
Time-Course Lipidomics of Ornithine-Induced Severe Acute Pancreatitis Model Reveals the Free Fatty Acids Centered Lipids Dysregulation Characteristics

 ,
,
Abstract
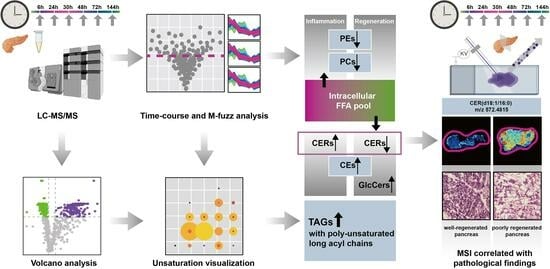
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animal Ethical Clearance, Acquisition, and Husbandry
2.2. Animal Experiments and Sample Collection
2.3. Histopathology and Biochemical Indexes Assessment
2.4. Lipidomics Study
2.5. Mass Spectrometry Imaging
3. Results
3.1. Time-Course Biochemical Indexes and Pathological Changes of SAP Induced by L-Ornithine
3.2. Time-Course Change in Lipids Perturbations in the Pancreas during Orn-SAP
3.3. Lipids Changed at Certain Timepoints in the Pancreas during Orn-SAP
3.4. Time-Course Change in Lipids Perturbations in the Serum during Orn-SAP
3.5. FFAs-Centered Lipids Perturbations in the Pancreas during Orn-SAP
3.6. Histopathological Heterogeneity Findings Correlated with In Situ Lipid Levels of CERs at 144 h of SAP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mederos, M.A.; Reber, H.A.; Girgis, M.D. Acute Pancreatitis: A Review. JAMA 2021, 325, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Banks, P.A.; Bollen, T.L.; Dervenis, C.; Gooszen, H.G.; Johnson, C.D.; Sarr, M.G.; Tsiotos, G.G.; Vege, S.S. Classification of acute pancreatitis−−2012: Revision of the Atlanta classification and definitions by international consensus. Gut 2013, 62, 102–111. [Google Scholar] [CrossRef]
- Garg, P.K.; Singh, V.P. Organ Failure Due to Systemic Injury in Acute Pancreatitis. Gastroenterology 2019, 156, 2008–2023. [Google Scholar] [CrossRef] [PubMed]
- Barreto, S.G.; Habtezion, A.; Gukovskaya, A.; Lugea, A.; Jeon, C.; Yadav, D.; Hegyi, P.; Venglovecz, V.; Sutton, R.; Pandol, S.J. Critical thresholds: Key to unlocking the door to the prevention and specific treatments for acute pancreatitis. Gut 2021, 70, 194–203. [Google Scholar] [CrossRef]
- Kui, B.; Balla, Z.; Végh, E.T.; Pallagi, P.; Venglovecz, V.; Iványi, B.; Takács, T.; Hegyi, P.; Rakonczay, Z., Jr. Recent advances in the investigation of pancreatic inflammation induced by large doses of basic amino acids in rodents. Lab. Investig. 2014, 94, 138–149. [Google Scholar] [CrossRef]
- van Westerloo, D.J. The better the model, the nearer the cure. Crit. Care Med. 2008, 36, 2208–2209. [Google Scholar] [CrossRef] [PubMed]
- Walter, F.R.; Harazin, A.; Tóth, A.E.; Veszelka, S.; Santa-Maria, A.R.; Barna, L.; Kincses, A.; Biczó, G.; Balla, Z.; Kui, B.; et al. Blood-brain barrier dysfunction in L-ornithine induced acute pancreatitis in rats and the direct effect of L-ornithine on cultured brain endothelial cells. Fluids Barriers CNS 2022, 19, 16. [Google Scholar] [CrossRef]
- Yang, J.; Shi, N.; Wang, S.; Wang, M.; Huang, Y.; Wang, Y.; Liang, G.; Yang, J.; Rong, J.; Ma, Y.; et al. Multi-dimensional metabolomic profiling reveals dysregulated ornithine metabolism hallmarks associated with a severe acute pancreatitis phenotype. Transl. Res. 2023; in press. [Google Scholar] [CrossRef]
- Biczó, G.; Hegyi, P.; Sinervirta, R.; Berczi, S.; Dósa, S.; Siska, A.; Iványi, B.; Venglovecz, V.; Takács, T.; Alhonen, L.; et al. Characterization of polyamine homeostasis in l-ornithine-induced acute pancreatitis in rats. Pancreas 2010, 39, 1047–1056. [Google Scholar] [CrossRef]
- Peng, Y.; Hong, J.; Raftery, D.; Xia, Q.; Du, D. Metabolomic-based clinical studies and murine models for acute pancreatitis disease: A review. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2021, 1867, 166123. [Google Scholar] [CrossRef]
- Martin-Perez, M.; Urdiroz-Urricelqui, U.; Bigas, C.; Benitah, S.A. The role of lipids in cancer progression and metastasis. Cell Metab. 2022, 34, 1675–1699. [Google Scholar] [CrossRef]
- Wagner, R.; Eckstein, S.S.; Yamazaki, H.; Gerst, F.; Machann, J.; Jaghutriz, B.A.; Schürmann, A.; Solimena, M.; Singer, S.; Königsrainer, A.; et al. Metabolic implications of pancreatic fat accumulation. Nat. Rev. Endocrinol. 2022, 18, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.L.; McNabb-Baltar, J. Hypertriglyceridemia and acute pancreatitis. Pancreatology 2020, 20, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Valdivielso, P.; Ramírez-Bueno, A.; Ewald, N. Current knowledge of hypertriglyceridemic pancreatitis. Eur. J. Intern. Med. 2014, 25, 689–694. [Google Scholar] [CrossRef]
- Dong, X.; Pan, S.; Zhang, D.; Hong, W.; Chen, T.; Zhang, B.; Huang, Z.; Chen, C. Hyperlipemia pancreatitis onset time affects the association between elevated serum triglyceride levels and disease severity. Lipids Health Dis. 2022, 21, 49. [Google Scholar] [CrossRef]
- Hidalgo, N.J.; Pando, E.; Alberti, P.; Mata, R.; Fernandes, N.; Adell, M.; Villasante, S.; Blanco, L.; Balsells, J.; Charco, R. The role of high serum triglyceride levels on pancreatic necrosis development and related complications. BMC Gastroenterol. 2023, 23, 51. [Google Scholar] [CrossRef]
- Navina, S.; Acharya, C.; DeLany, J.P.; Orlichenko, L.S.; Baty, C.J.; Shiva, S.S.; Durgampudi, C.; Karlsson, J.M.; Lee, K.; Bae, K.T.; et al. Lipotoxicity causes multisystem organ failure and exacerbates acute pancreatitis in obesity. Sci. Transl. Med. 2011, 3, 107ra110. [Google Scholar] [CrossRef]
- Yang, N.; Li, B.; Pan, Y.; Tu, J.; Liu, G.; Lu, G.; Li, W. Hypertriglyceridaemia delays pancreatic regeneration after acute pancreatitis in mice and patients. Gut 2019, 68, 378–380. [Google Scholar] [CrossRef]
- Khatua, B.; El-Kurdi, B.; Patel, K.; Rood, C.; Noel, P.; Crowell, M.; Yaron, J.R.; Kostenko, S.; Guerra, A.; Faigel, D.O.; et al. Adipose saturation reduces lipotoxic systemic inflammation and explains the obesity paradox. Sci. Adv. 2021, 7, eabd6449. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Y.; Tao, Y.; Hu, L.; Qiu, Q.; Pu, Q.; Yang, J.; Wang, S.; Huang, Y.; Chen, X.; et al. Temporal Proteomic and Lipidomic Profiles of Cerulein-Induced Acute Pancreatitis Reveal Novel Insights for Metabolic Alterations in the Disease Pathogenesis. ACS Omega 2023, 8, 12310–12326. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. The foundations and development of lipidomics. J. Lipid Res. 2022, 63, 100164. [Google Scholar] [CrossRef]
- Ahluwalia, K.; Ebright, B.; Chow, K.; Dave, P.; Mead, A.; Poblete, R.; Louie, S.G.; Asante, I. Lipidomics in Understanding Pathophysiology and Pharmacologic Effects in Inflammatory Diseases: Considerations for Drug Development. Metabolites 2022, 12, 333. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Han, X. Lipidomics: Techniques, Applications, and Outcomes Related to Biomedical Sciences. Trends Biochem. Sci. 2016, 41, 954–969. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, B.; Lam, S.M.; Shui, G. Integration of lipidomics and metabolomics for in-depth understanding of cellular mechanism and disease progression. J. Genet. Genom. 2020, 47, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xue, C. Ghrelin inhibits the development of acute pancreatitis and nuclear factor kappaB activation in pancreas and liver. Pancreas 2009, 38, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Rakonczay, Z., Jr.; Hegyi, P.; Dósa, S.; Iványi, B.; Jármay, K.; Biczó, G.; Hracskó, Z.; Varga, I.S.; Karg, E.; Kaszaki, J.; et al. A new severe acute necrotizing pancreatitis model induced by L-ornithine in rats. Crit. Care Med. 2008, 36, 2117–2127. [Google Scholar] [CrossRef] [PubMed]
- Nathan, J.D.; Romac, J.; Peng, R.Y.; Peyton, M.; Macdonald, R.J.; Liddle, R.A. Transgenic expression of pancreatic secretory trypsin inhibitor-I ameliorates secretagogue-induced pancreatitis in mice. Gastroenterology 2005, 128, 717–727. [Google Scholar] [CrossRef]
- Dawra, R.; Ku, Y.S.; Sharif, R.; Dhaulakhandi, D.; Phillips, P.; Dudeja, V.; Saluja, A.K. An improved method for extracting myeloperoxidase and determining its activity in the pancreas and lungs during pancreatitis. Pancreas 2008, 37, 62–68. [Google Scholar] [CrossRef]
- Zang, Q.; Wang, M.; Zhu, Y.; Wang, L.; Luo, Z.; Li, X.; He, J.; Zhang, R.; Abliz, Z. Enhanced On-Tissue Chemical Derivatization with Hydrogel Assistance for Mass Spectrometry Imaging. Anal. Chem. 2021, 93, 15373–15380. [Google Scholar] [CrossRef]
- Biczo, G.; Vegh, E.T.; Shalbueva, N.; Mareninova, O.A.; Elperin, J.; Lotshaw, E.; Gretler, S.; Lugea, A.; Malla, S.R.; Dawson, D.; et al. Mitochondrial Dysfunction, Through Impaired Autophagy, Leads to Endoplasmic Reticulum Stress, Deregulated Lipid Metabolism, and Pancreatitis in Animal Models. Gastroenterology 2018, 154, 689–703. [Google Scholar] [CrossRef]
- Kiss, L.; Fűr, G.; Pisipati, S.; Rajalingamgari, P.; Ewald, N.; Singh, V.; Rakonczay, Z., Jr. Mechanisms linking hypertriglyceridemia to acute pancreatitis. Acta Physiol. 2023, 237, e13916. [Google Scholar] [CrossRef]
- Chang, T.Y.; Chang, C.C.; Cheng, D. Acyl-coenzyme A:cholesterol acyltransferase. Annu. Rev. Biochem. 1997, 66, 613–638. [Google Scholar] [CrossRef]
- Andrieu-Abadie, N.; Gouazé, V.; Salvayre, R.; Levade, T. Ceramide in apoptosis signaling: Relationship with oxidative stress. Free. Radic. Biol. Med. 2001, 31, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chiang, Y.P.; He, M.; Worgall, T.S.; Zhou, H.; Jiang, X.C. Liver sphingomyelin synthase 1 deficiency causes steatosis, steatohepatitis, fibrosis, and tumorigenesis: An effect of glucosylceramide accumulation. iScience 2021, 24, 103449. [Google Scholar] [CrossRef]
- Zimmermann, A.; Gloor, B.; Kappeler, A.; Uhl, W.; Friess, H.; Büchler, M.W. Pancreatic stellate cells contribute to regeneration early after acute necrotising pancreatitis in humans. Gut 2002, 51, 574–578. [Google Scholar] [CrossRef]
- Ho, Q.W.C.; Zheng, X.; Ali, Y. Ceramide Acyl Chain Length and Its Relevance to Intracellular Lipid Regulation. Int. J. Mol. Sci. 2022, 23, 9697. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, B.; Summers, S.A. Ceramides in Metabolism: Key Lipotoxic Players. Annu. Rev. Physiol. 2021, 83, 303–330. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Wang, M.; Qiu, Q.; Huang, Y.; Wang, Y.; Pu, Q.; Jiang, N.; Wang, R.; Wen, L.; Zhang, X.; et al. Time-Course Lipidomics of Ornithine-Induced Severe Acute Pancreatitis Model Reveals the Free Fatty Acids Centered Lipids Dysregulation Characteristics. Metabolites 2023, 13, 993. https://doi.org/10.3390/metabo13090993
Yang J, Wang M, Qiu Q, Huang Y, Wang Y, Pu Q, Jiang N, Wang R, Wen L, Zhang X, et al. Time-Course Lipidomics of Ornithine-Induced Severe Acute Pancreatitis Model Reveals the Free Fatty Acids Centered Lipids Dysregulation Characteristics. Metabolites. 2023; 13(9):993. https://doi.org/10.3390/metabo13090993
Chicago/Turabian StyleYang, Jinxi, Manjiangcuo Wang, Qi Qiu, Yan Huang, Yiqin Wang, Qianlun Pu, Na Jiang, Rui Wang, Li Wen, Xiaoying Zhang, and et al. 2023. "Time-Course Lipidomics of Ornithine-Induced Severe Acute Pancreatitis Model Reveals the Free Fatty Acids Centered Lipids Dysregulation Characteristics" Metabolites 13, no. 9: 993. https://doi.org/10.3390/metabo13090993