
Lignocellulose Fermentation Products Generated by Giant Panda Gut Microbiomes Depend Ultimately on pH Rather than Portion of Bamboo: A Preliminary Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
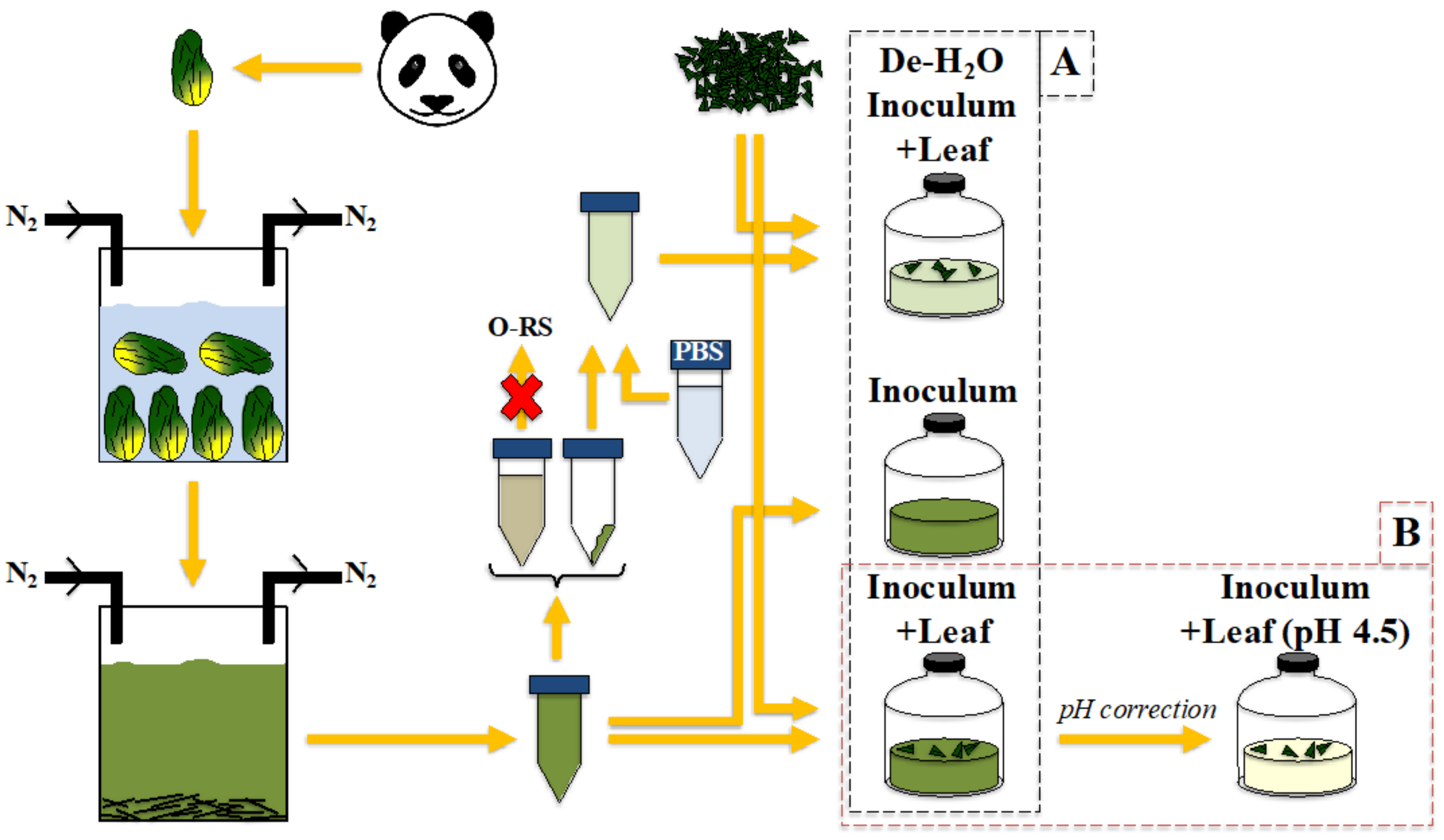
2.1. Inoculum Collection, Preparation and Cultivation System
2.2. Microbiological Analysis
2.3. Molecular Analysis
2.4. Metaproteomics
2.5. Chemical Analysis
2.6. Statistical Analysis
3. Results
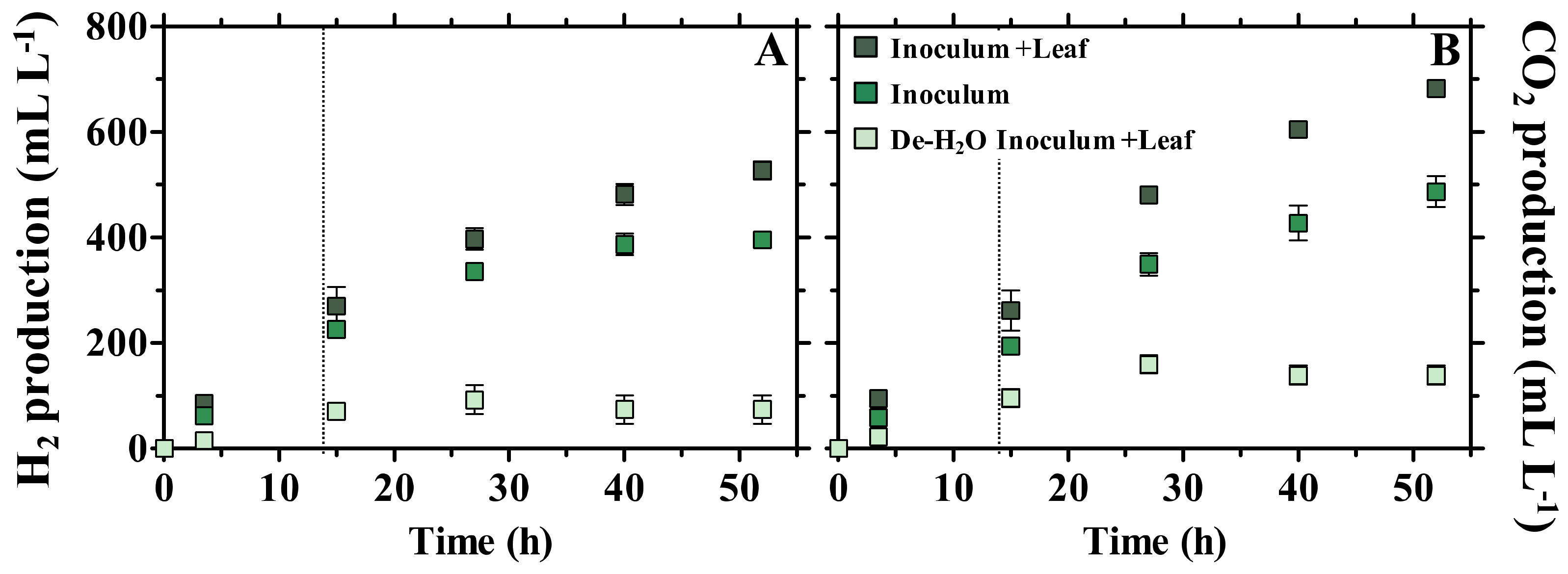
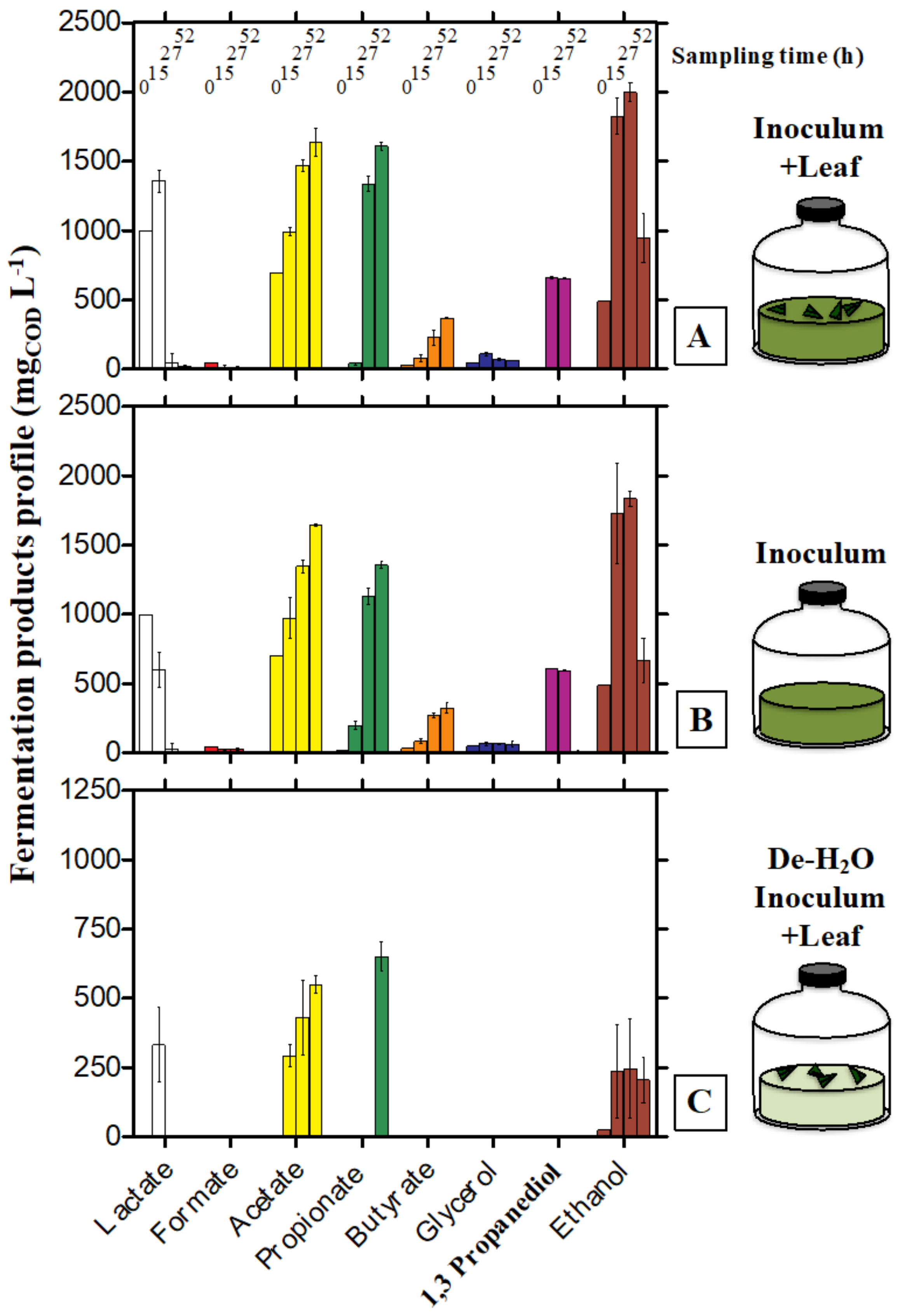
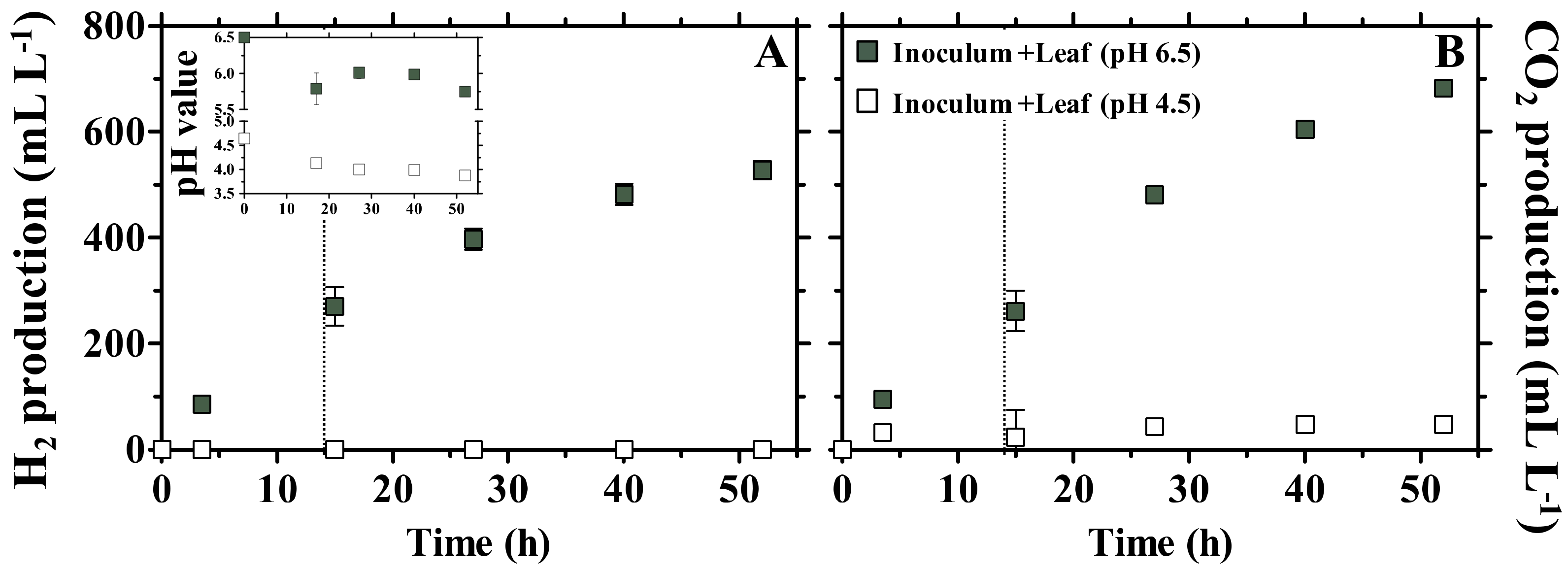
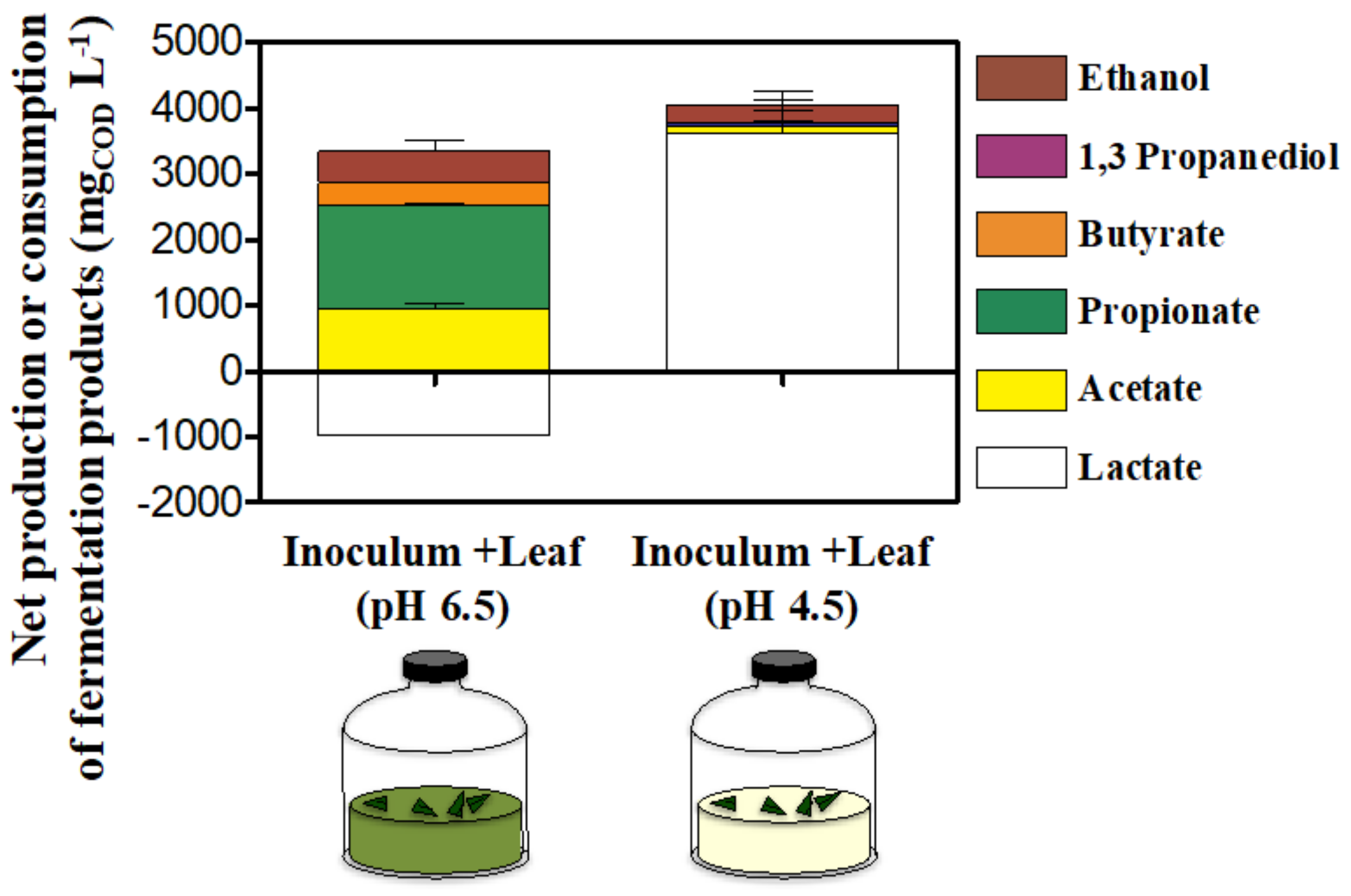
3.1. Fermentation Products
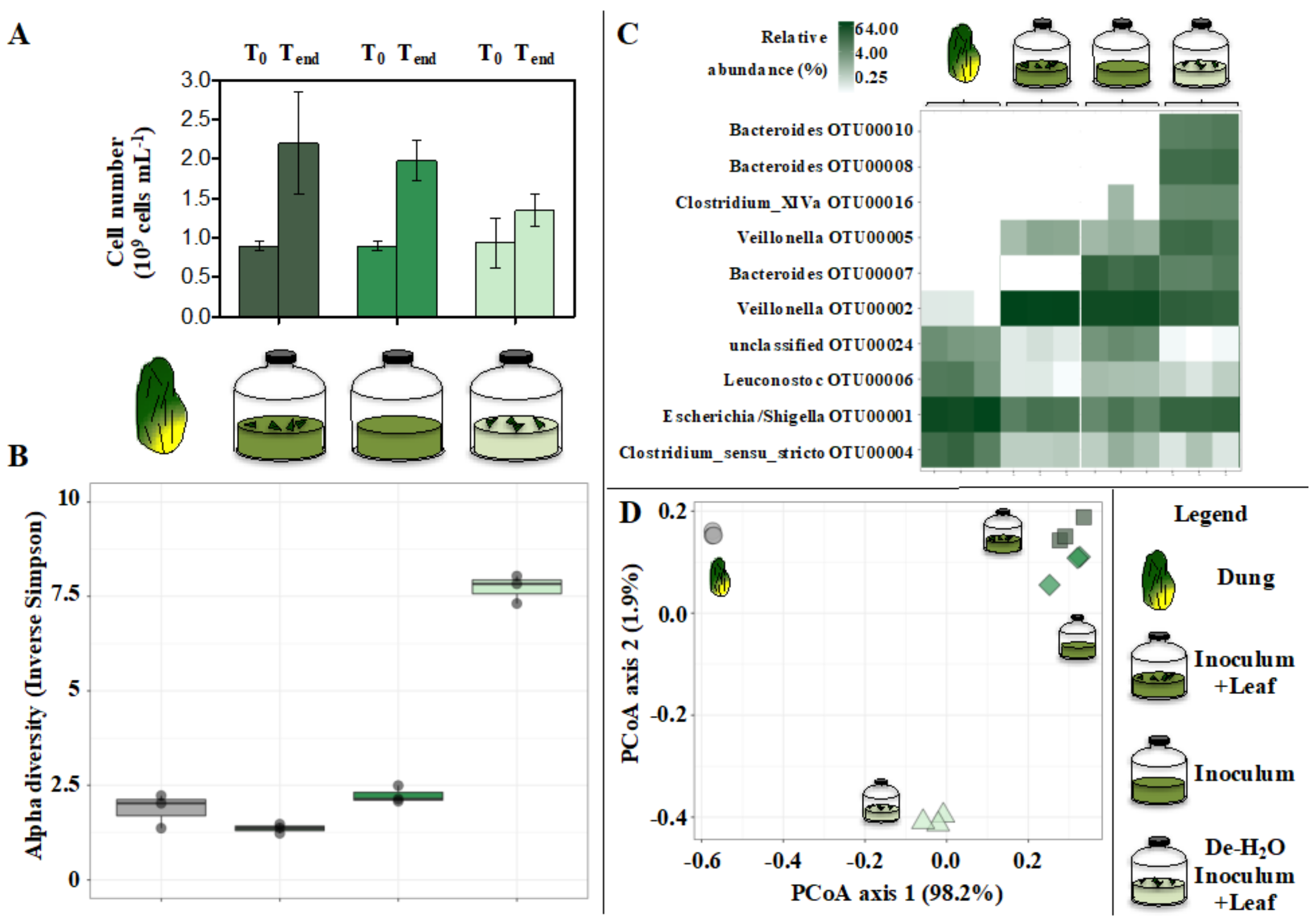
3.2. Metaproteomics
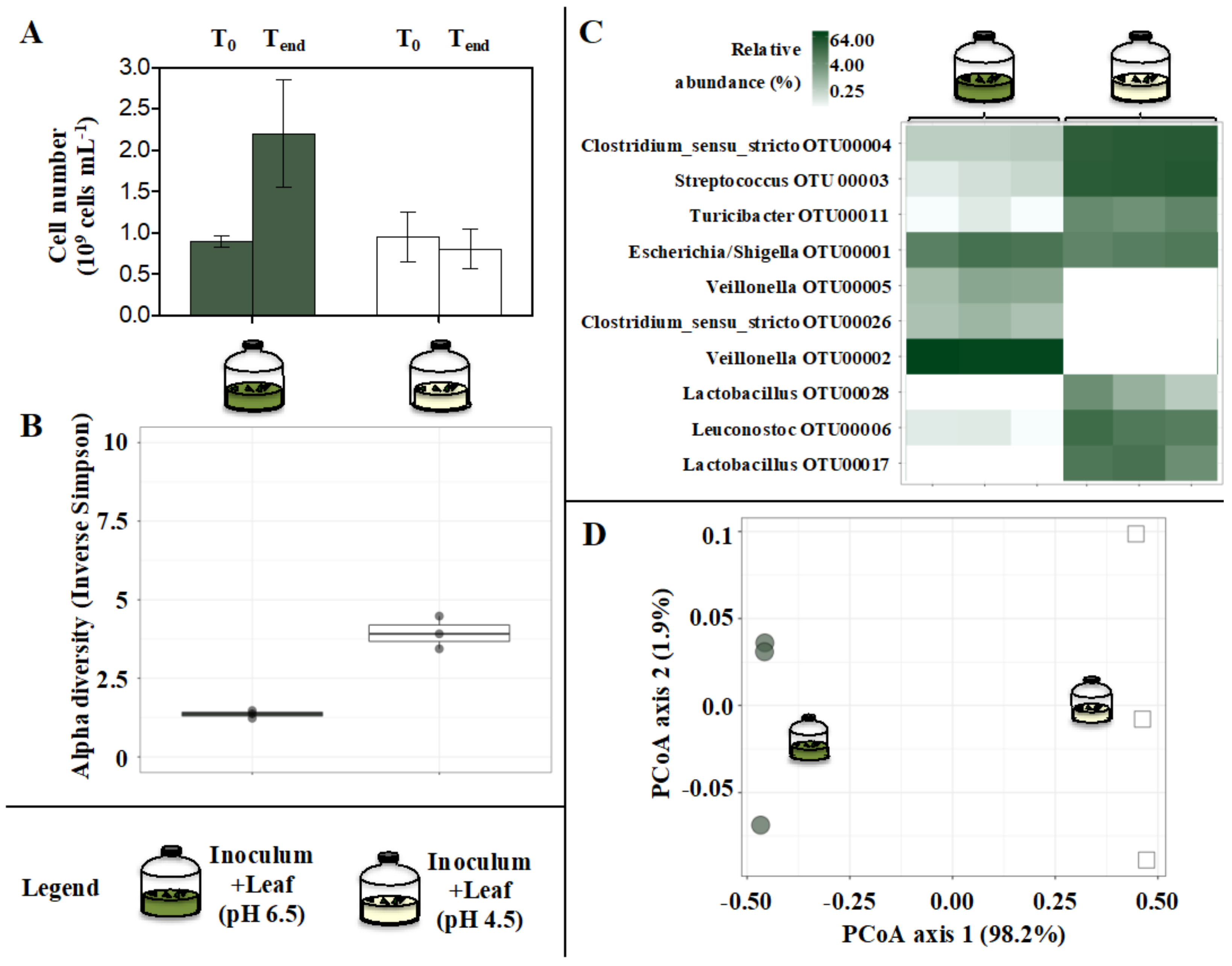
3.3. Microbial Communities
3.4. Impact of pH on Microbial Bamboo Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hungate, R.E. The Rumen and Its Microbes; Elsevier: Amsterdam, The Netherlands, 1966. [Google Scholar]
- Wildt, D.E.; Zhang, A.; Zhang, H.; Janssen, D.L.; Ellis, S. Giant Pandas—Biology, Veterinary Medicine and Management; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Dierenfeld, E.S.; Hintz, H.F.; Robertson, J.B.; Van Soest, P.J.; Oftedalt, A.T. Utilization of bamboo by the giant panda. J. Nutr. 1982, 112, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Mainka, S.A.; Guanlu, Z.; Mao, L. Utilization of a bamboo, sugar cane, and gruel diet by two juvenile giant pandas (Ailuropoda melanoleuca). J. Zoo Wildlife Med. 1989, 20, 39–44. [Google Scholar]
- Hansen, R.L.; Carr, M.M.; Apanavicius, C.J.; Jiang, P.; Bissell, H.A.; Gocinski, B.L.; Maury, F.; Himmelreich, M.; Beard, S.; Ouellette, J.R.; et al. Seasonal shifts in giant panda feeding behavior: Relationships to bamboo plant part consumption. Zoo Biol. 2010, 29, 470–483. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.L.; Willard, S.; Kouba, A.; Sparks, D.; Holmes, W.; Falcone, J.; Williams, C.H.; Brown, A. Dietary shifts affect the gastrointestinal microflora of the giant panda (Ailuropoda melanoleuca). J. Anim. Physiol. Anim. Nutr. 2013, 97, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Knott, K.K.; Christian, A.L.; Falcone, J.F.; Vance, C.K.; Bauer, L.L.; Fahey, G.C., Jr.; Kouba, A.J. Phenological changes in bamboo carbohydrates explain the preference for culm over leaves by giant pandas (Ailuropoda melanoleuca) during spring. PLoS ONE 2017, 12, e0177582. [Google Scholar] [CrossRef] [Green Version]
- Schaller, G.; Hu, J.; Pan, W.; Zhu, J. The Giant Pandas of Wolong; University of Chicago Press: Chicago, IL, USA, 1985. [Google Scholar]
- Taylor, A.H.; Zisheng, Q. Culm dynamics and dry matter production of bamboos in the Wolong and Tangjiahe giant panda reserves, Sichuan, China. J. Appl. Ecol. 1987, 24, 419–433. [Google Scholar] [CrossRef]
- Rybiski Tarou, L.; Williams, J.; Powell, D.M.; Tabet, R.; Allen, M. Behavioral preferences for bamboo in a pair of captive giant pandas (Ailuropoda melanoleuca). Zoo Biol. 2005, 24, 177–183. [Google Scholar] [CrossRef]
- Senshu, T.; Ohya, A.; Ide, K.; Mikogai, J.; Morita, M.; Nakao, T.; Imazu, K.; Lan, J.; Liu, X.; Lu, W.; et al. Studies on the digestion in the giant panda, Ailuropoda melanoleuca, fed feedstuffs including bamboo. Mammal Study 2007, 32, 139–149. [Google Scholar] [CrossRef]
- Sims, J.A.; Parsons, J.L.; Bissell, H.A.; Sikes, R.S.; Ouellette, J.R.; Rude, B.J. Determination of bamboo-diet digestibility and fecal output by giant pandas. Ursus 2007, 18, 38–45. [Google Scholar] [CrossRef]
- Finley, T.G.; Sikes, R.S.; Parsons, J.L.; Rude, B.J.; Bissell, H.A.; Ouellette, J.R. Energy digestibility of giant pandas on bamboo-only and on supplemented diets. Zoo Biol. 2011, 30, 121–133. [Google Scholar]
- Dierenfeld, E.S. Chemical composition of bamboo in relation to giant panda nutrition. In The Bamboos; Chapman, G.P., Ed.; Academic Press: London, UK, 1997; pp. 205–211. [Google Scholar]
- Li, R.; Fan, W.; Tian, G.; Zhu, H.; He, L.; Cai, J.; Huang, Q.; Cai, Q.; Li, B.; Bai, Y.; et al. The sequence and de novo assembly of the giant panda genome. Nature 2010, 463, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.; Wei, F.; Zhou, W.; Hu, Y.; Senior, A.M.; Wu, Q.; Li, Y.; Raubenheimer, D. Giant pandas are macronutritional carnivores. Curr. Biol. 2019, 29, 1677–1682. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Zhang, W.; Wang, L.; Hou, R.; Zhang, M.; Fei, L.; Zhang, X.; Huang, H.; Bridgewater, L.C.; Jiang, Y.; et al. The bamboo-eating giant panda harbors a carnivore-like gut microbiota, with excessive seasonal variations. mBio 2015, 6, e00022-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helander, M.; Jia, R.; Huitu, O.; Sieber, T.N.; Jia, J.; Niemelä, P.; Saikkonen, K. Endophytic fungi and silica content of different bamboo species in giant panda diet. Symbiosis 2013, 61, 13–22. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, X.; Ding, Y.; Hu, Y.; Nie, Y.; Wei, W.; Ma, S.; Li, Y.; Zhu, L.; Wei, F. Seasonal variation in nutrient utilization shapes gut microbiome structure and function in wild giant pandas. Proc. R. Soc. B Biol. Sci. 2017, 284, 1862. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, W.; Han, S.; Kong, F.; Wang, C.; Li, D.; Zhang, H.; Yang, M.; Xu, H.; Zeng, B.; et al. The evolution of the gut microbiota in the giant and the red pandas. Sci. Rep. 2015, 5, 10185. [Google Scholar] [CrossRef] [Green Version]
- Wei, G.; Lu, H.; Zhou, Z.; Xie, H.; Wang, A.; Nelson, K.; Zhao, L. The microbial community in the feces of the giant panda (Ailuropoda melanoleuca) as determined by PCR-TGGE profiling and clone library analysis. Microb. Ecol. 2007, 54, 194–202. [Google Scholar] [CrossRef]
- Tun, H.M.; Mauroo, N.F.; Yuen, C.S.; Ho, J.C.; Wong, M.T.; Leung, F.C. Microbial diversity and evidence of novel homoacetogens in the gut of both geriatric and adult giant pandas (Ailuropoda melanoleuca). PLoS ONE 2014, 9, e79902. [Google Scholar] [CrossRef]
- Peng, Z.; Zeng, D.; Wang, Q.; Niu, L.; Ni, X.; Zou, F.; Yang, M.; Sun, H.; Zhou, Y.; Liu, Q.; et al. Decreased microbial diversity and Lactobacillus group in the intestine of geriatric giant pandas (Ailuropoda melanoleuca). World J. Microbiol. Biotechnol. 2016, 32, 79. [Google Scholar] [CrossRef]
- Zhu, L.; Wu, Q.; Dai, J.; Zhang, S.; Wei, F. Evidence of cellulose metabolism by the giant panda gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 17714–17719. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Fang, Z.; Zhou, P.; Chang, F.; Hong, Y.; Zhang, X.; Peng, H.; Xiao, Y. Evidence for lignin oxidation by the giant panda fecal microbiome. PLoS ONE 2012, 7, e50312. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Mishra, S.; Zhao, J.; Tang, J.; Zeng, B.; Kong, F.; Ning, R.; Li, M.; Zhang, H.; Zeng, Y.; et al. Metagenomic Study Suggests That the Gut Microbiota of the Giant Panda (Ailuropoda melanoleuca) May Not Be Specialized for Fiber Fermentation. Front. Microbiol. 2018, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, W.; Hou, R.; Zhang, L.; Schmitz-Esser, S.; Sun, H.; Xie, J.; Zhang, Y.; Wang, C.; Li, L.; et al. Age-associated microbiome shows the giant panda lives on hemicelluloses, not on cellulose. ISME J. 2018, 12, 1319–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Fei, L.; Li, F.; Niu, L.; Zhang, Z. Analysis of the digestive enzyme activities in the digestive tract of giant pandas. Sichuan J. Zool. 2009, 28, 397–400. [Google Scholar]
- Wang, H.; Zhong, H.; Hou, R.; Ayala, J.; Liu, G.; Yuan, S.; Yan, Z.; Zhang, W.; Liu, Y.; Cai, K.; et al. A Diet Diverse in Bamboo Parts is Important for Giant Panda (Ailuropoda melanoleuca) Metabolism and Health. Sci. Rep. 2017, 7, 3377. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yin, Y.; Chen, X.; Chen, C.; Xia, Y.; Qi, H.; Baker, P.N.; Zhang, H.; Han, T.-L. Evaluating different extraction solvents for GC-MS based metabolomic analysis of the fecal metabolome of adult and baby giant pandas. Sci. Rep. 2019, 9, 12017. [Google Scholar] [CrossRef]
- Zhu, C.; Laghi, L.; Zhang, Z.; He, Y.; Wu, D.; Zhang, H.; Huang, Y.; Li, C.; Zou, L. First Steps toward the Giant Panda Metabolome Database: Untargeted Metabolomics of Feces, Urine, Serum, and Saliva by (1)H NMR. J. Proteome Res. 2020, 19, 1052–1059. [Google Scholar] [CrossRef]
- Scoma, A.; Khor, W.C.; Coma, M.; Heyer, R.; Props, R.; Schoelynck, J.; Bouts, T.; Benndorf, D.; Li, D.; Zhang, H.; et al. Substrate-Dependent Fermentation of Bamboo in Giant Panda Gut Microbiomes: Leaf Primarily to Ethanol and Pith to Lactate. Front. Microbiol. 2020, 11, 530. [Google Scholar] [CrossRef]
- Heyer, R.; Kohrs, F.; Benndorf, D.; Rapp, E.; Kausmann, R.; Heiermann, M.; Klocke, M.; Reichl, U. Metaproteome analysis of the microbial communities in agricultural biogas plants. New Biotechnol. 2013, 30, 614–622. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.C.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; pp. 1–3871. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggerth, A.H.; Gagnon, B.H. The Bacteroides of Human Feces. J. Bacteriol. 1933, 25, 389–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ose, R.; Hirano, K.; Maeno, S.; Nakagawa, J.; Salminen, S.; Tochio, T.; Endo, A. The ability of human intestinal anaerobes to metabolize different oligosaccharides: Novel means for microbiota modulation? Anaerobe 2018, 51, 110–119. [Google Scholar] [CrossRef]
- Scholten-Koerselman, I.; Houwaard, F.; Janssen, P.; Zehnder, A.J. Bacteroides xylanolyticus sp. nov., a xylanolytic bacterium from methane producing cattle manure. Antonie Van Leeuwenhoek 1986, 52, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Shah, H.N.; Collins, M.D. Genus Bacteroides. A chemotaxonomical perspective. J. Appl. Bacteriol. 1983, 55, 403–416. [Google Scholar] [CrossRef]
- Ishikawa, K.; Matsui, I.; Honda, K.; Nakatani, H. Substrate-dependent shift of optimum pH in porcine pancreatic alpha-amylase-catalyzed reactions. Biochemistry 1990, 29, 7119–7123. [Google Scholar] [CrossRef]
- Ishikawa, K.; Matsui, I.; Honda, K.; Kobayashi, S.; Nakatani, H. The pH dependence of the action pattern in porcine pancreatic alpha-amylase-catalyzed reaction for maltooligosaccharide substrates. Arch. Biochem. Biophys. 1991, 289, 124–129. [Google Scholar] [CrossRef]
- Funke, I.; Melzig, M.F. Effect of different phenolic compounds on a-amylase activity: Screening by microplate-reader based kinetic assay. Pharmazie 2005, 60, 796–797. [Google Scholar]
- Keski-Saari, S.; Ossipov, V.; Julkunen-Tiitto, R.; Jia, J.; Danell, K.; Veteli, T.; Guiquan, Z.; Yaowu, X.; Niemelä, P. Phenolics from the culms of five bamboo species in the Tangjiahe and Wolong Giant Panda Reserves, Sichuan, China. Biochem. Syst. Ecol. 2008, 36, 758–765. [Google Scholar] [CrossRef]
- Williams, C.L.; Dill-McFarland, K.A.; Vandewege, M.W.; Sparks, D.L.; Willard, S.T.; Kouba, A.J.; Suen, G.; Brown, A.E. Dietary Shifts May Trigger Dysbiosis and Mucous Stools in Giant Pandas (Ailuropoda melanoleuca). Front. Microbiol. 2016, 7, 661. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Neuhoff, V.; Stamm, R.; Eibl, H. Clear Background and Highly Sensitive Protein Staining with Coomassie Blue Dyes in Polyacrylamide Gels—A Systematic Analysis. Electrophoresis 1985, 6, 427–448. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scoma, A.; Khor, W.C.; Coma, M.; Heyer, R.; Props, R.; Bouts, T.; Benndorf, D.; Li, D.; Zhang, H.; Rabaey, K. Lignocellulose Fermentation Products Generated by Giant Panda Gut Microbiomes Depend Ultimately on pH Rather than Portion of Bamboo: A Preliminary Study. Microorganisms 2022, 10, 978. https://doi.org/10.3390/microorganisms10050978
Scoma A, Khor WC, Coma M, Heyer R, Props R, Bouts T, Benndorf D, Li D, Zhang H, Rabaey K. Lignocellulose Fermentation Products Generated by Giant Panda Gut Microbiomes Depend Ultimately on pH Rather than Portion of Bamboo: A Preliminary Study. Microorganisms. 2022; 10(5):978. https://doi.org/10.3390/microorganisms10050978
Chicago/Turabian StyleScoma, Alberto, Way Cern Khor, Marta Coma, Robert Heyer, Ruben Props, Tim Bouts, Dirk Benndorf, Desheng Li, Hemin Zhang, and Korneel Rabaey. 2022. "Lignocellulose Fermentation Products Generated by Giant Panda Gut Microbiomes Depend Ultimately on pH Rather than Portion of Bamboo: A Preliminary Study" Microorganisms 10, no. 5: 978. https://doi.org/10.3390/microorganisms10050978