
Cross-Sectional Blood Metabolite Markers of Hypertension: A Multicohort Analysis of 44,306 Individuals from the COnsortium of METabolomics Studies

 , , , , ,
, , , , ,  , , , ,
, , , ,  , and add
Show full author list
, and add
Show full author list
Abstract
:1. Introduction
2. Results
2.1. Study Demographics
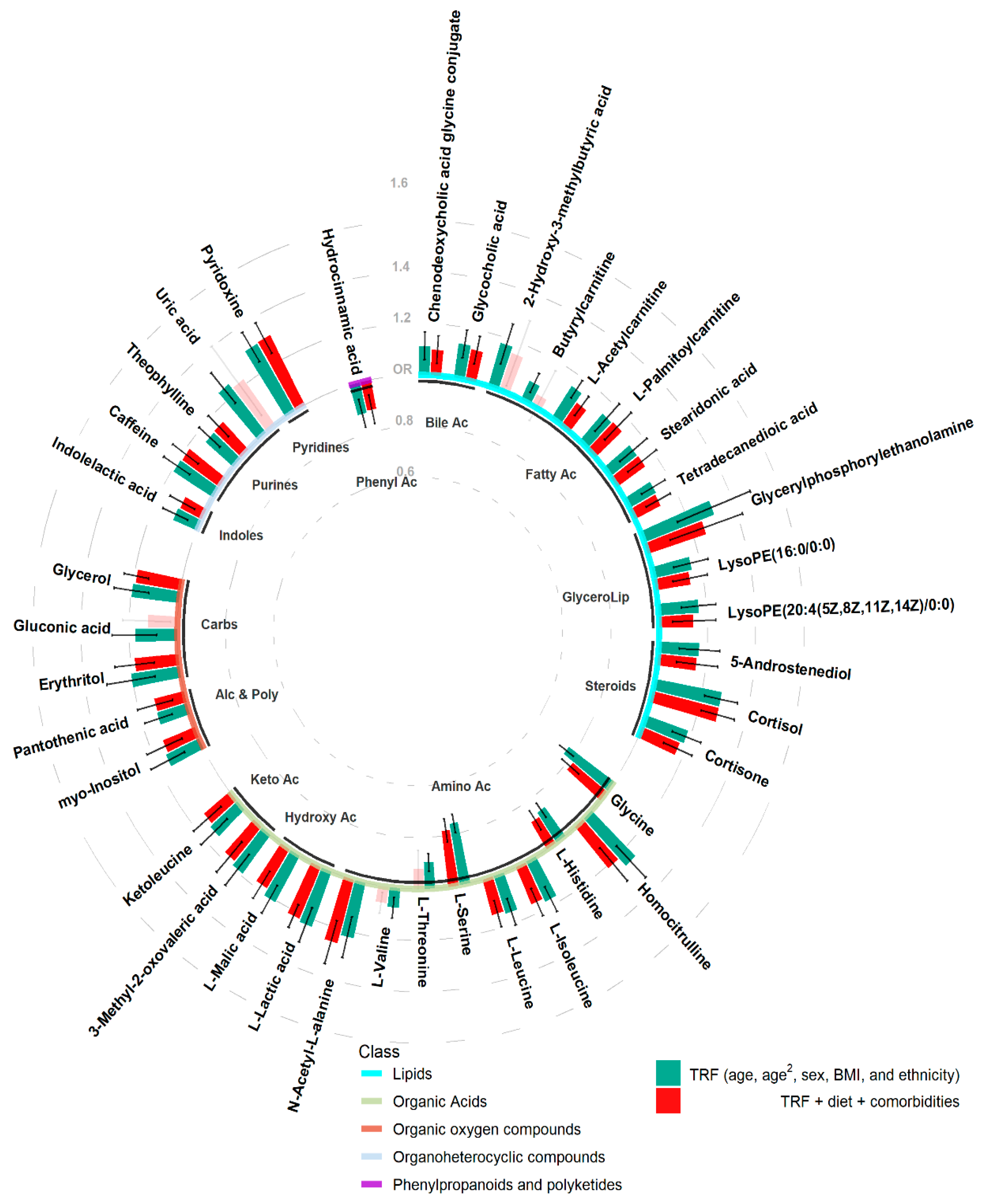
2.2. Discovery Analysis
2.3. Sensitivity Analysis
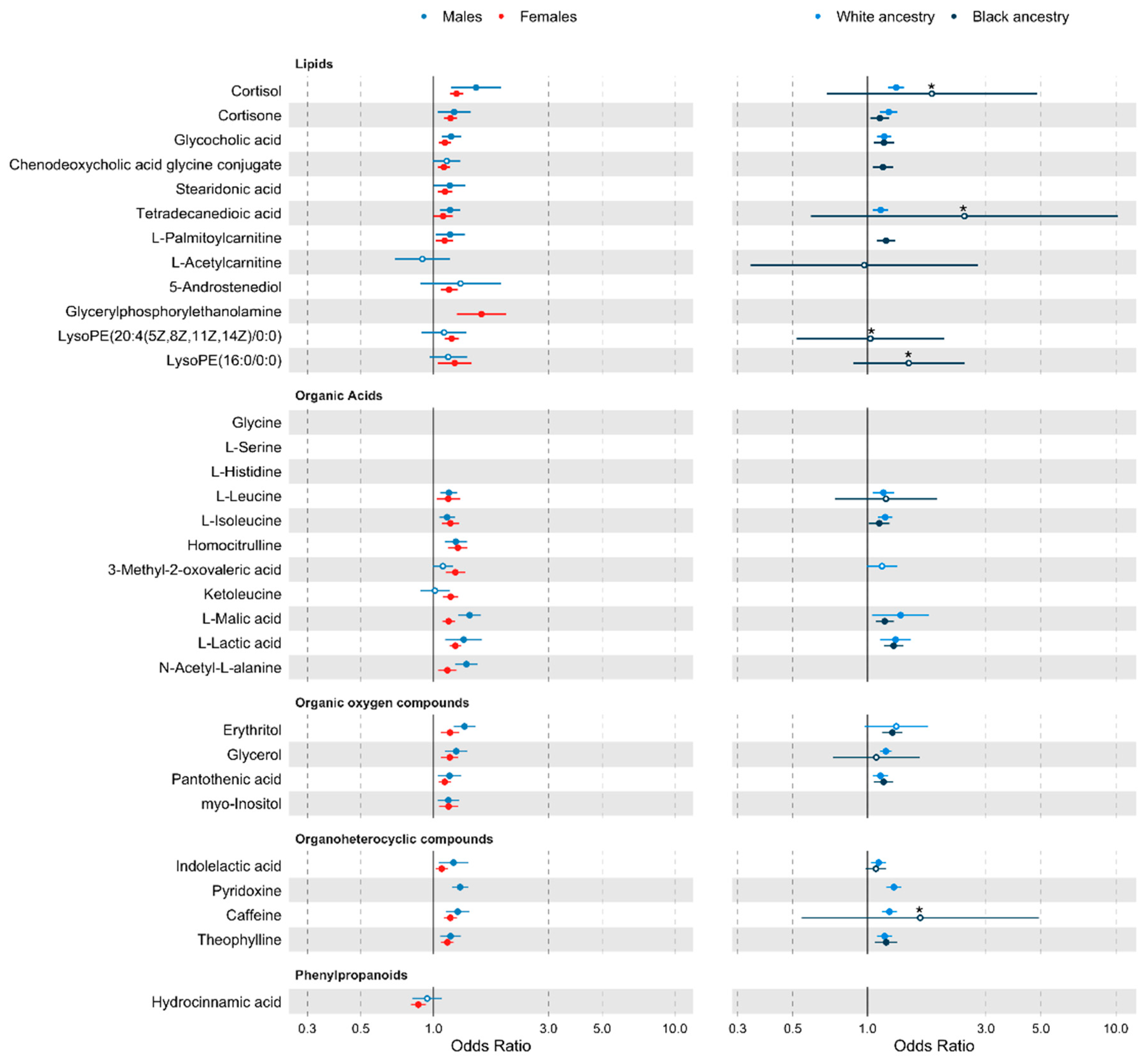
2.4. Stratified Analyses
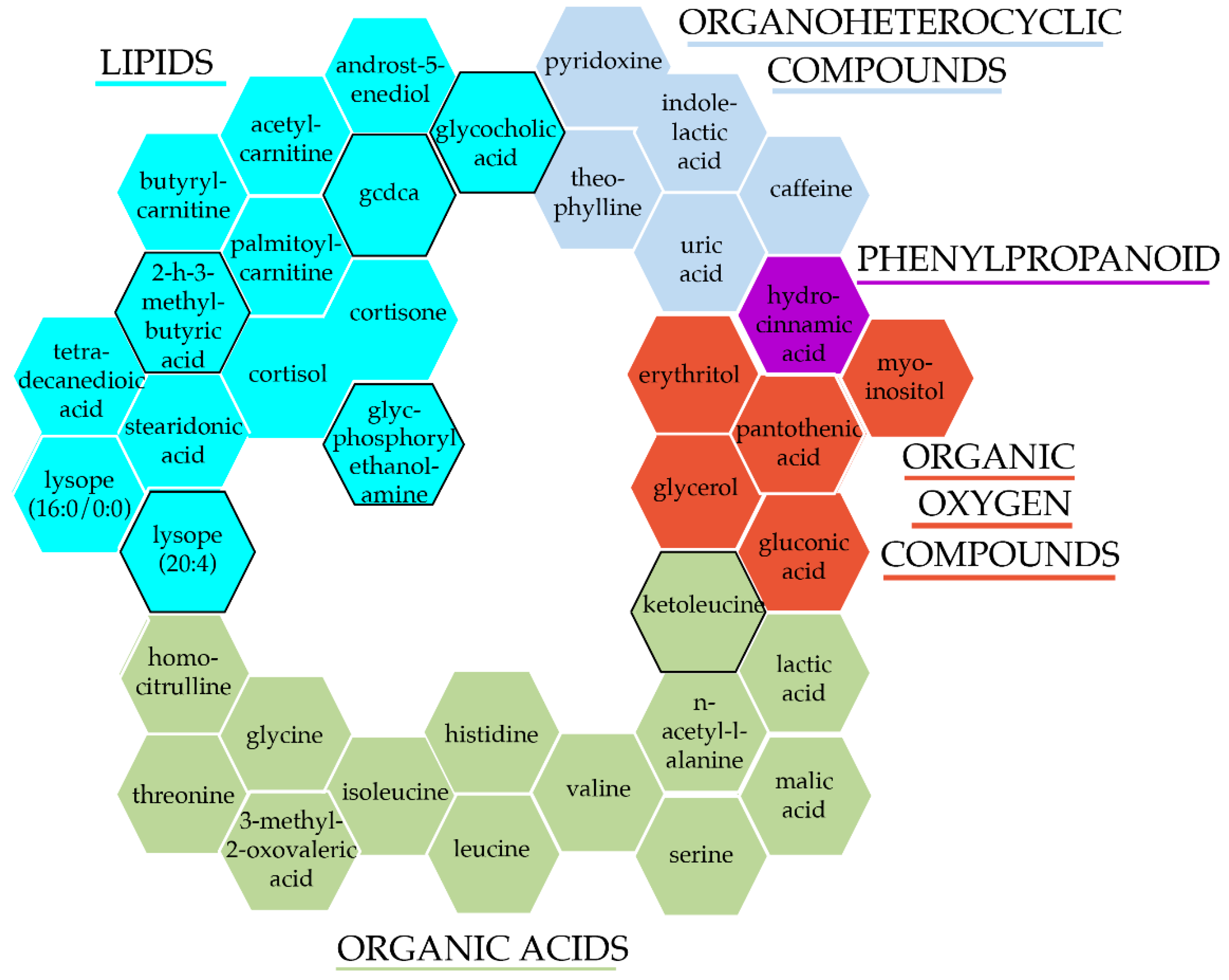
2.5. Pathway Analysis
3. Discussion
4. Methods
4.1. Study Populations
4.2. Phenotypes
4.2.1. Metabolomics
4.2.2. Blood Pressure
4.2.3. Covariates
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stergiou, G.S.; Palatini, P.; Parati, G.; O’Brien, E.; Januszewicz, A.; Lurbe, E.; Persu, A.; Mancia, G.; Kreutz, R.; on behalf of the European Society of Hypertension Council and the European Society of Hypertension Working Group on Blood Pressure Monitoring and Cardiovascular Variability; et al. 2021 European Society of Hypertension practice guidelines for office and out-of-office blood pressure measurement. J. Hypertens. 2021, 39, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Perel, P.; Mensah, G.A.; Ezzati, M. Global epidemiology, health burden and effective interventions for elevated blood pressure and hypertension. Nat. Rev. Cardiol. 2021, 18, 785–802. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, S.; Dominiczak, A.F. Genomics of hypertension: The road to precision medicine. Nat. Rev. Cardiol. 2021, 18, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Louca, P.; Menni, C.; Padmanabhan, S. Genomic Determinants of Hypertension With a Focus on Metabolomics and the Gut Microbiome. Am. J. Hypertens. 2020, 33, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, S.; Caulfield, M.; Dominiczak, A.F. Genetic and Molecular Aspects of Hypertension. Circ. Res. 2015, 116, 937–959. [Google Scholar] [CrossRef] [Green Version]
- Dzau, V.J.; Balatbat, C.A. Future of Hypertension. Hypertension 2019, 74, 450–457. [Google Scholar] [CrossRef]
- Menni, C.; Mangino, M.; Cecelja, M.; Psatha, M.; Brosnan, M.J.; Trimmer, J.; Mohney, R.P.; Chowienczyk, P.; Padmanabhan, S.; Spector, T.D.; et al. Metabolomic study of carotid-femoral pulse-wave velocity in women. J. Hypertens. 2015, 33, 791–796. [Google Scholar] [CrossRef] [Green Version]
- Menni, C.; Fauman, E.; Erte, I.; Perry, J.R.; Kastenmüller, G.; Shin, S.Y.; Petersen, A.K.; Hyde, C.; Psatha, M.; Ward, K.J.; et al. Biomarkers for type 2 diabetes and impaired fasting glucose using a nontargeted metabolomics approach. Diabetes 2013, 62, 4270–4276. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, S.; Floegel, A.; Weikert, C.; Prehn, C.; Adamski, J.; Pischon, T.; Boeing, H.; Drogan, D. Identification of Serum Metabolites Associated With Incident Hypertension in the European Prospective Investigation Into Cancer and Nutrition-Potsdam Study. Hypertension 2016, 68, 471–477. [Google Scholar] [CrossRef]
- Menni, C.; Graham, D.; Kastenmüller, G.; Alharbi, N.H.; Alsanosi, S.M.; McBride, M.; Mangino, M.; Titcombe, P.; Shin, S.Y.; Psatha, M.; et al. Metabolomic identification of a novel pathway of blood pressure regulation involving hexadecanedioate. Hypertension 2015, 66, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, G.S.; Mathur, R.S.; Margolius, H.S. Sex steroid hormones are altered in essential hypertension. J. Hypertens. 1989, 7, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yu, B.; Alexander, D.; Mosley, T.H.; Heiss, G.; Nettleton, J.A.; Boerwinkle, E. Metabolomics and Incident Hypertension Among Blacks. Hypertension 2013, 62, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Li, A.H.; Metcalf, G.A.; Muzny, D.M.; Morrison, A.C.; White, S.; Mosley, T.H.; Gibbs, R.A.; Boerwinkle, E. Loss-of-function variants influence the human serum metabolome. Sci. Adv. 2016, 2, e1600800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Yu, B.; Alexander, D.; Manolio, T.A.; Aguilar, D.; Coresh, J.; Heiss, G.; Boerwinkle, E.; Nettleton, J.A. Associations between Metabolomic Compounds and Incident Heart Failure Among African Americans: The ARIC Study. Am. J. Epidemiol. 2013, 178, 534–542. [Google Scholar] [CrossRef]
- Yu, B.; Zanetti, K.A.; Temprosa, M.; Albanes, D.; Appel, N.; Barrera, C.B.; Ben-Shlomo, Y.; Boerwinkle, E.; Casas, J.P.; Clish, C.; et al. The Consortium of Metabolomics Studies (COMETS): Metabolomics in 47 Prospective Cohort Studies. Am. J. Epidemiol. 2019, 188, 991–1012. [Google Scholar] [CrossRef] [Green Version]
- Bujak, R.; Mateo, J.; Blanco, I.; Izquierdo-García, J.L.; Dudzik, D.; Markuszewski, M.J.; Peinado, V.I.; Laclaustra, M.; Barberá, J.A.; Barbas, C.; et al. New Biochemical Insights into the Mechanisms of Pulmonary Arterial Hypertension in Humans. PLoS ONE 2016, 11, e0160505. [Google Scholar] [CrossRef]
- Pablo Arab, J.; Barrera, F.; Arrese, M. Bile Acids and Portal Hypertension. Ann. Hepatol. 2017, 16, S83–S86. [Google Scholar] [CrossRef]
- Tominaga, T.; Suzuki, H.; Ogata, Y.; Imafuku, T.; Saruta, T. Bile acids are able to reduce blood pressure by attenuating the vascular reactivity in spontaneously hypertensive rats. Life Sci. 1988, 42, 1861–1868. [Google Scholar] [CrossRef]
- Liu, L.; Su, J.; Li, R.; Luo, F. Changes in Intestinal Flora Structure and Metabolites Are Associated With Myocardial Fibrosis in Patients With Persistent Atrial Fibrillation. Front. Nutr. 2021, 8, 702085. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, Y.; Liu, Y.; Wang, X.; Guo, Z.; Zhang, Y.; Liang, X.; Lai, W. Effects of cholic acid on blood pressure and production of vascular aldosterone and corticosterone. Steroids 1999, 64, 291–295. [Google Scholar] [CrossRef]
- Ferreira, M.; Coxito, P.M.; Sardão, V.A.; Palmeira, C.M.; Oliveira, P.J. Bile acids are toxic for isolated cardiac mitochondria. Cardiovasc. Toxicol. 2005, 5, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Mels, C.M.; Delles, C.; Louw, R.; Schutte, A.E. Central systolic pressure and a nonessential amino acid metabolomics profile: The African Prospective study on the Early Detection and Identification of Cardiovascular disease and Hypertension. J. Hypertens. 2019, 37, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liu, Y.; Li, Z.; Song, Y.; Cai, X.; Liu, Y.; Zhang, T.; Yang, L.; Li, L.; Gao, S.; et al. Identification of essential hypertension biomarkers in human urine by non-targeted metabolomics based on UPLC-Q-TOF/MS. Clin. Chim. Acta 2018, 486, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Flores-Guerrero, J.L.; Groothof, D.; Connelly, M.A.; Otvos, J.D.; Bakker, S.J.L.; Dullaart, R.P.F. Concentration of Branched-Chain Amino Acids Is a Strong Risk Marker for Incident Hypertension. Hypertension 2019, 74, 1428–1435. [Google Scholar] [CrossRef]
- Akbaraly, T.; Würtz, P.; Singh-Manoux, A.; Shipley, M.J.; Haapakoski, R.; Lehto, M.; Desrumaux, C.; Kähönen, M.; Lehtimäki, T.; Mikkilä, V.; et al. Association of circulating metabolites with healthy diet and risk of cardiovascular disease: Analysis of two cohort studies. Sci. Rep. 2018, 8, 8620. [Google Scholar] [CrossRef]
- Polak-Iwaniuk, A.; Harasim-Symbor, E.; Gołaszewska, K.; Chabowski, A. How Hypertension Affects Heart Metabolism. Front. Physiol. 2019, 10, 435. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, G.; Hao, H.; Huang, Q.; Yan, B.; Zha, W.; Gu, S.; Ren, H.; Zhang, Y.; Fan, X.; et al. Gas chromatography/time-of-flight mass spectrometry based metabonomic approach to differentiating hypertension- and age-related metabolic variation in spontaneously hypertensive rats. Rapid Commun. Mass Spectrom. 2008, 22, 2882–2888. [Google Scholar] [CrossRef]
- Whitworth, J.A.; Williamson, P.M.; Mangos, G.; Kelly, J.J. Cardiovascular consequences of cortisol excess. Vasc. Health Risk Manag. 2005, 1, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Whitworth, J.A.; Saines, D.; Scoggins, B.A. Blood pressure and metabolic effects of cortisol and deoxycorticosterone in man. Clin. Exp. Hypertens. A 1984, 6, 795–809. [Google Scholar] [CrossRef]
- Robinson, O.; Carter, A.R.; Ala-Korpela, M.; Casas, J.P.; Chaturvedi, N.; Engmann, J.; Howe, L.D.; Hughes, A.D.; Järvelin, M.-R.; Kähönen, M.; et al. Metabolic profiles of socio-economic position: A multi-cohort analysis. Int. J. Epidemiol. 2020, 50, 768–782. [Google Scholar] [CrossRef] [PubMed]
- Al Kuwari, H.; Al Thani, A.; Al Marri, A.; Al Kaabi, A.; Abderrahim, H.; Afifi, N.; Qafoud, F.; Chan, Q.; Tzoulaki, I.; Downey, P.; et al. The Qatar Biobank: Background and methods. BMC Public Health 2015, 15, 1208. [Google Scholar] [CrossRef] [Green Version]
- Boyd, A.; Golding, J.; Macleod, J.; Lawlor, D.A.; Fraser, A.; Henderson, J.; Molloy, L.; Ness, A.; Ring, S.; Davey Smith, G. Cohort Profile: The ‘Children of the 90s’—The index offspring of the Avon Longitudinal Study of Parents and Children. Int. J. Epidemiol. 2012, 42, 111–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, A.; Macdonald-Wallis, C.; Tilling, K.; Boyd, A.; Golding, J.; Davey Smith, G.; Henderson, J.; Macleod, J.; Molloy, L.; Ness, A.; et al. Cohort Profile: The Avon Longitudinal Study of Parents and Children: ALSPAC mothers cohort. Int. J. Epidemiol. 2012, 42, 97–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northstone, K.; Lewcock, M.; Groom, A.; Boyd, A.; Macleod, J.; Timpson, N.; Wells, N. The Avon Longitudinal Study of Parents and Children (ALSPAC): An update on the enrolled sample of index children in 2019 [version 1; peer review: 2 approved]. Wellcome Open Res. 2019, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.A.; Taylor, R.; Thielke, R.; Payne, J.; Gonzalez, N.; Conde, J.G. Research electronic data capture (REDCap)—A metadata-driven methodology and workflow process for providing translational research informatics support. J. Biomed. Inform. 2009, 42, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.; Small, N.; Raynor, P.; Tuffnell, D.; Bhopal, R.; Cameron, N.; Fairley, L.; Lawlor, D.A.; Parslow, R.; Petherick, E.S.; et al. Cohort Profile: The Born in Bradford multi-ethnic family cohort study. Int. J. Epidemiol. 2012, 42, 978–991. [Google Scholar] [CrossRef] [Green Version]
- Garden, F.L.; Toelle, B.G.; Mihrshahi, S.; Webb, K.L.; Almqvist, C.; Tovey, E.R.; Brew, B.K.; Ayer, J.G.; Skilton, M.R.; Jones, G.; et al. Cohort profile: The Childhood Asthma Prevention Study (CAPS). Int. J. Epidemiol. 2018, 47, 1736–1736k. [Google Scholar] [CrossRef]
- Riboli, E.; Hunt, K.J.; Slimani, N.; Ferrari, P.; Norat, T.; Fahey, M.; Charrondière, U.R.; Hémon, B.; Casagrande, C.; Vignat, J.; et al. European Prospective Investigation into Cancer and Nutrition (EPIC): Study populations and data collection. Public Health Nutr. 2002, 5, 1113–1124. [Google Scholar] [CrossRef]
- Verdi, S.; Abbasian, G.; Bowyer, R.C.E.; Lachance, G.; Yarand, D.; Christofidou, P.; Mangino, M.; Menni, C.; Bell, J.T.; Falchi, M.; et al. TwinsUK: The UK Adult Twin Registry Update. Twin Res. Hum. Genet. 2019, 22, 523–529. [Google Scholar] [CrossRef] [Green Version]
- Marmot, M.; Brunner, E. Cohort Profile: The Whitehall II study. Int. J. Epidemiol. 2005, 34, 251–256. [Google Scholar] [CrossRef]
- Wright, J.D.; Folsom, A.R.; Coresh, J.; Sharrett, A.R.; Couper, D.; Wagenknecht, L.E.; Mosley, T.H.; Ballantyne, C.M.; Boerwinkle, E.A.; Rosamond, W.D.; et al. The ARIC (Atherosclerosis Risk In Communities) Study: JACC Focus Seminar 3/8. J. Am. Coll. Cardiol. 2021, 77, 2939–2959. [Google Scholar] [CrossRef] [PubMed]
- Santanasto, A.J.; Goodpaster, B.H.; Kritchevsky, S.B.; Miljkovic, I.; Satterfield, S.; Schwartz, A.V.; Cummings, S.R.; Boudreau, R.M.; Harris, T.B.; Newman, A.B. Body Composition Remodeling and Mortality: The Health Aging and Body Composition Study. J. Gerontol. Ser. A 2016, 72, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temprosa, M.; Moore, S.C.; Zanetti, K.A.; Appel, N.; Ruggieri, D.; Mazzilli, K.M.; Chen, K.L.; Kelly, R.S.; Lasky-Su, J.A.; Loftfield, E.; et al. COMETS Analytics: An online tool for analyzing and meta-analyzing metabolomics data in large research consortia. Am. J. Epidemiol. 2021, 191, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2017, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 3.6.3; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Rust, P.; Ekmekcioglu, C. Impact of Salt Intake on the Pathogenesis and Treatment of Hypertension. Adv. Exp. Med. Biol. 2017, 956, 61–84. [Google Scholar] [CrossRef]
- Al-Maqbali, S.R.; Mula-Abed, W.A. Comparison between Three Different Equations for the Estimation of Glomerular Filtration Rate in Omani Patients with Type 2 Diabetes Mellitus. Sultan Qaboos Univ. Med. J. 2014, 14, e197–e203. [Google Scholar]
- Bliss, C.I.; Greenwood, M.L.; White, E.S. A Rankit Analysis of Paired Comparisons for Measuring the Effect of Sprays on Flavor. Biometrics 1956, 12, 381–403. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef]
- Taylor, K.; McBride, N.; JGoulding, N.; Burrows, K.; Mason, D.; Pembrey, L.; Yang, T.; Azad, R.; Wright, J.; ALawlor, D. Metabolomics datasets in the Born in Bradford cohort [version 2; peer review: 1 approved, 1 approved with reservations]. Wellcome Open Res. 2021, 5, 264. [Google Scholar] [CrossRef]
- Goïta, Y.; Chao de la Barca, J.M.; Keïta, A.; Diarra, M.B.; Dembélé, K.C.; Chabrun, F.; Dramé, B.S.I.; Kassogué, Y.; Diakité, M.; Mirebeau-Prunier, D.; et al. Sexual Dimorphism of Metabolomic Profile in Arterial Hypertension. Sci. Rep. 2020, 10, 7517. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhong, S.; Hu, S.; Cheng, B.; Qiu, H.; Hu, Z. Changes of gut microbiome composition and metabolites associated with hypertensive heart failure rats. BMC Microbiol. 2021, 21, 141. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Tian, Y.; Gu, J.; Qiu, M.; Lu, Y.; Sun, W.; Kong, X. Comparative Study of Metabolite Changes after Antihypertensive Therapy with Calcium Channel Blockers or Angiotensin Type 1 Receptor Blockers. J. Cardiovasc. Pharmacol. 2021, 77, 228–237. [Google Scholar] [CrossRef]
- Hao, Y.; Wang, Y.; Xi, L.; Li, G.; Zhao, F.; Qi, Y.; Liu, J.; Zhao, D. A Nested Case-Control Study of Association between Metabolome and Hypertension Risk. BioMed Res. Int. 2016, 2016, 7646979. [Google Scholar] [CrossRef]
- Toba, H.; Nakamori, A.; Tanaka, Y.; Yukiya, R.; Tatsuoka, K.; Narutaki, M.; Tokitaka, M.; Hariu, H.; Kobara, M.; Nakata, T. Oral L-histidine exerts antihypertensive effects via central histamine H3 receptors and decreases nitric oxide content in the rostral ventrolateral medulla in spontaneously hypertensive rats. Clin. Exp. Pharm. Physiol. 2010, 37, 62–68. [Google Scholar] [CrossRef]
- Schwabedal, P.E.; Pietrzik, K.; Wittkowski, W. Pantothenic Acid Deficiency as a Factor Contributing to the Development of Hypertension. Cardiology 1985, 72 (Suppl. S1), 187–189. [Google Scholar] [CrossRef]
- Ho, A.; Sinick, J.; Esko, T.; Fischer, K.; Menni, C.; Zierer, J.; Matey-Hernandez, M.; Fortney, K.; Morgen, E.K. Circulating glucuronic acid predicts healthspan and longevity in humans and mice. Aging 2019, 11, 7694–7706. [Google Scholar] [CrossRef]
- Fregly, M.J.; Cade, J.R. Effect of Pyridoxine and Tryptophan, Alone and Combined, on the Development of Deoxycorticosterone Acetate-Induced Hypertension in Rats. Pharmacology 1995, 50, 298–306. [Google Scholar] [CrossRef]
- Dzúrik, R.; Fekovská, N.; Brimichová, G.; Tiso, P. Blood Pressure, 5-OH Indoleacetic Acid, and Vanilmandelic Acid Excretion and Blood Platelet Aggregation in Hypertensive Patients Treated with Ketanserin. J. Cardiovasc. Pharmacol. 1985, 7, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A. Anti-hypertensive Effect of Cereal Antioxidant Ferulic Acid and Its Mechanism of Action. Front. Nutr. 2019, 6, 121. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cohort | ALSPAC | ARIC | BIB | CaPS | EPIC | HealthABC | QBB | TwinsUK | Whitehall II |
|---|---|---|---|---|---|---|---|---|---|
| n | 9396 | 3293 | 1795 | 989 | 16,418 | 232 | 2906 | 4427 | 4850 |
| mean (SD) | mean (SD) | mean (SD) | mean (SD) | mean (SD) | mean (SD) | mean (SD) | mean (SD) | mean (SD) | |
| Age | 40.9 (12.5) | 52.9 (5.5) | 28 (5.7) | 61.4 (4.4) | 56.1 (8) | 74.7 (2.8) | 39 (12) | 54 (13.3) | 56.2 (6) |
| BMI | 26.1 (5) | 28.9 (5.9) | 26.8 (5.9) | 26.8 (3.7) | 26.7 (4.1) | 27.1 (4.4) | 28.9 (5.9) | 26.1 (4.8) | 26.3 (3.9) |
| SBP | 120.3 (13.8) | 129.8 (23.3) | 110.6 (11.8) | 148.6 (24.4) | 138.3 (22.5) | 138.1 (23.3) | 115.9 (17) | 127.4 (18.6) | 125.2 (18.3) |
| DBP | 71.1 (9.1) | 79.9 (13.5) | 65.6 (8.5) | 84.1 (13.2) | 84.8 (12.6) | 77.9 (14.3) | 74 (11.2) | 78.5 (10.8) | 78.6 (11.3) |
| n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | |
| HTN cases | 1007 (10.7) | 1501 (45.6) | 18 (1) | 631 (63.8) | 6897 (42) | 48 (20.7) | 409 (14.1) | 1322 (29.9) | 646 (13.3) |
| Non-HTN controls | 8389 (89.3) | 1792 (54.4) | 1777 (99) | 358 (36.2) | 9521 (58) | 184 (79.3) | 2497 (85.9) | 3105 (70.1) | 4204 (86.7) |
| Sex | |||||||||
| Males | 3062 (33) | 1375 (41.8) | 0 | 989 (100) | 8659 (52.7) | 232 (100) | 1457 (50.1) | 339 (7.7) | 3329 (68.6) |
| Females | 6334 (67) | 1918 (58.2) | 1795 (100) | 0 | 7759 (47.3) | 0 | 1449 (49.9) | 4088 (92.3) | 1521 (31.4) |
| Ancestry | |||||||||
| White | 8436 (89.8) | 1053 (32) | 842 (46.9) | 989 (100) | NA | 0 | 0 | 3653 (82.5) | 4475 (92.3) |
| Black | 40 (0.4) | 2240 (68) | 0 | 0 | NA | 232 (100) | 0 | 16 (0.4) | 103 (2.1) |
| Asian/Hispanic | 43 (0.5) | 0 | 953 (53.1) | 0 | NA | 0 | 2906 (100) | 27 (0.6) | 234 (4.8) |
| Other | 37 (0.4) | 0 | 0 | 0 | NA | 0 | 0 | 0 | 37 (0.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Louca, P.; Nogal, A.; Moskal, A.; Goulding, N.J.; Shipley, M.J.; Alkis, T.; Lindbohm, J.V.; Hu, J.; Kifer, D.; Wang, N.; et al. Cross-Sectional Blood Metabolite Markers of Hypertension: A Multicohort Analysis of 44,306 Individuals from the COnsortium of METabolomics Studies. Metabolites 2022, 12, 601. https://doi.org/10.3390/metabo12070601
Louca P, Nogal A, Moskal A, Goulding NJ, Shipley MJ, Alkis T, Lindbohm JV, Hu J, Kifer D, Wang N, et al. Cross-Sectional Blood Metabolite Markers of Hypertension: A Multicohort Analysis of 44,306 Individuals from the COnsortium of METabolomics Studies. Metabolites. 2022; 12(7):601. https://doi.org/10.3390/metabo12070601
Chicago/Turabian StyleLouca, Panayiotis, Ana Nogal, Aurélie Moskal, Neil J. Goulding, Martin J. Shipley, Taryn Alkis, Joni V. Lindbohm, Jie Hu, Domagoj Kifer, Ni Wang, and et al. 2022. "Cross-Sectional Blood Metabolite Markers of Hypertension: A Multicohort Analysis of 44,306 Individuals from the COnsortium of METabolomics Studies" Metabolites 12, no. 7: 601. https://doi.org/10.3390/metabo12070601