
Medium Roasting and Brewing Methods Differentially Modulate Global Metabolites, Lipids, Biogenic Amines, Minerals, and Antioxidant Capacity of Hawai‘i-Grown Coffee (Coffea arabica)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Green Coffee Grounds
2.2. Coffee Roasting
2.3. Drip Filter Paper (FP) Method
2.4. Drip Metallic Filter Mesh (FM) Method
2.5. French Press (FrP) Method
2.6. Cold Brew (CB) Method
2.7. Brewed Coffee Omics (Global Metabolites, Lipids, and Biogenic Amines)
2.8. Mineral and Metal Analysis of Brewed Coffee
2.9. Antioxidant Capacity of Brewed Coffee
2.10. Statistical Data Analysis
3. Results
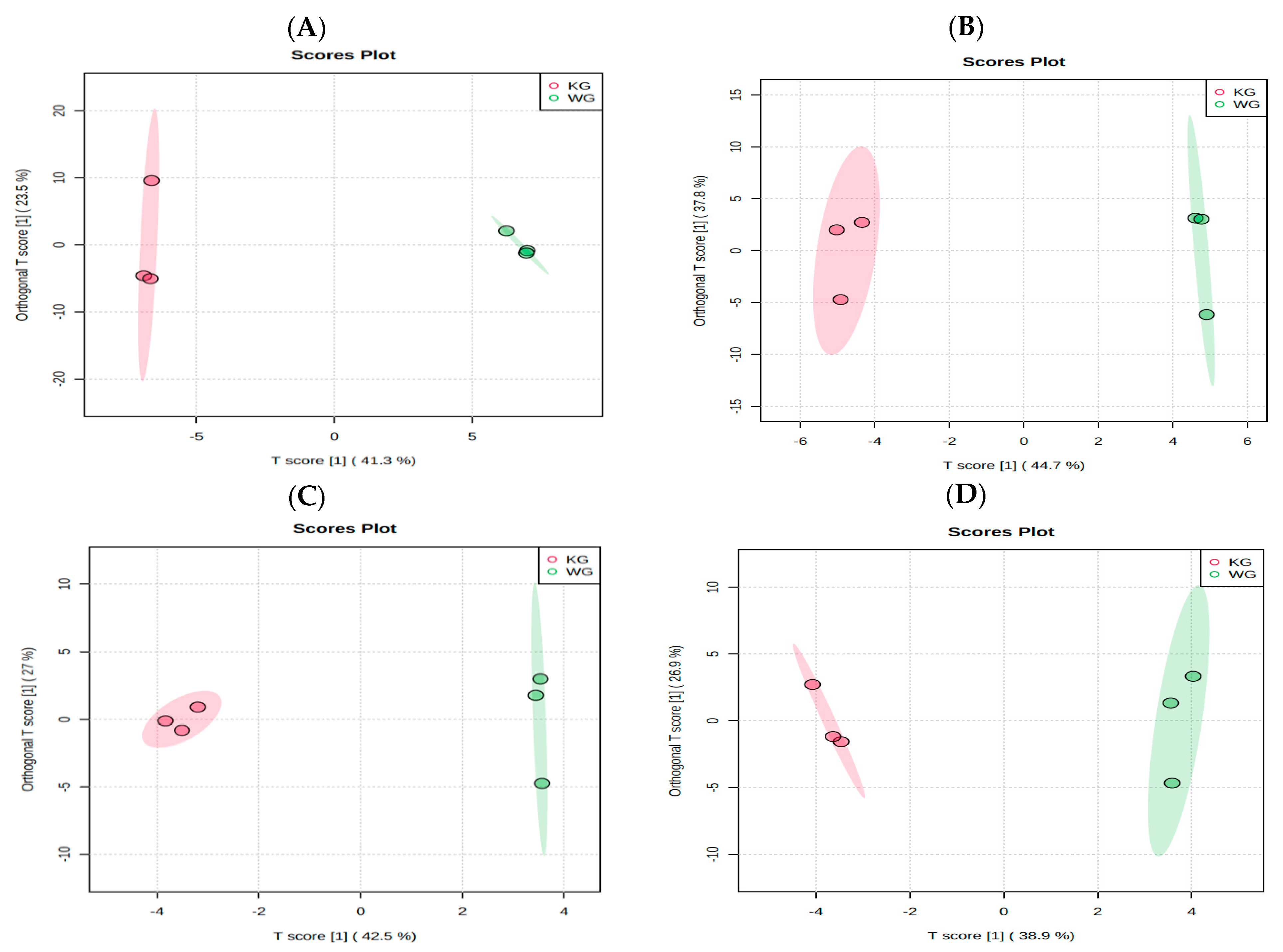
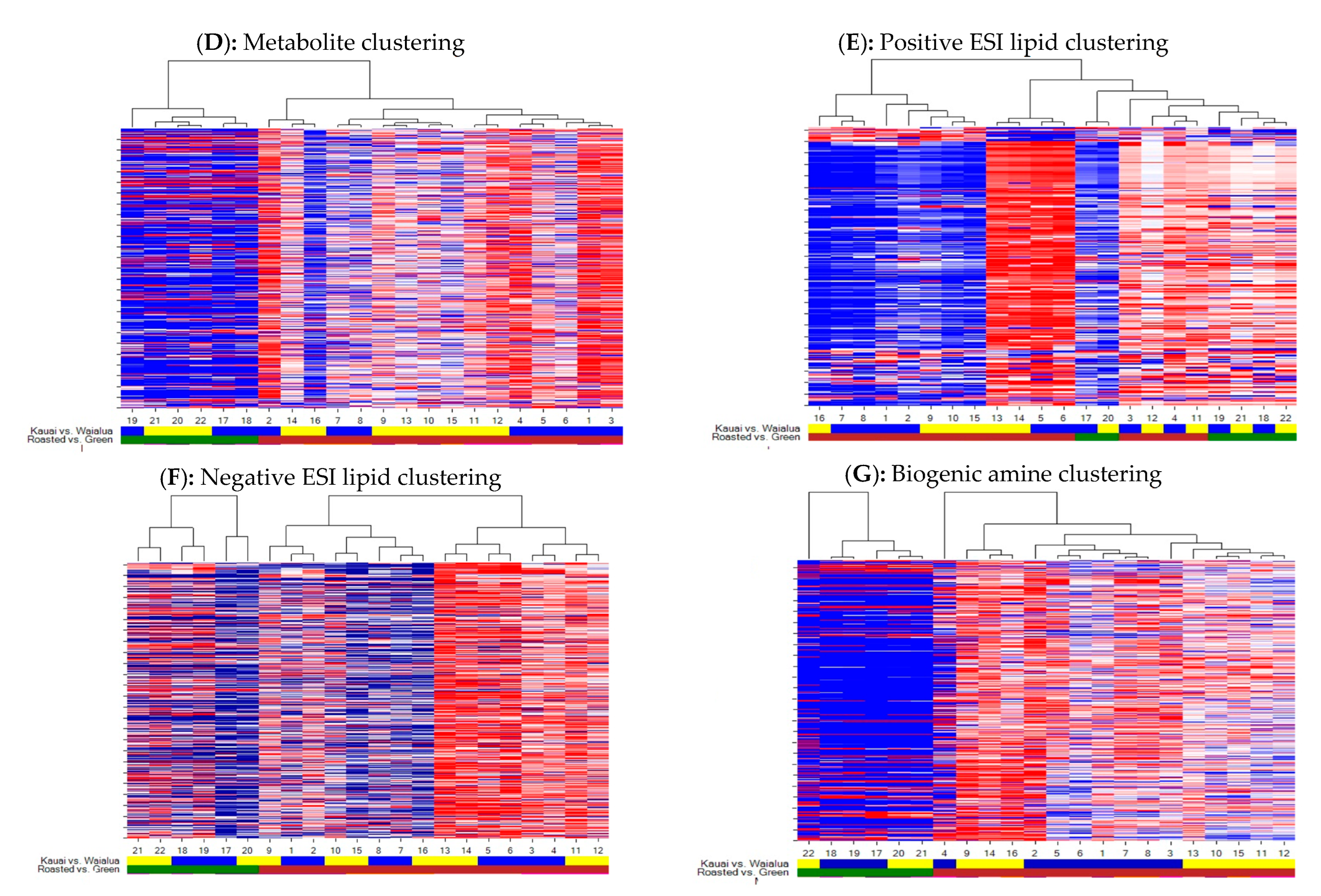
3.1. Differences in Global Metabolites, Lipids, and Biogenic Amines among the Green and Roasted Coffee Varieties of “Kona Typica” (Waialua) and “Yellow Catuai” (Kaua’i) Brews
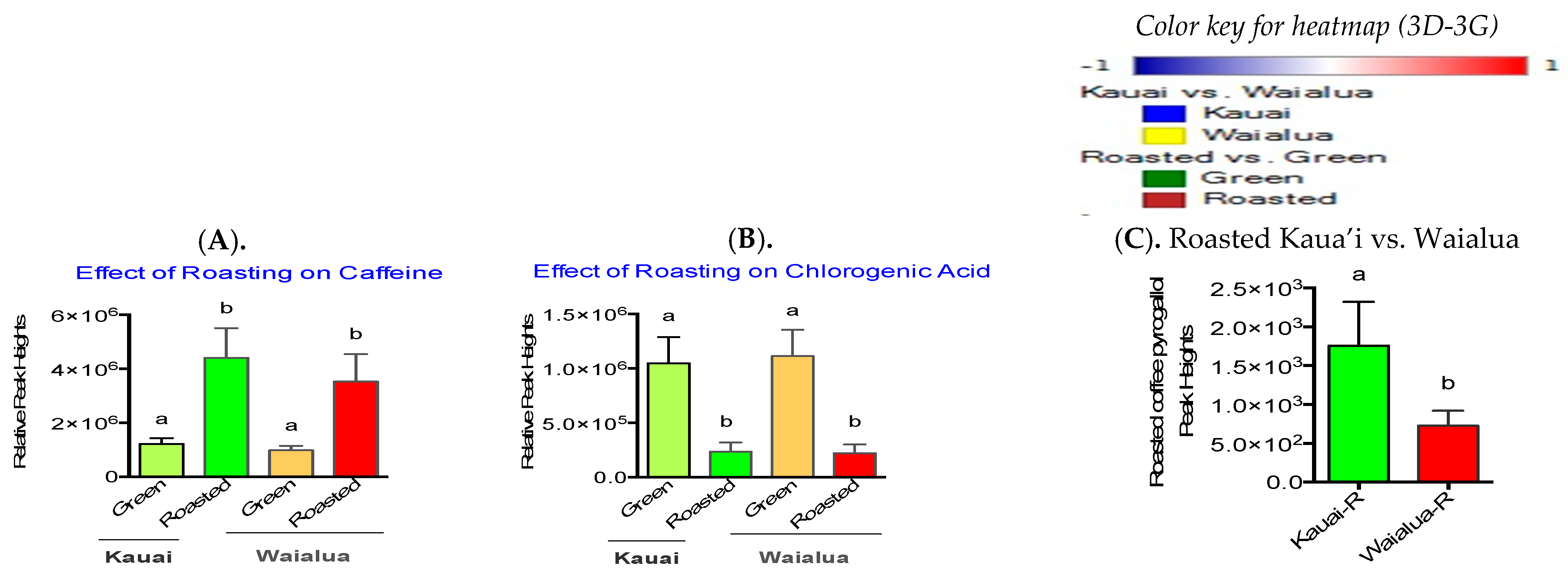
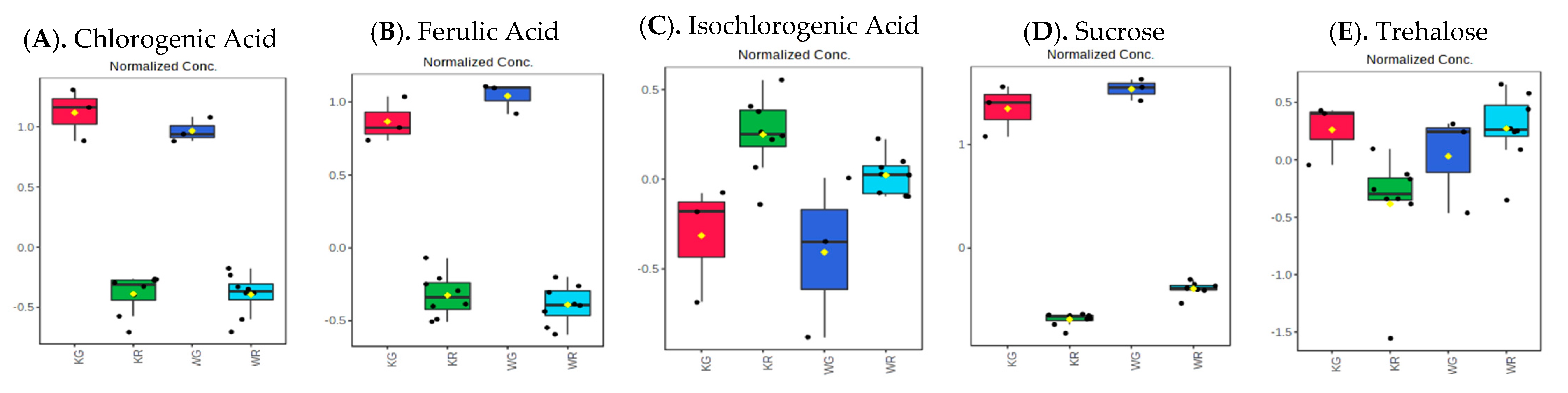
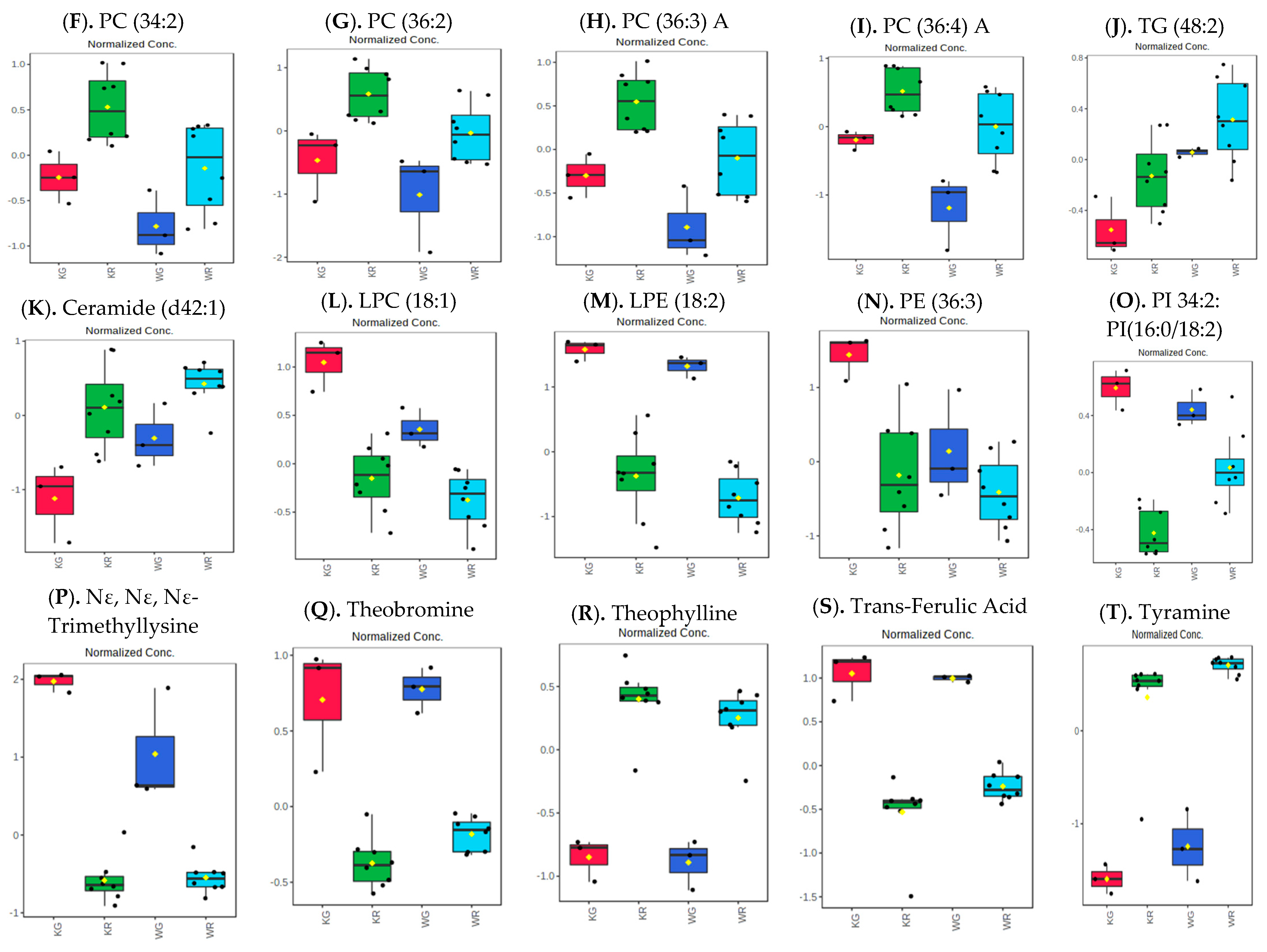
3.2. Influence of Roasting on Metabolites, Lipids, and Biogenic Amines of “Kona Typica” (Waialua) and “Yellow Catuai” (Kaua’i) Coffee Brews
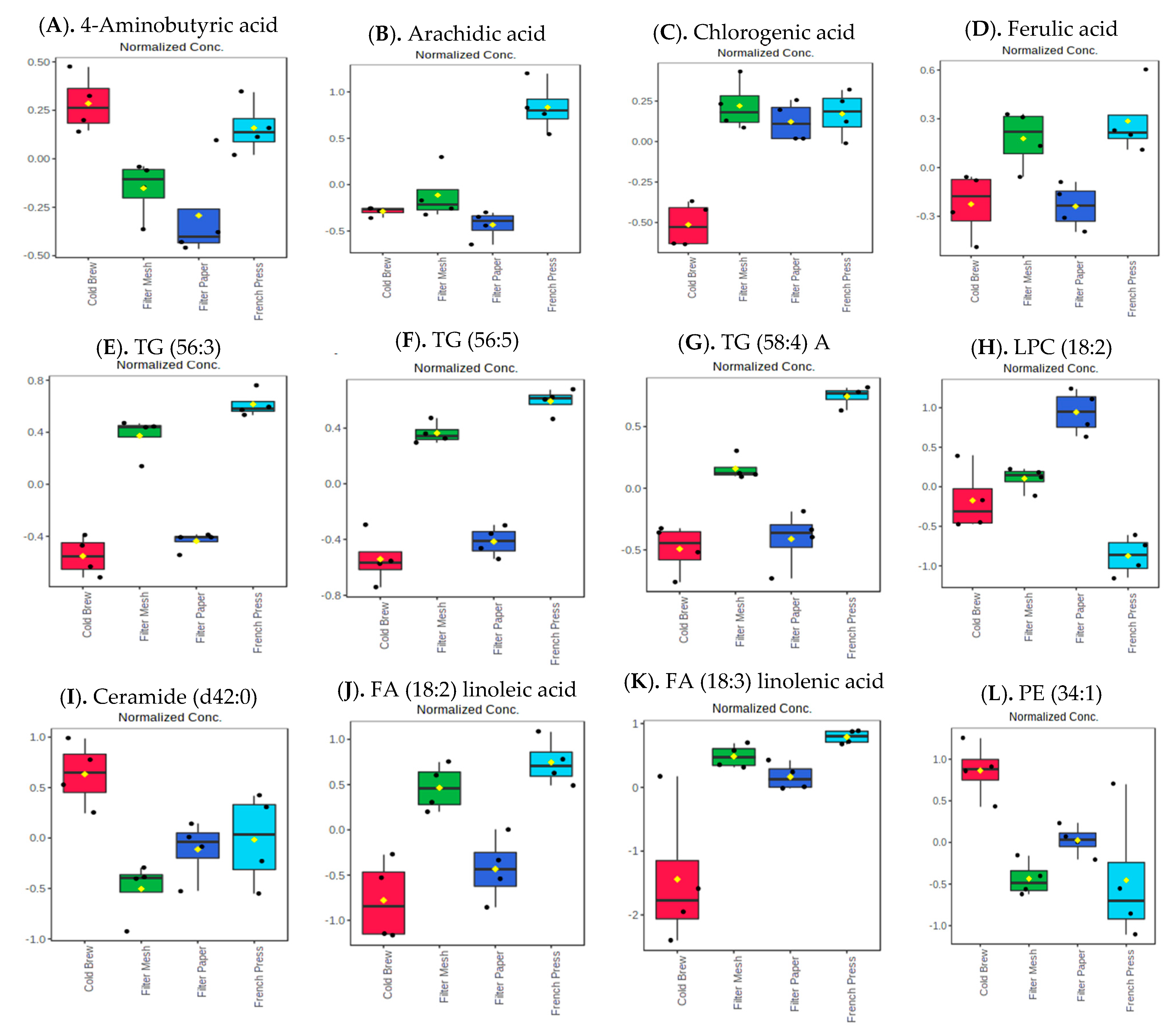
3.3. Influence of Brewing Methods on Metabolites, Lipids, and Biogenic Amines of “Kona Typica” (Waialua) and “Yellow Catuai” (Kaua’i) Coffee Brews
3.4. Mineral and Metal Analysis of “Kona Typica” (Waialua) and “Yellow Catuai” (Kaua’i) Coffee Brews
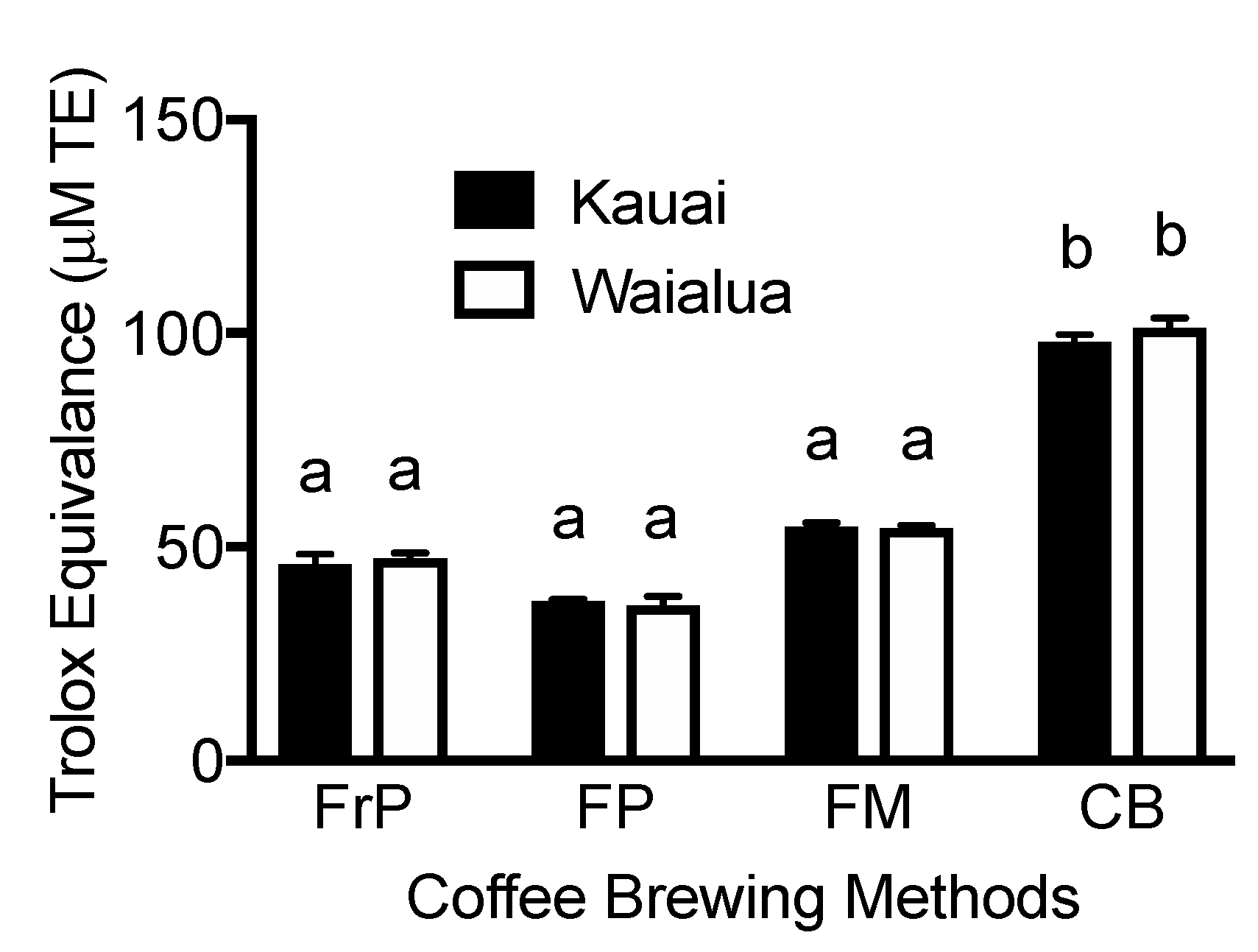
3.5. Antioxidant Capacity of “Kona Typica” (Waialua) and “Yellow Catuai” (Kaua’i) Coffee Brews
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- News, L.A. Global Coffee Production Expected to Recover in 2022/23 with Brazil Crop. Available online: https://www.riotimesonline.com/brazil-news/rio-business/global-coffee-production-expected-to-recover-in-2022-23-with-brazil-crop/ (accessed on 5 January 2023).
- Halstead, T. Coffee: World Markets and Trade. Worls Production, Markets and Trade Report. Available online: https://apps.fas.usda.gov/psdonline/circulars/coffee.pdf (accessed on 5 January 2023).
- Gunter, M. 22 Coffee Consumption Statistics and Facts in 2022: Fascinating Data! Available online: https://coffeeaffection.com/coffee-consumption-statistics/ (accessed on 5 January 2023).
- Nerurkar, P.V.; Gandhi, K.; Chen, J.J. Correlations between Coffee Consumption and Metabolic Phenotypes, Plasma Folate, and Vitamin B12: NHANES 2003 to 2006. Nutrients 2021, 13, 1348. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourshahidi, L.K.; Navarini, L.; Petracco, M.; Strain, J.J. A Comprehensive Overview of the Risks and Benefits of Coffee Consumption. Compr. Rev. Food Sci. Food Saf. 2016, 15, 671–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, R.M.; Lima, D.R. Coffee consumption, obesity and type 2 diabetes: A mini-review. Eur. J. Nutr. 2016, 55, 1345–1358. [Google Scholar] [CrossRef]
- Cho, H.J.; Yoo, J.Y.; Kim, A.N.; Moon, S.; Choi, J.; Kim, I.; Ko, K.P.; Lee, J.E.; Park, S.K. Association of coffee drinking with all-cause and cause-specific mortality in over 190,000 individuals: Data from two prospective studies. Int. J. Food Sci. Nutr. 2022, 73, 513–521. [Google Scholar] [CrossRef]
- Liu, D.; Li, Z.H.; Shen, D.; Zhang, P.D.; Song, W.Q.; Zhang, W.T.; Huang, Q.M.; Chen, P.L.; Zhang, X.R.; Mao, C. Association of Sugar-Sweetened, Artificially Sweetened, and Unsweetened Coffee Consumption with All-Cause and Cause-Specific Mortality: A Large Prospective Cohort Study. Ann. Intern. Med. 2022, 175, 909–917. [Google Scholar] [CrossRef]
- Surma, S.; Banach, M. Coffee and caffeine consumption and overall mortality. Pleasure with restrictions-where do we really stand in 2022? Nutrition 2022, 102, 111747. [Google Scholar] [CrossRef]
- Zheng, H.; Lin, F.; Xin, N.; Yang, L.; Zhu, P. Association of Coffee, Tea, and Caffeine Consumption with All-Cause Risk and Specific Mortality for Cardiovascular Disease Patients. Front. Nutr. 2022, 9, 842856. [Google Scholar] [CrossRef]
- Nordestgaard, A.T. Causal relationship from coffee consumption to diseases and mortality: A review of observational and Mendelian randomization studies including cardiometabolic diseases, cancer, gallstones and other diseases. Eur. J. Nutr. 2022, 61, 573–587. [Google Scholar] [CrossRef]
- Sakamaki, T.; Kayaba, K.; Kotani, K.; Namekawa, M.; Hamaguchi, T.; Nakaya, N.; Ishikawa, S. Coffee consumption and mortality in Japan with 18 years of follow-up: The Jichi Medical School Cohort Study. Public Health 2021, 191, 23–30. [Google Scholar] [CrossRef]
- Curb, J.D.; Reed, D.M.; Kautz, J.A.; Yano, K. Coffee, caffeine, and serum cholesterol in Japanese men in Hawaii. Am. J. Epidemiol. 1986, 123, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.R.; Curb, J.D.; Borhani, N.O.; Prineas, R.J.; Molteni, A. Coffee consumption and serum cholesterol in the hypertension detection and follow-up program. Am. J. Epidemiol. 1988, 128, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Hallstrom, H.; Byberg, L.; Glynn, A.; Lemming, E.W.; Wolk, A.; Michaelsson, K. Long-term coffee consumption in relation to fracture risk and bone mineral density in women. Am. J. Epidemiol. 2013, 178, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Kujawska, A.; Kujawski, S.; Hajec, W.; Skierkowska, N.; Kwiatkowska, M.; Husejko, J.; Newton, J.L.; Simoes, J.A.; Zalewski, P.; Kedziora-Kornatowska, K. Coffee Consumption and Blood Pressure: Results of the Second Wave of the Cognition of Older People, Education, Recreational Activities, Nutrition, Comorbidities, and Functional Capacity Studies (COPERNICUS). Nutrients 2021, 13, 3372. [Google Scholar] [CrossRef]
- Greenop, K.R.; Miller, M.; Attia, J.; Ashton, L.J.; Cohn, R.; Armstrong, B.K.; Milne, E. Maternal consumption of coffee and tea during pregnancy and risk of childhood brain tumors: Results from an Australian case-control study. Cancer Causes Control. 2014, 25, 1321–1327. [Google Scholar] [CrossRef]
- Cheng, J.; Su, H.; Zhu, R.; Wang, X.; Peng, M.; Song, J.; Fan, D. Maternal coffee consumption during pregnancy and risk of childhood acute leukemia: A metaanalysis. Am. J. Obstet. Gynecol. 2014, 210, 151.e1–151.e10. [Google Scholar] [CrossRef]
- Bonaventure, A.; Rudant, J.; Goujon-Bellec, S.; Orsi, L.; Leverger, G.; Baruchel, A.; Bertrand, Y.; Nelken, B.; Pasquet, M.; Michel, G.; et al. Childhood acute leukemia, maternal beverage intake during pregnancy, and metabolic polymorphisms. Cancer Causes Control. 2013, 24, 783–793. [Google Scholar] [CrossRef]
- Thomopoulos, T.P.; Ntouvelis, E.; Diamantaras, A.A.; Tzanoudaki, M.; Baka, M.; Hatzipantelis, E.; Kourti, M.; Polychronopoulou, S.; Sidi, V.; Stiakaki, E.; et al. Maternal and childhood consumption of coffee, tea and cola beverages in association with childhood leukemia: A meta-analysis. Cancer Epidemiol. 2015, 39, 1047–1059. [Google Scholar] [CrossRef]
- Virtanen, S.M.; Uusitalo, L.; Kenward, M.G.; Nevalainen, J.; Uusitalo, U.; Kronberg-Kippila, C.; Ovaskainen, M.L.; Arkkola, T.; Niinisto, S.; Hakulinen, T.; et al. Maternal food consumption during pregnancy and risk of advanced beta-cell autoimmunity in the offspring. Pediatr. Diabetes 2011, 12, 95–99. [Google Scholar] [CrossRef]
- Pei, L.; Kang, Y.; Cheng, Y.; Yan, H. The Association of Maternal Lifestyle with Birth Defects in Shaanxi Province, Northwest China. PLoS ONE 2015, 10, e0139452. [Google Scholar] [CrossRef] [Green Version]
- Parazzini, F.; Chiaffarino, F.; Chatenoud, L.; Tozzi, L.; Cipriani, S.; Chiantera, V.; Fedele, L. Maternal coffee drinking in pregnancy and risk of small for gestational age birth. Eur. J. Clin. Nutr. 2005, 59, 299–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, L.M.; Lonnerdal, B.; Keen, C.L.; Dewey, K.G. Coffee consumption as a factor in iron deficiency anemia among pregnant women and their infants in Costa Rica. Am. J. Clin. Nutr. 1998, 48, 645–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bech, B.H.; Nohr, E.A.; Vaeth, M.; Henriksen, T.B.; Olsen, J. Coffee and fetal death: A cohort study with prospective data. Am. J. Epidemiol. 2005, 162, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Wisborg, K.; Kesmodel, U.; Bech, B.H.; Hedegaard, M.; Henriksen, T.B. Maternal consumption of coffee during pregnancy and stillbirth and infant death in first year of life: Prospective study. BMJ 2003, 326, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leviton, A. Biases Inherent in Studies of Coffee Consumption in Early Pregnancy and the Risks of Subsequent Events. Nutrients 2018, 10, 1152. [Google Scholar] [CrossRef] [Green Version]
- Restuccia, D.; Spizzirri, U.G.; Parisi, O.I.; Cirillo, G.; Picci, N. Brewing effect on levels of biogenic amines in different coffee samples as determined by LC-UV. Food Chem. 2015, 175, 143–150. [Google Scholar] [CrossRef]
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Torres, J.; Falconi, C.; Moccand, C.; Weckx, S.; De Vuyst, L. Following Coffee Production from Cherries to Cup: Microbiological and Metabolomic Analysis of Wet Processing of Coffea arabica. Appl. Environ. Microbiol. 2019, 85, e02635-18. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.Z.A.; Lau, H.; Lim, S.Y.; Li, S.F.Y.; Liu, S.Q. Untargeted LC-QTOF-MS/MS based metabolomics approach for revealing bioactive components in probiotic fermented coffee brews. Food Res. Int. 2021, 149, 110656. [Google Scholar] [CrossRef]
- Hu, G.L.; Wang, X.; Zhang, L.; Qiu, M.H. The sources and mechanisms of bioactive ingredients in coffee. Food Funct. 2019, 10, 3113–3126. [Google Scholar] [CrossRef]
- Kim, C.H.; Park, S.J.; Yu, J.S.; Lee, D.Y. Interactive effect of post-harvest processing method, roasting degree, and brewing method on coffee metabolite profiles. Food Chem. 2022, 397, 133749. [Google Scholar] [CrossRef]
- Nosal, B.M.; Sakaki, J.R.; Kim, D.O.; Chun, O.K. Impact of coffee preparation on total phenolic content in brewed coffee extracts and their contribution to the body’s antioxidant status. Food Sci. Biotechnol. 2022, 31, 1081–1088. [Google Scholar] [CrossRef]
- Ciaramelli, C.; Palmioli, A.; Airoldi, C. Coffee variety, origin and extraction procedure: Implications for coffee beneficial effects on human health. Food Chem. 2019, 278, 47–55. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Loftfield, E.; Wedekind, R.; Freedman, N.; Kambanis, C.; Scalbert, A.; Sinha, R. A Metabolomic Study of the Variability of the Chemical Composition of Commonly Consumed Coffee Brews. Metabolites 2019, 9, 17. [Google Scholar] [CrossRef]
- Chang, K.L.; Ho, P.C. Gas chromatography time-of-flight mass spectrometry (GC-TOF-MS)-based metabolomics for comparison of caffeinated and decaffeinated coffee and its implications for Alzheimer’s disease. PLoS ONE 2014, 9, e104621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amalia, F.; Aditiawati, P.; Yusianto; Putri, S.P.; Fukusaki, E. Gas chromatography/mass spectrometry-based metabolite profiling of coffee beans obtained from different altitudes and origins with various postharvest processing. Metabolomics 2021, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Jumhawan, U.; Putri, S.P.; Yusianto; Bamba, T.; Fukusaki, E. Quantification of coffee blends for authentication of Asian palm civet coffee (Kopi Luwak) via metabolomics: A proof of concept. J. Biosci. Bioeng. 2016, 122, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Putri, S.P.; Irifune, T.; Yusianto; Fukusaki, E. GC/MS based metabolite profiling of Indonesian specialty coffee from different species and geographical origin. Metabolomics 2019, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Ongo, E.A.; Montevecchi, G.; Antonelli, A.; Sberveglieri, V.; Sevilla Iii, F. Metabolomics fingerprint of Philippine coffee by SPME-GC-MS for geographical and varietal classification. Food Res. Int. 2020, 134, 109227. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, B.; Boulanger, R.; Dussert, S.; Ribeyre, F.; Berthiot, L.; Descroix, F.; Joet, T. Climatic factors directly impact the volatile organic compound fingerprint in green Arabica coffee bean as well as coffee beverage quality. Food Chem. 2012, 135, 2575–2583. [Google Scholar] [CrossRef]
- Ali, A.; Zahid, H.F.; Cottrell, J.J.; Dunshea, F.R. A Comparative Study for Nutritional and Phytochemical Profiling of Coffea arabica (C. arabica) from Different Origins and Their Antioxidant Potential and Molecular Docking. Molecules 2022, 27, 5126. [Google Scholar] [CrossRef]
- Villarreal, D.; Laffargue, A.; Posada, H.; Bertrand, B.; Lashermes, P.; Dussert, S. Genotypic and environmental effects on coffee (Coffea arabica L.) bean fatty acid profile: Impact on variety and origin chemometric determination. J. Agric. Food Chem. 2009, 57, 11321–11327. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Zayed, A.; Sallam, I.E.; Abdelwareth, A.; Wessjohann, L.A. Metabolomics-Based Approach for Coffee Beverage Improvement in the Context of Processing, Brewing Methods, and Quality Attributes. Foods 2022, 11, 864. [Google Scholar] [CrossRef] [PubMed]
- Tassew, A.A.; Yadessa, G.B.; Bote, A.D.; Obso, T.K. Influence of location, elevation gradients, processing methods, and soil quality on the physical and cup quality of coffee in the Kafa Biosphere Reserve of SW Ethiopia. Heliyon 2021, 7, e07790. [Google Scholar] [CrossRef] [PubMed]
- van Mullem, J.J.; de Sousa Bueno Filho, J.S.; Dias, D.R.; Schwan, R.F. Chemical and sensory characterization of coffee from Coffea arabica cv. Mundo Novo and cv. Catuai Vermelho obtained by four different post-harvest processing methods. J. Sci. Food Agric. 2022, 102, 6687–6695. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Yang, M.; Zhang, J.; Zhang, L.; Tian, Y.; Li, C.; Bao, L.; Ma, C.; Abd El-Aty, A.M. Feasibility of Ultrasound-Assisted Extraction for Accelerated Cold Brew Coffee Processing: Characterization and Comparison with Conventional Brewing Methods. Front. Nutr. 2022, 9, 849811. [Google Scholar] [CrossRef] [PubMed]
- Niseteo, T.; Komes, D.; Belscak-Cvitanovic, A.; Horzic, D.; Budec, M. Bioactive composition and antioxidant potential of different commonly consumed coffee brews affected by their preparation technique and milk addition. Food Chem. 2012, 134, 1870–1877. [Google Scholar] [CrossRef] [PubMed]
- Villalon-Lopez, N.; Serrano-Contreras, J.I.; Tellez-Medina, D.I.; Gerardo Zepeda, L. An H NMR-based metabolomic approach to compare the chemical profiling of retail samples of ground roasted and instant coffees. Food Res. Int. 2018, 106, 263–270. [Google Scholar] [CrossRef]
- Perez-Miguez, R.; Castro-Puyana, M.; Sanchez-Lopez, E.; Plaza, M.; Marina, M.L. Untargeted HILIC-MS-Based Metabolomics Approach to Evaluate Coffee Roasting Process: Contributing to an Integrated Metabolomics Multiplatform. Molecules 2020, 25, 887. [Google Scholar] [CrossRef] [Green Version]
- Perez-Miguez, R.; Sanchez-Lopez, E.; Plaza, M.; Castro-Puyana, M.; Marina, M.L. A non-targeted metabolomic approach based on reversed-phase liquid chromatography-mass spectrometry to evaluate coffee roasting process. Anal. Bioanal. Chem. 2018, 410, 7859–7870. [Google Scholar] [CrossRef]
- Perez-Miguez, R.; Sanchez-Lopez, E.; Plaza, M.; Marina, M.L.; Castro-Puyana, M. Capillary electrophoresis-mass spectrometry metabolic fingerprinting of green and roasted coffee. J. Chromatogr. A 2019, 1605, 360353. [Google Scholar] [CrossRef]
- Angeloni, G.; Guerrini, L.; Masella, P.; Bellumori, M.; Daluiso, S.; Parenti, A.; Innocenti, M. What kind of coffee do you drink? An investigation on effects of eight different extraction methods. Food Res. Int. 2019, 116, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lao, F.; Xu, Z.; Wang, X.; Chen, F.; Liao, X.; Chen, A.; Yang, S. Use of liquid chromatography quadrupole time-of-flight mass spectrometry and metabolomic approach to discriminate coffee brewed by different methods. Food Chem. 2019, 286, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, X.; Liu, P.; Zhang, T.; Jelderks, J.A.; Corke, H. Preliminary Characterization of Phytochemicals and Polysaccharides in Diverse Coffee Cascara Samples: Identification, Quantification and Discovery of Novel Compounds. Foods 2022, 11, 1710. [Google Scholar] [CrossRef] [PubMed]
- Aditiawati, P.; Astuti, D.I.; Kriswantoro, J.A.; Khanza, S.M.; Kamarisima; Irifune, T.; Amalia, F.; Fukusaki, E.; Putri, S.P. GC/MS-based metabolic profiling for the evaluation of solid state fermentation to improve quality of Arabica coffee beans. Metabolomics 2020, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, N.; Pataquiva, L.; Osorio, C.; Moreno, F.L.M.; Ruiz, R.Y. Effect of grinding, extraction time and type of coffee on the physicochemical and flavour characteristics of cold brew coffee. Sci. Rep. 2019, 9, 8440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressanello, D.; Liberto, E.; Cordero, C.; Rubiolo, P.; Pellegrino, G.; Ruosi, M.R.; Bicchi, C. Coffee aroma: Chemometric comparison of the chemical information provided by three different samplings combined with GC-MS to describe the sensory properties in cup. Food Chem. 2017, 214, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Bressanello, D.; Liberto, E.; Cordero, C.; Sgorbini, B.; Rubiolo, P.; Pellegrino, G.; Ruosi, M.R.; Bicchi, C. Chemometric Modeling of Coffee Sensory Notes through Their Chemical Signatures: Potential and Limits in Defining an Analytical Tool for Quality Control. J. Agric. Food Chem. 2018, 66, 7096–7109. [Google Scholar] [CrossRef]
- Bressanello, D.; Marengo, A.; Cordero, C.; Strocchi, G.; Rubiolo, P.; Pellegrino, G.; Ruosi, M.R.; Bicchi, C.; Liberto, E. Chromatographic Fingerprinting Strategy to Delineate Chemical Patterns Correlated to Coffee Odor and Taste Attributes. J. Agric. Food Chem. 2021, 69, 4550–4560. [Google Scholar] [CrossRef]
- Liberto, E.; Bressanello, D.; Strocchi, G.; Cordero, C.; Ruosi, M.R.; Pellegrino, G.; Bicchi, C.; Sgorbini, B. HS-SPME-MS-Enose Coupled with Chemometrics as an Analytical Decision Maker to Predict In-Cup Coffee Sensory Quality in Routine Controls: Possibilities and Limits. Molecules 2019, 24, 4515. [Google Scholar] [CrossRef] [Green Version]
- Caprioli, G.; Cortese, M.; Sagratini, G.; Vittori, S. The influence of different types of preparation (espresso and brew) on coffee aroma and main bioactive constituents. Int. J. Food Sci. Nutr. 2015, 66, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Janda, K.; Jakubczyk, K.; Baranowska-Bosiacka, I.; Kapczuk, P.; Kochman, J.; Rebacz-Maron, E.; Gutowska, I. Mineral Composition and Antioxidant Potential of Coffee Beverages Depending on the Brewing Method. Foods 2020, 9, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hawary, E.A.; Zayed, A.; Laub, A.; Modolo, L.V.; Wessjohann, L.; Farag, M.A. How Does LC/MS Compare to UV in Coffee Authentication and Determination of Antioxidant Effects? Brazilian and Middle Eastern Coffee as Case Studies. Antioxidants 2022, 11, 131. [Google Scholar] [CrossRef] [PubMed]
- Seremet, D.; Fabecic, P.; Vojvodic Cebin, A.; Mandura Jaric, A.; Pudic, R.; Komes, D. Antioxidant and Sensory Assessment of Innovative Coffee Blends of Reduced Caffeine Content. Molecules 2022, 27, 448. [Google Scholar] [CrossRef] [PubMed]
- Stanek, N.; Zarebska, M.; Bilos, L.; Barabosz, K.; Nowakowska-Bogdan, E.; Semeniuk, I.; Blaszkiewicz, J.; Kulesza, R.; Matejuk, R.; Szkutnik, K. Influence of coffee brewing methods on the chromatographic and spectroscopic profiles, antioxidant and sensory properties. Sci. Rep. 2021, 11, 21377. [Google Scholar] [CrossRef] [PubMed]
- Doo, T.; Morimoto, Y.; Steinbrecher, A.; Kolonel, L.N.; Maskarinec, G. Coffee intake and risk of type 2 diabetes: The Multiethnic Cohort. Public Health Nutr. 2014, 17, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Gelber, R.P.; Petrovitch, H.; Masaki, K.H.; Ross, G.W.; White, L.R. Coffee intake in midlife and risk of dementia and its neuropathologic correlates. J. Alzheimer Dis. 2011, 23, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Ross, G.W.; Abbott, R.D.; Petrovitch, H.; Morens, D.M.; Grandinetti, A.; Tung, K.H.; Tanner, C.M.; Masaki, K.H.; Blanchette, P.L.; Curb, J.D.; et al. Association of coffee and caffeine intake with the risk of Parkinson disease. JAMA 2000, 283, 2674–2679. [Google Scholar] [CrossRef]
- Setiawan, V.W.; Wilkens, L.R.; Lu, S.C.; Hernandez, B.Y.; Le Marchand, L.; Henderson, B.E. Association of coffee intake with reduced incidence of liver cancer and death from chronic liver disease in the US multiethnic cohort. Gastroenterology 2015, 148, 118–125; quiz e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawaii. Available online: https://hfbf.org/mac-nuts/ (accessed on 5 January 2023).
- Top 20 Agricultural Commodities Produced State of Hawaii. Available online: https://hdoa.hawaii.gov/add/files/2021/12/Top-20-Commodities-2019_SOH_01.28.21.pdf (accessed on 5 January 2023).
- Kostenko, A. Coffee Industry Valued Higher Now. Available online: https://hawaiicoffeeassoc.org/News/10075301 (accessed on 5 January 2023).
- Sky, A. 33+ Coffee Industry Statistics [2022]: Cafes, Consumption, And Market Trends. Available online: https://www.zippia.com/advice/coffee-industry-statistics/ (accessed on 5 January 2023).
- A Guide to Hawaiian Coffee Production. Available online: https://perfectdailygrind.com/2022/01/a-guide-to-hawaiian-coffee-production/ (accessed on 5 January 2023).
- Grembecka, M.; Malinowska, E.; Szefer, P. Differentiation of market coffee and its infusions in view of their mineral composition. Sci. Total Environ. 2007, 383, 59–69. [Google Scholar] [CrossRef]
- Stelmach, E.; Pohl, P.; Szymczycha-Madeja, A. The suitability of the simplified method of the analysis of coffee infusions on the content of Ca, Cu, Fe, Mg, Mn and Zn and the study of the effect of preparation conditions on the leachability of elements into the coffee brew. Food Chem. 2013, 141, 1956–1961. [Google Scholar] [CrossRef]
- Stelmach, E.; Szymczycha-Madeja, A.; Pohl, P. A simplified determination of total concentrations of Ca, Fe, Mg and Mn in addition to their bioaccessible fraction in popular instant coffee brews. Food Chem. 2016, 197, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Wolska, J.; Janda, K.; Jakubczyk, K.; Szymkowiak, M.; Chlubek, D.; Gutowska, I. Levels of Antioxidant Activity and Fluoride Content in Coffee Infusions of Arabica, Robusta and Green Coffee Beans in According to their Brewing Methods. Biol. Trace Elem. Res. 2017, 179, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, N.; Kitts, D.D. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Girolamo, F.G.; Mazzucco, S.; Situlin, R.; Mohorko, N.; Jenko-Praznikar, Z.; Petelin, A.; Tence, M.; Pisot, R.; Navarini, L.; Biolo, G. Roasting intensity of naturally low-caffeine Laurina coffee modulates glucose metabolism and redox balance in humans. Nutrition 2016, 32, 928–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.; Kim, M.H.; Park, J.H.; Jeong, Y.; Ko, K.S. Cellular Antioxidant and Anti-Inflammatory Effects of Coffee Extracts with Different Roasting Levels. J. Med. Food 2017, 20, 626–635. [Google Scholar] [CrossRef]
- Ho, K.V.; Schreiber, K.L.; Park, J.; Vo, P.H.; Lei, Z.; Sumner, L.W.; Brown, C.R.; Lin, C.H. Identification and Quantification of Bioactive Molecules Inhibiting Pro-inflammatory Cytokine Production in Spent Coffee Grounds Using Metabolomics Analyses. Front. Pharmacol. 2020, 11, 229. [Google Scholar] [CrossRef]
- Angeloni, G.; Guerrini, L.; Masella, P.; Innocenti, M.; Bellumori, M.; Parenti, A. Characterization and comparison of cold brew and cold drip coffee extraction methods. J. Sci. Food Agric. 2019, 99, 391–399. [Google Scholar] [CrossRef]
- Rao, N.Z.; Fuller, M.; Grim, M.D. Physiochemical Characteristics of Hot and Cold Brew Coffee Chemistry: The Effects of Roast Level and Brewing Temperature on Compound Extraction. Foods 2020, 9, 902. [Google Scholar] [CrossRef]
- Cajka, T.; Fiehn, O. Toward Merging Untargeted and Targeted Methods in Mass Spectrometry-Based Metabolomics and Lipidomics. Anal. Chem. 2016, 88, 524–545. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.4.1–30.4.32. [Google Scholar] [CrossRef] [Green Version]
- Sud, M.; Fahy, E.; Cotter, D.; Azam, K.; Vadivelu, I.; Burant, C.; Edison, A.; Fiehn, O.; Higashi, R.; Nair, K.S.; et al. Metabolomics Workbench: An international repository for metabolomics data and metadata, metabolite standards, protocols, tutorials and training, and analysis tools. Nucleic Acids Res. 2016, 44, D463–D470. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.M.; Rosario, R.; Brito, A.; Agrawal, K.; Fanter, R.; Lietz, G.; Haskell, M.; Engle-Stone, R.; Newman, J.W.; La Frano, M.R. Multiassay nutritional metabolomics profiling of low vitamin A status versus adequacy is characterized by reduced plasma lipid mediators among lactating women in the Philippines: A pilot study. Nutr. Res. 2022, 104, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Granda, M.L.; Prince, D.K.; Fiehn, O.; Chen, Y.; Rajabi, T.; Yeung, C.K.; Hoofnagle, A.N.; Kestenbaum, B. Metabolomic Profiling Identifies New Endogenous Markers of Tubular Secretory Clearance. Kidney360 2023, 4, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Miao, G.; Deen, J.; Struzeski, J.B.; Chen, M.; Zhang, Y.; Cole, S.A.; Fretts, A.M.; Lee, E.T.; Howard, B.V.; Fiehn, O.; et al. Plasma lipidomic profile of depressive symptoms: A longitudinal study in a large sample of community-dwelling American Indians in the strong heart study. Mol. Psychiatry 2023, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mazi, T.A.; Borkowski, K.; Newman, J.W.; Fiehn, O.; Bowlus, C.L.; Sarkar, S.; Matsukuma, K.; Ali, M.R.; Kieffer, D.A.; Wan, Y.Y.; et al. Ethnicity-specific alterations of plasma and hepatic lipidomic profiles are related to high NAFLD rate and severity in Hispanic Americans, a pilot study. Free. Radic. Biol. Med. 2021, 172, 490–502. [Google Scholar] [CrossRef]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Kind, T.; Okazaki, Y.; Saito, K.; Fiehn, O. LipidBlast templates as flexible tools for creating new in-silico tandem mass spectral libraries. Anal. Chem. 2014, 86, 11024–11027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- ORAC Antioxidant Assay Kit. Available online: http://www.zen-bio.com/products/kits/orac-antioxidant_screening.php (accessed on 29 May 2018).
- Pertuzatti, P.B.; Barcia, M.T.; Rodrigues, D.; da Cruz, P.N.; Hermosín-Gutiérrez, I.; Smith, R.; Godoy, H.T. Antioxidant activity of hydrophilic and lipophilic extracts of Brazilian blueberries. Food Chem. 2014, 164, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant Activity/Capacity Measurement. 2. Hydrogen Atom Transfer (HAT)-Based, Mixed-Mode (Electron Transfer (ET)/HAT), and Lipid Peroxidation Assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef]
- Pilipczuk, T.; Kusznierewicz, B.; Zielinska, D.; Bartoszek, A. The influence of roasting and additional processing on the content of bioactive components in special purpose coffees. J. Food Sci. Technol. 2015, 52, 5736–5744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toci, A.T.; Farah, A. Volatile fingerprint of Brazilian defective coffee seeds: Corroboration of potential marker compounds and identification of new low quality indicators. Food Chem. 2014, 153, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.-J.; Jeong, H.-J.; Moon, H.; Kim, H.-N.; Cho, J.-H.; Lee, J.-E.; Hong, K.S.; Hong, Y.-S. Assessment of green coffee beanmetabolites dependent on coffee quality using a 1H NMR-based metabolomics approach. Food Res. Int. 2015, 67, 175–182. [Google Scholar] [CrossRef]
- Malta, M.R.; Fassio, L.O.; Liska, G.R.; Carvalho, G.R.; Pereira, A.A.; Botelho, C.E.; Ferraz, V.P.; Silva, A.D.; Pedrosa, A.W.; Alvaro, L.N.; et al. Discrimination of genotypes coffee by chemical composition of the beans: Potential markers in natural coffees. Food Res. Int. 2020, 134, 109219. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Braceschi, G.P.; Odello, L.; Bertuzzi, T.; Trevisan, M.; Lucini, L. Identification of markers of sensory quality in ground coffee: An untargeted metabolomics approach. Metabolomics 2020, 16, 127. [Google Scholar] [CrossRef] [PubMed]
- Happyana, N.; Pratiwi, A.; Hakim, E.H. Metabolite Profiles of the Green Beans of Indonesian Arabica Coffee Varieties. Int. J. Food Sci. 2021, 2021, 5782578. [Google Scholar] [CrossRef]
- Wongsa, P.; Khampa, N.; Horadee, S.; Chaiwarith, J.; Rattanapanone, N. Quality and bioactive compounds of blends of Arabica and Robusta spray-dried coffee. Food Chem. 2019, 283, 579–587. [Google Scholar] [CrossRef]
- Colzi, I.; Taiti, C.; Marone, E.; Magnelli, S.; Gonnelli, C.; Mancuso, S. Covering the different steps of the coffee processing: Can headspace VOC emissions be exploited to successfully distinguish between Arabica and Robusta? Food Chem. 2017, 237, 257–263. [Google Scholar] [CrossRef]
- Monakhova, Y.B.; Ruge, W.; Kuballa, T.; Ilse, M.; Winkelmann, O.; Diehl, B.; Thomas, F.; Lachenmeier, D.W. Rapid approach to identify the presence of Arabica and Robusta species in coffee using 1H NMR spectroscopy. Food Chem. 2015, 182, 178–184. [Google Scholar] [CrossRef]
- Pereira, T.S.; Batista, N.N.; Santos Pimenta, L.P.; Martinez, S.J.; Ribeiro, L.S.; Oliveira Naves, J.A.; Schwan, R.F. Self-induced anaerobiosis coffee fermentation: Impact on microbial communities, chemical composition and sensory quality of coffee. Food Microbiol. 2022, 103, 103962. [Google Scholar] [CrossRef]
- Król, K.; Gantner, M.; Tatarak, A.; Hallmann, E. The content of polyphenols in coffee beans as roasting, origin and storage effect. Eur. Food Res. Technol. 2020, 246, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Bruges, G.; Venturini, W.; Crespo, G.; Zambrano, M.L. Pyrogallol Induces Apoptosis in Human Platelets. Folia Biol. 2018, 64, 23–30. [Google Scholar]
- Kanai, N.; Yoshihara, N.; Kawamura, I. Solid-state NMR characterization of triacylglycerol and polysaccharides in coffee beans. Biosci. Biotechnol. Biochem. 2019, 83, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Anagbogu, C.F.; Zhou, J.; Olasupo, F.O.; Baba Nitsa, M.; Beckles, D.M. Lipidomic and metabolomic profiles of Coffea canephora L. beans cultivated in Southwestern Nigeria. PLoS ONE 2021, 16, e0234758. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.P.; Borem, F.M.; Ribeiro, F.C.; Giomo, G.S.; Henrique, d.S.J.; Malta, M.R. Fatty acid profiles and parameters of quality of specialty coffees produced in different Brazilian regions. Afr. J. Agric. Res. 2015, 10, 3484–3493. [Google Scholar] [CrossRef]
- Martin, M.J.; Pablos, F.; Gonzalez, A.G.; Valdenebro, M.S.; Leon-Camacho, M. Fatty acid profiles as discriminant parameters for coffee varieties differentiation. Talanta 2001, 54, 291–297. [Google Scholar] [CrossRef]
- Joet, T.; Laffargue, A.; Descroix, F.; Doulbeau, S.; Bertrand, B.; de Kochko, A.; Dussert, S. Influence of environmental factors, wet processing and their interactions on the biochemical composition of green Arabica coffee beans. Food Chem. 2010, 118, 693–701. [Google Scholar] [CrossRef]
- Mehari, B.; Redi-Abshiro, M.; Chandravanshi, B.S.; Combrinck, S.; McCrindle, R.; Atlabachew, M. GC-MS profiling of fatty acids in green coffee (Coffea arabica L.) beans and chemometric modeling for tracing geographical origins from Ethiopia. J. Sci. Food Agric. 2019, 99, 3811–3823. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.C.R.; da Silva, C.C.; Garrett, R.; Rezende, C.M. Comprehensive lipid analysis of green Arabica coffee beans by LC-HRMS/MS. Food Res. Int. 2020, 137, 109727–109736. [Google Scholar] [CrossRef]
- Cossignani, L.; Montesano, D.; Simonetti, M.S.; Blasi, F. Authentication of Coffea arabica according to Triacylglycerol Stereospecific Composition. J. Anal. Methods Chem. 2016, 2016, 7482620. [Google Scholar] [CrossRef] [Green Version]
- Romano, R.; Santini, A.; Le Grottaglie, L.; Manzo, N.; Visconti, A.; Ritien, i.A. Identification markers based on fatty acid composition to differentiate between roasted Arabica and Canephora (Robusta) coffee varieties in mixtures. J. Food Compos. Anal. 2014, 35, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.L.F.; Nascimento, G.O.; Lopes, G.S.; Matos, W.O.; Cunha, R.L.; Malta, M.R.; Liska, G.R.; Owen, R.W.; Trevisan, M.T.S. The concentration of polyphenolic compounds and trace elements in the Coffea arabica leaves: Potential chemometric pattern recognition of coffee leaf rust resistance. Food Res. Int. 2020, 134, 109221. [Google Scholar] [CrossRef] [PubMed]
- Visciano, P.; Schirone, M. Update on Biogenic Amines in Fermented and Non-Fermented Beverages. Foods 2022, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, M.; Lambropoulou, D.; Morrison, C.; Kłodzínska, E.; Namièsnik, J.; Płotka-Wasylka, J. Literature update of analytical methods for biogenic amines determination in food and beverages. Trends Anal. Chem. 2018, 98, 128–142. [Google Scholar] [CrossRef] [Green Version]
- Kalac, P. Recent advances in the research on biological roles of dietary polyamines in man. J. Appl. Biomed. 2009, 7, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, A.R. Significance of biogenic amines to food safety and human health. Food Res. Int. 1996, 29, 675–690. [Google Scholar] [CrossRef]
- Choi, M.-Y.; Choi, W.; Park, J.H.; Lim, J.; Kwon, S.W. Determination of coffee origins by integrated metabolomic approach of combining multiple analytical data. Food Chem. 2010, 121, 1260–1268. [Google Scholar] [CrossRef]
- Amorim, H.V.; Basso, L.C.; Crocomo, O.J.; Teixeira, A.A. Polyamines in green and roasted coffee. J. Agric. Food Chem. 1977, 25, 957–958. [Google Scholar] [CrossRef]
- Casal, S.; Mendes, E.; Oliveira, M.B.P.P.; Ferreira, M.A. Determination of biogenic amines in coffee by an optimized liquid chromatographic method. J. Liq. Chromatogr. Relat. Technol. 2002, 25, 2535–2549. [Google Scholar] [CrossRef]
- Casal, S.; Mendes, E.; Alves, M.R.; Alves, R.C.; Beatriz, M.; Oliveira, P.P.; Ferreira, M.A. Free and conjugated biogenic amines in green and roasted coffee beans. J. Agric. Food Chem. 2004, 52, 6188–6192. [Google Scholar] [CrossRef]
- Cirilo, M.P.G.; Coelho, A.F.S.; Araujo, C.M.; Goncalves, F.R.B.; Nogueira, F.D.; Gloria, M.B.A. Profiles and levels of bioactive amines in green and roasted coffee. Food Chem. 2003, 82, 397–402. [Google Scholar] [CrossRef]
- Dias, E.C.; Pereira, R.G.; Borem, F.M.; Mendes, E.; de Lima, R.R.; Fernandes, J.O.; Casal, S. Biogenic amine profile in unripe Arabica coffee beans processed according to dry and wet methods. J. Agric. Food Chem. 2012, 60, 4120–4125. [Google Scholar] [CrossRef] [PubMed]
- Priftis, A.; Stagos, D.; Konstantinopoulos, K.; Tsitsimpikou, C.; Spandidos, D.A.; Tsatsakis, A.M.; Tzatzarakis, M.N.; Kouretas, D. Comparison of antioxidant activity between green and roasted coffee beans using molecular methods. Mol. Med. Rep. 2015, 12, 7293–7302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opitz, S.E.; Goodman, B.A.; Keller, M.; Smrke, S.; Wellinger, M.; Schenker, S.; Yeretzian, C. Understanding the Effects of Roasting on Antioxidant Components of Coffee Brews by Coupling On-line ABTS Assay to High Performance Size Exclusion Chromatography. Phytochem. Anal. 2017, 28, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Perez-Martinez, M.; Caemmerer, B.; De Pena, M.P.; Cid, C.; Kroh, L.W. Influence of brewing method and acidity regulators on the antioxidant capacity of coffee brews. J. Agric. Food Chem. 2010, 58, 2958–2965. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Calderon, J.; Mejia-Diaz, D.; Martinez-Castano, M.; Bedoya-Ramirez, D.; Lopez-Rojas, N.; Gomez-Narvaez, F.; Medina-Pineda, Y.; Vega-Castro, O. Evaluation of antioxidant capacity in coffees marketed in Colombia: Relationship with the extent of non-enzymatic browning. Food Chem. 2016, 209, 162–170. [Google Scholar] [CrossRef]
- Trandafir, I.; Nour, V.; Ionica, M.E. Antioxidant capacity, phenolic acids and caffeine contents of some commercial coffees available on the Romanian market. Arch. Latinoam. Nutr. 2013, 63, 87–94. [Google Scholar]
- Parras, P.; Martínez-Tomè, M.; Jimènez, A.M.; Murcia, M.A. Antioxidant capacity of coffees of several origins brewed following three different procedures. Food Chem. 2007, 102, 582–592. [Google Scholar] [CrossRef]
- Nicoli, M.C.; Anese, M.; Manzocco, L.; Lerici, C.R. Antioxidant properties of coffee brews in relation to the roasting degree. Food Sci. Technol.-Lebensm. Wiss. Technol. 1997, 30, 292–297. [Google Scholar] [CrossRef]
- Lopez-Galilea, I.; de Peña, M.P.; Cid, C. Application of multivariate analysis to investigate potential antioxidants in conventional and torrefacto roasted coffee. Eur. Food Res. Technol. 2008, 227, 141–149. [Google Scholar] [CrossRef]
- Yashin, A.; Yashin, Y.; Wang, J.Y.; Nemzer, B. Antioxidant and Antiradical Activity of Coffee. Antioxidants 2013, 2, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Ramadan-Hassanien, M.F. Total antioxidant potential of juices, beverages and hot drinks consumed in Egypt screened by DPPH in vitro assay. Grasas Aceites 2008, 59, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Abdelwareth, A.; Zayed, A.; Farag, M.A. Chemometrics-based aroma profiling for revealing origin, roasting indices, and brewing method in coffee seeds and its commercial blends in the Middle East. Food Chem. 2021, 349, 129162. [Google Scholar] [CrossRef] [PubMed]

 = Kaua’i roasted (KR) and
= Kaua’i roasted (KR) and  = Waialua roasted (WR).
= Kaua’i roasted (KR) and = Waialua roasted (WR).
= Waialua roasted (WR).
= Kaua’i roasted (KR) and = Waialua roasted (WR).


 KG = Kaua’i green,
KG = Kaua’i green,  KR = Kaua’i roasted,
KR = Kaua’i roasted,  WG = Waialua green, and
WG = Waialua green, and  WR = Waialua roasted.
KG = Kaua’i green, KR = Kaua’i roasted, WG = Waialua green, and WR = Waialua roasted.
WR = Waialua roasted.
KG = Kaua’i green, KR = Kaua’i roasted, WG = Waialua green, and WR = Waialua roasted.

 cold brew;
cold brew;  filter mesh;
filter mesh;  filter paper;
filter paper;  French press.
cold brew; filter mesh; filter paper; French press.
French press.
cold brew; filter mesh; filter paper; French press.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mineral | Average RDA, AI, and UL in Males and Females in mg/day (19-70 years) | Minerals in 8 oz of Kaua’i Coffee (mg) | Minerals in 8 oz of Waialua Coffee (mg) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| French Press (K-FrP) | Filter Paper (K-FP) | Filter Mesh (K-FM) | Cold Brew (K-CB) | French Press (W-FrP) | Filter Paper (W-FP) | Filter Mesh (W-FM) | Cold Brew (W-CB) | ||
| P (mg.day−1) 1 | 700 * 1250 ** | 5.721 ± 0.26 a | 3.352 ± 0.42 a | 4.524 ± 1.46 a | 12.128 ± 1.30 b | 5.511 ± 0.004 a | 4.315 ± 0.13 a | 4.572 ± 0.45 a | 14.074 ± 1.36 b |
| K (mg.day−1) 2 | 2600 * 4700 ** | 110.395 ± 9.70 a,b | 79.83 ± 7.16 a | 108.60 ± 33.17 a,b | 137.26 ± 16.88 b | 112.26 ± 2.57 a,b | 101.65 ± 3.42 a,b | 112.74 ± 8.60 a,b | 171.61 ± 17.77 c |
| Ca (mg.day−1) 1 | 1000 * 1300 ** | 2.883 ± 0.19 a | 2.116 ± 0.23 a | 2.363 ± 0.80 a | 5.711 ± 0.56 b | 2.299 ± 0.10 a | 2.032 ± 0.18 a | 1.946 ± 0.12 a | 6.871 ± 0.36 c |
| Mg (mg.day−1) 1 | 310–320 * 400–420 * | 11.17 ± 1.28 a | 7.31 ± 0.14 b | 8.69 ± 2.26 a,b | 16.78 ± 0.97 c | 8.92 ± 0.18 a,b | 7.45 ± 0.42 b | 7.562± 0.65 b | 18.61 ± 1.73 c |
| Na (mg.day−1) 2 | 2300 * | 7.00 ± 0.008 a | 5.384 ± 0.20 a | 5.977 ± 1.63 a | 8.773 ± 0.23 b,c | 6.654 ± 0.05 a,b | 5.92 ± 0.13 a | 6.309 ± 0.86 a | 9.99 ± 1.26 c |
| Fe (mg.day−1) 1 | 18 * 8 * | 0.0084 ± 0.0011 a,d | 0.0073 ± 0.0007 d | 0.01 ± 0.002 a,d | 0.017 ± 0.001 b | 0.011 ± 0.0008 a | 0.011 ± 0.00001 a,d | 0.012 ± 0.001 a | 0.029 ± 0.003 c |
| Mn (mg.day−1) 2 | 2.3 ** | 0.056 ± 0.004 a | 0.036 ± 0.008 a | 0.05 ± 0.02 a | 0.156 ± 0.03 b | 0.057 ± 0.0004 a | 0.046 ± 0.003 a | 0.05 ± 0.003 a | 0.252 ± 0.009 c |
| Zn (mg.day−1) 1 | 8 * 11 * | 0.006 ± 0.0002 a | 0.004 ± 0.001 a | 0.006 ± 0.001 a | 0.014 ± 0.003 b | 0.006 ± 0.001 a | 0.004 ± 0.00002 a | 0.007 ± 0.003 a,b | 0.023 ± 0.006 c |
| Cu (μg.day−1) 1 | 0.9 ** | 0.005 ± 0.0001 a | 0.004 ± 0.0004 a | 0.006 ± 0.001 a | 0.009 ± 0.001 a | 0.005 ± 0.0001 a | 0.006 ± 0.001 a | 0.006 ± 0.001 a | 0.034 ± 0.023 b |
| B (mg.day−1) 3 | 1.5 | 0.042 ± 0.003 a,c | 0.035 ± 0.003 a,b | 0.048± 0.01 a.c | 0.025 ± 0.003 b | 0.044 ± 0.0008 a,c | 0.048 ± 0.003 a,c | 0.052± 0.003 c | 0.039 ± 0.004 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nerurkar, P.V.; Yokoyama, J.; Ichimura, K.; Kutscher, S.; Wong, J.; Bittenbender, H.C.; Deng, Y. Medium Roasting and Brewing Methods Differentially Modulate Global Metabolites, Lipids, Biogenic Amines, Minerals, and Antioxidant Capacity of Hawai‘i-Grown Coffee (Coffea arabica). Metabolites 2023, 13, 412. https://doi.org/10.3390/metabo13030412
Nerurkar PV, Yokoyama J, Ichimura K, Kutscher S, Wong J, Bittenbender HC, Deng Y. Medium Roasting and Brewing Methods Differentially Modulate Global Metabolites, Lipids, Biogenic Amines, Minerals, and Antioxidant Capacity of Hawai‘i-Grown Coffee (Coffea arabica). Metabolites. 2023; 13(3):412. https://doi.org/10.3390/metabo13030412
Chicago/Turabian StyleNerurkar, Pratibha V., Jennifer Yokoyama, Kramer Ichimura, Shannon Kutscher, Jamie Wong, Harry C. Bittenbender, and Youping Deng. 2023. "Medium Roasting and Brewing Methods Differentially Modulate Global Metabolites, Lipids, Biogenic Amines, Minerals, and Antioxidant Capacity of Hawai‘i-Grown Coffee (Coffea arabica)" Metabolites 13, no. 3: 412. https://doi.org/10.3390/metabo13030412