
Evaluation of the Pathogenic Potential of Escherichia coli Strains Isolated from Eye Infections


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Bacterial Strains
2.3. Phylogenetic Analysis, Clonal Relationship, and Sequence Typing
2.4. Virulence Genes Profiles
2.5. Hemolytic Activity
2.6. Antibiotic Susceptibility
2.7. Cell Culture and In-Vitro Adherence to Human Corneal Epithelial Cells Assay
3. Results
3.1. Phylogenetic Analysis, Clonal Relationship, and Sequence Typing
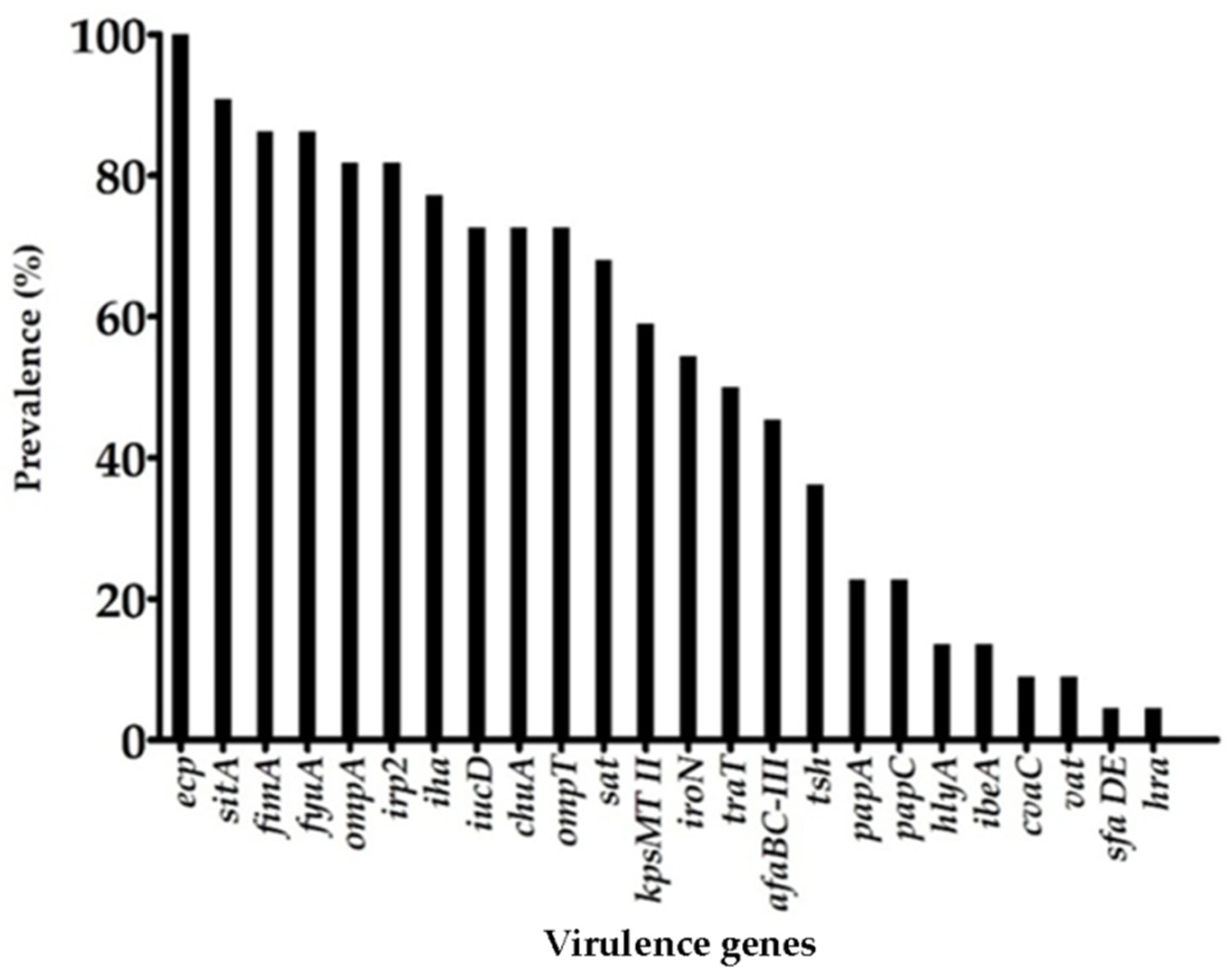
3.2. Virulence Genes Profile
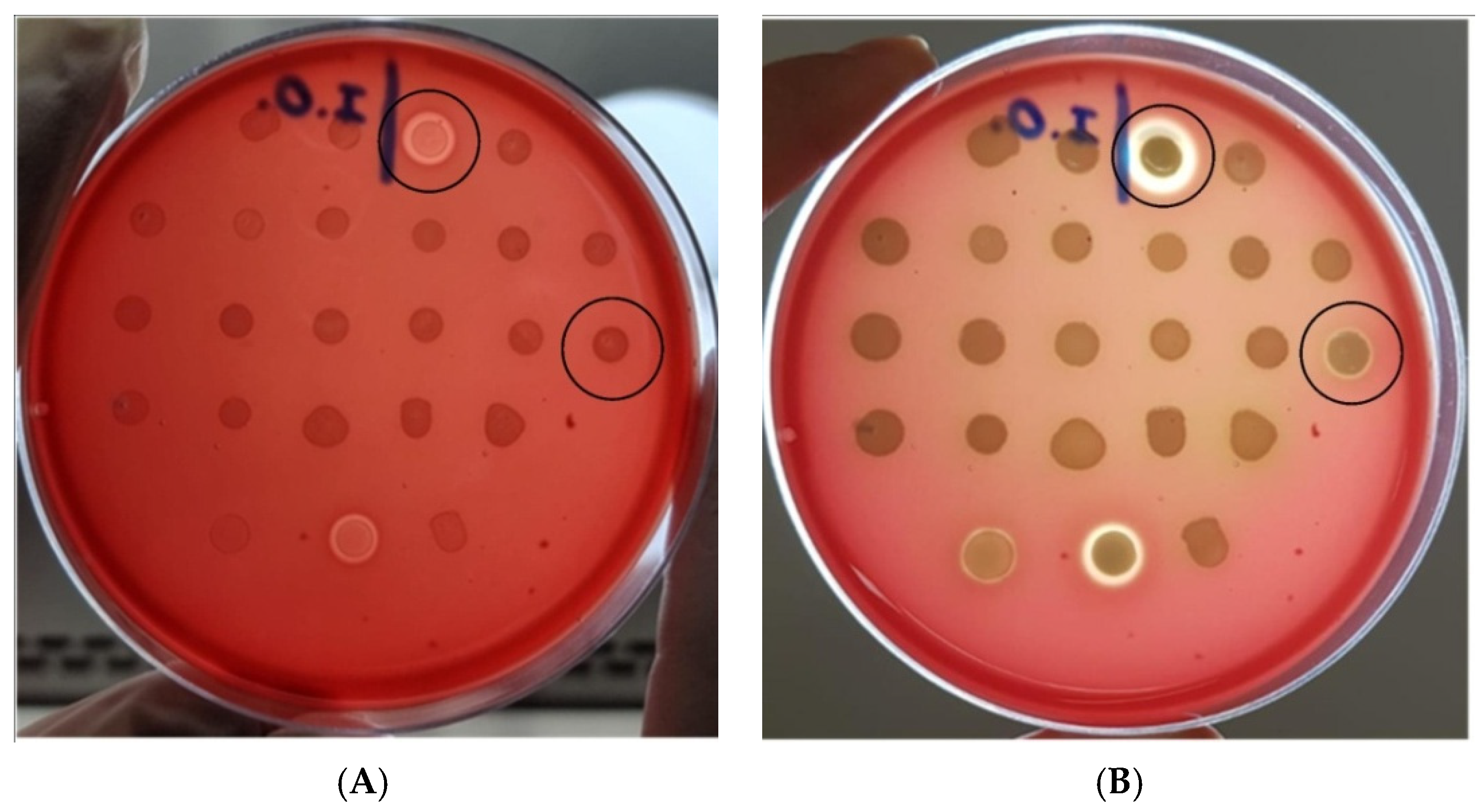
3.3. Hemolytic Activity
3.4. Antibiotic Susceptibility
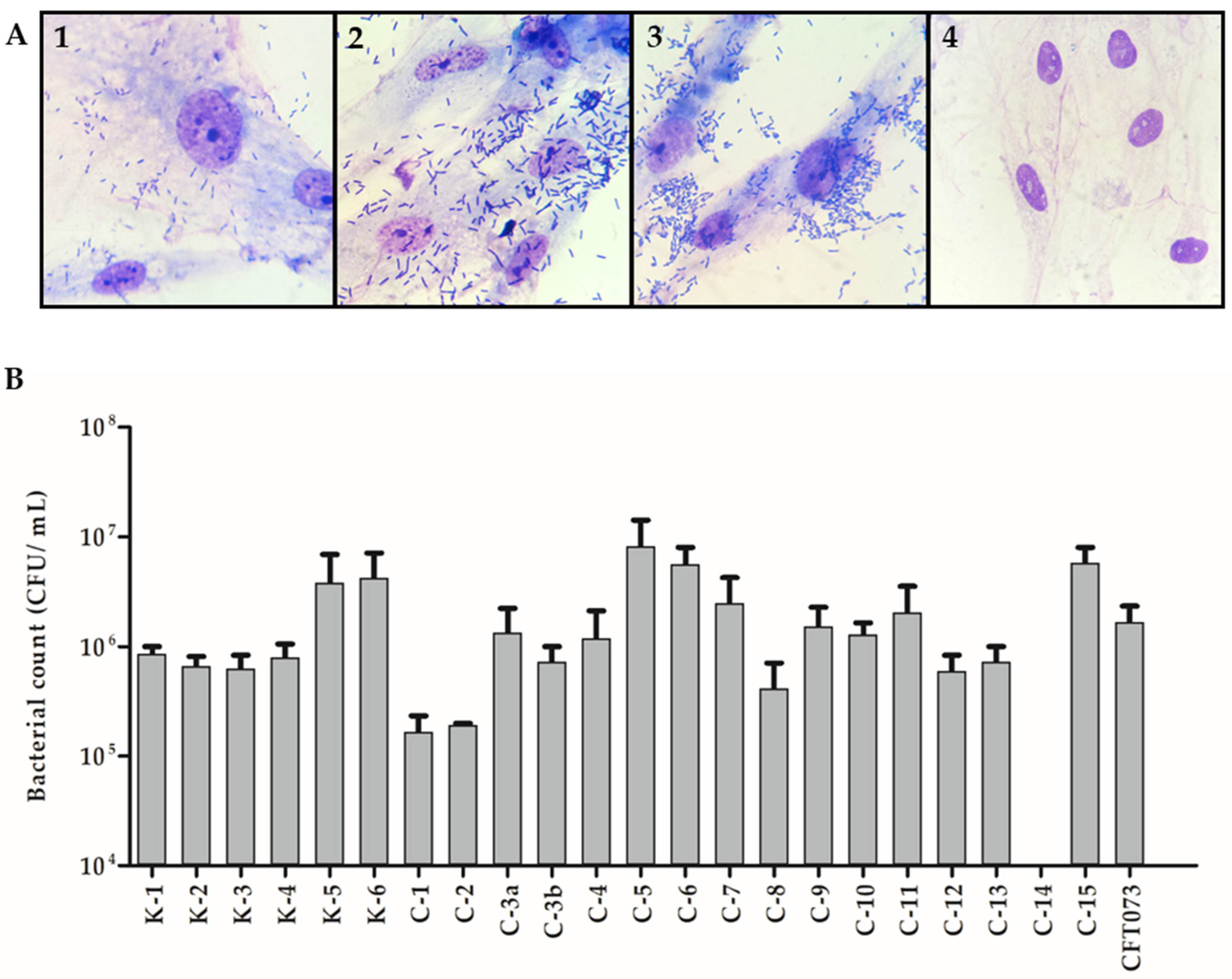
3.5. In-Vitro Adherence to Human Corneal Epithelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willcox, M.D. Characterization of the normal microbiota of the ocular surface. Exp. Eye Res. 2013, 117, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, F.; Pignataro, D.; Lavano, M.A.; Santella, B.; Folliero, V.; Zannella, C.; Astarita, C.; Gagliano, C.; Franci, G.; Avitabile, T.; et al. Current evidence on the ocular surface microbiota and related diseases. Microorganisms 2020, 8, 1033. [Google Scholar] [CrossRef] [PubMed]
- Teweldemedhin, M.; Gebreyesus, H.; Atsbaha, A.H.; Asgedom, S.W.; Saravanan, M. Bacterial profile of ocular infections: A systematic review. BMC Ophthalmol. 2017, 17, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.P.; Woodford, N. Extra-intestinal pathogenic Escherichia coli (ExPEC): Disease, carriage and clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [Green Version]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.-L.; Barnich, N.; Bringer, M.-A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.-F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The population genetics of pathogenic Escherichia coli. Nat. Rev. Genet. 2020, 19, 37–54. [Google Scholar] [CrossRef]
- Johnson, J.R.; Russo, T.A. Molecular Epidemiology of Extraintestinal Pathogenic Escherichia coli. EcoSal Plus 2018, 8. [Google Scholar] [CrossRef]
- Johnson, J.R.; Kuskowski, M.A.; Owens, K.; Gajewski, A.; Winokur, P. Phylogenetic Origin and Virulence Genotype in Relation to Resistance to Fluoroquinolones and/or Extended—Spectrum Cephalosporins and Cephamycins among Escherichia coli Isolates from Animals and Humans. J. Infect. Dis. 2003, 188, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Spurbeck, R.R.; Dinh, P.C., Jr.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.C.M.; Zidko, A.C.M.; Pignatari, A.C.; Silva, R.M. Assessing the diversity of the virulence potential of Escherichia coli isolated from bacteremia in São Paulo, Brazil. Braz. J. Med. Biol. Res. 2013, 46, 968–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.C.M.; Santos, F.F.; Silva, R.M.; Gomes, T.A.T. Diversity of Hybrid- and Hetero-Pathogenic Escherichia coli and their potential implication in more severe diseases. Front. Cell. Infect. Microbiol. 2020, 10, 339. [Google Scholar] [CrossRef] [PubMed]
- Mariani-Kurkdjian, P.; Lemaître, C.; Bidet, P.; Perez, D.; Boggini, L.; Kwon, T.; Bonacorsi, S. Haemolytic-uraemic syndrome with bacteraemia caused by a new hybrid Escherichia coli pathotype. New Microbes New Infect. 2014, 2, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.; Nisa, S.; Hazen, T.H.; Horneman, A.; Amoroso, A.; Rasko, D.A.; Donnenberg, M.S. Diarrhea, bacteremia and multiorgan dysfunction due to an extraintestinal pathogenic Escherichia coli strain with enteropathogenic E. coli genes. Pathog. Dis. 2015, 73, ftv076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gati, N.S.; Middendorf-Bauchart, B.; Bletz, S.; Dobrindt, U.; Mellmann, A. Origin and Evolution of Hybrid Shiga Toxin-Producing and Uropathogenic Escherichia coli Strains of Sequence Type 141. J. Clin. Microbiol. 2019, 58, e01309-19. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Pacheco, A.B.F.; Guth, B.E.C.; Scares, K.C.C.; De Almeida, D.F.; Ferreira, L.C.S. Clonal relationships among Escherichia coli serogroup O6 isolates based on RAPD. FEMS Microbiol. Lett. 1997, 148, 255–260. [Google Scholar] [CrossRef]
- Nielsen, K.L.; Dynesen, P.; Larsen, P.; Frimodt-Møller, N. Faecal Escherichia coli from patients with E. coli urinary tract infection and healthy controls who have never had a urinary tract infection. J. Med. Microbiol. 2014, 63, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Doumith, M.; Day, M.; Ciesielczuk, H.; Hope, R.; Underwood, A.; Reynolds, R.; Wain, J.; Livermore, D.M.; Woodford, N. Rapid Identification of Major Escherichia coli Sequence Types Causing Urinary Tract and Bloodstream Infections. J. Clin. Microbiol. 2015, 53, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, J.A.S.; Santos, F.F.; Santos-Neto, J.F.; Trovão, L.O.; Valiatti, T.B.; Pinaffi, I.C.; Vieira, M.A.M.; Silva, R.M.; Falsetti, I.N.; Santos, A.C.M.; et al. Molecular Epidemiology and Presence of Hybrid Pathogenic Escherichia coli among Isolates from Community-Acquired Urinary Tract Infection. Microorganisms 2022, 10, 302. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Montenegro, M.A.; Orskov, I.; Orskov, F.; Prada, J.; Zimmermann, S.; Stephan, R. Close association of verotoxin (Shiga-like toxin) production with enterohemolysin production in strains of Escherichia coli. J. Clin. Microbiol. 1989, 27, 2559–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Valiatti, T.B.; Santos, F.F.; Santos, A.C.M.; Nascimento, J.A.S.; Silva, R.M.; Carvalho, E.; Sinigaglia, R.; Gomes, T.A.T. Genetic and Virulence Characteristics of a Hybrid Atypical Enteropathogenic and Uropathogenic Escherichia coli (aEPEC/UPEC) Strain. Front. Cell. Infect. Microbiol. 2020, 10, 492. [Google Scholar] [CrossRef] [PubMed]
- Van Tyne, D.; Ciolino, J.B.; Wang, J.; Durand, M.L.; Gilmore, M.S. Novel Phagocytosis-Resistant Extended-Spectrum β-Lactamase-Producing Escherichia coli From Keratitis. JAMA Ophthalmol. 2016, 134, 1306–1309. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Shen, L.; Wang, A.; Li, M.; Yang, C. Bacterial Culture of Tear Duct Infections Secondary to Congenital Nasolacrimal Duct Obstructions. J. Ophthalmol. 2022, 2022, 9954634. [Google Scholar] [CrossRef]
- Manente, R.; Santella, B.; Pagliano, P.; Santoro, E.; Casolaro, V.; Borrelli, A.; Capunzo, M.; Galdiero, M.; Franci, G.; Boccia, G. Prevalence and Antimicrobial Resistance of Causative Agents to Ocular Infections. Antibiotics 2022, 11, 463. [Google Scholar] [CrossRef]
- Saralaya, V.; Shenoy, S.; Baliga, S.; Hegde, A.; Adhikari, P.; Chakraborty, A. Characterization of Escherichia coli phylogenetic groups associated with extraintestinal infections in South Indian population. Ann. Med. Health Sci. Res. 2015, 5, 241. [Google Scholar] [CrossRef] [Green Version]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection? Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Kohoutova, D.; Smajs, D.; Moravkova, P.; Cyrany, J.; Moravkova, M.; Forstlova, M.; Cihak, M.; Rejchrt, S.; Bures, J. Escherichia coli strains of phylogenetic group B2 and D and bacteriocin production are associated with advanced colorectal neoplasia. BMC Infect. Dis. 2014, 14, 733. [Google Scholar] [CrossRef] [Green Version]
- Nowrouzian, F.L.; Wold, A.E.; Adlerberth, I. Escherichia coli Strains Belonging to Phylogenetic Group B2 Have Superior Capacity to Persist in the Intestinal Microflora of Infants. J. Infect. Dis. 2005, 191, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended Virulence Genotypes of Escherichia coli Strains from Patients with Urosepsis in Relation to Phylogeny and Host Compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahwash, S.M.; Raheema, R.H.; Albahadili, M.A.; Maslat, A.H. Distribution of Phylogenetics and Virulence Genes of Uropathogenic Escherichia coli Among Urinary Tract Infection in Pregnant Women. Biochem. Cell Arch. 2021, 21, 449–456. [Google Scholar]
- Iranpour, D.; Hassanpour, M.; Ansari, H.; Tajbakhsh, S.; Khamisipour, G.; Najafi, A. Phylogenetic Groups of Escherichia coli Strains from Patients with Urinary Tract Infection in Iran Based on the New Clermont Phylotyping Method. BioMed Res. Int. 2015, 2015, 846219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezatofighi, S.E.; Mirzarazi, M.; Salehi, M. Virulence genes and phylogenetic groups of uropathogenic Escherichia coli isolates from patients with urinary tract infection and uninfected control subjects: A case-control study. BMC Infect. Dis. 2021, 21, 361. [Google Scholar] [CrossRef]
- Regua-Mangia, A.H.; Irino, K.; Pacheco, R.D.S.; Bezerra, R.M.P.; Périssé, A.R.S.; Teixeira, L.M. Molecular characterization of uropathogenic and diarrheagenic Escherichia coli pathotypes. J. Basic Microbiol. 2010, 50, S107–S115. [Google Scholar] [CrossRef]
- Grude, N.; Potaturkina-Nesterova, N.I.; Jenkins, A.; Strand, L.; Nowrouzian, F.L.; Nyhus, J.; Kristiansen, B.-E. A comparison of phylogenetic group, virulence factors and antibiotic resistance in Russian and Norwegian isolates of Escherichia coli from urinary tract infection. Clin. Microbiol. Infect. 2007, 13, 208–211. [Google Scholar] [CrossRef] [Green Version]
- Nicolas-Chanoine, M.-H.; Bertrand, X.; Madec, J.-Y. Escherichia coli ST131, an Intriguing Clonal Group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Johnson, J.R. A new clone sweeps clean: The enigmatic emergence of Escherichia coli Sequence Type 131. Antimicrob. Agents Chemother. 2014, 58, 4997–5004. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.D.; DeVinney, R. Escherichia coli ST131: A multidrug-resistant clone primed for global domination. F1000Research 2017, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Colpan, A.; Johnston, B.; Porter, S.; Clabots, C.; Anway, R.; Thao, L.; Kuskowski, M.A.; Tchesnokova, V.; Sokurenko, E.V.; Johnson, J.R.; et al. Escherichia coli sequence type 131 (ST131) subclone H30 as an emergent multidrug-resistant pathogen among US veterans. Clin. Infect. Dis. 2013, 57, 1256–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Ewers, C.; Nandanwar, N.; Guenther, S.; Jadhav, S.; Wieler, L.H.; Ahmed, N. Multiresistant Uropathogenic Escherichia coli from a Region in India Where Urinary Tract Infections Are Endemic: Genotypic and Phenotypic Characteristics of Sequence Type 131 Isolates of the CTX-M-15 Extended-Spectrum-β-Lactamase-Producing Lineage. Antimicrob. Agents Chemother. 2012, 56, 6358–6365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, D.A.; Freeman, J.T.; Porter, S.; Roberts, S.; Wiles, S.; Paterson, D.L.; Johnson, J.R. Clinical and molecular correlates of virulence in Escherichia coli causing bloodstream infection following transrectal ultrasound-guided (TRUS) prostate biopsy. J. Antimicrob. Chemother. 2013, 68, 2898–2906. [Google Scholar] [CrossRef] [PubMed]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-PLoSkonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sannes, M.R.; Kuskowski, M.A.; Owens, K.; Gajewski, A.; Johnson, J.R. Virulence Factor Profiles and Phylogenetic Background of Escherichia coli Isolates from Veterans with Bacteremia and Uninfected Control Subjects. J. Infect. Dis. 2004, 190, 2121–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khasheii, B.; Anvari, S.; Jamalli, A. Frequency evaluation of genes encoding siderophores and the effects of different concentrations of Fe ions on growth rate of uropathogenic Escherichia coli. Iran J. Microbiol. 2016, 8, 359–365. [Google Scholar]
- Searle, L.J.; Méric, G.; Porcelli, I.; Sheppard, S.K.; Lucchini, S. Variation in siderophore biosynthetic gene distribution and production across environmental and faecal populations of Escherichia coli. PLoS ONE 2015, 10, e0117906. [Google Scholar] [CrossRef] [Green Version]
- Sabri, M.; Léveillé, S.; Dozois, C.M. A SitABCD homologue from an avian pathogenic Escherichia coli strain mediates transport of iron and manganese and resistance to hydrogen peroxide. Microbiology 2006, 152, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Wijetunge, D.S.S.; Gongati, S.; DebRoy, C.; Kim, K.S.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Kariyawasam, S. Characterizing the pathotype of neonatal meningitis causing Escherichia coli (NMEC). BMC Microbiol. 2015, 15, 211. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, J.; Willcox, M. Role of lactoferrin in the tear film. Biochimie 2009, 91, 35–43. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Stapleton, A.E.; Johnson, J.R.; Walk, S.T.; Hooton, T.M.; Mobley, H.L. Fimbrial profiles predict virulence of uropathogenic Escherichia coli Strains: Contribution of Ygi and Yad fimbriae. Infect. Immun. 2011, 79, 4753–4763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.; Blanco, J.E.; Alonso, M.P.; Mora, A.; Balsalobre, C.; Munoa, F.; Juárez, A.; Blanco, J. Detection of pap, sfa and afa adhesin-encoding operons in uropathogenic Escherichia coli strains: Relationship with expression of adhesins and production of toxins. Res. Microbiol. 1997, 148, 745–755. [Google Scholar] [CrossRef]
- Antão, E.M.; Wieler, L.H.; Ewers, C. Adhesive threads of extraintestinal pathogenic Escherichia coli. Gut Pathog. 2009, 1, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendón, M.A.; Saldaña, Z.; Erdem, A.L.; Monteiro-Neto, V.; Vázquez, A.; Kaper, J.B.; Puente, J.L.; Girón, J.A. Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proc. Natl. Acad. Sci. USA 2007, 104, 10637–10642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Koch, W.H.; Cebula, T.A. Detection and characterization of the fimA gene of Escherichia coli O157:H7. Mol. Cell Probes. 1997, 11, 397–406. [Google Scholar] [CrossRef]
- Müller, C.M.; Åberg, A.; Straseviçiene, J.; Emődy, L.; Uhlin, B.E.; Balsalobre, C. Type 1 fimbriae, a colonization factor of uropathogenic Escherichia coli, are controlled by the metabolic sensor CRP-cAMP. PLoS Pathog. 2009, 5, e1000303. [Google Scholar] [CrossRef] [Green Version]
- Connell, I.; Agace, W.; Klemm, P.; Schembri, M.; Mărild, S.; Svanborg, C. Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract. Proc. Natl. Acad. Sci. USA 1996, 93, 9827–9832. [Google Scholar] [CrossRef] [Green Version]
- Hull, R.A.; Donovan, W.H.; Del Terzo, M.; Stewart, C.; Rogers, M.; Darouiche, R.O. Role of Type 1 Fimbria- and P Fimbria-Specific Adherence in Colonization of the Neurogenic Human Bladder by Escherichia coli. Infect. Immun. 2002, 70, 6481–6484. [Google Scholar] [CrossRef] [Green Version]
- Vizcarra, I.A.; Hosseini, V.; Kollmannsberger, P.; Meier, S.; Weber, S.S.; Arnoldini, M.; Ackermann, M.; Vogel, V. How type 1 fimbriae help Escherichia coli to evade extracellular antibiotics. Sci. Rep. 2016, 6, 18109. [Google Scholar] [CrossRef] [Green Version]
- Labbate, M.; Zhu, H.; Thung, L.; Bandara, R.; Larsen, M.R.; Willcox, M.D.P.; Givskov, M.; Rice, S.A.; Kjelleberg, S. Quorum-Sensing Regulation of Adhesion in Serratia marcescens MG1 Is Surface Dependent. J. Bacteriol. 2007, 189, 2702–2711. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Wan, Z.S.; Chen, Y.H.; Jong, A.Y.; Kim, K.S. Further characterization of Escherichia coli brain microvascular endothelial cell invasion gene ibeA by deletion, complementation, and protein expression. J. Infect. Dis. 2001, 183, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieza, R.J.; Hu, J.; Ross, B.N.; Sbrana, E.; Torres, A.G. The IbeA invasin of adherent-invasive Escherichia coli mediates interaction with intestinal epithelia and macrophages. Infect. Immun. 2015, 83, 1904–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flechard, M.; Cortes, M.A.M.; Reperant, M.; Germon, P. New Role for the ibeA Gene in H2O2 Stress Resistance of Escherichia coli. J. Bacteriol. 2012, 194, 4550–4560. [Google Scholar] [CrossRef] [Green Version]
- Che, X.; Chi, F.; Wang, L.; Jong, T.D.; Wu, C.H.; Wang, X.; Huang, S.H. Involvement of IbeA in meningitic Escherichia coli K1-induced polymorphonuclear leukocyte transmigration across brain endothelial cells. Brain Pathol. 2011, 21, 389–404. [Google Scholar] [CrossRef] [PubMed]
- Wiles, T.J.; Mulvey, M.A. The RTX pore-forming toxin α-hemolysin of uropathogenic Escherichia coli: Progress and perspectives. Futur. Microbiol. 2013, 8, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, Q.; Lv, J.; Sun, X.; Cao, Y.; Yu, K.; Miao, C.; Zhang, Z.-S.; Yao, Z.; Wang, Q. Alpha-hemolysin of uropathogenic Escherichia coli induces GM-CSF-mediated acute kidney injury. Mucosal. Immunol. 2019, 13, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.J.A.; Koronakis, V.; Schmoll, T.; Hughes, C. Escherichia coli HIyT protein, a transcriptional activator of haemolysin synthesis and secretion, is encoded by the rfaH (sfrB) locus required for expression of sex factor and lipopolysaccharide genes. Mol. Microbiol. 1992, 6, 1003–1012. [Google Scholar] [CrossRef]
- Kerényi, M.; Allison, H.E.; Bátai, I.; Sonnevend, A.; Emödy, L.; Plaveczky, N.; Pál, T. Occurrence of hlyA and sheA genes in extraintestinal Escherichia coli strains. J. Clin. Microbiol. 2005, 43, 2965–2968. [Google Scholar] [CrossRef] [Green Version]
- Beutin, L.; Prada, J.; Zimmermann, S.; Stephan, R.; Ørskov, I.; Ørskov, F. Enterohemolysin, a new type of hemolysin produced by some strains of enteropathogenic E. coli (EPEC). Zent. Bakteriol. Mikrobiol. Hygiene. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1988, 267, 576–588. [Google Scholar] [CrossRef]
- Boczek, L.A.; Johnson, C.H.; Rice, E.W.; Kinkle, B.K. The Widespread Occurrence of the Enterohemolysin Gene ehlyA Among Environmental Strains of Escherichia coli. Proc. Water Environ. Fed. 2009, 2009, 2992–2995. [Google Scholar] [CrossRef]
- Schwidder, M.; Heinisch, L.; Schmidt, H. Genetics, Toxicity, and Distribution of Enterohemorrhagic Escherichia coli Hemolysin. Toxins 2019, 11, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putra, I.; Rabiee, B.; Anwar, K.N.; Gidfar, S.; Shen, X.; Babalooee, M.; Ghassemi, M.; Afsharkhamseh, N.; Bakhsh, S.; Missiakas, D.; et al. Staphylococcus aureus alpha-hemolysin impairs corneal epithelial wound healing and promotes intracellular bacterial invasion. Exp. Eye Res. 2019, 181, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.K.; Allen, J.H. The role of hemolysin in corneal infections with Pseudomonas aeruginosa. Investig. Ophthalmol. Vis. Sci. 1978, 17, 480–483. [Google Scholar]
- Stevens, S.X.; Jensen, H.G.; Jett, B.D.; Gilmore, M.S. A hemolysin-encoding plasmid contributes to bacterial virulence in experimental Enterococcus faecalis endophthalmitis. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1650–1656. [Google Scholar]
- Beecher, D.J.; Pulido, J.S.; Barney, N.P.; Wong, A.C. Extracellular virulence factors in Bacillus cereus endophthalmitis: Methods and implication of involvement of hemolysin BL. Infect. Immun. 1995, 63, 632–639. [Google Scholar] [CrossRef] [Green Version]
- Callegan, M.C.; Jett, B.D.; Hancock, L.E.; Gilmore, M.S. Role of Hemolysin BL in the Pathogenesis of Extraintestinal Bacillus cereus Infection Assessed in an Endophthalmitis Model. Infect. Immun. 1999, 67, 3357–3366. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.A.; Ali, M.M.; Zenebe, M.H. Bacterial etiology of ocular and periocular infections, antimicrobial susceptibility profile and associated factors among patients attending eye unit of Shashemene comprehensive specialized hospital, Shashemene, Ethiopia. BMC Ophthalmol. 2020, 20, 124. [Google Scholar] [CrossRef]
- Getahun, E.; Gelaw, B.; Assefa, A.; Assefa, Y.; Amsalu, A. Bacterial pathogens associated with external ocular infections alongside eminent proportion of multidrug resistant isolates at the University of Gondar Hospital, northwest Ethiopia. BMC Ophthalmol. 2017, 17, 151. [Google Scholar] [CrossRef] [Green Version]
- Ranjith, K.; Arunasri, K.; Reddy, G.S.; Adicherla, H.; Sharma, S.; Shivaji, S. Global gene expression in Escherichia coli, isolated from the diseased ocular surface of the human eye with a potential to form biofilm. Gut Pathog. 2017, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, S.; Zhang, S.; Zhao, Z.; Cao, Y.; Chen, M.; Li, B. A Comparative Study of Fluoroquinolone-Resistant Escherichia coli Lineages Portrays Indistinguishable Pathogenicity- and Survivability-Associated Phenotypic Characteristics Between ST1193 and ST131. Infect. Drug Resist. 2020, 13, 4167–4175. [Google Scholar] [CrossRef]
- Okesola, A.; Salako, A. Microbiological profile of bacterial conjunctivitis in Ibadan, Nigeria. Ann. Ib. Postgrad. Med. 2011, 8, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, H.; Bispo, P.J.; Kobashi, H.; Jacobs, D.; Gilmore, M.S.; Ciolino, J.B. Resolution of fluoroquinolone-resistant Escherichia coli keratitis with a PROSE device for enhanced targeted antibiotic delivery. Am. J. Ophthalmol. Case Rep. 2018, 12, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Rameshkumar, G.; Ramakrishnan, R.; Shivkumar, C.; Meenakshi, R.; Anitha, V.; Reddy, Y.C.V.; Maneksha, V. Prevalence and antibacterial resistance patterns of extended-spectrum beta-lactamase producing Gram-negative bacteria isolated from ocular infections. Indian J. Ophthalmol. 2016, 64, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Asbell, P.A.; Sanfilippo, C.M.; Mah, F.S. Antibiotic susceptibility of bacterial pathogens isolated from the aqueous and vitreous humor in the Antibiotic Resistance Monitoring in Ocular micRoorganisms (ARMOR) Surveillance Study: 2009–2020 update. J. Glob. Antimicrob. Resist. 2022, 29, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Sobel, J.D. Bacterial Adherence in the Pathogenesis of Urinary Tract Infection: A Review. Clin. Infect. Dis. 1987, 9, 470–487. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Disease | Phylogroup | Sequence Type a |
|---|---|---|---|
| K-1 | Keratitis | A | - |
| K-2 | Keratitis | B1 | - |
| K-3 | Keratitis | B2 | - |
| K-4 | Keratitis | B2 | - |
| K-5 | Keratitis | B2 | ST131 |
| K-6 | Keratitis | C | - |
| C-1 | Conjunctivitis | A | - |
| C-2 | Conjunctivitis | A | - |
| C-3a | Conjunctivitis | B2 | ST131 |
| C-3b | Conjunctivitis | B2 | ST131 |
| C-4 | Conjunctivitis | B2 | ST131 |
| C-5 | Conjunctivitis | B2 | ST131 |
| C-6 | Conjunctivitis | B2 | ST131 |
| C-7 | Conjunctivitis | B2 | ST131 |
| C-8 | Conjunctivitis | B2 | ST131 |
| C-9 | Conjunctivitis | B2 | ST131 |
| C-10 | Conjunctivitis | B2 | ST131 |
| C-11 | Conjunctivitis | B2 | ST131 |
| C-12 | Conjunctivitis | D | - |
| C-13 | Conjunctivitis | D | - |
| C-14 | Conjunctivitis | F | - |
| C-15 | Conjunctivitis | F | - |
| Strain | Phylogroup | Virulence Markers | Intrinsic Virulence a | Uropathogenicity a | Predicted Pathotype |
|---|---|---|---|---|---|
| K-1 | A | ecp, sitA, fimA, ompA, ompT, cvaC | iucD, kpsMTII, papA | - | ExPEC |
| K-2 | B1 | ecp, fimA, ompA, ompT | - | - | - |
| K-3 | B2 | ecp, fimA, ompA, irp2, iha, ompT, sat, traT, hra | kpsMTII, papA, papC, sfaDE | fyuA, chuA, yfcV, vat | ExPEC/UPEC |
| K-4 | B2 | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat, tsh | iucD, kpsMTII | fyuA, chuA, yfcV, vat | ExPEC/UPEC |
| K-5 | B2 | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat, iroN, traT, tsh | iucD, papC | fyuA, chuA | ExPEC |
| K-6 | C | ecp, sitA, fimA, ompA, irp2, ompT, cvaC | papA | fyuA, yfcV | - |
| C-1 | A | ecp, sitA, fimA, irp2, iha, sat, iroN, traT, tsh | iucD, afaBCIII | fyuA | ExPEC |
| C-2 | A | ecp, sitA, ompA, irp2, iha | iucD, afaBCIII | fyuA | ExPEC |
| C-3a | B2 | ecp, sitA, fimA, ompA, irp2, iha, sat, iroN, ibeA, hlyA | afaBCIII | fyuA, chuA, yfcV | UPEC |
| C-3b | B2 | ecp, sitA, fimA, ompA, iha, ompT, sat, iroN, hlyA | kpsMTII, afaBCIII | fyuA, chuA | ExPEC |
| C-4 | B2 | ecp, sitA, fimA, ompA, iha, ompT, sat, iroN, traT, ibeA, hlyA | iucD, kpsMTII, afaBC III, papC | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-5 | B2 | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat, iroN, traT | iucD | chuA, yfcV | - |
| C-6 | B2 | ecp, sitA, fimA, irp2, iha, ompT, tsh | iucD, kpsMTII | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-7 | B2 | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat, iroN, traT, tsh | iucD, kpsMTII, afaBCIII | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-8 | B2 | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat | iucD, kpsMTII | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-9 | B2 | ecp, sitA, fimA, ompA, irp2, iha, sat, tsh | iucD | fyuA, chuA, yfcV | UPEC |
| C-10 | B2 | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat, iroN, traT, tsh | iucD, kpsMTII, afaBCIII | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-11 | B2 | ecp, sitA, fimA, irp2, iha, ompT, sat, iroN, traT, tsh, ibeA | iucD, kpsMTII | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-12 | D | ecp, sitA, ompA, irp2, iroN, traT | iucD, kpsMTII, afaBCIII | fyuA | ExPEC |
| C-13 | D | ecp, sitA, fimA, irp2, iha, iroN, traT | iucD, kpsMTII, afaBCIII | fyuA, chuA | ExPEC |
| C-14 | F | ecp, sitA, fimA, ompA, irp2, iha, ompT, sat | iucD, kpsMTII, papA, papC | fyuA, chuA, yfcV | ExPEC/UPEC |
| C-15 | F | ecp, sitA, ompA, irp2, ompT, sat, iroN, traT | kpsMTII, afaBCIII, papA, papC | fyuA, chuA, yfcV | ExPEC/UPEC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, P.H.S.; Valiatti, T.B.; Santos, A.C.d.M.; Nascimento, J.A.d.S.; Santos-Neto, J.F.; Rocchetti, T.T.; Yu, M.C.Z.; Hofling-Lima, A.L.; Gomes, T.A.T. Evaluation of the Pathogenic Potential of Escherichia coli Strains Isolated from Eye Infections. Microorganisms 2022, 10, 1084. https://doi.org/10.3390/microorganisms10061084
Nunes PHS, Valiatti TB, Santos ACdM, Nascimento JAdS, Santos-Neto JF, Rocchetti TT, Yu MCZ, Hofling-Lima AL, Gomes TAT. Evaluation of the Pathogenic Potential of Escherichia coli Strains Isolated from Eye Infections. Microorganisms. 2022; 10(6):1084. https://doi.org/10.3390/microorganisms10061084
Chicago/Turabian StyleNunes, Pedro Henrique Soares, Tiago Barcelos Valiatti, Ana Carolina de Mello Santos, Júllia Assis da Silva Nascimento, José Francisco Santos-Neto, Talita Trevizani Rocchetti, Maria Cecilia Zorat Yu, Ana Luisa Hofling-Lima, and Tânia Aparecida Tardelli Gomes. 2022. "Evaluation of the Pathogenic Potential of Escherichia coli Strains Isolated from Eye Infections" Microorganisms 10, no. 6: 1084. https://doi.org/10.3390/microorganisms10061084