68 b | 35.76 c | 34.92 c | 35.06 c |
| ∑MUFA | 75.66 e | 77.45 e | 77.03 e | 54.72 d | 55.23 d | 55.85 d |
| ∑PUFA | 4.30 a | 4.37 a | 5.08 a | 6.75 a | 6.68 a | 6.72 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gientka, I.; Wirkowska-Wojdyła, M.; Ostrowska-Ligęza, E.; Janowicz, M.; Reczek, L.; Synowiec, A.; Błażejak, S. Enhancing Red Yeast Biomass Yield and Lipid Biosynthesis by Using Waste Nitrogen Source by Glucose Fed-Batch at Low Temperature. Microorganisms 2022, 10, 1253. https://doi.org/10.3390/microorganisms10061253
Gientka I, Wirkowska-Wojdyła M, Ostrowska-Ligęza E, Janowicz M, Reczek L, Synowiec A, Błażejak S. Enhancing Red Yeast Biomass Yield and Lipid Biosynthesis by Using Waste Nitrogen Source by Glucose Fed-Batch at Low Temperature. Microorganisms. 2022; 10(6):1253. https://doi.org/10.3390/microorganisms10061253
Chicago/Turabian StyleGientka, Iwona, Magdalena Wirkowska-Wojdyła, Ewa Ostrowska-Ligęza, Monika Janowicz, Lidia Reczek, Alicja Synowiec, and Stanisław Błażejak. 2022. "Enhancing Red Yeast Biomass Yield and Lipid Biosynthesis by Using Waste Nitrogen Source by Glucose Fed-Batch at Low Temperature" Microorganisms 10, no. 6: 1253. https://doi.org/10.3390/microorganisms10061253