
Experimental Polymorphism Survey in Intergenic Regions of the icaADBCR Locus in Staphylococcus aureus Isolates from Periprosthetic Joint Infections

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Oligonucleotides and Culture Conditions
2.2. DNA Manipulations
2.3. Whole-Genome Sequencing and Genomic Analysis
2.4. Generation of Transcriptional Fusions of Ica Regulatory Sequences with gfp
2.5. Western Blotting
2.6. Biofilm Assay and PIA/PNAG Detection
2.7. Galleria Mellonella Survival Assay
2.8. Statistical Analyses
3. Results
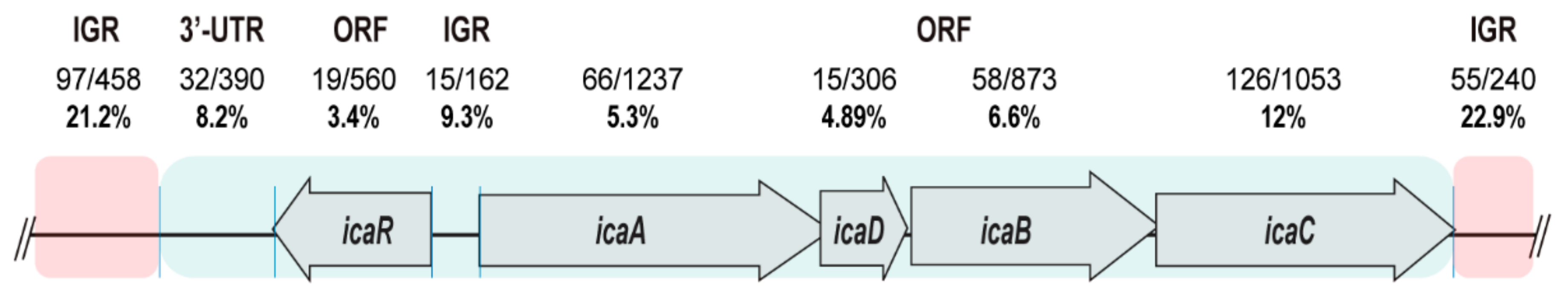
3.1. The SNPs Occurrence Rate Is Low in the Regulatory IGRs of the icaADBCR Locus
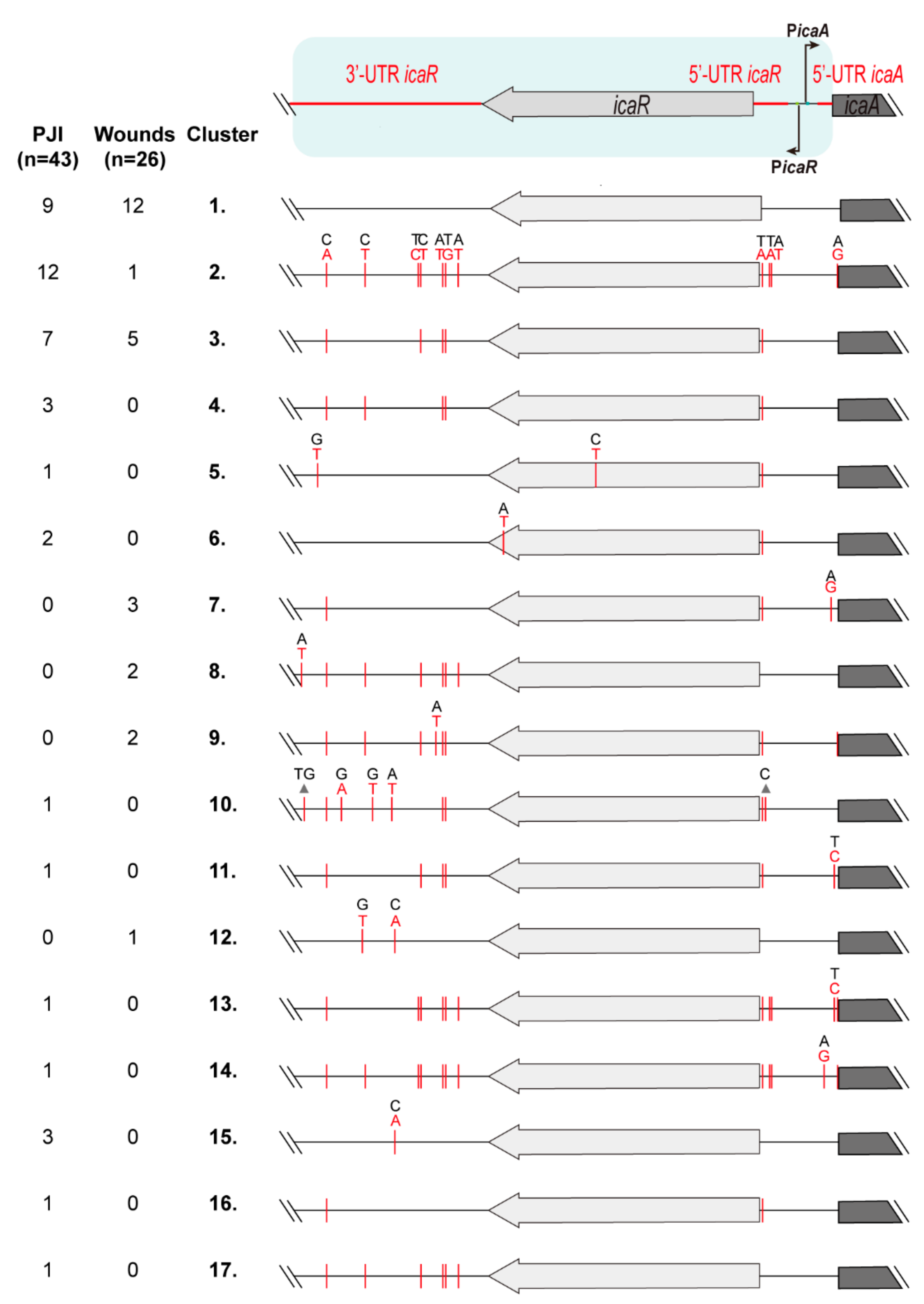
3.2. Identification of Genetic Variations in the Regulatory IGRs of the icaADBCR Locus
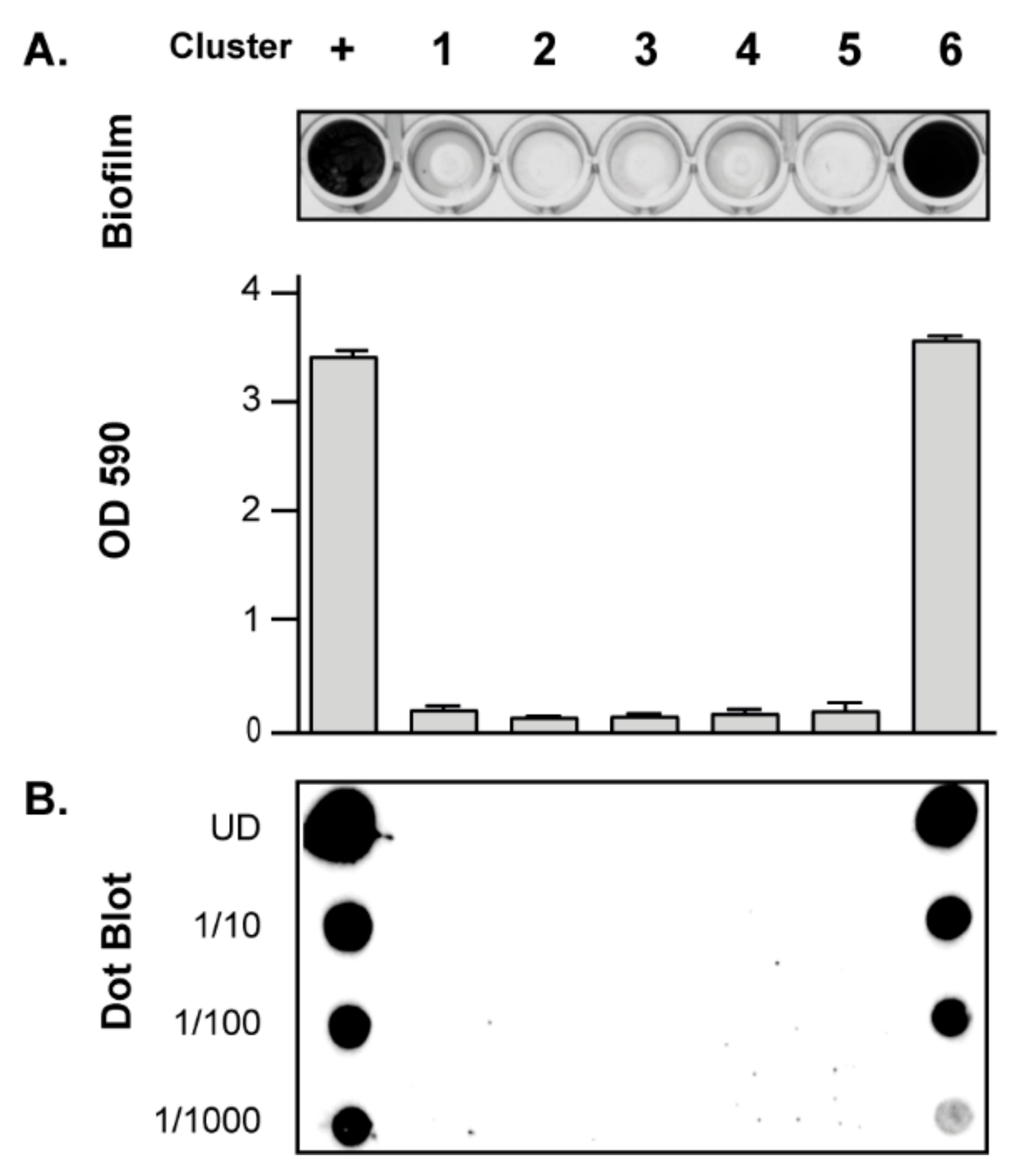
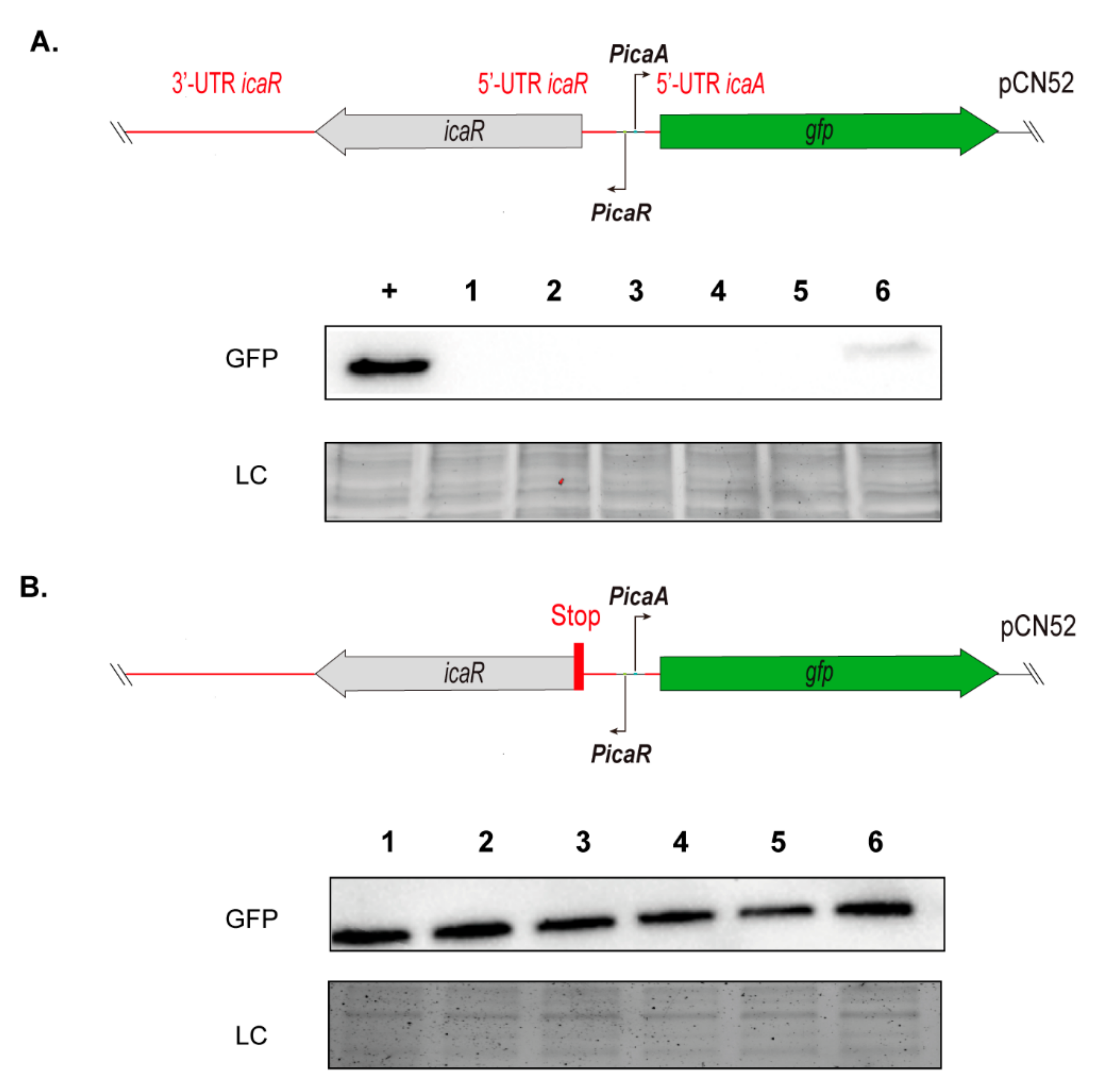
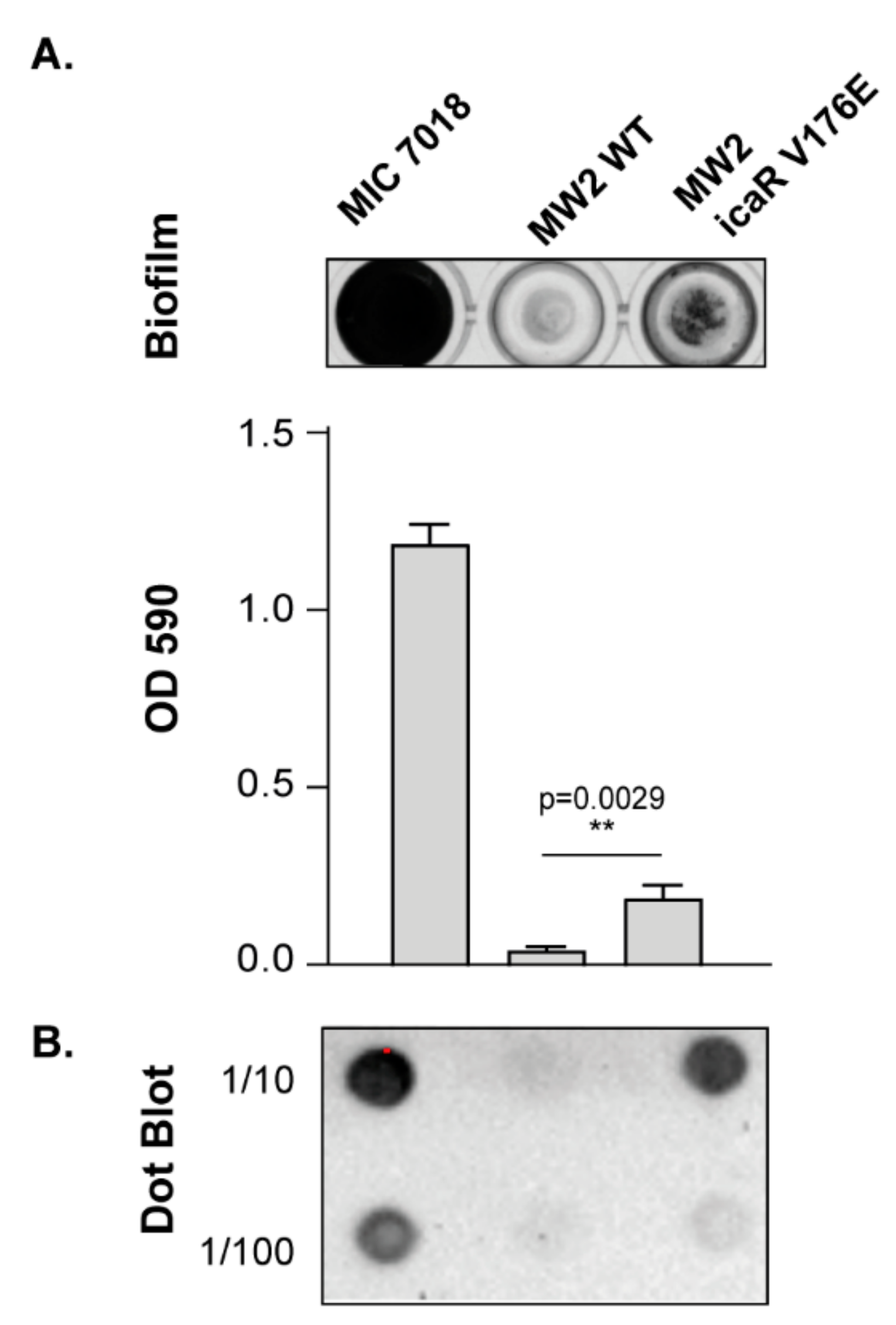
3.3. Contribution of SNPs in the Regulatory IGRs of the icaADBCR Locus to PIA/PNAG Production Capacity
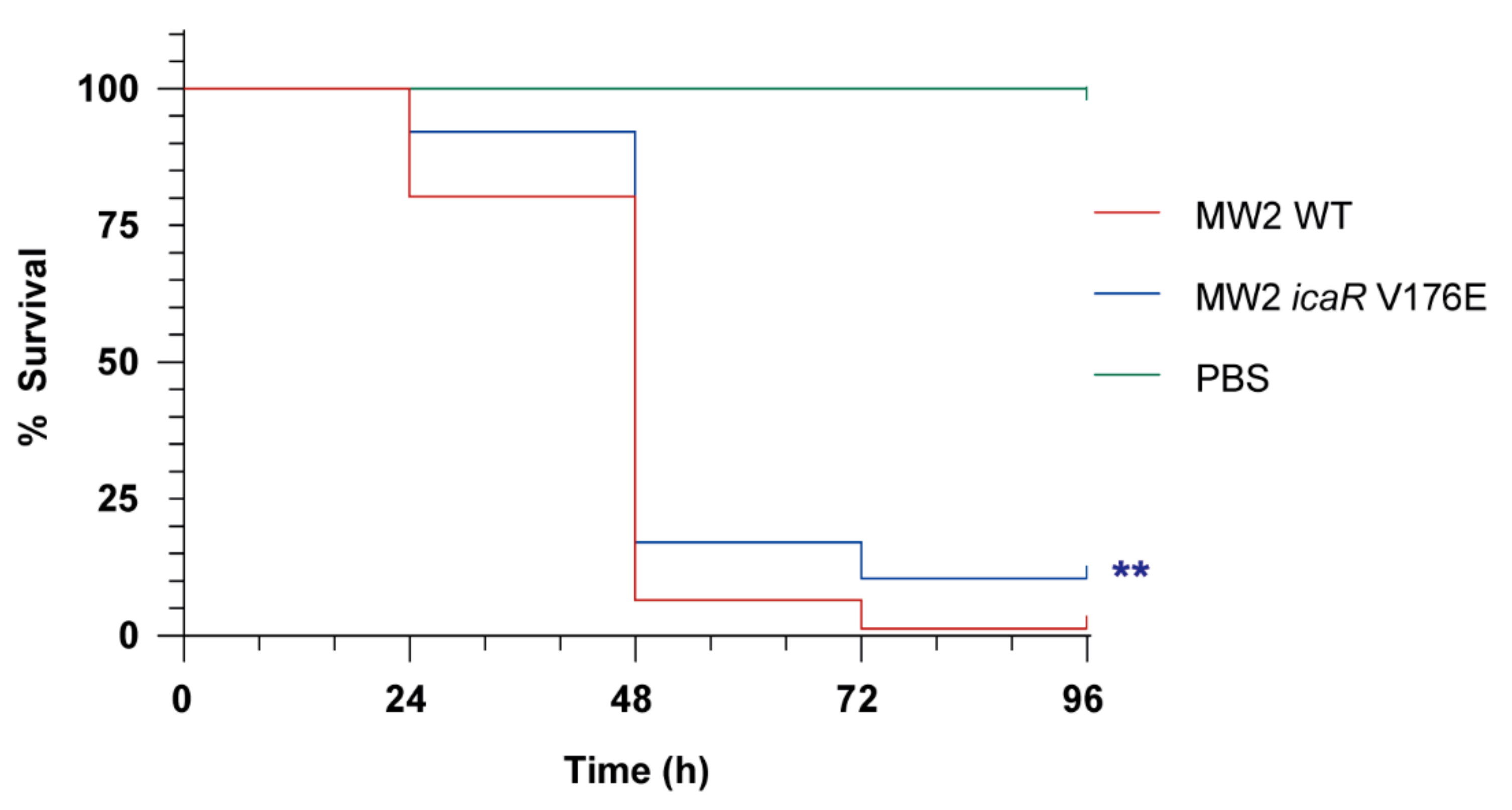
3.4. Increasing PIA/PNAG Production Leads to a Reduction in Virulence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bozic, K.J.; Kurtz, S.M.; Lau, E.; Ong, K.; Chiu, V.; Vail, T.P.; Rubash, H.E.; Berry, D.J. The epidemiology of revision total knee arthroplasty in the United States. Clin. Orthop. Relat. Res. 2010, 468, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tande, A.J.; Patel, R. Prosthetic Joint Infection. Clin. Microbiol. Rev. 2014, 27, 302–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Hou, Y.; Zhang, B.; Chen, Y.; Wang, Q.; Wang, K.; Chen, Z.; Li, X.; Lin, J. Identifying common pathogens in periprosthetic joint infection and testing drug-resistance rate for different antibiotics: A prospective, single center study in Beijing. Orthop. Surg. 2018, 10, 235–240. [Google Scholar] [CrossRef]
- Ricciardi, B.F.; Muthukrishnan, G.; Masters, E.; Ninomiya, M.; Lee, C.C.; Schwarz, E.M. Staphylococcus aureus evasion of host Immunity in the setting of prosthetic joint infection: Biofilm and beyond. Curr. Rev. Musculoskelet. Med. 2018, 11, 389–400. [Google Scholar] [CrossRef]
- Valle, J.; Burgui, S.; Langheinrich, D.; Gil, C.; Solano, C.; Toledo-Arana, A.; Helbig, R.; Lasagni, A.; Lasa, I. Evaluation of surface microtopography engineered by direct laser interference for bacterial anti-biofouling. Macromol. Biosci. 2015, 15, 1060–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speziale, P.; Pietrocola, G.; Foster, T.J.; Geoghegan, J.A. Protein-based biofilm matrices in Staphylococci. Front. Cell Infect. Microbiol. 2014, 4, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, T.J. Surface proteins of Staphylococcus epidermidis. Front. Microbiol. 2020, 11, 1829. [Google Scholar] [CrossRef]
- O’Gara, J.P. ica and beyond: Biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. Fems. Microbiol. Lett. 2007, 270, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Rice, K.C.; Mann, E.E.; Endres, J.L.; Weiss, E.C.; Cassat, J.E.; Smeltzer, M.S.; Bayles, K.W. The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2007, 104, 8113–8118. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcal biofilms. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Schilcher, K.; Horswill, A.R. Staphylococcal biofilm development: Structure, regulation, and treatment strategies. Microbiol. Mol. Biol. Rev. 2020, 84. [Google Scholar] [CrossRef] [PubMed]
- Gotz, F. Staphylococcus and Biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Gotz, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle, J.; Toledo-Arana, A.; Berasain, C.; Ghigo, J.; Amorena, B.; Penadés, J.R.; Lasa, I. SarA and not σB is essential for biofilm development by Staphylococcus aureus. Mol. Microbiol. 2003, 48, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K.; Pier, D.B.; Goldmann, D.A.; Pier, G.B. The Teicoplanin-associated locus regulator (TcaR) and the intercellular adhesin locus regulator (IcaR) are transcriptional inhibitors of the ica locus in Staphylococcus aureus. J. Bacteriol. 2004, 186, 2449–2456. [Google Scholar] [CrossRef] [Green Version]
- Cue, D.; Lei, M.G.; Lee, C.Y. Genetic regulation of the intercellular adhesion locus in Staphylococci. Front. Cell Infect. Microbiol. 2012, 2, 38. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Z.; Zheng, F.; Chew, J.Y.; Pei, Y.; Zhou, J.; Wen, K.; Han, M.; Lemieux, M.J.; Hwang, P.M.; Wen, Y. Deciphering the activation and recognition mechanisms of Staphylococcus aureus response regulator ArlR. Nucleic Acids Res. 2019, 47, 11418–11429. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, J.; Xu, T.; Liu, J.; Yu, W.; Lou, Q.; Zhu, T.; He, N.; Ben, H.; Hu, J.; et al. The two-component signal transduction system ArlRS regulates Staphylococcus epidermidis biofilm formation in an ica-dependent manner. PLoS ONE 2012, 7, e40041. [Google Scholar] [CrossRef]
- Conlon, K.M.; Humphreys, H.; O’Gara, J.P. icaR encodes a transcriptional repressor involved in environmental regulation of ica operon expression and biofilm formation in Staphylococcus epidermidis. J. Bacteriol. 2002, 184, 4400–4408. [Google Scholar] [CrossRef] [Green Version]
- Månsson, E.; Johannesen, T.B.; Nilsdotter-Augustinsson, Å.; Söderquist, B.; Stegger, M. Comparative genomics of Staphylococcus epidermidis from prosthetic-joint infections and nares highlights genetic traits associated with antimicrobial resistance, not virulence. Microb. Genom. 2021, 7. [Google Scholar] [CrossRef]
- Wildeman, P.; Tevell, S.; Eriksson, C.; Lagos, A.C.; Söderquist, B.; Stenmark, B. Genomic characterization and outcome of prosthetic joint infections caused by Staphylococcus aureus. Sci. Rep. 2020, 10, 5938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwood, D.N.; Loughran, A.J.; Courtney, A.P.; Anthony, A.C.; Meeker, D.G.; Spencer, H.J.; Gupta, R.K.; Lee, C.Y.; Beenken, K.E.; Smeltzer, M.S. Comparative impact of diverse regulatory loci on Staphylococcus aureus biofilm formation. Microbiologyopen 2015, 4, 436–451. [Google Scholar] [CrossRef] [Green Version]
- Marvig, R.L.; Sommer, L.M.; Molin, S.; Johansen, H.K. Convergent evolution and adaptation of Pseudomonas aeruginosa within patients with cystic fibrosis. Nat. Genet. 2015, 47, 57–64. [Google Scholar] [CrossRef]
- Osborne, S.E.; Walthers, D.; Tomljenovic, A.M.; Mulder, D.T.; Silphaduang, U.; Duong, N.; Lowden, M.J.; Wickham, M.E.; Waller, R.F.; Kenney, L.J.; et al. Pathogenic adaptation of intracellular bacteria by rewiring a Cis-regulatory input function. Proc. Natl. Acad. Sci. USA 2009, 106, 3982–3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Li, Y.; Cheng, S.; Yang, H.; Lu, J.; Hu, Z.; Ge, B. Mutations in the embC-embA intergenic region contribute to Mycobacterium tuberculosis resistance to ethambutol. Antimicrob. Agents Chemother. 2014, 58, 6837–6843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marvig, R.L.; Damkiær, S.; Khademi, S.M.H.; Markussen, T.M.; Molin, S.; Jelsbak, L. Within-host evolution of Pseudomonas aeruginosa reveals adaptation toward iron acquisition from hemoglobin. Mbio 2014, 5, e00966-14. [Google Scholar] [CrossRef] [Green Version]
- Khademi, S.M.H.; Sazinas, P.; Jelsbak, L. Within-host adaptation mediated by intergenic evolution in Pseudomonas aeruginosa. Genome Biol. Evol. 2019, 11, 1385–1397. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, H.A.; Bayliss, S.C.; Hurst, L.D.; Feil, E.J. Comparative analyses of selection operating on non-translated intergenic regions of diverse bacterial species. Genetics 2017, 206, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Laabei, M.; Recker, M.; Rudkin, J.K.; Aldeljawi, M.; Gulay, Z.; Sloan, T.J.; Williams, P.; Endres, J.L.; Bayles, K.W.; Fey, P.D.; et al. Predicting the virulence of MRSA from its genome sequence. Genome Res. 2014, 24, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, H.A.; Bayliss, S.C.; Sheppard, S.K.; Feil, E.J. Piggy: A rapid, large-scale pan-genome analysis tool for intergenic regions in bacteria. Gigascience 2018, 7, giy015. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, K.K.; Cramton, S.E.; Götz, F.; Pier, G.B. Identification of a 5-nucleotide sequence that controls expression of the ica locus in Staphylococcus aureus and characterization of the DNA-binding properties of IcaR. Mol. Microbiol. 2003, 48, 889–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson Malchau, K.; Tillander, J.; Zaborowska, M.; Hoffman, M.; Lasa, I.; Thomsen, P.; Mal-chau, H.; Rolfson, O.; Trobos, M. Biofilm properties in relation to treatment outcome in patients with first-time periprosthetic hip or knee joint infection. J Orthop Translat. 2021, 30, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Trobos, M.; Firdaus, R.; Svensson Malchau, K.; Tillander, J.; Arnellos, D.; Rolfson, O.; Thomsen, P.; Lasa, I. Genomics of Staphylococcus aureus and Staphylococcus epidermidis from periprosthetic joint infections and correlation to clinical outcome. submitted.
- Monk, I.R.; Tree, J.J.; Howden, B.P.; Stinear, T.P.; Foster, T.J. Complete bypass of restriction systems for major Staphylococcus aureus lineages. Mbio 2015, 6, e00308-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, T.; Takeuchi, F.; Kuroda, M.; Yuzawa, H.; Aoki, K.; Oguchi, A.; Nagai, Y.; Iwama, N.; Asano, K.; Naimi, T.; et al. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet 2002, 359, 1819–1827. [Google Scholar] [CrossRef]
- Vergara-Irigaray, M.; Valle, J.; Merino, N.; Latasa, C.; García, B.; de los Mozos, I.R.; Solano, C.; Toledo-Arana, A.; Penadés, J.R.; Lasa, I. Relevant role of fibronectin-binding proteins in Staphylococcus aureus biofilm-associated foreign-body infections. Infect. Immun. 2009, 77, 3978–3991. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, M.; Chastanet, A.; Débarbouillé, M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, Gram-positive bacteria. Appl. Environ. Microb. 2004, 70, 6887–6891. [Google Scholar] [CrossRef] [Green Version]
- Charpentier, E.; Anton, A.I.; Barry, P.; Alfonso, B.; Fang, Y.; Novick, R.P. Novel cassette-based shuttle vector system for Gram-positive bacteria. Appl. Environ. Microb. 2004, 70, 6076–6085. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Schenk, S.; Laddaga, R. Improved method for electroporation of Staphylococcus aureus. FEMS Microbiol. Lett. 1992, 73, 1338. [Google Scholar]
- Heilmann, C.; Gerke, C.; Perdreau-Remington, F.; Gotz, F. Characterization of Tn917 insertion mutants of Staphylococcus epidermidis affected in biofilm formation. Infect. Immun. 1996, 64, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Kropec, A.; Maira-Litran, T.; Jefferson, K.K.; Grout, M.; Cramton, S.E.; Götz, F.; Goldmann, D.A.; Pier, G.B. Poly-N-Acetylglucosamine production in Staphylococcus aureus is essential for virulence in murine models of systemic infection. Infect. Immun. 2005, 73, 6868–6876. [Google Scholar] [CrossRef] [Green Version]
- de los Mozos, I.R.; Vergara-Irigaray, M.; Segura, V.; Villanueva, M.; Bitarte, N.; Saramago, M.; Domingues, S.; Arraiano, C.M.; Fechter, P.; Romby, P.; et al. Base pairing interaction between 5′- and 3′-UTRs controls icaR mRNA translation in Staphylococcus aureus. PLoS Genet 2013, 9, e1004001. [Google Scholar] [CrossRef]
- Pulido, L.; Ghanem, E.; Joshi, A.; Purtill, J.J.; Parvizi, J. Periprosthetic joint infection: The incidence, timing, and predisposing factors. Clin. Orthop. Relat. Res. 2008, 466, 1710–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flurin, L.; Greenwood-Quaintance, K.E.; Patel, R. Microbiology of polymicrobial prosthetic joint infection. Diagn. Micr. Infec. Dis. 2019, 94, 255–259. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.J.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asthana, S.; Roytberg, M.; Stamatoyannopoulos, J.; Sunyaev, S. Analysis of sequence conservation at nucleotide resolution. PLoS Comput. Biol. 2007, 3, e254. [Google Scholar] [CrossRef] [PubMed]
- Dermitzakis, E.T.; Reymond, A.; Antonarakis, S.E. Conserved non-genic sequences—an unexpected feature of mammalian genomes. Nat. Rev. Genet. 2005, 6, 151–157. [Google Scholar] [CrossRef]
- Cooper, G.M.; Brown, C.D. Qualifying the relationship between sequence conservation and molecular function. Genome Res. 2008, 18, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Jeng, W.-Y.; Ko, T.-P.; Liu, C.-I.; Guo, R.-T.; Liu, C.-L.; Shr, H.-L.; Wang, A.H.-J. Crystal structure of IcaR, a repressor of the TetR family implicated in biofilm formation in Staphylococcus epidermidis. Nucleic Acids Res. 2008, 36, 1567–1577. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.L.; Jefferson, K.K. Phase variation of Poly-N-Acetylglucosamine expression in Staphylococcus aureus. PLoS Pathog. 2014, 10, e1004292. [Google Scholar] [CrossRef]
- Schwartbeck, B.; Birtel, J.; Treffon, J.; Langhanki, L.; Mellmann, A.; Kale, D.; Kahl, J.; Hirschhausen, N.; Neumann, C.; Lee, J.C.; et al. Dynamic in vivo mutations within the ica operon during persistence of Staphylococcus aureus in the airways of cystic fibrosis patients. PLoS Pathog. 2016, 12, e1006024. [Google Scholar] [CrossRef]
- Coombes, B.K. Regulatory evolution at the host–pathogen interface. Can J. Microbiol. 2013, 59, 365–367. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Characteristic(s) | MIC a | Reference |
|---|---|---|---|
| E. coli IM01B | E. coli K12 DH10B Δdcm and containing the hsdS gene of MW2 integrated between the essQ and cspB genes. Used for cloning experiments and isolation of plasmids that are transformed into S. aureus CC1 strains at high efficiency | 5694 | [34] |
| S. aureus 15981 | MSSA clinical isolate; strong biofilm producer; PNAG-dependent biofilm matrix | 0532 | [14] |
| S. aureus MW2 | Community-acquired strain of MRSA, which was isolated in 1998 in North Dakota, USA | 3566 | [35] |
| S. aureus MW2 icaR V176E | MW2 strain harbouring a mutation in the icaR gene, coding for a V176E IcaR variant | 7983 | This study |
| S. aureus 132 ica::tet | S. aureus 132 containing a tetracycline resistance cassette that replaces ica genes | 3343 | [36] |
| S. aureus clinical isolate | Sahlgrenska University Hospital (Sweden). Periprosthetic joint infection (hip) | 6924 | [32,33] |
| S. aureus clinical isolate | Sahlgrenska University Hospital (Sweden). Periprosthetic joint infection (hip) | 6934 | [32,33] |
| S. aureus clinical isolate | Sahlgrenska University Hospital (Sweden). Periprosthetic joint infection (hip) | 6935 | [32,33] |
| S. aureus clinical isolate | Sahlgrenska University Hospital (Sweden). Periprosthetic joint infection (hip) | 6936 | [32,33] |
| S. aureus clinical isolate | Sahlgrenska University Hospital (Sweden). Periprosthetic joint infection (hip) | 6948 | [32,33] |
| S. aureus clinical isolate | Sahlgrenska University Hospital (Sweden). Periprosthetic joint infection (knee). Strong biofilm producer | 7018 | [32,33] |
| S. aureus clinical isolates | Culture Collection at University of Gothenburg, (CCGU), Sweden. Wounds | 7032–7050 7166–7172 | This study |
| Plasmids | Relevant Characteristics | Reference |
|---|---|---|
| pJET1.2 | Cloning vector. AmpR. | Thermo Scientific |
| pMAD | E. coli-S. aureus shuttle vector with a thermosensitive origin of replication for Gram-positive bacteria. AmpR EryR | [37] |
| pMAD::icaRV176E | pMAD plasmid containing the DNA sequence for icaR V176E mutation | This study |
| pCN52 | E. coli-S. aureus shuttle vector with promoterless gfpmut2. EryR | [38] |
| pCN52::IGRica C1 | 3′UTR icaR-icaR—IGR icaR/icaA amplified from MIC 6935 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRica C2 | 3′UTR icaR-icaR—IGR icaR/icaA amplified from MIC 6924 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRica C3 | 3′UTR icaR-icaR—IGR icaR/icaA amplified from MIC 6934 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRica C4 | 3′UTR icaR-icaR—IGR icaR/icaA amplified from MIC 6936 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRica C5 | 3′UTR icaR-icaR—IGR icaR/icaA amplified from MIC 6948 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRica C6 | 3′UTR icaR-icaR—IGR icaR/icaA amplified from MIC 7018 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRStopica C1 | 3′UTR icaR-icaRSTOP—IGR icaR/icaA amplified from MIC 6935 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRStopica C2 | 3′UTR icaR-icaRSTOP—IGR icaR/icaA amplified from MIC 6924 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRStopica C3 | 3′UTR icaR-icaRSTOP—IGR icaR/icaA amplified from MIC 6934 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRStopica C4 | 3′UTR icaR-icaRSTOP—IGR icaR/icaA amplified from MIC 6936 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRStopica C5 | 3′UTR icaR-icaRSTOP—IGR icaR/icaA amplified from MIC 6948 and fused to promoterless gfpmut2 in pCN52 | This study |
| pCN52::IGRStopica C6 | 3′UTR icaR-icaRSTOP—IGR icaR/icaA amplified from MIC 7018 and fused to promoterless gfpmut2 in pCN52 | This study |
| Oligonucleotides | Sequencea | |
| Au56 | ATGCCTGCAGGTCGACCGAGTAGAAGCATCATCATTACTTGATT | |
| Au73 | GGATCCTAAGCCATATGGTAATTGATAG | |
| Au76 | ACGAATTCGAGCTCGGTACCTTTCTTTACCTACCTTTCGTTAGTTAGGTTG | |
| Au78 | TTATTGATAACGCAATAACCTTATAAGGATCCTTTTCAGAGAAGGGGTATGACGG | |
| Au77 | CCGTCATACCCCTTCTCTGAAAAGGATCCTTATAAGGTTATTGCGTTATCAATAA | |
| LM9 | CAAAGATGAAGAGTACTCGCTACTAAATA | |
| LM10 | TAGTAGCGAGTACTCTTCATCTTTGAATTG | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Laverde, L.; Echeverz, M.; Trobos, M.; Solano, C.; Lasa, I. Experimental Polymorphism Survey in Intergenic Regions of the icaADBCR Locus in Staphylococcus aureus Isolates from Periprosthetic Joint Infections. Microorganisms 2022, 10, 600. https://doi.org/10.3390/microorganisms10030600
Morales-Laverde L, Echeverz M, Trobos M, Solano C, Lasa I. Experimental Polymorphism Survey in Intergenic Regions of the icaADBCR Locus in Staphylococcus aureus Isolates from Periprosthetic Joint Infections. Microorganisms. 2022; 10(3):600. https://doi.org/10.3390/microorganisms10030600
Chicago/Turabian StyleMorales-Laverde, Liliana, Maite Echeverz, Margarita Trobos, Cristina Solano, and Iñigo Lasa. 2022. "Experimental Polymorphism Survey in Intergenic Regions of the icaADBCR Locus in Staphylococcus aureus Isolates from Periprosthetic Joint Infections" Microorganisms 10, no. 3: 600. https://doi.org/10.3390/microorganisms10030600