
Deciphering the Microbiota and Volatile Profiles of Algerian Smen, a Traditional Fermented Butter

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.1.1. Collected Smen Samples
2.1.2. Dynamic Follow-Up of a Smen Preparation
2.2. Microbiological Analyses
2.2.1. Safety and Hygienic Quality
2.2.2. Enumeration and Isolation of Lactic Acid Bacteria and Yeasts
2.2.3. Identification of Lactic Acid Bacteria and Yeast Isolates
2.3. Metataxonomic Analysis of Bacterial and Fungal Communities
2.3.1. Sample Preparation and DNA Extraction
2.3.2. PCR and Metagenetic Sequencing Conditions
2.3.3. Bioinformatic and Phylogenetic Analysis
2.3.4. Biodiversity and Statistical Analysis
2.4. Physicochemical Analysis
2.5. Correlation Analyses between Microbial Diversity and Volatile Profiles
3. Results
3.1. Microbial Diversity and Biochemical Profiles of Smen Samples Produced in Algerian Households
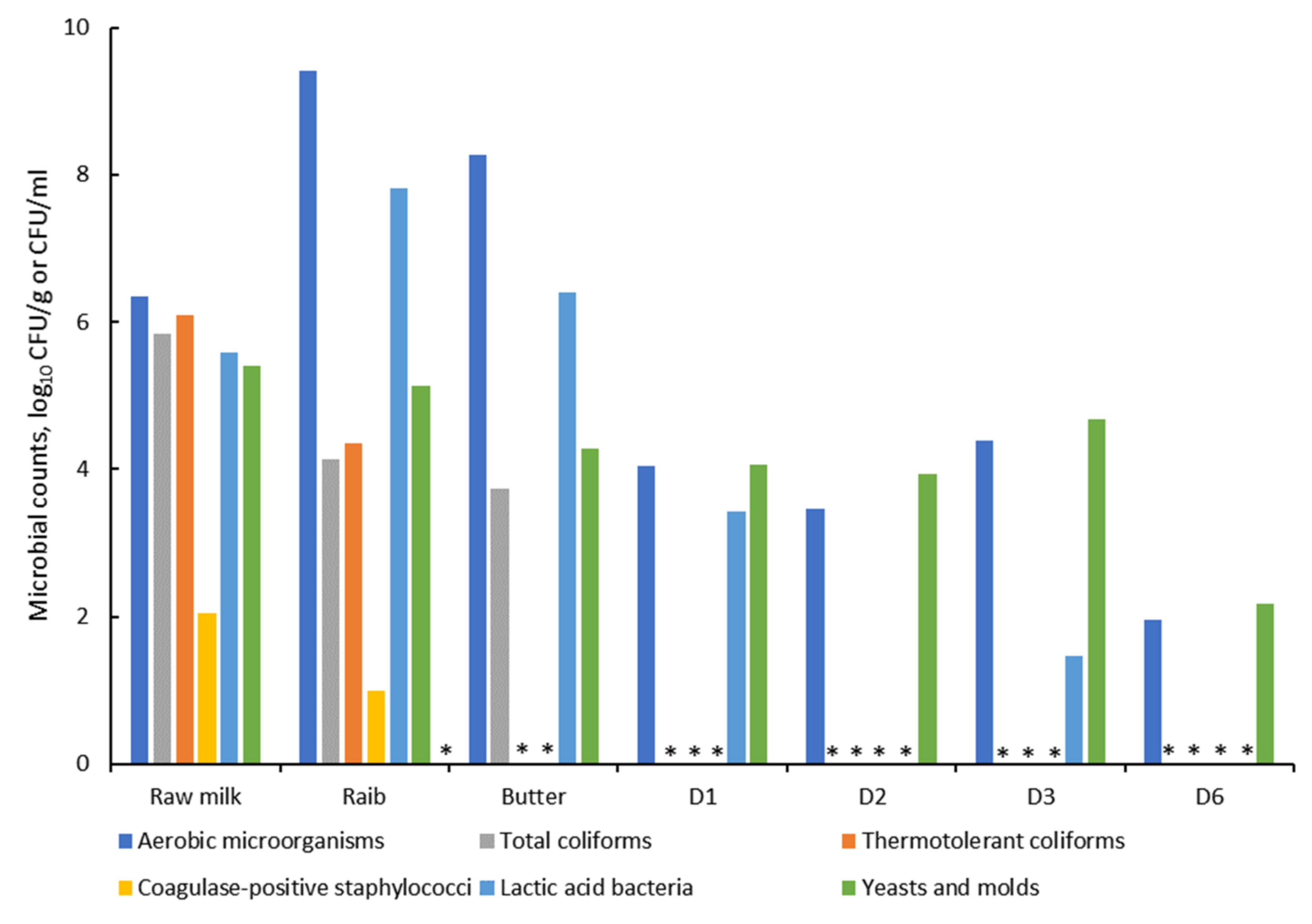
3.1.1. Safety and Hygienic Quality
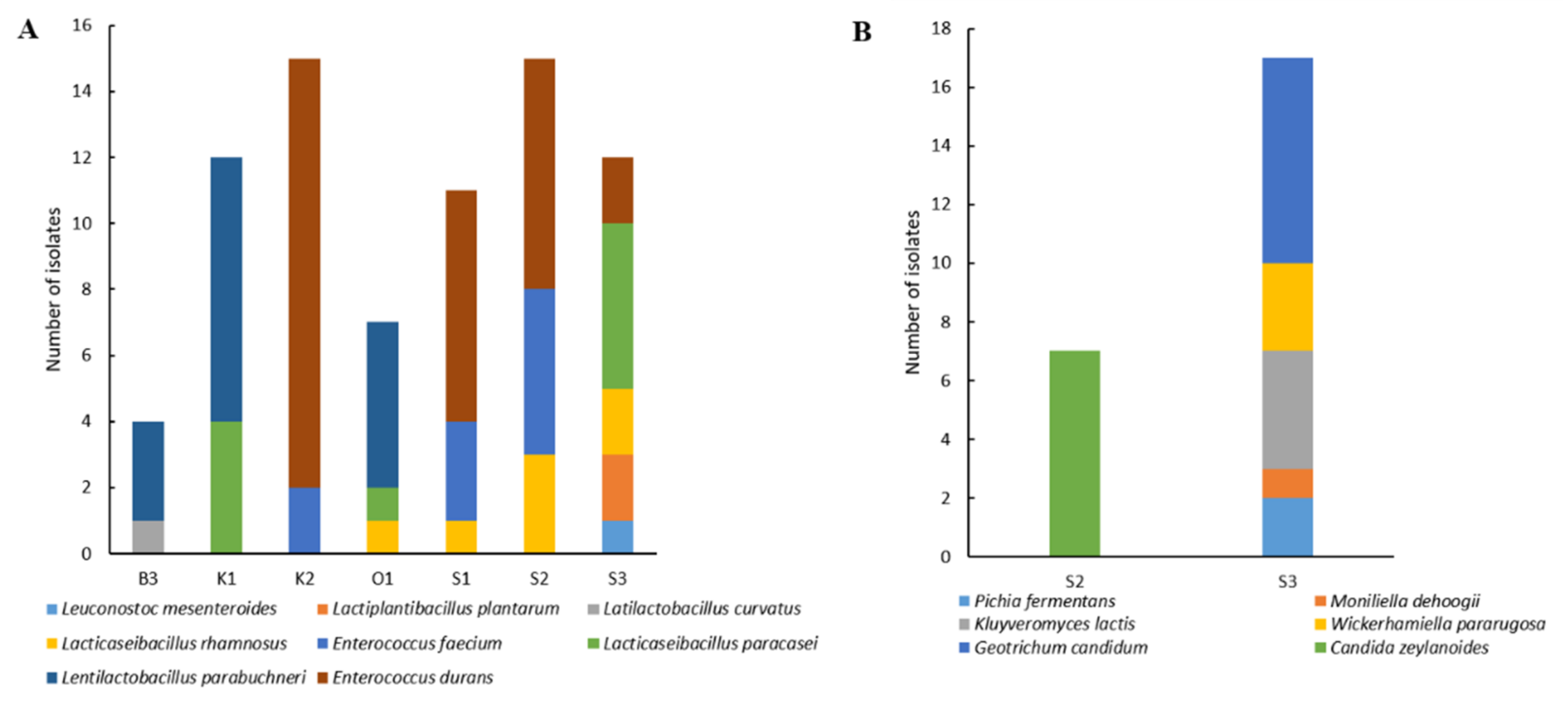
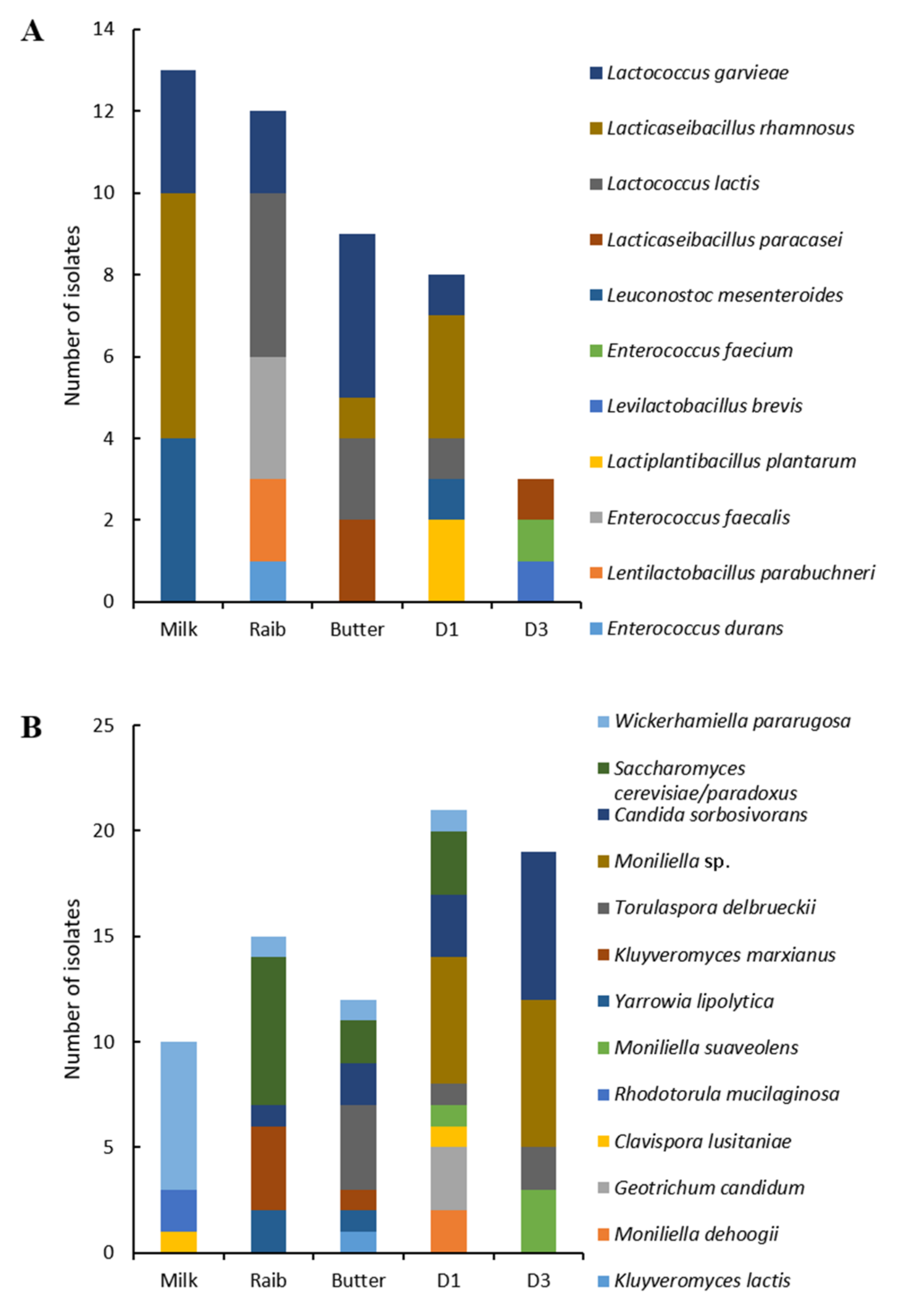
3.1.2. Lactic Acid Bacteria and Yeast Diversity Using Culture-Dependent Analysis
3.1.3. Microbial Diversity Using Metagenetic Analyses
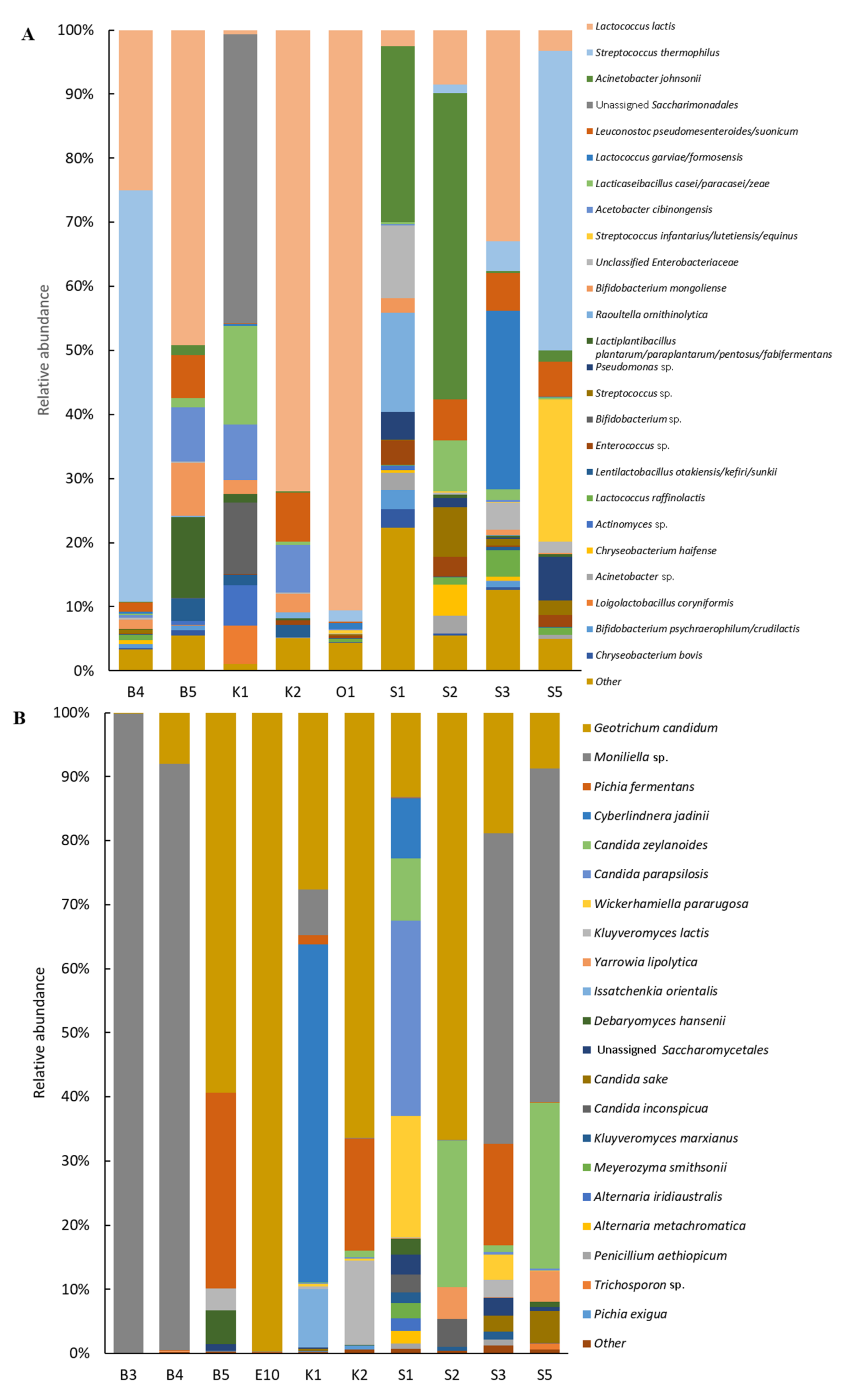
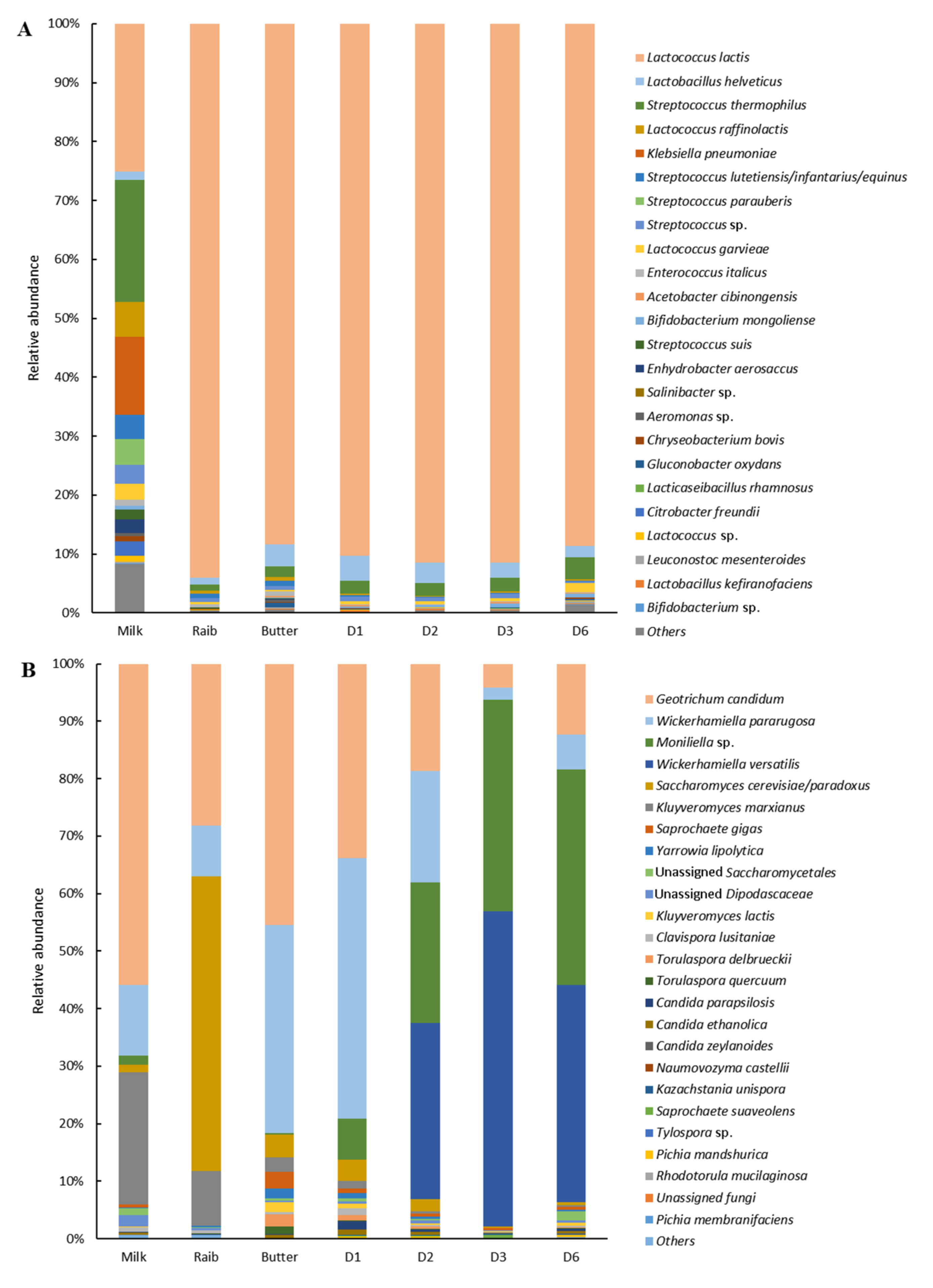
Bacterial Diversity
Fungal Diversity
3.1.4. Physico-Chemical Analysis and Volatile Profiles
3.2. Microbial Dynamics and Diversity and Biochemical Profiles throughout Different Preparation Stages of Smen
3.2.1. Safety and Hygienic Quality
3.2.2. Lactic Acid Bacteria and Yeast Diversity Using Culture-Dependent Analysis
3.2.3. Microbial Diversity Using Culture-Independent Analysis
Bacterial Diversity Using V3–V4 Metabarcoding Analysis
Fungal Diversity Using ITS2 Metabarcoding Analysis
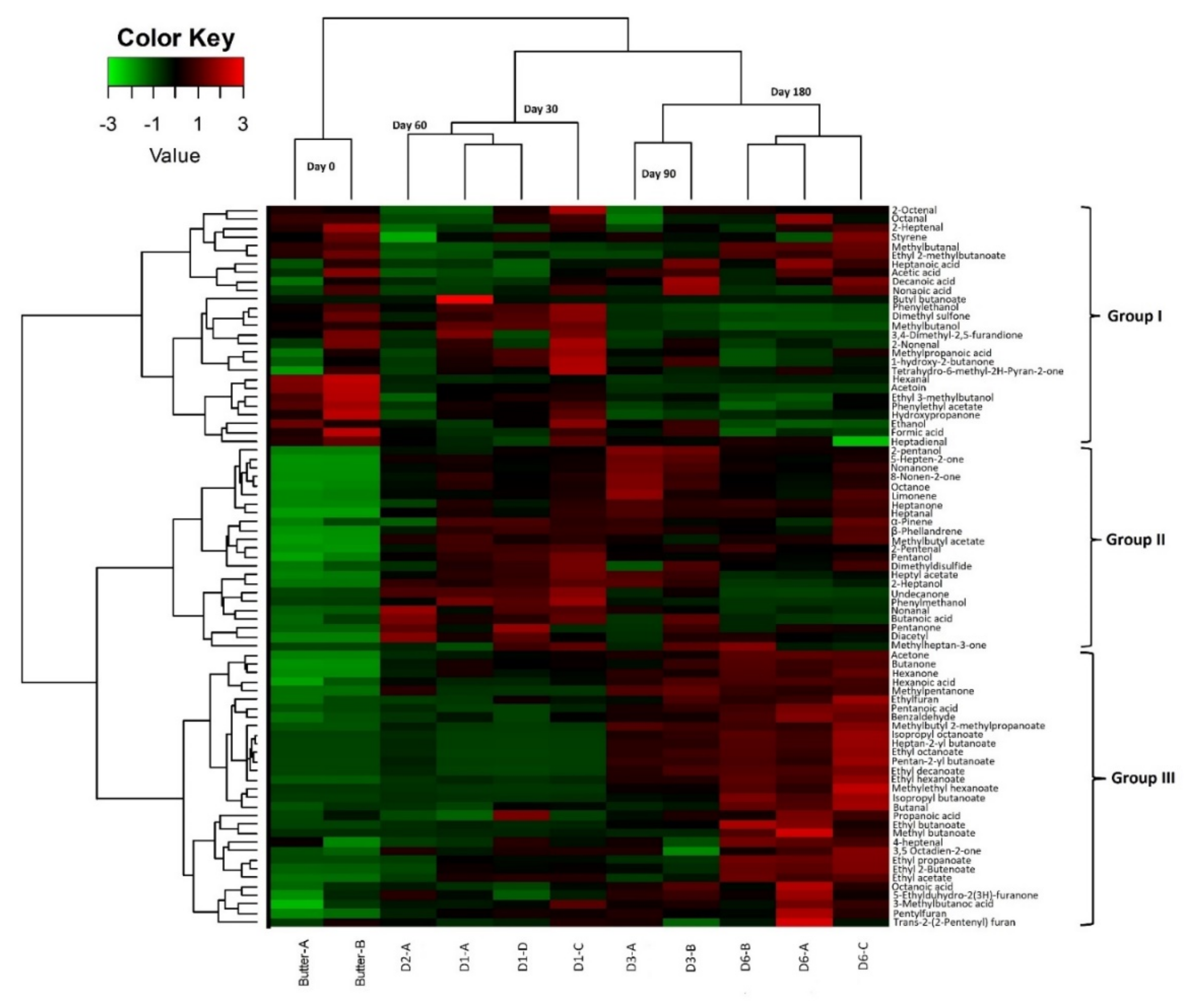
3.2.4. Physico-Chemical Analysis and Volatile Profiles
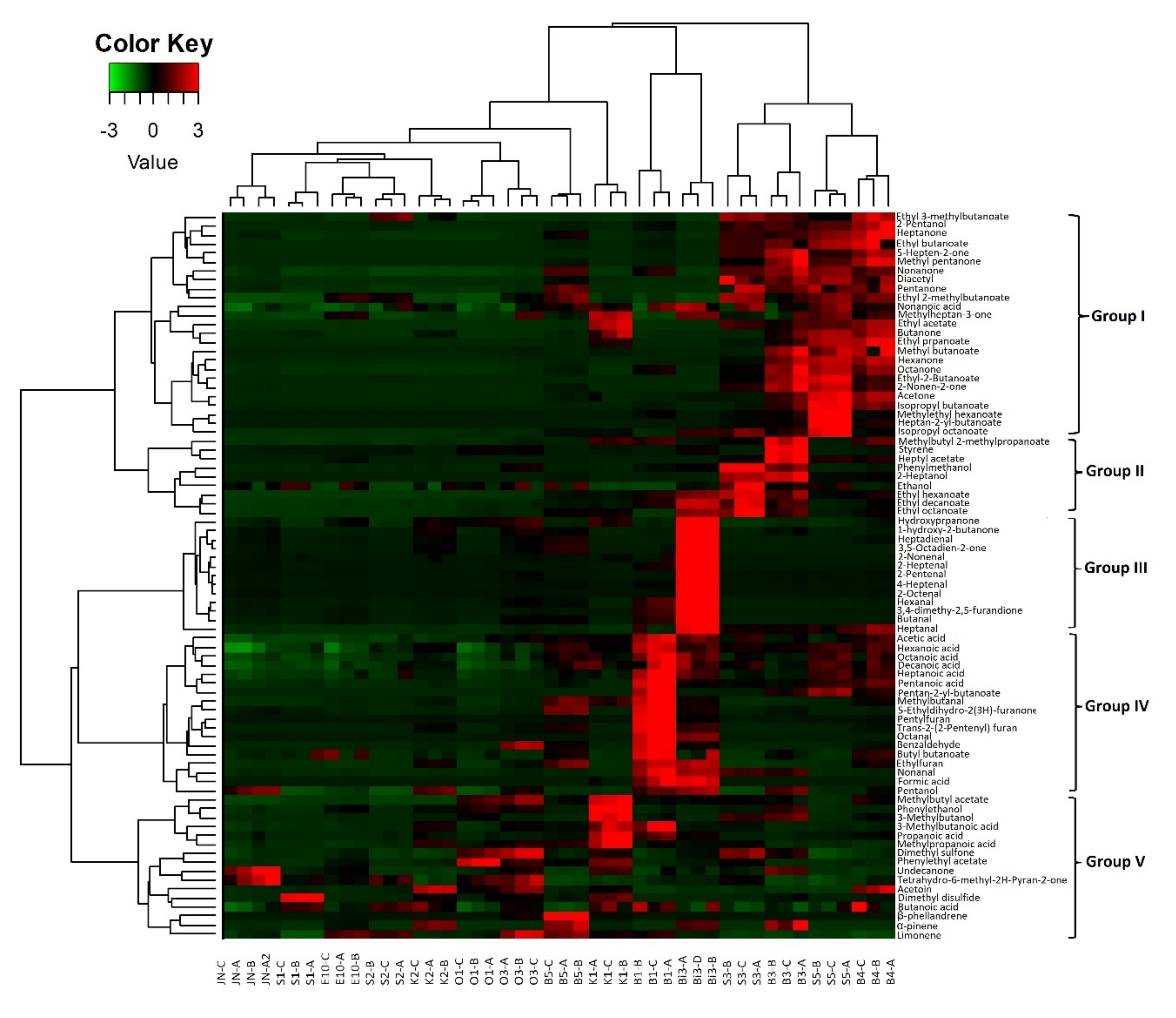
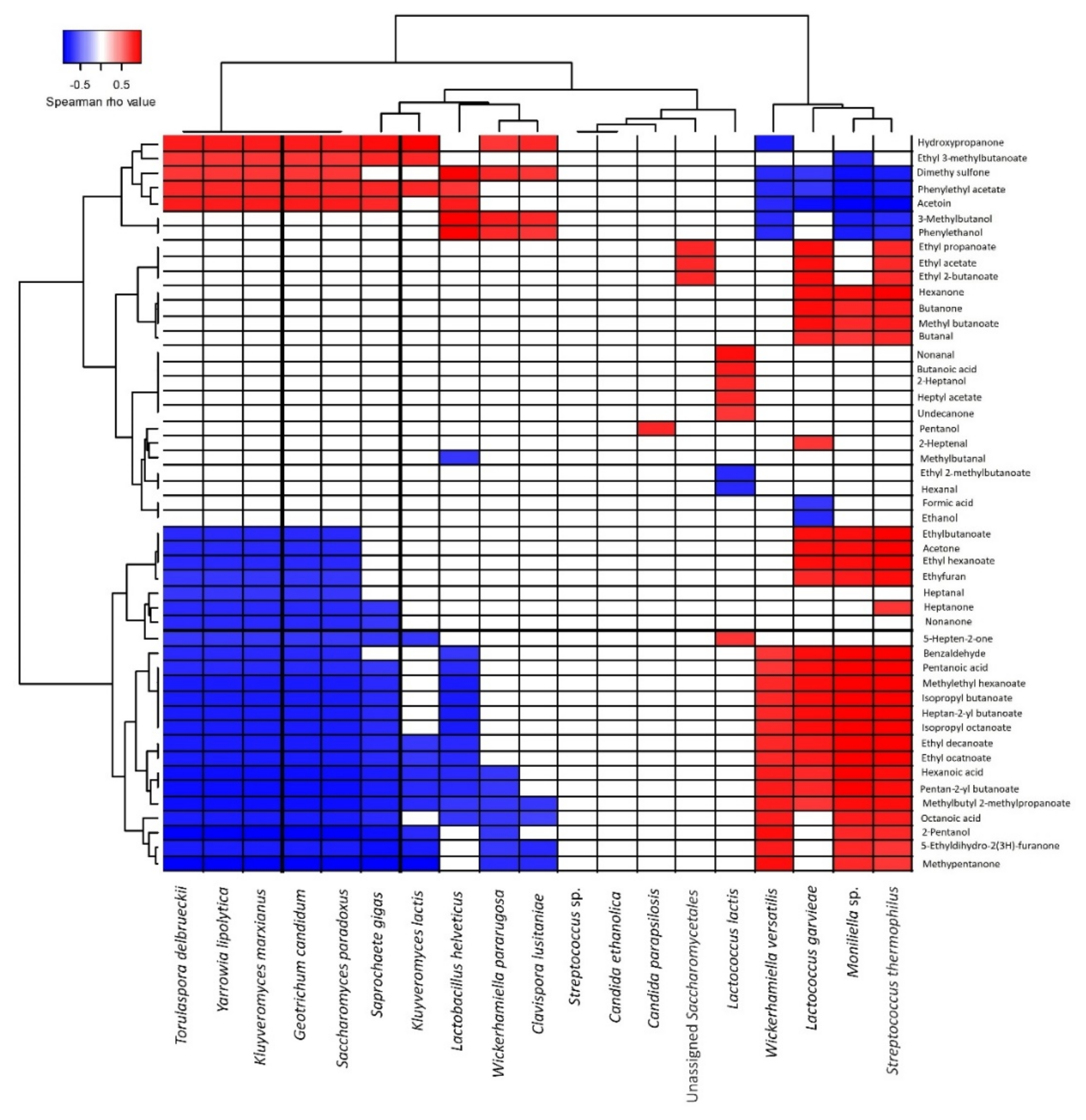
3.2.5. Correlation between Volatile Profiles and Microbiota during Maturation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tamang, J.P.; Kailasapathy, K. Fermented Foods and Beverages of the World; CRC Press, Taylor and Francis Group: New York, NY, USA, 2010. [Google Scholar]
- El Sheikha, A.F.; Hu, D.-M. Molecular techniques reveal more secrets of fermented foods. Crit. Rev. Food Sci. Nutr. 2018, 60, 11–32. [Google Scholar] [CrossRef] [PubMed]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Kasase, C.; Toarta, C.; Kassen, R.; Poulain, A.J. Microbial Community Structure of Three Traditional Zambian Fermented Products: Mabisi, Chibwantu and Munkoyo. PLoS ONE 2013, 8, e63948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraffa, G. Studying the dynamics of microbial populations during food fermentation. FEMS Microbiol. Rev. 2004, 28, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocolin, L.; Ercolini, D. Zooming into Food-Associated Microbial Consortia: A ‘Cultural’ Evolution. Curr. Opin. Food Sci. 2015, 2, 43–50. [Google Scholar] [CrossRef]
- Tamang, J.P.; Thapa, N.; Tamang, B.; Rai, A.; Chettri, R. Microorganisms in Fermented Foods and Beverages. In Health Benefits of Fermented Foods and Beverages; Tamang, J.P., Ed.; CRC Press, Taylor and Francis Group: New York, NY, USA, 2015; pp. 2–87. [Google Scholar]
- Diaz, M.; Kellingray, L.; Akinyemi, N.; Adefiranye, O.O.; Olaonipekun, A.B.; Bayili, G.R.; Ibezim, J.; Du Plessis, A.S.; Houngbédji, M.; Kamya, D.; et al. Comparison of the microbial composition of African fermented foods using amplicon sequencing. Sci. Rep. 2019, 9, 13863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wafula, E.N.; Franz, C.M.; Rohn, S.; Huch, M.; Mathara, J.M.; Trierweiler, B. Fermentation of African indigenous leafy vegetables to lower post-harvest losses, maintain quality and increase product safety. Afr. J. Hortic. Sci. 2016, 9, 1–13. [Google Scholar]
- Benkerroum, N. Traditional Fermented Foods of North African Countries: Technology and Food Safety Challenges with Regard to Microbiological Risks. Compr. Rev. Food Sci. Food Saf. 2013, 12, 54–89. [Google Scholar] [CrossRef]
- Boussekine, R.; Merabti, R.; Barkat, M.; Becila, F.-Z.; Belhoula, N.; Mounier, J.; Bekhouche, F. Traditional Fermented Butter Smen/Dhan: Current Knowledge, Production and Consumption in Algeria. J. Food Res. 2020, 9, 71–82. [Google Scholar] [CrossRef]
- Iradukunda, C.; Aida, W.M.W.; Ouafi, A.T.; Barkouch, Y.; Boussaid, A. Aroma profile of a traditionally fermented butter (smen). J. Dairy Res. 2018, 85, 114–120. [Google Scholar] [CrossRef]
- El Marrakchi, A.; Tantaoui-Elarraki, A.; El Mane, A.; Tifrit, L. La flore microbienne du smen marocain I. Flore naturelle et flore d’intérêt hygiénique. Le Lait 1988, 68, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Triqui, R.; Guth, H. Potent odorants in “Smen”, a traditional fermented butter product. Eur. Food Res. Technol. 2001, 212, 292–295. [Google Scholar] [CrossRef]
- Bensalah, F.; Labtar, A.; Delorme, C.; Renault, P. Occurrence, isolation and DNA identification of Streptococcus thermophilus involved in algerian traditional butter ‘Smen’. Afr. J. Biotechnol. 2011, 10, 17251–17257. [Google Scholar] [CrossRef]
- Bettache, G.; Fatma, A.; Miloud, H.; Mebrouk, K. Isolation and identification of lactic acid bacteria from Dhan, a traditional butter and their major technological traits. World Appl. Sci. J. 2012, 17, 480–488. [Google Scholar]
- Labtar, A.; Larouci, S.; Guermouche, A.; Bensalah, F. Study on molecular identification of lactic acid bacteria from fermented milks and" Smen"(a traditional steppe butter) and their enzyme producing attributes. SAJEB 2019, 9, 95–103. [Google Scholar] [CrossRef]
- Ongol, M.P.; Asano, K. Main microorganisms involved in the fermentation of Ugandan ghee. Int. J. Food Microbiol. 2009, 133, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Bezuidt, O.; Gomri, M.A.; Pierneef, R.; Van Goethem, M.; Kharroub, K.; Cowan, D.; Makhalanyane, T.P. Draft genome sequence of Thermoactinomyces sp. strain AS95 isolated from a Sebkha in Thamelaht, Algeria. Stand. Genom. Sci. 2016, 11, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Idoui, T.; Karam, N.E. Lactic acid bacteria from Jijel’s traditional butter: Isolation, identification and major technological traits. Grasas Aceites 2008, 59, 361–367. [Google Scholar] [CrossRef]
- ISO 4833:2003; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Microorganisms—Colony-Count Technique at 30 Degrees C. ISO: Geneva, Switzerland. Available online: https://www.iso.org/standard/34524.html (accessed on 21 March 2021).
- ISO 4832:2006; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Coliforms—Colony-Count Technique. ISO: Geneva, Switzerland. Available online: https://www.iso.org/standard/38280.html (accessed on 21 March 2021).
- ISO 7937:2004; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Clostridium Perfringens—Colony-Count Technique. ISO: Geneva, Switzerland. Available online: https://standards.iteh.ai/catalog/standards/cen/4f03b11b-ede0-4640-ad00-7169594d489b/en-iso-7937-2004 (accessed on 21 March 2021).
- ISO 6888-1:1999; Microbiology of Food and Animal Feeding Stuffs Horizontal Method for the Enumeration of Coagulase Positive Staphylococci (Staphylococcus aureus and Other Species)—Part 1: Technique Using Baird-Parker Agar Medium. ISO: Geneva, Switzerland. Available online: https://www.iso.org/standard/23036.html (accessed on 21 March 2021).
- Salmonella, A.G. Food Microbiology Protocols; Spencer, J.F.T., Ragout de Spencer, A.L., Eds.; Humana Press Inc.: Totowa, NJ, USA, 2001; pp. 113–118. [Google Scholar]
- Guinebretiere, M.H.; Nguyen-The, C. Sources of Bacillus cereus contamination in a pasteurized zucchini puree processing line, differentiated by two PCR-based methods. FEMS Microbiol. Ecol. 2003, 43, 207–215. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification of clinically important ascomycetous yeasts based on nucleotide divergence in the 5’ end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Penland, M.; Falentin, H.; Parayre, S.; Pawtowski, A.; Maillard, M.-B.; Thierry, A.; Mounier, J.; Coton, M.; Deutsch, S.-M. Linking Pélardon artisanal goat cheese microbial communities to aroma compounds during cheese-making and ripening. Int. J. Food Microbiol. 2021, 345, 109130. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewski, M.; Proietti, C.; Ellis, J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome–environment interactions. Bioinformatics 2016, 33, 782–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquot, C. IUPAC Standard Methods for the Analysis of Oils, Fats and Derivatives, 6th ed.; Pergamon Press: Oxford, UK, 1979. [Google Scholar]
- Wolff, J.P. Manuel d’Analyse des Corps Gras; Azoulay: Paris, France, 1968. [Google Scholar]
- AOAC. Official Methods of Analysis, 12th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1975. [Google Scholar]
- Pogačić, T.; Maillard, M.-B.; Leclerc, A.; Hervé, C.; Chuat, V.; Yee, A.L.; Valence, F.; Thierry, A. A methodological approach to screen diverse cheese-related bacteria for their ability to produce aroma compounds. Food Microbiol. 2015, 46, 145–153. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. Available online: http://www.jstor.org/stable/2346101 (accessed on 21 March 2022). [CrossRef]
- Lê, S.; Josse, J.; Husson, F. “FactoMineR: An R Package for Multivariate Analysis”. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Ogier, J.C.; Serror, P. Safety assessment of dairy microorganisms: The Enterococcus genus. Int. J. Food Microbiol. 2008, 126, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Daza, M.V.B.; Cortimiglia, C.; Bassi, D.; Cocconcelli, P.S. Genome-based studies indicate that the Enterococcus faecium Clade B strains belong to Enterococcus lactis species and lack of the hospital infection associated markers. Int. J. Syst. Evol. Microbiol. 2021, 71, 004948. [Google Scholar] [CrossRef]
- Sakili, D.; Isoual, D. Les Bactéries Lactiques dans l’élaboration du Smen Marocain; Académie d’Agriculture de France: Paris, France, 2003. [Google Scholar]
- Savadogo, A.; Traore, A. La flore microbienne et les propriétés fonctionnelles des yaourts et laits fermentés. Int. J. Biol. Chem. Sci. 2012, 5, 2057. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, H.; Wüthrich, D.; Bruggmann, R.; Wechsler, D.; Fröhlich-Wyder, M.-T.; Irmler, S. Development of new methods for the quantitative detection and typing of Lactobacillus parabuchneri in dairy products. Int. Dairy J. 2017, 70, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Da Rosa, P.D.; Mattanna, P.; Carboni, D.; Amorim, L.; Richards, N.; Valente, P. Candida zeylanoides as a new yeast model for lipid metabolism studies: Effect of nitrogen sources on fatty acid accumulation. Folia Microbiol. 2014, 59, 477–484. [Google Scholar] [CrossRef]
- Čanak, I.; Berkics, A.; Bajcsi, N.; Kovacs, M.; Belak, A.; Teparić, R.; Maraz, A.; Mrša, V. Purification and Characterization of a Novel Cold-Active Lipase from the Yeast Candida zeylanoides. Microb. Physiol. 2015, 25, 403–411. [Google Scholar] [CrossRef]
- Tantaoui-Elaraki, A.; El Marrakchi, A. Study of Moroccan Dairy Products: Lben and Smen. Mircen J. 1987, 3, 211–220. [Google Scholar] [CrossRef]
- Abd-El-Malek, Y. Traditional Egyptian dairy fermentations. In Global Impact of Food Microbiology (GIAM). State of the Art: GIAM and Its Relevance to Development in Developing Countries; Stantan, W.R., Da Silva, E.J., Eds.; University of Malaya Press: Kuala Lumpur, Malaysia, 1978; pp. 198–208. [Google Scholar]
- Ben Amor, K.; Cornelieus, C.; Mahjoub, A.; Thonart, P. Identification de La Flore Lactique Du Lait Fermenté Traditionnel Tunisien (Lben) et Évaluation Des Composés Aromatisants. Microb. Hyg. Alim. 1998, 10, 31–36. [Google Scholar]
- Hamama, A.; Ha, D. Improvements of the manufacture of traditional fermented products in Morocco: Case of jben (moroccan traditional fresh cheese). In Emerging Technology Series, Food Processing Technologies for Africa; UNIDO: Vienna, Austria, 1997; pp. 85–102. [Google Scholar]
- Schmidt, V.S.; Kaufmann, V.; Kulozik, U.; Scherer, S.; Wenning, M. Microbial biodiversity, quality and shelf life of microfiltered and pasteurized extended shelf life (ESL) milk from Germany, Austria and Switzerland. Int. J. Food Microbiol. 2012, 154, 1–9. [Google Scholar] [CrossRef]
- Delavenne, E.; Mounier, J.; Déniel, F.; Barbier, G.; Le Blay, G. Biodiversity of antifungal lactic acid bacteria isolated from raw milk samples from cow, ewe and goat over one-year period. Int. J. Food Microbiol. 2012, 155, 185–190. [Google Scholar] [CrossRef]
- Wouters, J.T.; Ayad, E.H.; Hugenholtz, J.; Smit, G. Microbes from raw milk for fermented dairy products. Int. Dairy J. 2002, 12, 91–109. [Google Scholar] [CrossRef]
- Gardini, F.; Tofalo, R.; Belletti, N.; Iucci, L.; Suzzi, G.; Torriani, S.; Guerzoni, M.; Lanciotti, R. Characterization of yeasts involved in the ripening of Pecorino Crotonese cheese. Food Microbiol. 2006, 23, 641–648. [Google Scholar] [CrossRef]
- Sessou, P.; Keisam, S.; Tuikhar, N.; Gagara, M.; Farougou, S.; Jeyaram, K. High-Throughput Illumina MiSeq amplicon sequencing of yeast communities associated with indigenous dairy products from republics of benin and niger. Front. Microbiol. 2019, 10, 594. [Google Scholar] [CrossRef]
- Rosa, C.A.; Jindamorakot, S.; Limtong, S.; Nakase, T.; Lachance, M.-A.; Fidalgo-Jiménez, A.; Daniel, H.-M.; Pagnocca, F.C.; Inácio, J.; Morais, P.B. Synonymy of the yeast genera Moniliella and Trichosporonoides and proposal of Moniliella fonsecae sp. nov. and five new species combinations. Int. J. Syst. Evol. Microbiol. 2009, 59, 425–429. [Google Scholar] [CrossRef]
- Nakayama, M.; Hosoya, K.; Shimizu-Imanishi, Y.; Chibana, H.; Yaguchi, T. Development of Rapid Identification and Risk Analysis of Moniliella spp. in Acidic Processed Foods. Biocontrol Sci. 2016, 21, 73–80. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, G.S. Taxonomic review of Moniliella, Trichosporonoides and Hyalodendron. In The Black Yeasts, II: Moniliella and Allied Genera; Institute of the Royal Netherlands: Amsterdam, The Netherlands, 1979; pp. 1–36. [Google Scholar]
- Sarhir, S.T.; Amanpour, A.; Bouseta, A.; Selli, S. Fingerprint of aroma-active compounds and odor activity values in a traditional Moroccan fermented butter “Smen” using GC–MS–Olfactometry. J. Food Compost. Anal. 2021, 96, 103761. [Google Scholar] [CrossRef]
- Serio, A.; Chaves-López, C.; Paparella, A.; Suzzi, G. Evaluation of metabolic activities of enterococci isolated from Pecorino Abruzzese cheese. Int. Dairy J. 2010, 20, 459–464. [Google Scholar] [CrossRef]
- García-Cano, I.; Rocha-Mendoza, D.; Ortega-Anaya, J.; Wang, K.; Kosmerl, E.; Jiménez-Flores, R. Lactic acid bacteria isolated from dairy products as potential producers of lipolytic, proteolytic and antibacterial proteins. Appl. Microbiol. Biotechnol. 2019, 103, 5243–5257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliszewski, R.; Medina, R.B.; Gonzalez, S.N.; Perezchaia, A.; Chaia, A.B.P. Esterase activities of indigenous lactic acid bacteria from Argentinean goats’ milk and cheeses. Food Chem. 2007, 101, 1446–1450. [Google Scholar] [CrossRef]
- Boutrou, R.; Guéguen, M. Interests in Geotrichum candidum for cheese technology. Int. J. Food Microbiol. 2005, 102, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sakpuntoon, V.; Angchuan, J.; Boontham, W.; Khunnamwong, P.; Boonmak, C.; Srisuk, N. Grease Waste as a Reservoir of Lipase-Producing Yeast and Description of Limtongella siamensis gen. nov., sp. nov. Microorganisms 2019, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Collins, Y.F.; McSweeney, P.L.H.; Wilkinson, M.G. Lipolysis and free fatty acid catabolism in cheese: A review of current knowledge. Int. Dairy J. 2003, 13, 841–866. [Google Scholar] [CrossRef]
- Fernández, E.; Alegría, A.; Delgado, S.; Mayo, B. Phenotypic, genetic and technological characterization of Lactococcus garvieae strains isolated from a raw milk cheese. Int. Dairy J. 2010, 20, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Kalantzopoulos, G.; Tsakalidou, E.; Manolopoulou, E. Proteinase, peptidase and esterase activities of cell-free extracts from wild strains of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus salivarius subsp. thermophilus isolated from traditional Greek yogurt. J. Dairy Res. 1990, 57, 593–601. [Google Scholar] [CrossRef]
- Higuchi, T.; Aoki, T.; Uchida, K. Esterase activity in soy sauce Moromi as a factor hydrolyzing flavor esters. J. Ferment. Bioeng. 1991, 71, 163–167. [Google Scholar] [CrossRef]
- McSweeney, P.L.; Sousa, M.J. Biochemical pathways for the production of flavour compounds in cheeses during ripening: A review. Le Lait. 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Padilla, B.; Belloch, C.; López-Díez, J.J.; Flores, M.; Manzanares, P. Potential impact of dairy yeasts on the typical flavour of traditional ewes’ and goats’ cheeses. Int. Dairy J. 2014, 35, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Laufenberg, G.; Rosato, P.; Kunz, B. Adding value to vegetable waste: Oil press cakes as substrates for microbial decalactone production. Eur. J. Lipid Sci. Technol. 2004, 106, 207–217. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Milk Origin | Maturation Time | Region | Preparation Method | Storage Temperature |
|---|---|---|---|---|---|
| B1 | Cow | 10 years | Batna | Salting | Ambient |
| B3 | Cow | <1 year | Batna | Salting | Ambient |
| B4 | Cow | <1 year | Batna | Salting | Ambient |
| B5 | Goat | 2 years | Batna | Salting | Ambient |
| Bi3 | Ewe and goat | <1 year | Biskra | Heat-treatment | Refrigerated |
| E10 | Goat | 1 year | El Oued | Heat-treatment | Ambient |
| Jn | Cow | <3 months | Jijel | Heat-treatment | Ambient |
| K1 | Goat | 6 months | Khenchela | Salting | Ambient |
| K2 | Goat | 10 months | Khenchela | Salting | Ambient |
| O1 | Goat | 1 year | Ouargla | Heat-treatment | Refrigerated |
| O3 | Goat | 1 year | Ouargla | Heat-treatment | Ambient |
| S1 | Cow | 6 months | Setif | Salting | Ambient |
| S2 | Cow | 2 months | Setif | Salting | Ambient |
| S3 | Cow | 1 months | Setif | Salting | Ambient |
| S5 | Cow | 2 years | Setif | Salting | Ambient |
| Samples | pH | Moisture (%) | Acid Index (mg KOH/g) | Salt Content (%) | Total Aerobic Mesophilic Bacteria | Total Coliforms | Thermo-Tolerant Coliforms | LAB 1 | Yeast and Molds | Coagulase-Positive Staphylococci | Salmonella spp. | Sulphite-Reducing Anaerobic Bacteria |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | 3.1 ± 0.0 | 19.9 | 153.7 ± 1.7 | ND 1 | <1 | <1 | <1 | <1 | <2 | <2 | n.d. 3 | n.d. |
| B3 | 3.5 ± 0.0 | 12.2 | 35.9 ± 0.0 | ND | 4.5 | <1 | <1 | 2.3 | <2 | <2 | n.d. | n.d. |
| B4 | 3.4 ± 0.0 | 12 | 40.3 ± 0.1 | ND | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| B5 | 3.6 ± 0.0 | 12.6 | 31.4 ± 0.0 | ND | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| Bi3 | 3.8 ± 0.0 | 0.8 | 19.1 ± 1.1 | ND | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| E10 | ND | ND | ND | ND | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| Jn | 4.3 ± 0.0 | 0 | 3.9 ± 0.1 | 0.1 ± 0.0 | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| K1 | 3.3 ± 0.0 | 12.7 | 6.7 ± 0.0 | 3.3 ± 0.0 | 5.2 | <1 | <1 | 5.1 | <2 | <2 | n.d. | n.d. |
| K2 | 3.8 ± 0.0 | 9.0 | 14.6 ± 1.5 | 2.2 ± 0.0 | 4.1 | <1 | <1 | 3.1 | <2 | <2 | n.d. | n.d. |
| O1 | 5.2 ± 0.0 | 0 | 2.2 ± 0.0 | 0.1 ± 0.0 | 3.2 | <1 | <1 | 3.3 | <2 | <2 | n.d. | n.d. |
| O3 | ND | 0 | 2.2 ± 0.0 | ND | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| S1 | 3.3 ± 0.0 | 12.8 | 3.3 ± 0.0 | 7.4 ± 0.1 | 3.7 | <1 | <1 | 4.5 | <2 | <2 | n.d. | n.d. |
| S2 | 3.6 ± 0.0 | 7.3 | 3.6 ± 0.0 | 3.9 ± 0.9 | 5.3 | <1 | <1 | 5.5 | 4.2 | <2 | n.d. | n.d. |
| S3 | 4.4 ± 0.0 | 10.5 | 4.4 ± 0.6 | 1.4 ± 0.0 | 5.4 | <1 | <1 | 6.7 | 4.7 | <2 | n.d. | n.d. |
| S5 | ND | 2.5 | 43.8 ± 1.9 | ND | <1 | <1 | <1 | <1 | <2 | <2 | n.d. | n.d. |
| Sample | pH | Moisture (%) | Dornic Acidity (D°) | Salt Content (%) | Acid Index (mg KOH/g) |
|---|---|---|---|---|---|
| Milk | 6.7 ± 0.0 | ND | 16.3 ± 0.0 | ND | ND |
| Raib | 4.2 ± 0.0 | ND | 47.0 ± 0.0 | ND | ND |
| Butter | 4.5 ± 0.1 | 22.6 | ND | 0.20 ± 0.0 | 6.7 ± 0.0 |
| 1-month Smen (D1) | 4.0 ± 0.1 | 15.9 | ND | 1.10 ± 0.0 | 10.1 ± 1.1 |
| 2-month Smen (D2) | 4.3 ± 0.1 | 15.1 | ND | 1.10 ± 0.0 | 18.4 ± 0.0 |
| 3-month Smen (D3) | 4.1 ± 0.0 | 15.0 | ND | 1.20 ± 0.0 | 19.1 ± 1.1 |
| 6-month Smen (D6) | 4.0 ± 0.0 | 14.8 | ND | 1.30 ± 0.1 | 21.1 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boussekine, R.; Bekhouche, F.; Debaets, S.; Thierry, A.; Maillard, M.-B.; Falentin, H.; Pawtowski, A.; Barkat, M.; Coton, M.; Mounier, J. Deciphering the Microbiota and Volatile Profiles of Algerian Smen, a Traditional Fermented Butter. Microorganisms 2022, 10, 736. https://doi.org/10.3390/microorganisms10040736
Boussekine R, Bekhouche F, Debaets S, Thierry A, Maillard M-B, Falentin H, Pawtowski A, Barkat M, Coton M, Mounier J. Deciphering the Microbiota and Volatile Profiles of Algerian Smen, a Traditional Fermented Butter. Microorganisms. 2022; 10(4):736. https://doi.org/10.3390/microorganisms10040736
Chicago/Turabian StyleBoussekine, Rania, Farida Bekhouche, Stella Debaets, Anne Thierry, Marie-Bernadette Maillard, Hélène Falentin, Audrey Pawtowski, Malika Barkat, Monika Coton, and Jérôme Mounier. 2022. "Deciphering the Microbiota and Volatile Profiles of Algerian Smen, a Traditional Fermented Butter" Microorganisms 10, no. 4: 736. https://doi.org/10.3390/microorganisms10040736