
Nanosuspension of Foeniculum Vulgare Promotes Accelerated Sensory and Motor Function Recovery after Sciatic Nerve Injury

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Collection and Processing of Foeniculum vulgare
2.3. Preparation of Foeniculum vulgare Nanosuspensions
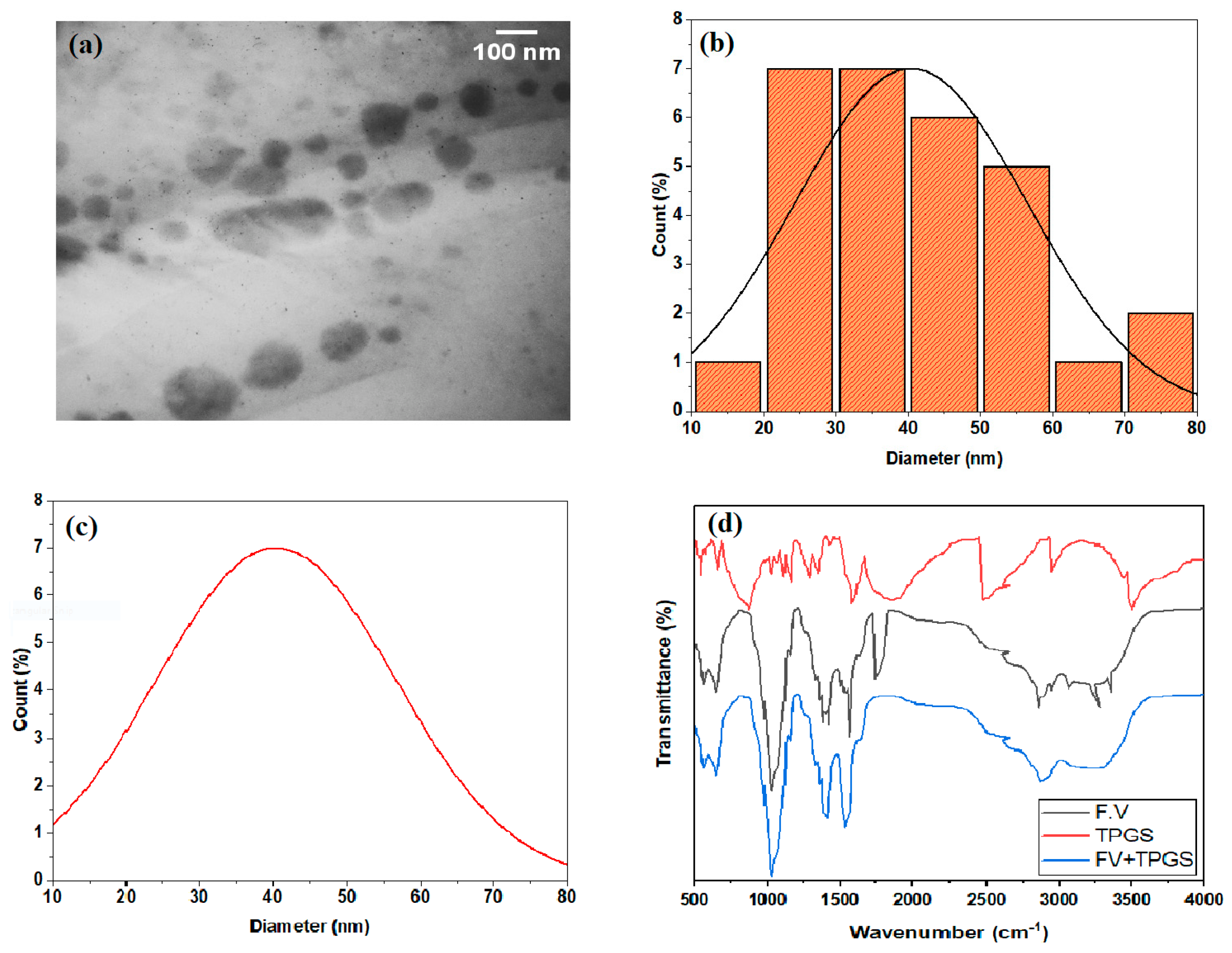
2.4. Characterization of Nanosuspension
2.5. Supplementation of FV Nanosuspension
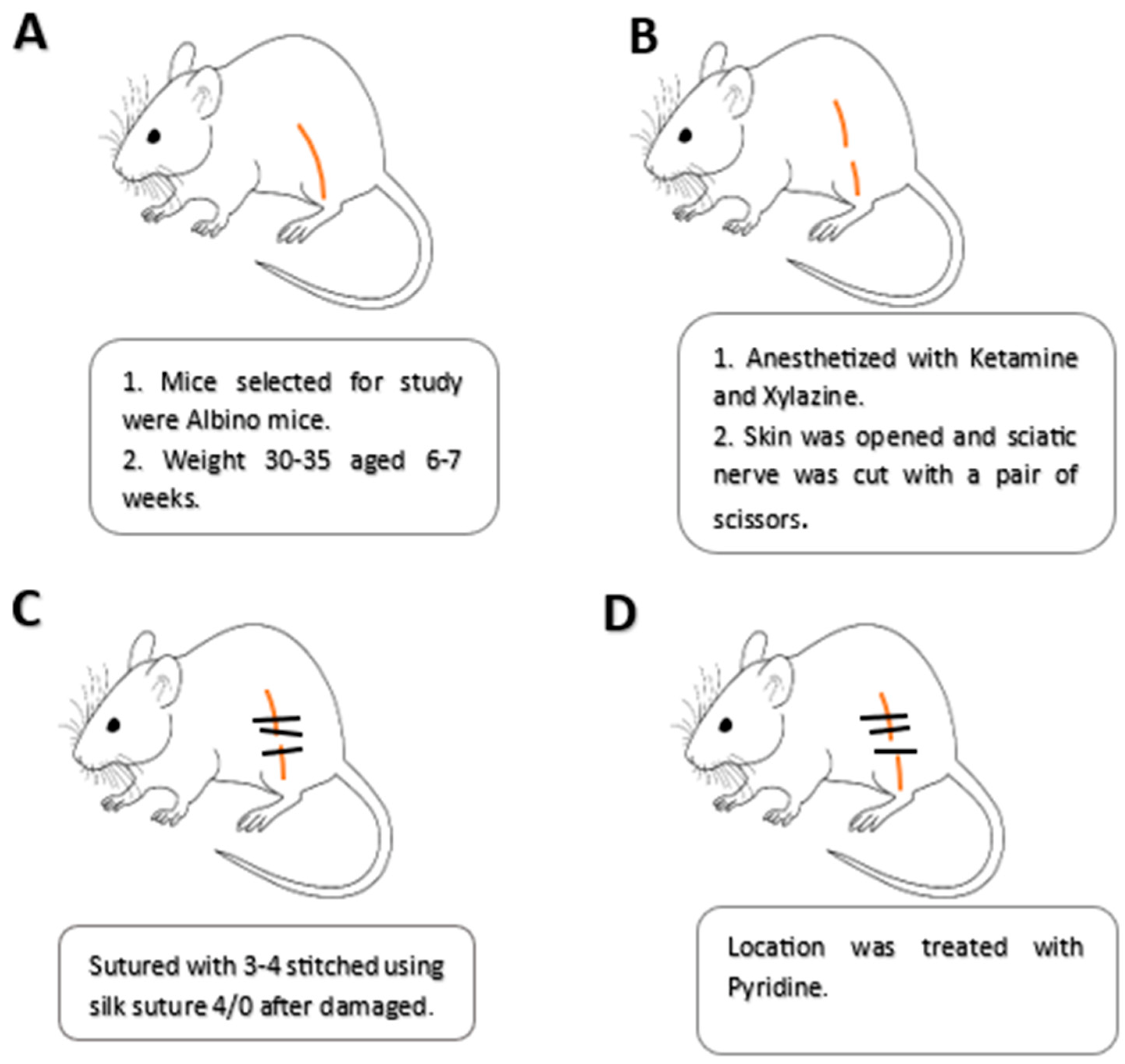
2.6. Induction of Sciatic Nerve Crush Injury
2.7. Behavior-Related Studies
2.7.1. Sciatic Functional Index
2.7.2. Hot Plate Test
2.7.3. Assessment of Grip Strength of Muscle
2.8. Biochemical Studies
2.8.1. Total Antioxidant Capacity
2.8.2. Total Oxidant Status (TOS)
2.8.3. Random Blood Glucose
3. Results
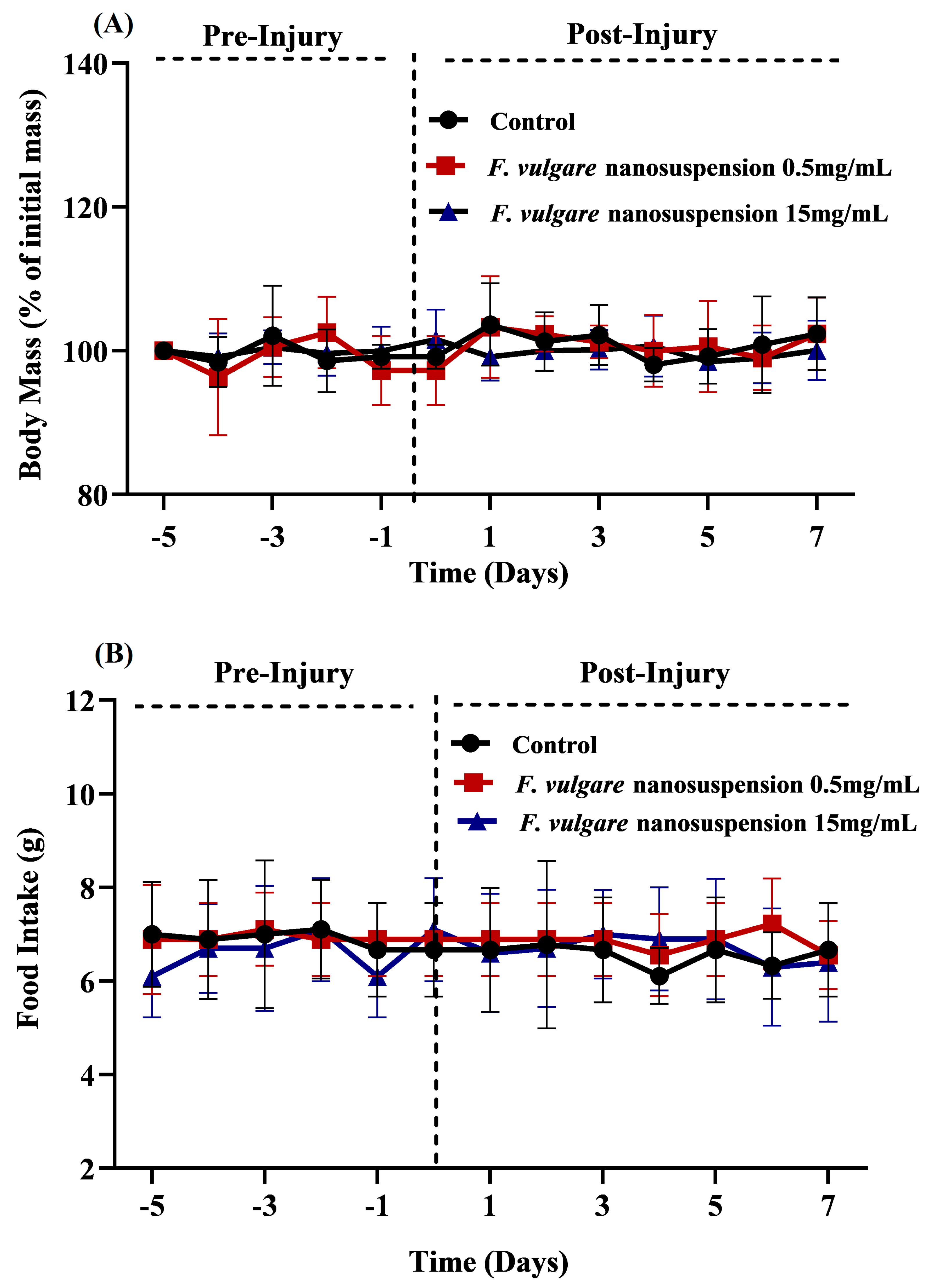
3.1. Effects of NS-Treatments on Body Mass and Food Consumption
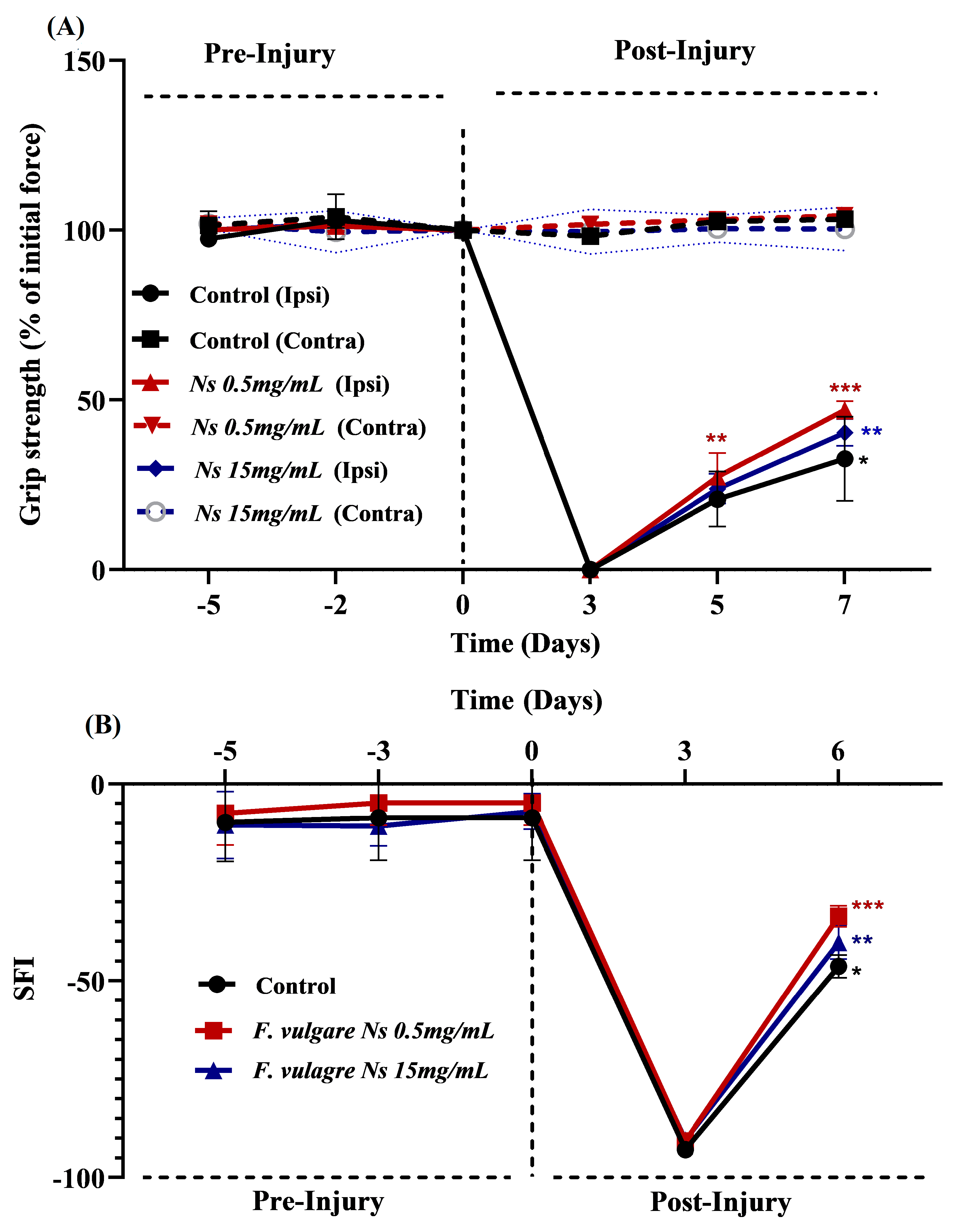
3.2. Effects of NS-Treatments on Regaining of Motor Functions and Muscle Strength
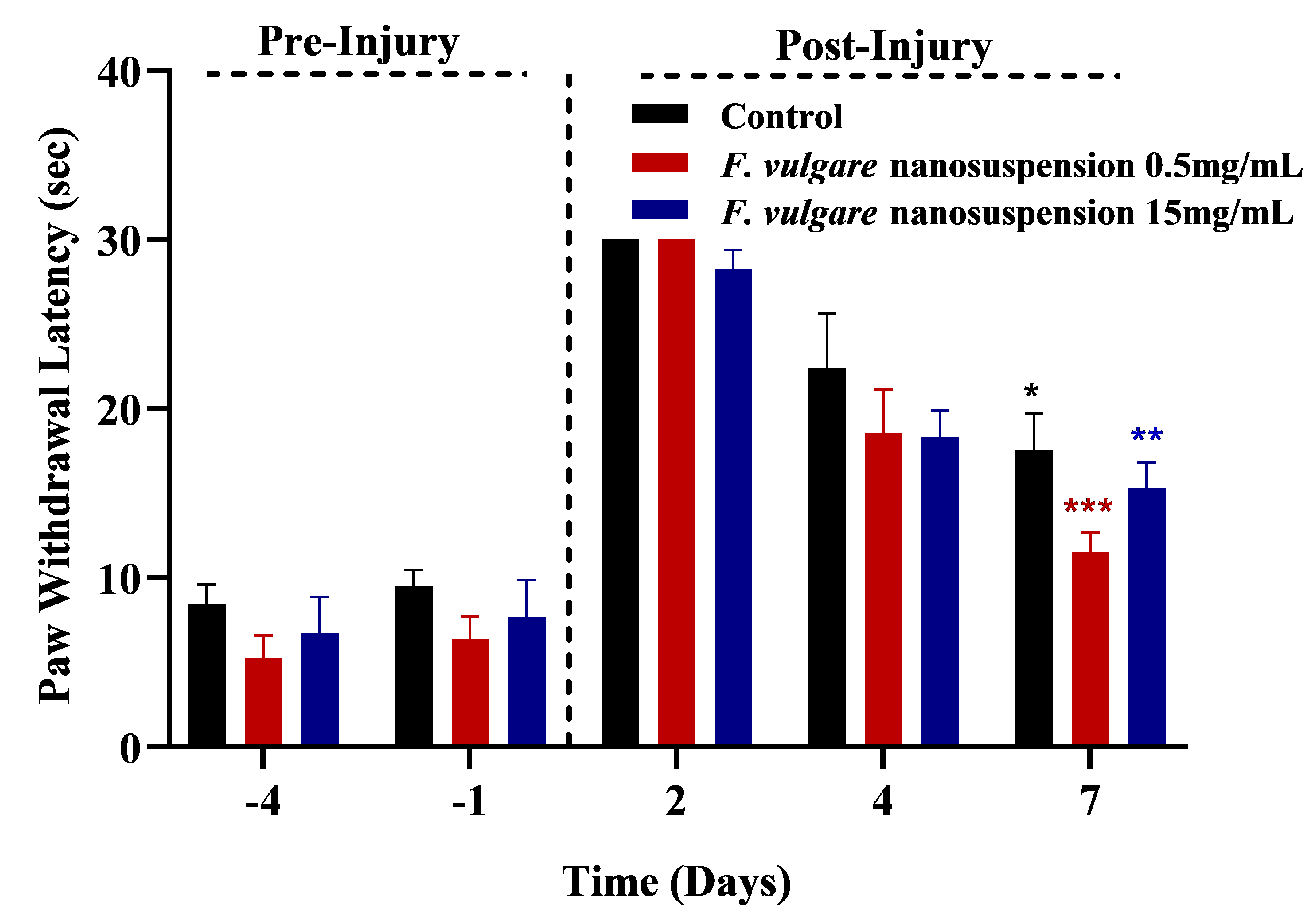
3.3. Effects of NS-Treatments on Regaining of Sensory Functions
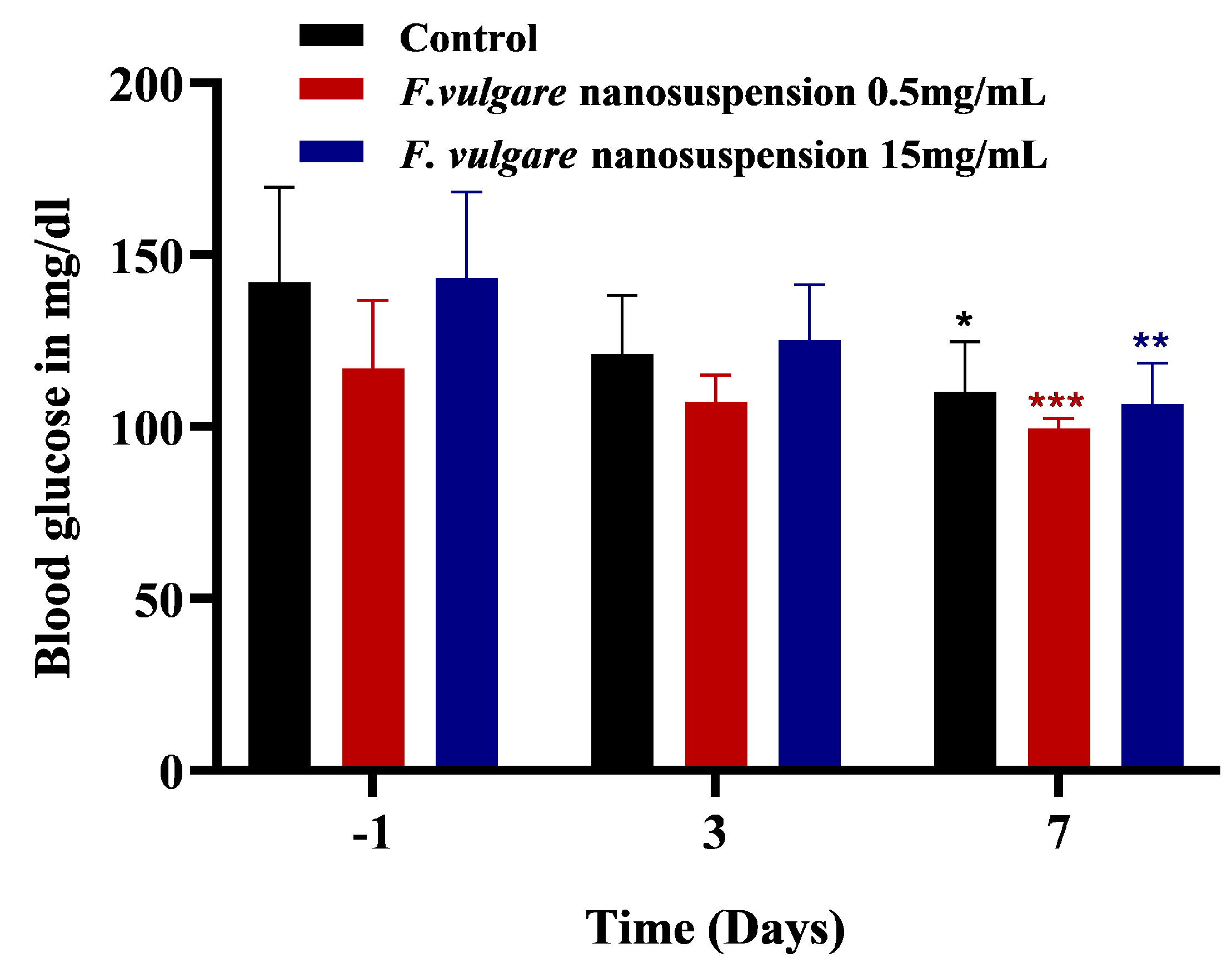
3.4. Effects of NS-Treatments on Glycemic Level
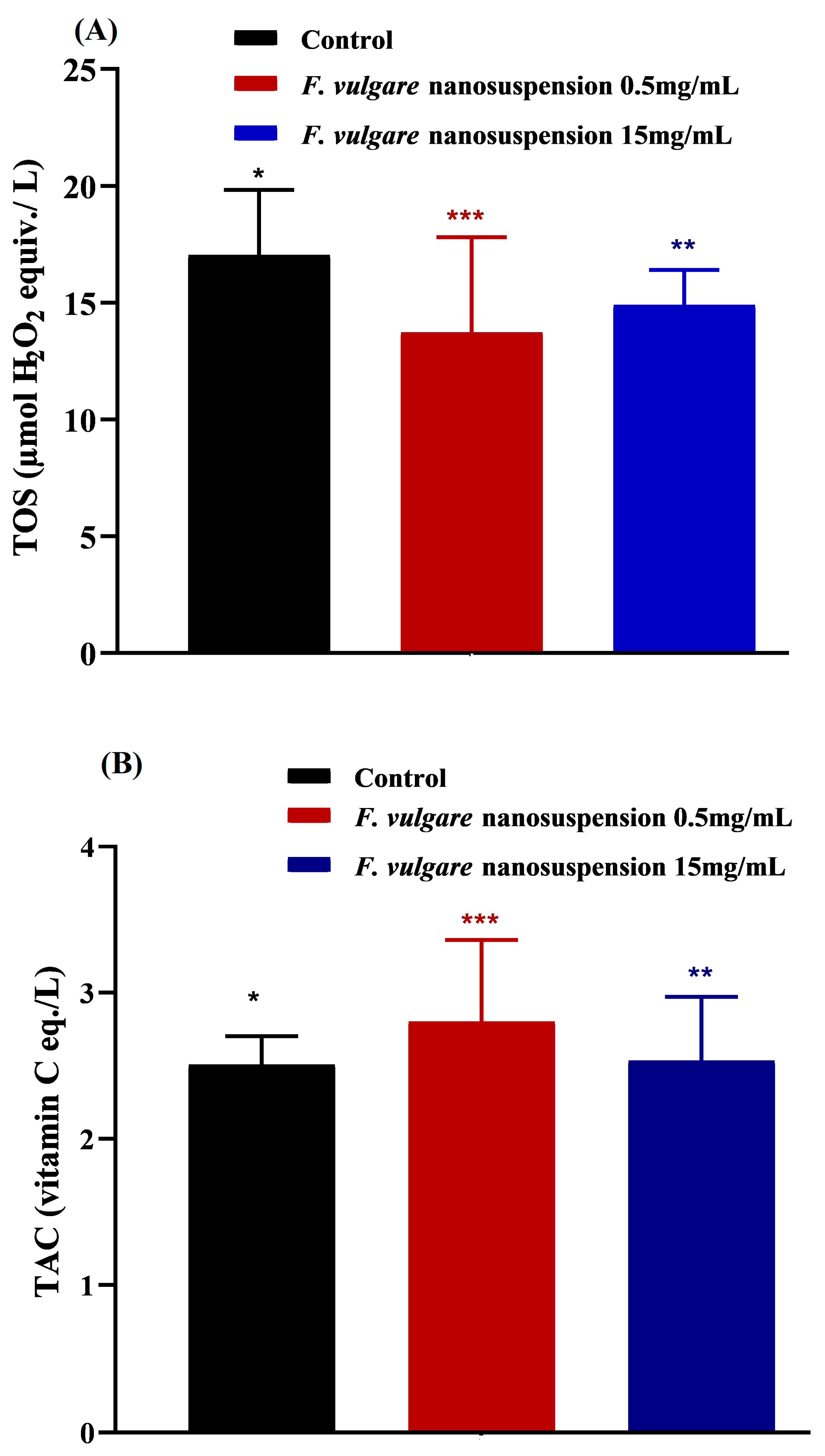
3.5. Effects of NS-Treatments on Oxidative Stress
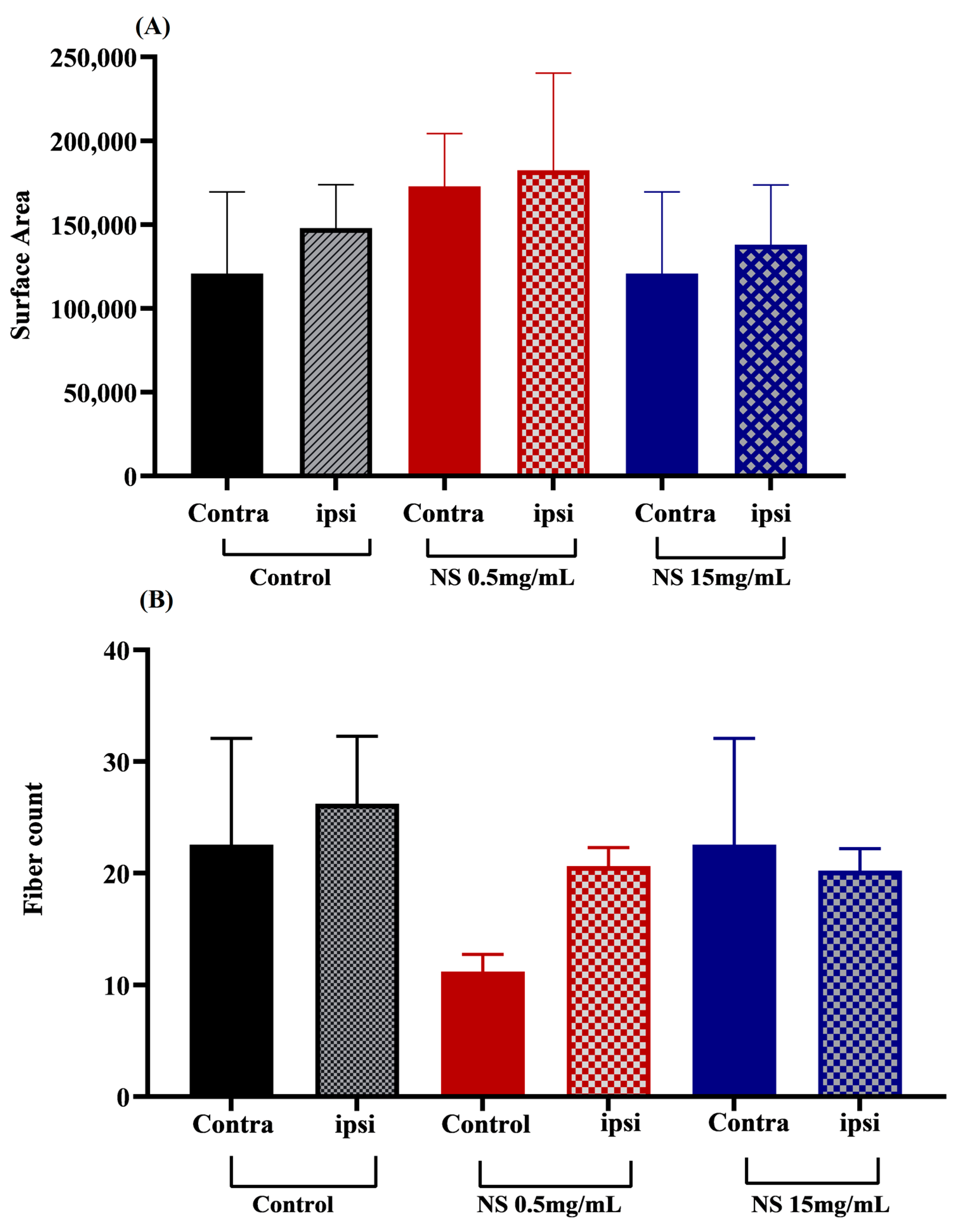
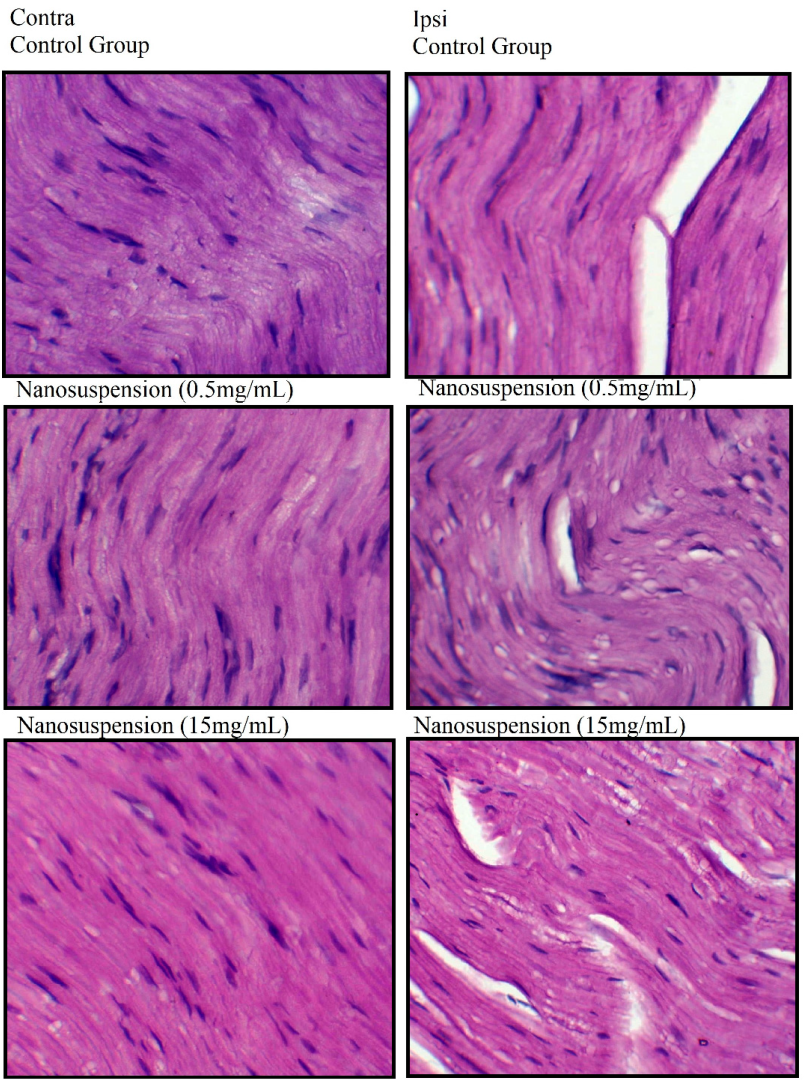
3.6. Effects of Nanosuspension on Muscle Histology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Namgung, U. The role of Schwann cell-axon interaction in peripheral nerve regeneration. Cells Tissues Organs 2014, 200, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R. The role of c-Jun and autocrine signaling loops in the control of repair Schwann cells and regeneration. Front. Cell. Neurosci. 2022, 15, 581. [Google Scholar] [CrossRef]
- Rodemer, W.; Gallo, G.; Selzer, M.E. Mechanisms of axon elongation following CNS injury: What is happening at the axon tip? Front. Cell. Neurosci. 2020, 14, 177. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xiao, H.; Zhao, J.; Ju, L.; Wen, Y.; Xu, Q.; Cui, X. GAP-43 is involved in the orientation of cell division by interacting with GAI during neurogenesis. Int. J. Neurosci. 2020, 130, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, D.-h.; Zhang, H.-y.; Wang, J.; Li, X.-k.; Xiao, J. Growth factors-based therapeutic strategies and their underlying signaling mechanisms for peripheral nerve regeneration. Acta Pharmacol. Sin. 2020, 41, 1289–1300. [Google Scholar] [CrossRef]
- Gordon, T. Peripheral nerve regeneration and muscle reinnervation. Int. J. Mol. Sci. 2020, 21, 8652. [Google Scholar] [CrossRef]
- Bolívar, S.; Navarro, X.; Udina, E. Schwann cell role in selectivity of nerve regeneration. Cells 2020, 9, 2131. [Google Scholar] [CrossRef]
- Yow, Y.-Y.; Goh, T.-K.; Nyiew, K.-Y.; Lim, L.-W.; Phang, S.-M.; Lim, S.-H.; Ratnayeke, S.; Wong, K.-H. Therapeutic Potential of Complementary and Alternative Medicines in Peripheral Nerve Regeneration: A Systematic Review. Cells 2021, 10, 2194. [Google Scholar] [CrossRef]
- Lopes, B.; Sousa, P.; Alvites, R.; Branquinho, M.; Sousa, A.C.; Mendonça, C.; Atayde, L.M.; Luís, A.L.; Varejão, A.S.P.; Maurício, A.C. Peripheral Nerve Injury Treatments and Advances: One Health Perspective. Int. J. Mol. Sci. 2022, 23, 918. [Google Scholar] [CrossRef]
- Messaoudi, M.; Begaa, S. Application of INAA technique for analysis of essential trace and toxic elements in medicinal seeds of Carum carvi L. & Foeniculum vul-gare Mill. used in Algeria. J. Appl. Res. Med. Aromat. Plants 2018, 9, 39–45. [Google Scholar] [CrossRef]
- Messaoudi, M.; Begaa, S. Dietary Intake and Content of Some Micronutrients and Toxic Elements in Two Algerian Spices (Coriandrum sativum L. and Cuminum cyminum L.). Biol. Trace Elem. Res. 2019, 188, 508–513. [Google Scholar] [CrossRef]
- Messaoudi, M.; Rebiai, A.; Sawicka, B.; Atanassova, M.; Ouakouak, H.; Larkem, I.; Egbuna, C.; Awuchi, C.G.; Boubekeur, S.; Ferhat, M.A.; et al. Effect of Extraction Methods on Polyphenols, Flavonoids, Mineral Elements, and Biological Activities of Essential Oil and Extracts of Mentha pulegium L. Molecules 2022, 27, 11. [Google Scholar] [CrossRef]
- Messaoudi, M.; Benarfa, A.; Ouakouak, H.; Begaa, S. Determination of Some Chemical Elements of Common Spices Used by Algerians and Possible Health Risk Assessment. Biol. Trace Elem. Res. 2022, 200, 2498–2509. [Google Scholar] [CrossRef]
- Parvez, M.K. Natural or plant products for the treatment of neurological disorders: Current knowledge. Curr. Drug Metab. 2018, 19, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Rane, A.R.; Paithankar, H.; Hosur, R.V.; Choudhary, S. Modulation of α-synuclein fibrillation by plant metabolites, daidzein, fisetin and scopoletin under physiological conditions. Int. J. Biol. Macromol. 2021, 182, 1278–1291. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Jeevanandam, J.; Egbuna, C.; Kumar, S.; Ifemeje, J.C. Phytochemicals as sources of drugs. In Phytochemistry: An In-Silico and In-Vitro Update; Springer: Berlin/Heidelberg, Germany, 2019; pp. 3–22. [Google Scholar]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.S.; Hussain, T.; Pati, S. Drug-lead anti-tuberculosis phytochemicals: A systematic review. Curr. Top. Med. Chem. 2021, 21, 1832–1868. [Google Scholar] [CrossRef]
- Khan, A.; Jahan, S.; Imtiyaz, Z.; Alshahrani, S.; Antar Makeen, H.; Mohammed Alshehri, B.; Kumar, A.; Arafah, A.; Rehman, M.U. Neuroprotection: Targeting multiple pathways by naturally occurring phytochemicals. Biomedicines 2020, 8, 284. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Lankatillake, C.; Dias, D.A.; Docea, A.O.; Mahomoodally, M.F.; Lobine, D.; Chazot, P.L.; Kurt, B.; Boyunegmez Tumer, T.; Catarina Moreira, A. Impact of natural compounds on neurodegenerative disorders: From preclinical to pharmacotherapeutics. J. Clin. Med. 2020, 9, 1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koulakiotis, N.S.; Purhonen, P.; Gikas, E.; Hebert, H.; Tsarbopoulos, A. Crocus-derived compounds alter the aggregation pathway of Alzheimer’s Disease-associated beta amyloid protein. Sci. Rep. 2020, 10, 18150. [Google Scholar] [CrossRef] [PubMed]
- Mehra, N.; Tamta, G.; Nand, V. A review on nutritional value, phytochemical and pharmacological attributes of Foeniculum vulgare Mill. J. Pharmacogn. Phytochem. 2021, 10, 1255–1263. [Google Scholar] [CrossRef]
- Kishore, N.; Verma, A.K. Foeniculum vulgare Mill: Flavoring, Pharmacological, Phytochemical, and Folklore Aspects. In Medicinal Plants; Apple Academic Press: Palm Bay, FL, USA, 2022; pp. 77–91. [Google Scholar]
- Shahsavari, M.; Mohammadabadi, M.; Khezri, A.; Borshch, O.; Babenko, O.; Kalashnyk, O.; Afanasenko, V.; Kondratiuk, V. Effect of Fennel (Foeniculum vulgare) Seed Powder Consumption on Insulin-like Growth Factor 1 Gene Expression in the Liver Tissue of Growing Lambs. Gene Expr. 2022, 21, 21–26. [Google Scholar] [CrossRef]
- Pohl, F.; Kong Thoo Lin, P. The potential use of plant natural products and plant extracts with antioxidant properties for the prevention/treatment of neurodegenerative diseases: In vitro, in vivo and clinical trials. Molecules 2018, 23, 3283. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.H.; Akter, R.; Kamal, M.A. Prospective function of different antioxidant containing natural products in the treatment of neurodegenerative diseases. CNS Neurol. Disord.-Drug Targets (Former. Curr. Drug Targets-CNS Neurol. Disord.) 2021, 20, 694–703. [Google Scholar]
- Maqbool, J.; Anwar, H.; Iqbal, J.; Rasul, A.; Imran, A.; Ahmad Malik, S.; Shabbir, A.; Ijaz, F.; Sajid, F.; Akram, R. Methanolic extract of Fennel (Foeniculum vulgare) escalates functional restoration following a compression injury to the sciatic nerve in a mouse model. Food Sci. Nutr. 2021, 9, 701–710. [Google Scholar] [CrossRef]
- Imran, A.; Xiao, L.; Ahmad, W.; Anwar, H.; Rasul, A.; Imran, M.; Aziz, N.; Razzaq, A.; Arshad, M.U.; Shabbir, A. Foeniculum vulgare (Fennel) promotes functional recovery and ameliorates oxidative stress following a lesion to the sciatic nerve in mouse model. J. Food Biochem. 2019, 43, e12983. [Google Scholar] [CrossRef]
- Čvorović, J.; Ziberna, L.; Fornasaro, S.; Tramer, F.; Passamonti, S. Bioavailability of flavonoids: The role of cell membrane transporters. In Polyphenols: Mechanisms of Action in Human Health and Disease; Elsevier: Amsterdam, The Netherlands, 2018; pp. 295–320. [Google Scholar]
- Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the encapsulation in bioavailability of phenolic compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.K.; Goyal, A.; Algın Yapar, E.; Cavalu, S. Bioactive compounds and nanodelivery perspectives for treatment of cardiovascular diseases. Appl. Sci. 2021, 11, 11031. [Google Scholar] [CrossRef]
- Vieira, I.R.S.; Conte-Junior, C.A. Nano-delivery systems for food bioactive compounds in cancer: Prevention, therapy, and clinical applications. Crit. Rev. Food Sci. Nutr. 2022, 1–26. [Google Scholar] [CrossRef]
- Wickramasinghe, A.S.D.; Kalansuriya, P.; Attanayake, A.P. Nanoformulation of plant-based natural products for type 2 diabetes mellitus: From formulation design to therapeutic applications. Curr. Ther. Res. 2022, 96, 100672. [Google Scholar] [CrossRef]
- Mohamad, R.H.; El-Bastawesy, A.M.; Abdel-Monem, M.G.; Noor, A.M.; Al-Mehdar, H.A.R.; Sharawy, S.M.; El-Merzabani, M.M. Antioxidant and anticarcinogenic effects of methanolic extract and volatile oil of fennel seeds (Foeniculum vulgare). J. Med. Food 2011, 14, 986–1001. [Google Scholar] [CrossRef]
- Parejo, I.; Viladomat, F.; Bastida, J.; Schmeda-Hirschmann, G.; Burillo, J.; Codina, C. Bioguided isolation and identification of the nonvolatile antioxidant compounds from fennel (Foeniculum vulgare Mill.) waste. J. Agric. Food Chem. 2004, 52, 1890–1897. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Zhao, J.; Ding, Y.; Li, L. A cost-effective method to prepare curcumin nanosuspensions with enhanced oral bioavailability. J. Colloid Interface Sci. 2017, 485, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Dar, B.A.; Sofi, S.N.; Bhat, B.A.; Qurishi, M.A. Foeniculum vulgare: A comprehensive review of its traditional use, phytochemistry, pharmacology, and safety. Arab. J. Chem. 2016, 9, S1574–S1583. [Google Scholar] [CrossRef] [Green Version]
- Nassar, M.I.; Aboutabl, E.-s.A.; Makled, Y.A.; El-Khrisy, E.-D.; Osman, A.F. Secondary metabolites and pharmacology of Foeniculum vulgare Mill. Subsp. Piperitum. Rev. Latinoam. De Química 2010, 38, 103–112. [Google Scholar]
- Sunila, E.S.; Kuttan, G. A Preliminary Study on Antimetastatic Activity of Thuja occidentalis L. in Mice Model. Immunopharmacol. Immunotoxicol. 2006, 28, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.-l.; Xiong, X.-b.; Xiang, M.-q. Effect of Arborvitae Seed on Cognitive Function and α7nAChR Protein Expression of Hippocampus in Model Rats with Alzheimer’s Disease. Cell Biochem. Biophys. 2013, 67, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Schmitt, F.; Henriques, A.; Lequeu, T.; Rene, F.; Bindler, F.; Dirrig-Grosch, S.; Oudart, H.; Palamiuc, L.; Metz-Boutigue, M.-H.; et al. Systemic Down-Regulation of Delta-9 Desaturase Promotes Muscle Oxidative Metabolism and Accelerates Muscle Function Recovery following Nerve Injury. PLoS ONE 2013, 8, e64525. [Google Scholar] [CrossRef] [Green Version]
- Naz, S.; Farooq, T.; Hameed, A.; Anwar, H.; Aslam, R.; Malik, S.A.; Akram, R.; Sajid, F.; Saeed, Z.; Hussain, G. Thuja Occidentalis l. Expedites Functional Recovery after Sciatic Nerve Crush Injury in Mice. Pak. J. Med. Health Sci. 2022, 16, 396. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar]
- Rubio, C.P.; Hernández-Ruiz, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Ceron, J.J. Spectrophotometric assays for total antioxidant capacity (TAC) in dog serum: An update. BMC Vet. Res. 2016, 12, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Feng, J.; Yang, Y.; Dai, C.; Lu, A.; Li, J.; Liao, Y.; Xiang, M.; Huang, Q.; Wang, D. Significance of serum total oxidant/antioxidant status in patients with colorectal cancer. PLoS ONE 2017, 12, e0170003. [Google Scholar] [CrossRef]
- Wang, M.L.; Rivlin, M.; Graham, J.G.; Beredjiklian, P.K. Peripheral nerve injury, scarring, and recovery. Connect. Tissue Res. 2019, 60, 3–9. [Google Scholar] [CrossRef]
- Fideles, S.O.M.; de Cássia Ortiz, A.; Buchaim, D.V.; de Souza Bastos Mazuqueli Pereira, E.; Parreira, M.J.B.M.; de Oliveira Rossi, J.; da Cunha, M.R.; de Souza, A.T.; Soares, W.C.; Buchaim, R.L. Influence of the Neuroprotective Properties of Quercetin on Regeneration and Functional Recovery of the Nervous System. Antioxidants 2023, 12, 149. [Google Scholar] [CrossRef]
- Muratori, L.; Fregnan, F.; Maurina, M.; Haastert-Talini, K.; Ronchi, G. The Potential Benefits of Dietary Polyphenols for Peripheral Nerve Regeneration. Int. J. Mol. Sci. 2022, 23, 5177. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Xiao, J.; Weng, Z.; Lu, X.; Shen, X.; Wang, F. A phenolic glycoside from Moringa oleifera Lam. improves the carbohydrate and lipid metabolisms through AMPK in db/db mice. Food Chem. 2020, 311, 125948. [Google Scholar] [CrossRef]
- Yang, Y.; Lin, L.; Zhao, M.; Yang, X. The hypoglycemic and hypolipemic potentials of Moringa oleifera leaf polysaccharide and polysaccharide-flavonoid complex. Int. J. Biol. Macromol. 2022, 210, 518–529. [Google Scholar] [CrossRef]
- Shah, K.H.; Oza, M.J. Comprehensive review of bioactive and molecular aspects of Moringa Oleifera lam. Food Rev. Int. 2022, 38, 1427–1460. [Google Scholar] [CrossRef]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; He, Z.; Ma, N.; Chen, Z.-Y. Beneficial effects of dietary polyphenols on high-fat diet-induced obesity linking with modulation of gut microbiota. J. Agric. Food Chem. 2019, 68, 33–47. [Google Scholar] [CrossRef]
- Safaei-Cherehh, A.; Rasouli, B.; Alaba, P.A.; Seidavi, A.; Hernández, S.R.; Salem, A.Z. Effect of dietary Foeniculum vulgare Mill. extract on growth performance, blood metabolites, immunity and ileal microflora in male broilers. Agrofor. Syst. 2020, 94, 1269–1278. [Google Scholar] [CrossRef]
- Mesfin, M.; Asres, K.; Shibeshi, W. Evaluation of anxiolytic activity of the essential oil of the aerial part of Foeniculum vulgare Miller in mice. BMC Complement. Altern. Med. 2014, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bawazir, A.; Bokhary, L.E. Study the Effect of Foeniculum vulgare Mill. Seeds Essential Oil on Some Neurotransmitters Content and Histological Structure of Cerebellar Cortex in Male Albino Rats. Adv. Environ. Biol. 2016, 10, 140–147. [Google Scholar]
- Sin, T.K.; Yung, B.Y.; Siu, P.M. Modulation of SIRT1-Foxo1 signaling axis by resveratrol: Implications in skeletal muscle aging and insulin resistance. Cell. Physiol. Biochem. 2015, 35, 541–552. [Google Scholar] [CrossRef]
- Salucci, S.; Falcieri, E. Polyphenols and their potential role in preventing skeletal muscle atrophy. Nutr. Res. 2020, 74, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Renno, W.M.; Al-Maghrebi, M.; Al-Banaw, A. (−)-Epigallocatechin-3-gallate (EGCG) attenuates functional deficits and morphological alterations by diminishing apoptotic gene overexpression in skeletal muscles after sciatic nerve crush injury. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 807–822. [Google Scholar] [CrossRef]
- Chen, M.-M.; Qin, J.; Chen, S.-J.; Yao, L.-M.; Zhang, L.-Y.; Yin, Z.-Q.; Liao, H. Quercetin promotes motor and sensory function recovery following sciatic nerve-crush injury in C57BL/6J mice. J. Nutr. Biochem. 2017, 46, 57–67. [Google Scholar] [CrossRef]
- Türedi, S.; Yuluğ, E.; Alver, A.; Bodur, A.; İnce, İ. A morphological and biochemical evaluation of the effects of quercetin on experimental sciatic nerve damage in rats. Exp. Ther. Med. 2018, 15, 3215–3224. [Google Scholar]
- Qiu, J.; Yang, X.; Wang, L.; Zhang, Q.; Ma, W.; Huang, Z.; Bao, Y.; Zhong, L.; Sun, H.; Ding, F. Isoquercitrin promotes peripheral nerve regeneration through inhibiting oxidative stress following sciatic crush injury in mice. Ann. Transl. Med. 2019, 7, 680. [Google Scholar] [CrossRef] [PubMed]
- Meotti, F.C.; Lemos de Andrade, E.; Calixto, J.B. TRP modulation by natural compounds. In Mammalian Transient Receptor Potential (TRP) Cation Channels: Volume II; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1177–1238. [Google Scholar]
- Bagood, M.D.; Isseroff, R.R. TRPV1: Role in skin and skin diseases and potential target for improving wound healing. Int. J. Mol. Sci. 2021, 22, 6135. [Google Scholar] [CrossRef] [PubMed]
- Coyoy-Salgado, A.; Segura-Uribe, J.J.; Guerra-Araiza, C.; Orozco-Suárez, S.; Salgado-Ceballos, H.; Feria-Romero, I.A.; Gallardo, J.M.; Orozco-Barrios, C.E. The Importance of Natural Antioxidants in the Treatment of Spinal Cord Injury in Animal Models: An Overview. Oxidative Med. Cell. Longev. 2019, 2019, 3642491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, W.; Wang, M.; Lin, C.; Li, G.; Zhou, X.; Luo, J.; Jin, D. Quercetin reduces neural tissue damage and promotes astrocyte activation after spinal cord injury in rats. J. Cell. Biochem. 2018, 119, 2298–2306. [Google Scholar] [CrossRef]
- Kouhestani, S.; Jafari, A.; Babaei, P. Kaempferol attenuates cognitive deficit via regulating oxidative stress and neuroinflammation in an ovariectomized rat model of sporadic dementia. Neural Regen. Res. 2018, 13, 1827. [Google Scholar]
- Kluska, M.; Juszczak, M.; Żuchowski, J.; Stochmal, A.; Woźniak, K. Effect of Kaempferol and Its Glycoside Derivatives on Antioxidant Status of HL-60 Cells Treated with Etoposide. Molecules 2022, 27, 333. [Google Scholar] [CrossRef] [PubMed]
- Gerhke, S.A.; Shibli, J.A.; Salles, M.B. Potential of the use of an antioxidant compound to promote peripheral nerve regeneration after injury. Neural Regenration Res. 2015, 10, 1063–1064. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Moradi, S.Z.; Cao, H.; Khan, H.; Xiao, J. Effects of polyphenols on oxidative stress, inflammation, and interconnected pathways during spinal cord injury. Oxidative Med. Cell. Longev. 2022, 2022, 8100195. [Google Scholar] [CrossRef]
- Bekiari, C.; Tekos, F.; Skaperda, Z.; Argyropoulou, A.; Skaltsounis, A.-L.; Kouretas, D.; Tsingotjidou, A. Antioxidant and Neuroprotective Effect of a Grape Pomace Extract on Oxaliplatin-Induced Peripheral Neuropathy in Rats: Biochemical, Behavioral and Histopathological Evaluation. Antioxidants 2022, 11, 1062. [Google Scholar] [CrossRef]
- Moradi, S.Z.; Jalili, F.; Farhadian, N.; Joshi, T.; Wang, M.; Zou, L.; Cao, H.; Farzaei, M.H.; Xiao, J. Polyphenols and neurodegenerative diseases: Focus on neuronal regeneration. Crit. Rev. Food Sci. Nutr. 2022, 62, 3421–3436. [Google Scholar] [CrossRef]
- El-Ouady, F.; Lahrach, N.; Ajebli, M.; Haidani, A.E.; Eddouks, M. Antihyperglycemic effect of the aqueous extract of Foeniculum vulgare in normal and streptozotocin-induced diabetic rats. Cardiovasc. Haematol. Disord.-Drug Targets (Former. Curr. Drug Targets-Cardiovasc. Hematol. Disord.) 2020, 20, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Zulifqar, S. Influence of Foeniculum vulgare Mill in the Management of Hyperglycemia. Int. J. Innov. Sci. Res. Technol. 2019, 4, 1117–1122. [Google Scholar]
- Parsaeyan, N. The effect of Foeniculum vulgare (fennel) extract on lipid profile, lipid peroxidation and liver enzymes of diabetic rat. Iran. J. Diabetes Obes. 2016, 8, 24–29. [Google Scholar]
- Yan, L.; Vaghari-Tabari, M.; Malakoti, F.; Moein, S.; Qujeq, D.; Yousefi, B.; Asemi, Z. Quercetin: An effective polyphenol in alleviating diabetes and diabetic complications. Crit. Rev. Food Sci. Nutr. 2022, 1–24. [Google Scholar] [CrossRef]
- AL-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Borger, A.; Stadlmayr, S.; Haertinger, M.; Semmler, L.; Supper, P.; Millesi, F.; Radtke, C. How miRNAs regulate schwann cells during peripheral nerve regeneration—A systemic review. Int. J. Mol. Sci. 2022, 23, 3440. [Google Scholar] [CrossRef]
- Rosso, G.; Wehner, D.; Schweitzer, C.; Möllmert, S.; Sock, E.; Guck, J.; Shahin, V. Matrix stiffness mechanosensing modulates the expression and distribution of transcription factors in Schwann cells. Bioeng. Transl. Med. 2022, 7, e10257. [Google Scholar] [PubMed]
- Ugwah-Oguejiofor, C.J.; Okoli, C.O.; Ugwah, M.O.; Umaru, M.L.; Ogbulie, C.S.; Mshelia, H.E.; Umar, M.; Njan, A.A. Acute and sub-acute toxicity of aqueous extract of aerial parts of Caralluma dalzielii NE Brown in mice and rats. Heliyon 2019, 5, e01179. [Google Scholar] [CrossRef] [Green Version]
- Yardim, A.; Kandemir, F.M.; Ozdemir, S.; Kucukler, S.; Comakli, S.; Gur, C.; Celik, H. Quercetin provides protection against the peripheral nerve damage caused by vincristine in rats by suppressing caspase 3, NF-κB, ATF-6 pathways and activating Nrf2, Akt pathways. Neurotoxicology 2020, 81, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Huang, C.-Y.; Tsai, F.-J.; Tsai, C.-C.; Yao, C.-H.; Chen, Y.-S. Growth-promoting effects of quercetin on peripheral nerves in rats. Int. J. Artif. Organs 2011, 34, 1095–1105. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajaber, M.A.; Hussain, G.; Farooq, T.; Noreen, R.; Ibrahim, M.; Umbreen, H.; Batool, S.; Rehman, K.; Hameed, A.; Farid, M.F.; et al. Nanosuspension of Foeniculum Vulgare Promotes Accelerated Sensory and Motor Function Recovery after Sciatic Nerve Injury. Metabolites 2023, 13, 391. https://doi.org/10.3390/metabo13030391
Bajaber MA, Hussain G, Farooq T, Noreen R, Ibrahim M, Umbreen H, Batool S, Rehman K, Hameed A, Farid MF, et al. Nanosuspension of Foeniculum Vulgare Promotes Accelerated Sensory and Motor Function Recovery after Sciatic Nerve Injury. Metabolites. 2023; 13(3):391. https://doi.org/10.3390/metabo13030391
Chicago/Turabian StyleBajaber, Majed A., Ghulam Hussain, Tahir Farooq, Razia Noreen, Muhammad Ibrahim, Huma Umbreen, Shaheera Batool, Kanwal Rehman, Arruje Hameed, Muhammad Fayyaz Farid, and et al. 2023. "Nanosuspension of Foeniculum Vulgare Promotes Accelerated Sensory and Motor Function Recovery after Sciatic Nerve Injury" Metabolites 13, no. 3: 391. https://doi.org/10.3390/metabo13030391