
The Impact of Pre- and Probiotic Product Combinations on Ex vivo Growth of Avian Pathogenic Escherichia coli and Salmonella Enteritidis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Pathogens and Media
2.4. Determination of Suitable Antibiotic Combinations
2.5. Pre- and Probiotic Products
2.6. Ex vivo Assay and Determination of Growth Parameters
2.7. 16S rDNA Sequencing
2.8. Statistical Analysis
3. Results
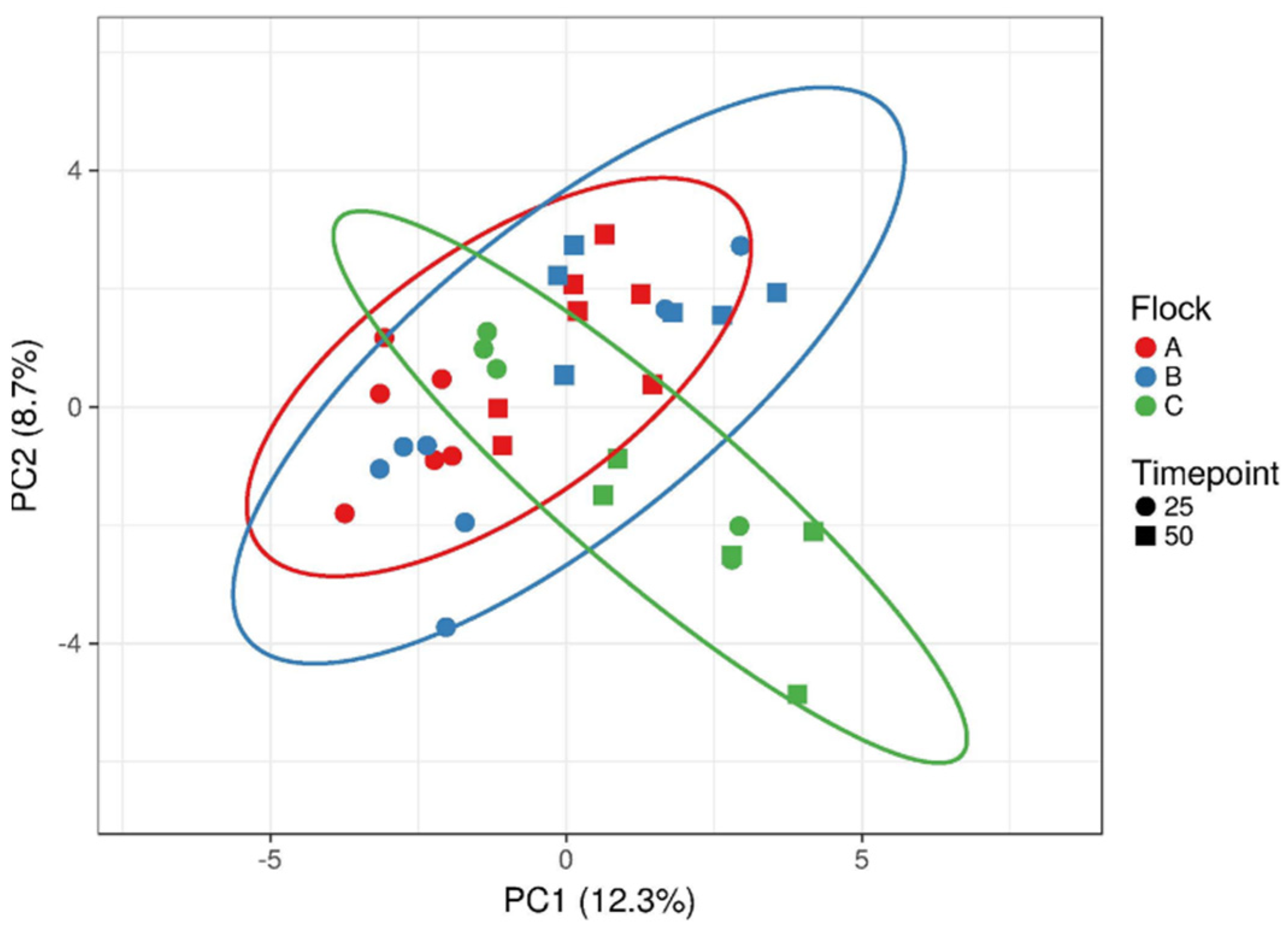
3.1. Original Bacterial Composition of Flock Caecal Contents
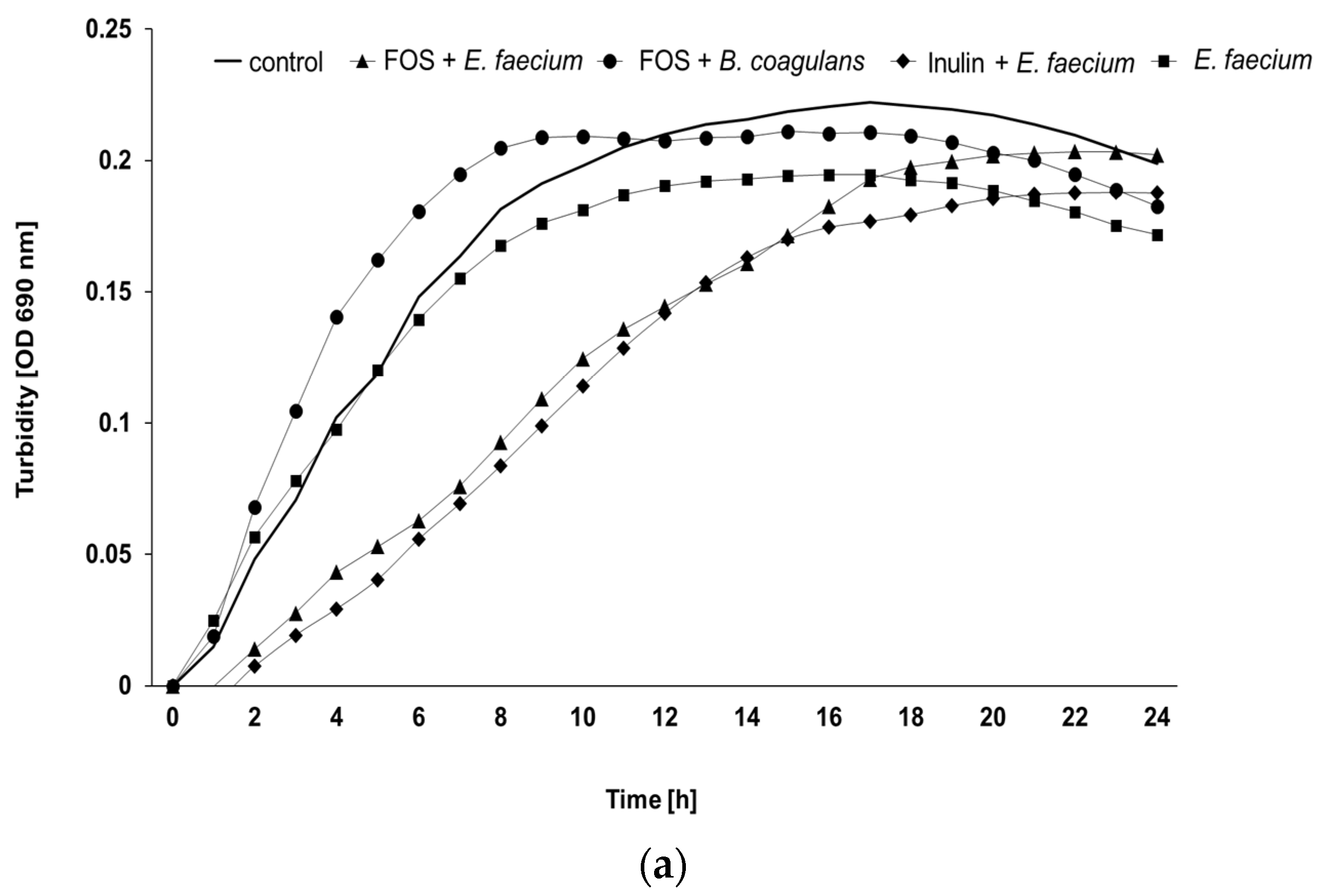
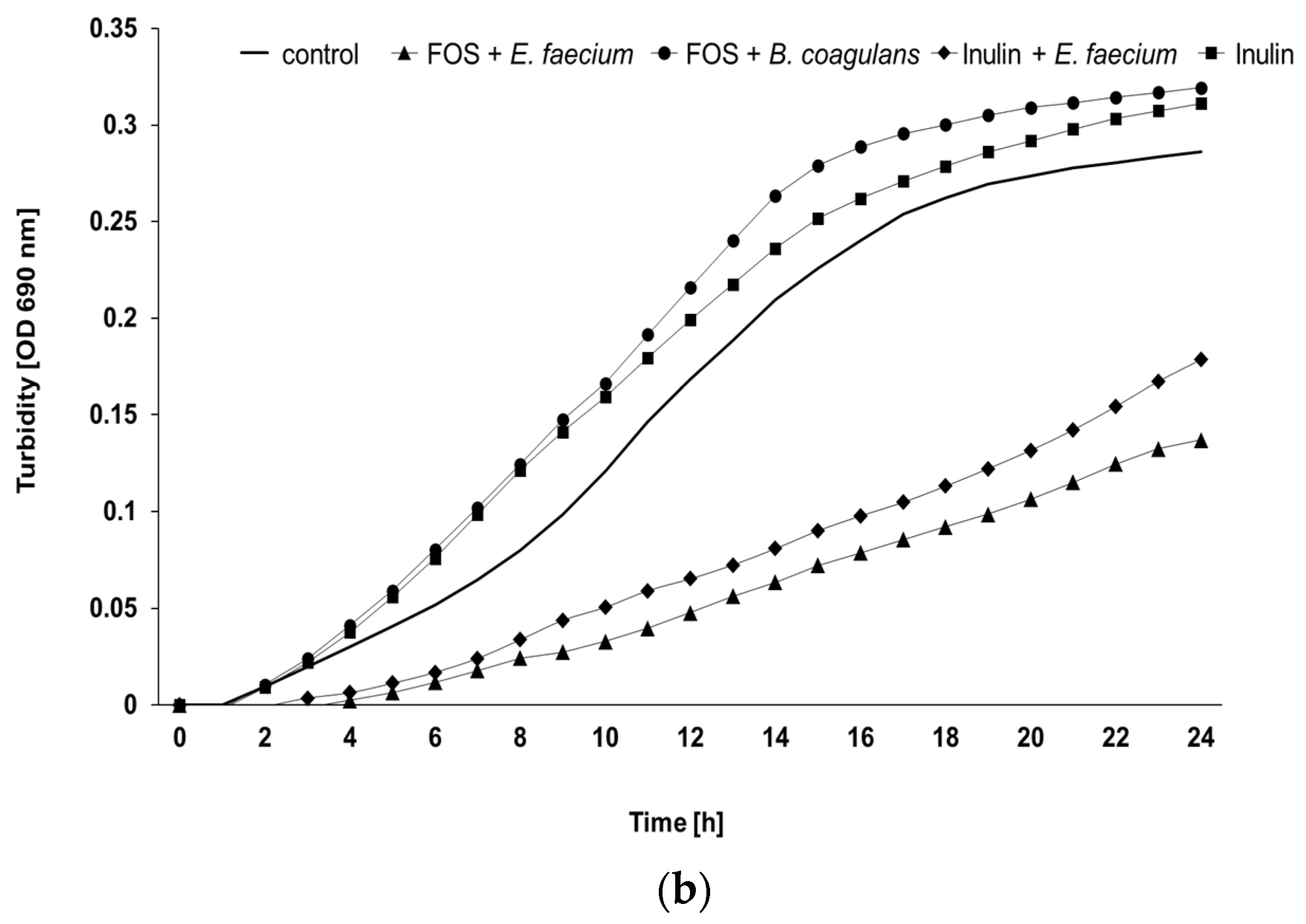
3.2. Ex Vivo Assay
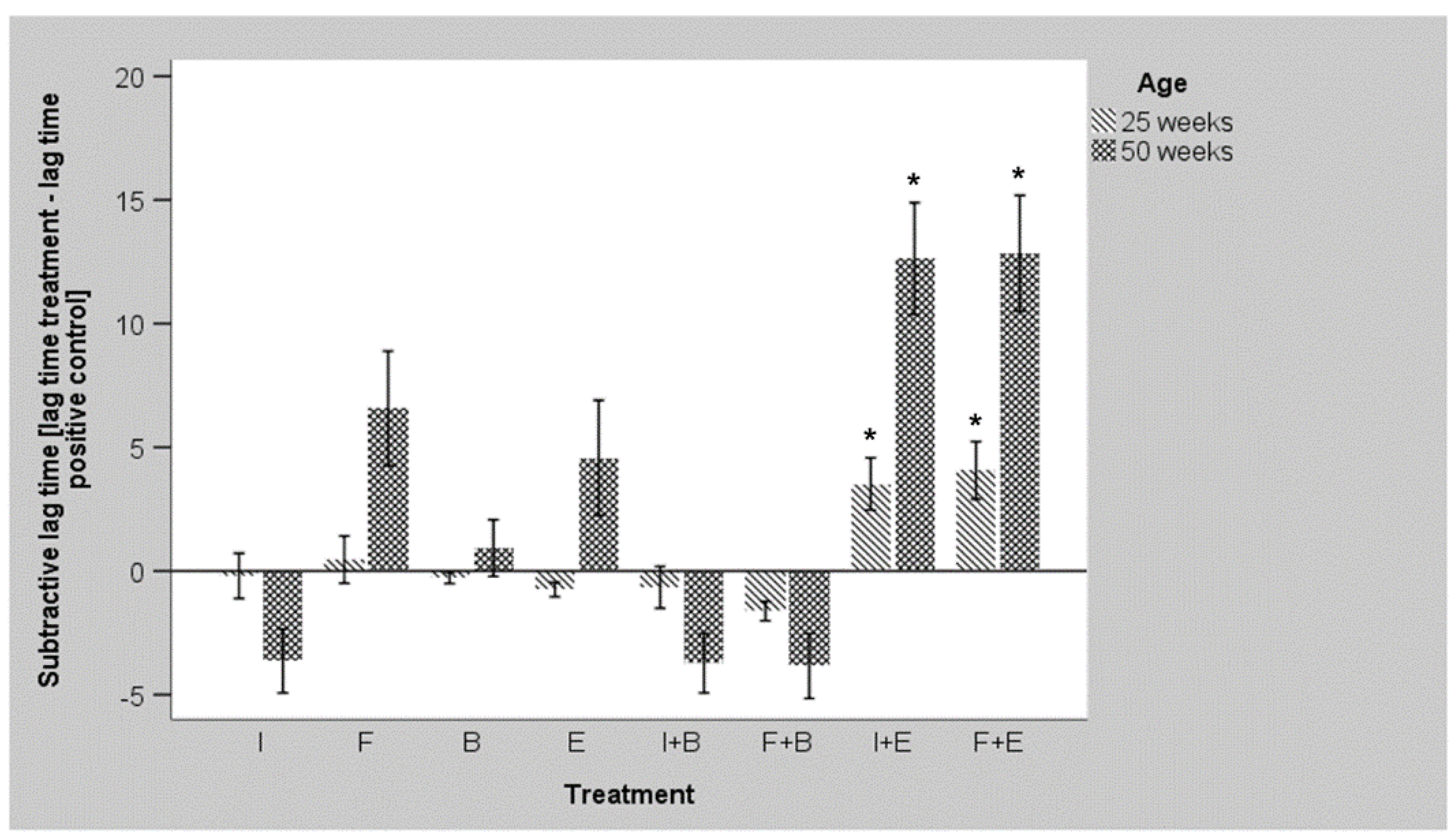
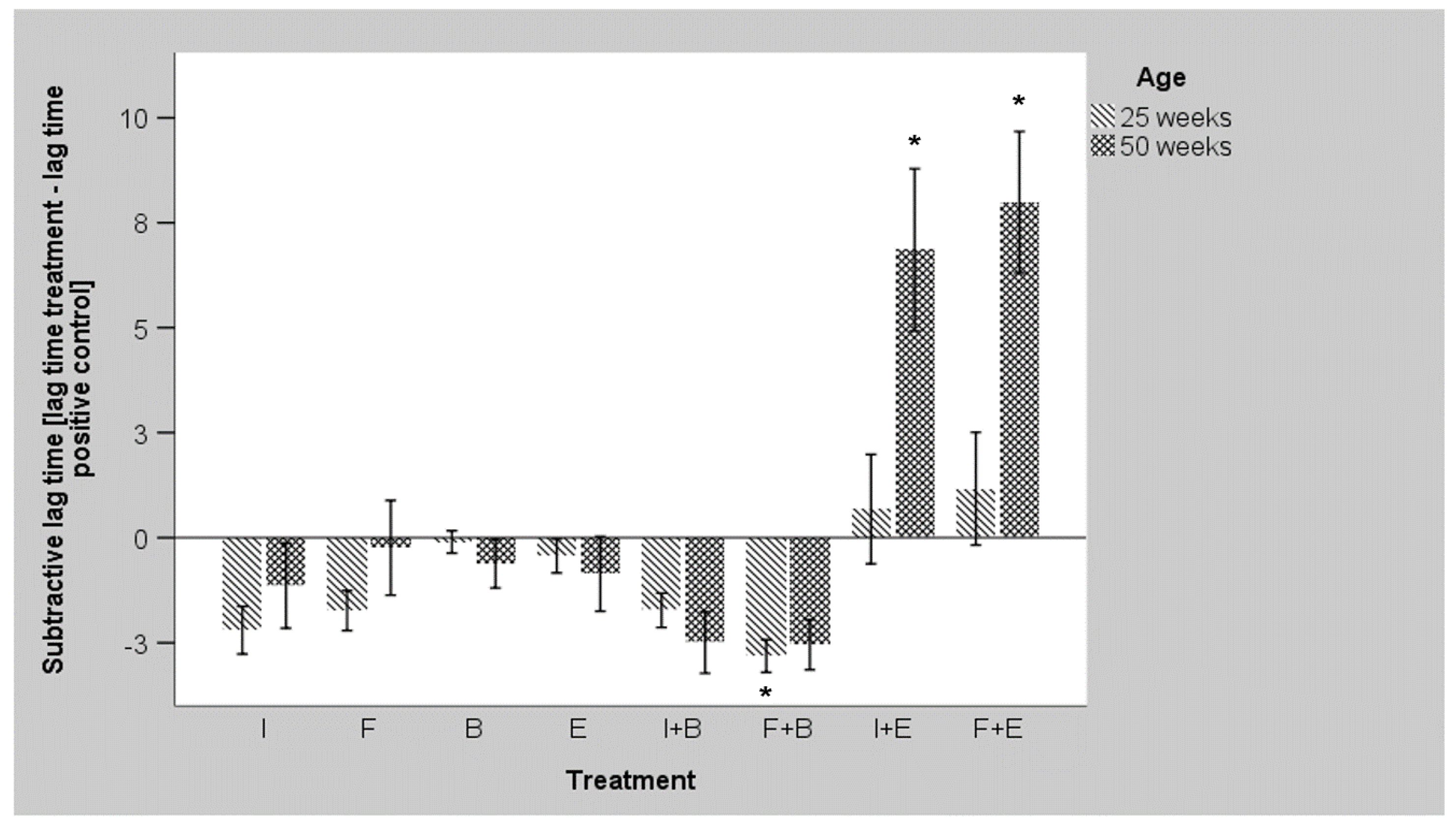
3.3. Ex Vivo Growth of E. coli O1/O18 after Incubation with the Additives
3.4. Ex Vivo Growth of S. Enteritidis after Incubation with the Additives
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eurostat. Agriculture, Forestry and Fishery Statistics, 2020 Edition. Available online: https://ec.europa.eu/eurostat/web/products-statistical-books/-/ks-fk-20-001 (accessed on 11 October 2021).
- Snow, L.C.; Davies, R.H.; Christiansen, K.H.; Carrique-Mas, J.J.; Cook, A.J.; Evans, S.J. Investigation of risk factors for Salmonella on commercial egg-laying farms in Great Britain, 2004–2005. Vet. Rec. 2010, 166, 579–586. [Google Scholar] [CrossRef] [Green Version]
- European Union. RASFF—The Rapid Alert System for Food and Feed—Annual Report 2020. Available online: https://ec.europa.eu/food/system/files/2021-08/rasff_pub_annual-report_2020.pdf (accessed on 1 November 2021).
- EFSA, European Food Safety Authority. The European Union One Health 2019 Zoonoses Report. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/6406 (accessed on 1 November 2021).
- Barnes, H.J.; Nolan, L.K.; Vaillancourt, J.P. Salmonella Infections. In Diseases of Poultry, 12th ed.; Saif, Y.M., Ed.; Blackwell Publishing: Ames, IA, USA, 2008; pp. 619–665. [Google Scholar]
- Chantziaras, I.; Boyen, F.; Callens, B.; Dewulf, J. Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: A report on seven countries. J. Antimicrob. Chemother. 2014, 69, 827–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliu, E.M.; Vahjen, W.; Zentek, J. Types and prevalence of extended-spectrum beta-lactamase producing Enterobacteriaceae in poultry. Anim. Health Res. Rev. 2017, 18, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, H.J.; Nolan, L.K.; Vaillancourt, J.P. Colibacillosis. In Diseases of Poultry, 12th ed.; Saif, Y.M., Ed.; Blackwell Publishing: Ames, IA, USA, 2008; pp. 691–732. [Google Scholar]
- Lutful Kabir, S.M. Avian colibacillosis and salmonellosis: A closer look at epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Public Health. 2010, 7, 89–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anadón, A. WS 14 The EU ban of antibiotics as feed additives (2006): Alternatives and consumer safety. J. Vet. Pharmacol. Ther. 2006, 29, 41–44. [Google Scholar] [CrossRef]
- More, S.J. European perspectives on efforts to reduce antimicrobial usage in food animal production. Ir. Vet. J. 2020, 73, 2. [Google Scholar] [CrossRef] [Green Version]
- IACG; Interagency Coordination Group on Antimicrobial Resistance. No Time to Wait: Securing the Future from Drug-Resistant Infections, Report to the Secretary-General of the United Nations. Available online: https://www.who.int/antimicrobial-resistance/interagency-coordination-group/IACG_final_report_EN.pdf?ua=1 (accessed on 10 March 2021).
- Clavijo, V.; Flórez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef] [PubMed]
- Methner, U.; Rösler, U. Efficacy of a competitive exclusion culture against extended-spectrum β-lactamase-producing Escherichia coli strains in broilers using a seeder bird model. BMC Vet. Res. 2020, 16, 143. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Sarangi, N.R.; Babu, L.K.; Kumar, A.; Pradhan, C.R.; Pati, P.K.; Mishra, J.P. Effect of dietary supplementation of prebiotic, probiotic, and synbiotic on growth performance and carcass characteristics of broiler chickens. Vet. World 2016, 9, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Al-Enzi, N.; Al-Sharrah, T.; Ragheb, G.; Al-Qalaf, S.; Mohammed, A. Effect of dietary probiotics and prebiotics on the performance of broiler chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Antonissen, G.; Eeckhaut, V.; Van Driessche, K.; Onrust, L.; Haesebrouck, F.; Ducatelle, R.; Moore, R.J.; Van Immerseel, F. Microbial shifts associated with necrotic enteritis. Avian Pathol. 2016, 45, 308–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeilinger, K.; Hellmich, J.; Zentek, J.; Vahjen, W. Novel ex vivo screening assay to preselect farm specific pre- and probiotics in pigs. Benef. Microbes 2021, 12, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Saliu, E.M.; Zentek, J.; Goodarzi Boroojeni, F.; Vahjen, W. Screening of Host Specific Lactic Acid Bacteria Active Against Escherichia coli From Massive Sample Pools With a Combination of in vitro and ex vivo Methods. Front. Microbiol. 2019, 10, 2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Huse, S.M.; Huber, J.A.; Morrison, H.G.; Sogin, M.L.; Welch, D.M. Accuracy and quality of massively parallel DNA pyrosequencing. Genome Biol. 2007, 8, R143. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vázquez-Baeza, Y.; Birmingham, A.; et al. Normalization and microbial differential abundance strategies depend upon data characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Classen, H.; Apajalahti, J.; Svihus, B.; Choct, M. The role of the crop in poultry production. World’s Poult. Sci. J. 2016, 72, 459–472. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Vahjen, W.; Dadi, T.; Saliu, E.M.; Boroojeni, F.G.; Zentek, J. Synergistic Effects of Probiotics and Phytobiotics on the Intestinal Microbiota in Young Broiler Chicken. Microorganisms 2019, 7, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyarzabal, O.A.; Conner, D.E.; Blevins, W.T. Fructooligosaccharide Utilization by Salmonellae (†) and Potential Direct-Fed-Microbial Bacteria for Poultry. J. Food Prot. 1995, 58, 1192–1196. [Google Scholar] [CrossRef]
- Rehman, H.; Böhm, J.; Zentek, J. Effects of differentially fermentable carbohydrates on the microbial fermentation profile of the gastrointestinal tract of broilers. J. Anim. Physiol. Anim. Nutr. 2008, 92, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Kumar, S.; Thippareddi, H.; Kim, W.K. Effect of Dietary Fructooligosaccharide (FOS) Supplementation on Ileal Microbiota in Broiler Chickens. Poult. Sci. 2018, 97, 3622–3634. [Google Scholar] [CrossRef]
- Nabizadeh, A. The effect of inulin on broiler chicken intestinal microflora, gut morphology, and performance. J. Anim. Feed Sci. 2012, 21, 725–734. [Google Scholar] [CrossRef]
- Samli, H.E.; Senkoylu, N.; Koc, F.; Kanter, M.; Agma, A. Effects of Enterococcus faecium and dried whey on broiler performance, gut histomorphology and intestinal microbiota. Arch. Anim. Nutr. 2007, 61, 42–49. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Yu, Y.; Zhang, R.; Wu, Y.; Yue, M.; Yang, C. Effects of Bacillus Coagulans on growth performance, antioxidant capacity, immunity function, and gut health in broilers. Poult. Sci. 2021, 100, 101168. [Google Scholar] [CrossRef]
- Reich, F.; Atanassova, V.; Klein, G. Extended-spectrum β-lactamase- and AmpC-producing enterobacteria in healthy broiler chickens, Germany. Emerg. Infect. Dis. 2013, 19, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in animal nutrition and health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Watzl, B.; Girrbach, S.; Roller, M. Inulin, oligofructose and immunomodulation. Br. J. Nutr. 2005, 93 (Suppl. 1), S49–S55. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Antão, E.M.; Diehl, I.; Philipp, H.C.; Wieler, L.H. Intestine and environment of the chicken as reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential. Appl. Environ. Microbiol. 2009, 75, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Brooks, L.A.; Bailey, M.A.; Krehling, J.T.; Chasteen, K.S.; Macklin, K.S. A Comparison of Colonizing Ability Between Salmonella Enteritidis and Salmonella Heidelberg in Broiler Chickens Challenged Through Feed Administration. Foodborne Pathog. Dis. 2021, 18, 784–789. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Böhm, J. Intestinal structure and function of broiler chickens on diets supplemented with a synbiotic containing Enterococcus faecium and oligosaccharides. Int. J. Mol. Sci. 2008, 9, 2205–2216. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.L.; Rebolé, A.; Velasco, S.; Ortiz, L.T.; Treviño, J.; Alzueta, C. Wheat- and barley-based diets with or without additives influence broiler chicken performance, nutrient digestibility and intestinal microflora. J. Sci. Food Agric. 2012, 92, 184–190. [Google Scholar] [CrossRef]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.; Hartke, A.; Huycke, M. The Physiology and Metabolism of Enterococci. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Oyarzabal, O.A.; Conner, D.E. In vitro fructooligosaccharide utilization and inhibition of Salmonella spp. by selected bacteria. Poult. Sci. 1995, 74, 1418–1425. [Google Scholar] [CrossRef]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef]
- Ohenhen, R.; Isibor, J.; Emonfonmwan, G.; Enabulele, S. Effects of PH and Storage Temperatures on Antibacterial Activity of Bacteriocin Produced by Lactic Acid Bacteria Isolated from OGI. Br. Microbiol. Res. J. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Freese, E.; Sheu, C.W.; Galliers, E. Function of lipophilic acids as antimicrobial food additives. Nature 1973, 241, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Poeta, P.; Costa, D.; Rodrigues, J.; Torres, C. Detection of genes encoding virulence factors and bacteriocins in fecal enterococci of poultry in Portugal. Avian Dis. 2006, 50, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Kim, I.H. Effects of probiotic supplementation in different energy and nutrient density diets on performance, egg quality, excreta microflora, excreta noxious gas emission, and serum cholesterol concentrations in laying hens. J. Anim. Sci. 2013, 91, 4781–4787. [Google Scholar] [CrossRef] [PubMed]
- Comerlato, C.B.; Ritter, A.C.; Miyamoto, K.N.; Brandelli, A. Proteomic study of Enterococcus durans LAB18S growing on prebiotic oligosaccharides. Food Microbiol. 2020, 89, 103430. [Google Scholar] [CrossRef]
- Zhen, W.; Shao, Y.; Gong, X.; Wu, Y.; Geng, Y.; Wang, Z.; Guo, Y. Effect of dietary Bacillus coagulans supplementation on growth performance and immune responses of broiler chickens challenged by Salmonella enteritidis. Poult. Sci. 2018, 97, 2654–2666. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, Z.; Xu, Y.; Ying, J.; Wang, B.; Majeed, M.; Majeed, S.; Pande, A.; Li, W. Application of Bacillus coagulans in Animal Husbandry and Its Underlying Mechanisms. Animals 2020, 10, 454. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shao, Y.; Song, B.; Zhen, W.; Wang, Z.; Guo, Y.; Shahid, M.S.; Nie, W. Effects of Bacillus coagulans supplementation on the growth performance and gut health of broiler chickens with Clostridium perfringens-induced necrotic enteritis. J. Anim. Sci. Biotechnol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Valdés-Varela, L.; Ruas-Madiedo, P.; Gueimonde, M. In vitro fermentation of different fructo-oligosaccharides by Bifidobacterium strains for the selection of synbiotic combinations. Int. J. Food Microbiol. 2017, 242, 19–23. [Google Scholar] [CrossRef]
- Ten Bruggencate, S.J.; Bovee-Oudenhoven, I.M.; Lettink-Wissink, M.L.; Katan, M.B.; Van Der Meer, R. Dietary fructo-oligosaccharides and inulin decrease resistance of rats to salmonella: Protective role of calcium. Gut 2004, 53, 530–535. [Google Scholar] [CrossRef] [Green Version]
- Lührmann, A.; Ovadenko, K.; Hellmich, J.; Sudendey, C.; Belik, V.; Zentek, J.; Vahjen, W. Characterization of the fecal microbiota of sows and their offspring from German commercial pig farms. PLoS ONE 2021, 16, e0256112. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Broiler Breeder Flock | |||||
|---|---|---|---|---|---|
| Age | A | B | C | SEM | p-Value * |
| 25 weeks | |||||
| Richness | 135.29 | 173.8 | 139.17 | 8.5 | 0.077 |
| Shannon-Index | 4.07 | 3.61 | 3.81 | 0.10 | 0.161 |
| Evenness | 0.833 b | 0.705 a | 0.775 a,b | 0.021 | 0.007 |
| 50 weeks | |||||
| Richness | 177.29 b | 131.5 a | 246.0 c | 14.0 | 0.016 |
| Shannon-Index | 4.40 b | 4.00 a | 4.64 b | 0.07 | 0.03 |
| Evenness | 0.852 b | 0.822 a | 0.849 b | 0.053 | 0.046 |
| Prebiotic Products | Probiotic Products | Bacillus and Prebiotic | Enterococcus and Prebiotic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Age (Weeks) | Flock | Control | Inulin | FOS | Bacillus | Enterococcus | Inulin | FOS | Inulin | FOS | p-Value Treatment |
| 25 | A | 4.99 a,b | 4.90 a,b,c | 5.29 a,b,c | 4.60 a | 4.02 a | 4.69 a | 3.51 a | 9.29 b,c | 9.28 c | 0.011 |
| B | 5.79 | 5.00 | 6.31 | 4.80 | 4.34 | 4.78 | 3.84 | 7.99 | 8.23 | 0.441 | |
| C | 4.28 | 4.52 | 4.84 | 4.78 | 4.47 | 4.57 | 2.84 | 8.32 | 9.76 | 0.137 | |
| All | 5.02 a,b | 4.81 a,b | 5.48 a,b | 4.73 a,b | 4.27 a | 4.68 a,b | 3.40 a | 8.53 b | 9.09 b | <0.001 | |
| 50 | A | 6.43 a,b | 4.39 a,b | 5.36 a,b | 9.84 b | 6.24 a,b | 3.60 a,b | 2.38 a | 12.48 b | 12.46 a,b | 0.009 |
| B | 5.95 a | 7.73 a,b | 17.70 a,b | 7.53 a,b | 17.13 a,b | 5.81 a,b | 5.91 a,b | 24.00 b | 24.00 b | <0.001 | |
| C | 7.35 a,b | 3.35 a,b | 16.36 b,c | 5.11 a,b | 10.06 a,b,c | 3.70 a | 5.72 a,b | 21.11 c | 21.75 c | 0.006 | |
| All | 6.57 a | 4.90 a | 13.14 a,b | 7.49 a,b | 11.15 a,b | 4.29 a | 4.10 a | 19.20 b | 19.41 b | <0.001 | |
| All | All | 5.80 a,b | 4.84 a,b | 9.31 b,c | 6.11 a,b | 7.71 a,b | 4.51 a,b | 3.68 a | 13.86 c | 14.25 c | <0.001 |
| p-value age | 0.250 | 0.726 | 0.016 | 0.005 | 0.015 | 0.796 | 0.643 | <0.001 | 0.003 | ||
| Prebiotic Products | Probiotic Products | Bacillus and Prebiotic | Enterococcus and Prebiotic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Age (Weeks) | Flock | Control | Inulin | FOS | Bacillus | Enterococcus | Inulin | FOS | Inulin | FOS | p-Value Treatment |
| 25 | A | 7.83 b | 5.25 a | 6.05 a,b | 7.78 b | 7.94 b | 6.47 a,b | 6.25 a,b | 6.35 a,b | 7.68 b | 0.043 |
| B | 9.24 | 7.48 | 7.36 | 9.42 | 9.15 | 7.20 | 6.08 | 12.34 | 12.92 | 0.083 | |
| C | 9.44 | 7.19 | 7.89 | 9.03 | 8.09 | 7.66 | 5.75 | 9.81 | 9.44 | 0.065 | |
| All | 8.84 b | 6.64 a,b | 7.10 a,b | 8.74 b | 8.39 a,b | 7.11 a,b | 6.03 a | 9.50 a,b | 10.20 b | <0.001 | |
| 50 | A | 10.72 a,b,c | 8.26 a | 9.27 a | 11.04 a,b,c | 9.63 a,b | 8.78 a | 8.49 a | 16.23 b,c | 16.87 c | 0.029 |
| B | 13.62 a,b | 15.90 a,b | 18.25 a,b | 11.80 a | 13.54 a,b | 11.53 a | 12.05 a | 24.00 b | 24.00 b | <0.001 | |
| C | 11.74 a,b | 8.53 a,b | 7.92 a | 11.43 a,b | 10.33 a,b | 8.29 a,b | 7.93 a | 16.41 a,b | 19.21 b | 0.011 | |
| All | 12.03 a,b,c | 10.90 a | 11.81 a,b | 11.42 a,b,c | 11.17 a,b | 9.54 a | 9.49 a | 18.88 b,c | 20.02 c | <0.001 | |
| All | All | 10.43 a,b,c | 8.77 a | 9.45 a,b | 10.08 a,b,c | 9.78 a,b,c | 8.32 a | 7.76 a | 14.19 b,c | 15.02 c | <0.001 |
| p-value age | 0.004 | 0.004 | 0.010 | 0.002 | 0.009 | 0.005 | 0.004 | <0.001 | <0.001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuhrmann, L.; Vahjen, W.; Zentek, J.; Günther, R.; Saliu, E.-M. The Impact of Pre- and Probiotic Product Combinations on Ex vivo Growth of Avian Pathogenic Escherichia coli and Salmonella Enteritidis. Microorganisms 2022, 10, 121. https://doi.org/10.3390/microorganisms10010121
Fuhrmann L, Vahjen W, Zentek J, Günther R, Saliu E-M. The Impact of Pre- and Probiotic Product Combinations on Ex vivo Growth of Avian Pathogenic Escherichia coli and Salmonella Enteritidis. Microorganisms. 2022; 10(1):121. https://doi.org/10.3390/microorganisms10010121
Chicago/Turabian StyleFuhrmann, Laura, Wilfried Vahjen, Jürgen Zentek, Ronald Günther, and Eva-Maria Saliu. 2022. "The Impact of Pre- and Probiotic Product Combinations on Ex vivo Growth of Avian Pathogenic Escherichia coli and Salmonella Enteritidis" Microorganisms 10, no. 1: 121. https://doi.org/10.3390/microorganisms10010121