
Fungal Depsides—Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities

 ,
,  ,
,  ,
,
Abstract
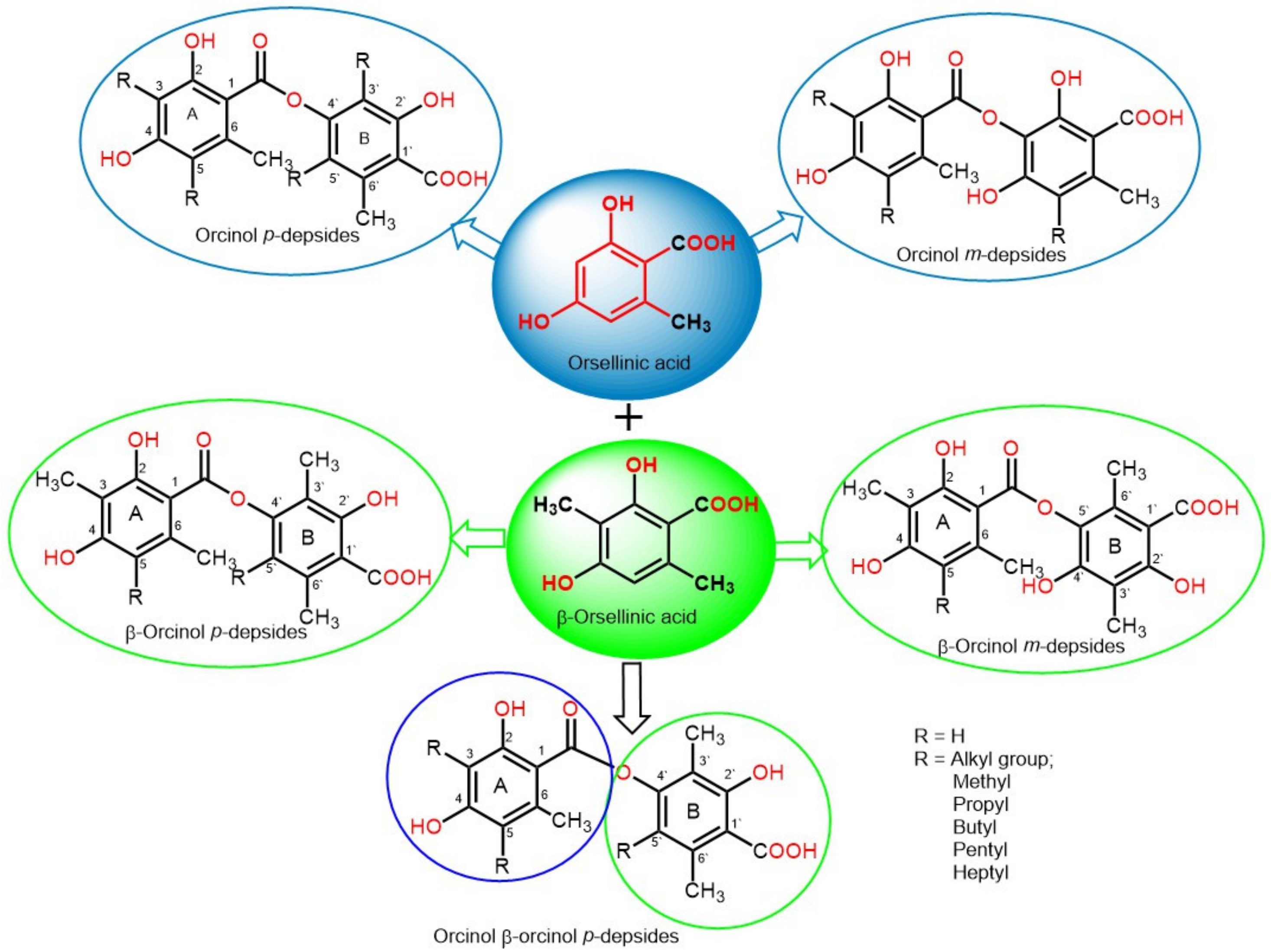
:1. Introduction
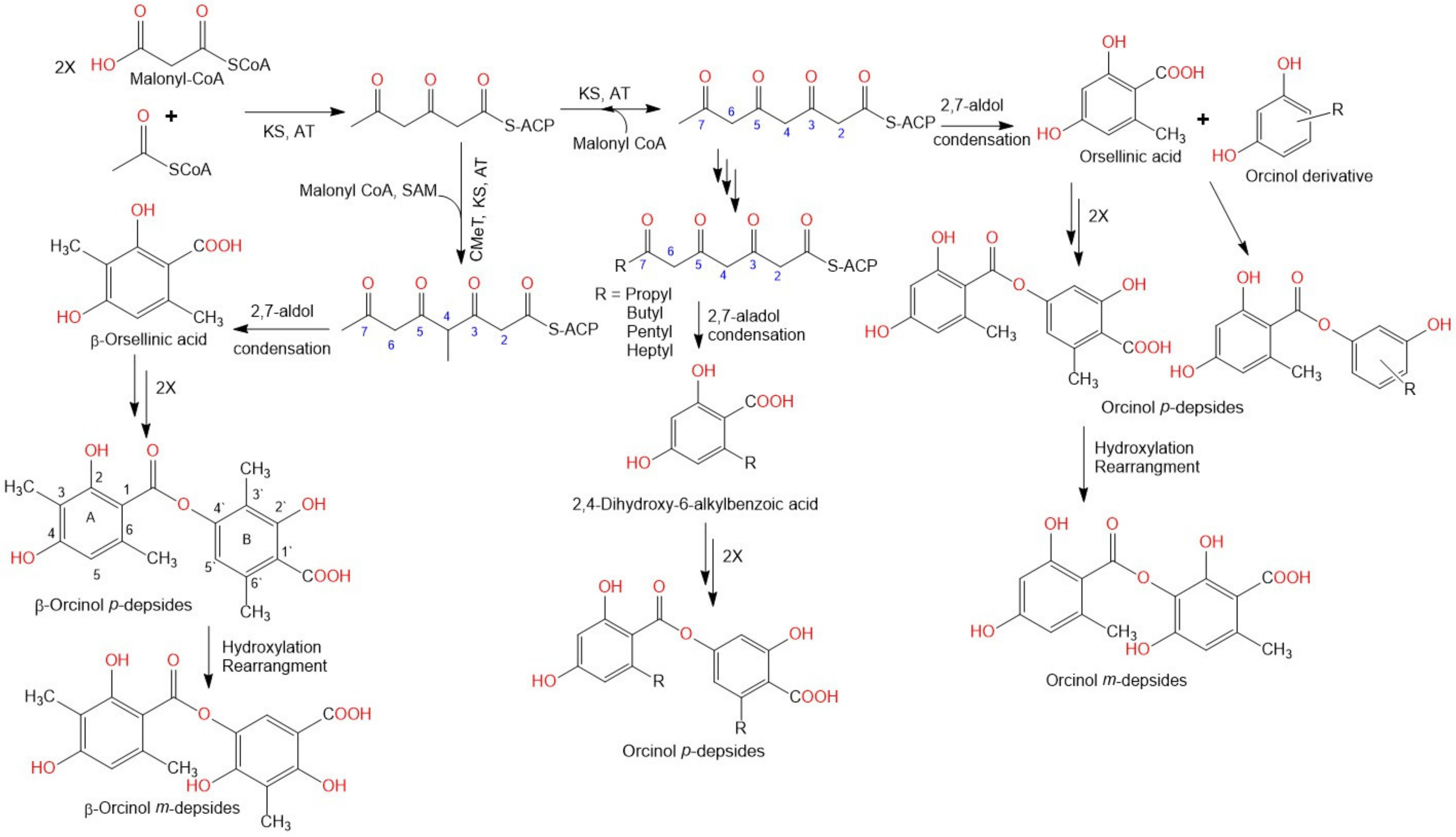
2. Biosynthesis of Depsides
3. Biological Activities
3.1. Antitumor Activity
3.2. Antimicrobial Activity
3.3. Antifouling Activity
3.4. Anti-Diabetic Activity
3.5. D-Glucose-6-Phosphate Phosphohydrolase Inhibitory Activity
3.6. α-Glucosidase Inhibitory (αGI) Activity
3.7. Protein Tyrosine Phosphatase Inhibitory (PTP1BI) Activity
3.8. Diacylglycerol Acyltransferase Inhibitory (DGATI) Activity
3.9. Activity of 11β-Hydroxysteroid Dehydrogenase Inhibitory (11β-HSDI) Enzyme
3.10. Anti-Inflammatory Activities
3.11. Antimalarial Activity
3.12. Antioxidant Activity
3.13. Ca2+/CaM Dependent Phosphodiesterase Inhibitory (CaM-PDEI) Activity
3.14. Antiviral Activity
3.15. Human Leukocyte Elastase (HLE) Inhibitory Activity
3.16. Indoleamine 2,3-Dioxygenase Inhibitory (IDOI) Activity
3.17. Adenosine Triphosphatase Inhibitory Activity
3.18. Proteasome Inhibitory Activity
3.19. Phospholipase Inhibitory Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spect. 2017, 5, 79–95. [Google Scholar] [CrossRef]
- Al-Rabia, M.W.; Mohamed, G.A.; Ibrahim, S.R.M.; Asfour, H.Z. Anti-inflammatory ergosterol derivatives from the endophytic fungus Fusarium chlamydosporum. Nat. Prod. Res. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Noor, A.O.; Almasri, D.M.; Bagalagel, A.A.; Abdallah, H.M.; Mohamed, S.G.A.; Mohamed, G.A.; Ibrahim, S.R.M. Naturally occurring isocoumarins derivatives from endophytic fungi: Sources, isolation, structural characterization, biosynthesis, and biological activities. Molecules 2020, 25, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Khayat, M.T.; Ibrahim, S.R.M.; Mohamed, G.A.; Abdallah, H.M. Anti-inflammatory metabolites from endophytic fungus Fusarium sp. Phytochem. Lett. 2019, 29, 104–109. [Google Scholar] [CrossRef]
- Sanchez, S.; Demain, A.L. Bioactive products from fungi. Food Bioact. 2017, 59–87. [Google Scholar]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.R.M.; Mohamed, G.A.; Kamal, H.M.K.; Mohamed, S.G.A.; Khedr, A.I.M. Terretonins from Aspergillus genus: Structures, biosynthesis, bioactivities, and structural elucidation. Mini-Rev. Org. Chem. 2021, 21, 2654. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Altyar, A.E.; Mohamed, S.G.A.; Mohamed, G.A. Genus Thielavia: Phytochemicals, industrial importance and biological relevance. Nat. Prod. Res. 2021, 5, 1–16. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Sindi, I.; Mohamed, G.A. Biologically active secondary metabolites and biotechnological applications of species of the family Chaetomiaceae (Sordariales): An updated review from 2016 to 2021. Mycol. Prog. 2021, 20, 595–639. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Altyar, A.E.; Mohamed, G.A. Humicola genus: Chemical constituents, industrial importance, and biological activities. Curr. Microbiol. 2021, 78, 2488–2509. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F.; Khayat, M.T. Biologically active fungal depsidones: Chemistry, biosynthesis, structural characterization, and bioactivities. Fitoterapia 2018, 129, 317–365. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Abdallah, H.M.; Elkhayat, E.S.; Al Musayeib, N.M.; Asfour, H.Z.; Zayed, M.F.; Mohamed, G.A. Fusaripeptide A: New antifungal and anti-malarial cyclodepsipeptide from the endophytic fungus Fusarium sp. J. Asian Nat. Prod. Res. 2018, 20, 75–85. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Zayed, M.F. Potential anti-malarial agents from endophytic fungi: A review. Mini. Rev. Med. Chem. 2018, 18, 1110–1132. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al Haidari, R.A.; El-Kholy, A.A.; Asfour, H.; Zayed, M.F. Fusaristerol A: A new ergosterol fatty acid ester from the endophytic fungus Fusarium sp. Pharmacog. Magaz. 2018, 14, 308–311. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Stocker-Wörgötter, E. Metabolic diversity of lichen-forming ascomycetous fungi: Culturing, polyketide and shikimate metabolite production, and PKS genes. Nat. Prod. Rep. 2008, 25, 188–200. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, L.S.; Abreu, L.M.; Nielsen, A.; Ingmer, H.; Larsen, T.O.; Nielsen, K.F.; Rodrigues-Filho, E. Dereplication-guided isolation of depsides thielavins S-T and lecanorins D-F from the endophytic fungus Setophoma sp. Phytochemistry 2015, 111, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Joshi, G.P.; Rawat, M.S.M. Lichens as a potential natural source of bioactive compounds: A review. Phytochem. Rev. 2010, 9, 303–314. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Tolstikov, G.A. Halogenated phenol compounds in lichens and fungi. Chem. Sustain. Dev. 2003, 11, 557–565. [Google Scholar]
- Seshadri, T.R. A Theory of Biogenesis of Lichen Depsides and Depsidones. Indian Acadademy Sci. 1944, 20, 1–14. [Google Scholar] [CrossRef]
- Jin, Q.; Hu, X.; Deng, Y.; Hou, J.; Lei, M.; Ji, H.; Zhou, J.; Qu, H.; Wu, W.; Guo, D. Four New Depsides isolated from Salvia miltiorrhiza and their significant nerve-protective activities. Molecules 2018, 23, 3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhang, X.; Cao, J.; Guo, Z.; Lou, Y.; Ding, M.; Zhao, Y. Depside derivatives with anti-hepatic fibrosis and anti-diabetic activities from Impatiens balsamina L. flowers. Fitoterapia 2015, 105, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhang, S.; Lu, X.; Ma, Y.; Fan, Y.; Shi, Y.; Dong, A.; Duan, B. Trivaric acid, a potent depside human leukocyte elastase inhibitor. Biol. Pharm. Bull. 2012, 35, 2247–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezawa, H.; Shibamoto, N.; Naganawa, H.; Ayukawa, S.; Matsuzaki, M.; Takeuchi, T.; Kono, K.; Sakamoto, T. Isolation of lecanoric acid, an inhibitor of histidine decarboxylase from a fungus. J. Antibiot. 1974, 27, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, Y.; Nishimura, K.; Kiriyama, N. Transformation of asterriquinone diacetate to asterriquinone monoalkyl ether via its monoacetal. Chem. Pharm. Bull. 1976, 24, 1853–1859. [Google Scholar] [CrossRef] [Green Version]
- Legaz, M.E.; de Armas, R.; Vicente, C. Bioproduction of Depsidones for Pharmaceutical Purposes; IntechOpen: Rijeka, Croatia, 2011; Volume 19, pp. 487–508. [Google Scholar]
- Muggia, L.; Grube, M. Type III polyketide synthases in lichen mycobionts. Fungal Biol. 2010, 114, 379–385. [Google Scholar] [PubMed]
- Schümann, J.; Hertweck, C. Advances in cloning, functional analysis and heterologous expression of fungal polyketide synthase genes. J. Biotechnol. 2006, 124, 690–703. [Google Scholar] [CrossRef]
- Rawlings, B. Biosynthesis of polyketides (other than Actinomycete Macrolides). Nat. Prod. Rep. 1999, 16, 425–484. [Google Scholar] [CrossRef]
- Elix, J.A. Biochemistry and secondary metabolites. In Lichen Biology; Cambridge University Press: Cambridge, UK, 1996; pp. 154–180. [Google Scholar]
- Elix, J.A.; Gaul, K.L. The interconversion of the lichen depsides para- and meta-scrobiculin, and the biosynthetic implications. Aust. J. Chem. 1986, 39, 613–624. [Google Scholar] [CrossRef]
- Nagai, H.; Kim, Y.H. Cancer prevention from the perspective of global cancer burden patterns. J. Thorac. Dis. 2017, 9, 448–451. [Google Scholar] [CrossRef]
- Ward, Z.J.; Scott, A.M.; Hricak, H.; Atun, R. Global costs, health benefits, and economic benefits of scaling up treatment and imaging modalities for survival of 11 cancers: A simulation-based analysis. Lancet Oncol. 2021, 22, 341–350. [Google Scholar] [CrossRef]
- Kornienko, A.; Evidente, A.; Vurro, M.; Mathieu, V.; Cimmino, A.; Evidente, M.; van Otterlo, W.A.; Dasari, R.; Lefranc, F.; Kiss, R. Toward a cancer drug of fungal rrigin. Med. Res. Rev. 2015, 35, 937–967. [Google Scholar] [CrossRef] [PubMed]
- Lünne, F.; Niehaus, E.M.; Lipinski, S.; Kunigkeit, J.; Kalinina, S.A.; Humpf, H.U. Identification of the polyketide synthase PKS7 responsible for the production of lecanoric acid and ethyl lecanorate in Claviceps purpurea. Fungal Genet. Biol. 2020, 145, 103481. [Google Scholar] [CrossRef]
- Phainuphong, P.; Rukachaisirikul, V.; Phongpaichit, S.; Sakayaroj, J.; Kanjanasirirat, P.; Borwornpinyo, S.; Akrimajirachoote, N.; Yimnual, C.; Muanprasat, C. Depsides and depsidones from the soil-derived fungus Aspergillus unguis PSU-RSPG204. Tetrahedron 2018, 74, 5691–5699. [Google Scholar] [CrossRef]
- Kurasawa, S.; Takeuchi, T.; Umezawa, H. Studies on glyoxalase inhibitor: Isolation of a new active agent, MS-3, from a mushroom. Agric. Biol. Chem. 1975, 39, 2003–2008. [Google Scholar] [CrossRef]
- Ma, K.; Bao, L.; Han, J.; Jin, T.; Yang, X.; Zhao, F.; Li, S.; Song, F.; Liu, M.; Liu, H. New benzoate derivatives and hirsutane type sesquiterpenoids with antimicrobial activity and cytotoxicity from the solid-state fermented rice by the medicinal mushroom Stereum hirsutum. Food Chem. 2014, 143, 239–245. [Google Scholar] [CrossRef]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Khonsanit, A.; Komwijit, S. Phenolic glycosides from the filamentous fungus Acremonium sp. BCC 14080. J. Nat. Prod. 2008, 71, 891–894. [Google Scholar] [CrossRef]
- Perlatti, B.; Lan, N.; Earp, C.E.; AghaAmiri, S.; Vargas, S.H.; Azhdarinia, A.; Bills, G.F.; Gloer, J.B. Arenicolins: C-glycosylated depsides from Penicillium arenicola. J. Nat. Prod. 2020, 83, 668–674. [Google Scholar] [CrossRef]
- Ko, H.R.; Kim, B.Y.; Oh, W.K.; Kang, D.O.; Lee, H.S.; Koshino, H.; Osada, H.; Mheen, T.I.; Ahn, J.S. CRM646-A and -B, novel fungal metabolites that inhibit heparinase. J. Antibiot. 2000, 53, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhang, Z.; Yu, B. Total synthesis of CRM646-A and -B, two fungal glucuronides with potent heparinase inhibition activities. J. Org. Chem. 2005, 70, 8884–8889. [Google Scholar] [CrossRef]
- Asami, Y.; Kim, S.O.; Jang, J.P.; Ko, S.K.; Kim, B.Y.; Osada, H.; Jang, J.H.; Ahn, J.S. CRM646-A, a fungal metabolite, induces nucleus condensation by increasing Ca2+ levels in rat 3Y1 fibroblast cells. J. Microbiol. Biotechnol. 2020, 30, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, W.; Zou, G.; Chen, S.; Pang, J.; She, Z. Bioactive polyketides from the mangrove endophytic fungi Phoma sp. SYSU-SK-7. Fitoterapia 2019, 139, 104369. [Google Scholar] [CrossRef] [PubMed]
- Togashi, K.; Ko, H.R.; Ahn, J.S.; Osada, H. Inhibition of telomerase activity by fungus metabolites, CRM646-A and thielavin B. Biosci. Biotechnol. Biochem. 2001, 65, 651–653. [Google Scholar] [CrossRef] [PubMed]
- Ayers, S.; Ehrmann, B.M.; Adcock, A.F.; Kroll, D.J.; Wani, M.C.; Pearce, C.J.; Oberlies, N.H. Thielavin B methyl ester: A cytotoxic benzoate trimer from an unidentified fungus (MSX 55526) from the order Sordariales. Tetrahedron Lett. 2011, 52, 5733–5735. [Google Scholar] [CrossRef] [Green Version]
- De Medeiros, L.S.; Murgu, M.; de Souza, A.Q.L.; Rodrigues-Fo, E. Antimicrobial depsides produced by Cladosporium uredinicola, an endophytic fungus isolated from Psidium guajava fruits. Helv. Chim. Acta. 2011, 94, 1077–1084. [Google Scholar] [CrossRef]
- Cao, S.; Lee, A.S.; Huang, Y.; Flotow, H.; Ng, S.; Butler, M.S.; Buss, A.D. Agonodepsides a and B: Two new depsides from a filamentous fungus F7524. J. Nat. Prod. 2002, 65, 1037–1038. [Google Scholar] [CrossRef]
- Nielsen, J.; Nielsen, P.H.; Frisvad, J.C. Fungal depside, guisinol, from a marine derived strain of Emericella unguis. Phytochemistry 1999, 50, 263–265. [Google Scholar] [CrossRef]
- Han, Z.; Li, Y.X.; Liu, L.L.; Lu, L.; Guo, X.R.; Zhang, X.X.; Zhang, X.Y.; Qi, S.H.; Xu, Y.; Qian, P.Y. Thielavins W-Z₇, New Antifouling Thielavins from the Marine-Derived Fungus Thielavia sp. UST030930-004. Mar. Drugs 2017, 15, 128. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.T.; Qi, Q.Y.; Ma, K.; Pei, Y.F.; Han, J.J.; Xu, W.; Li, E.W.; Liu, H.W. Depside α-glucosidase inhibitors from a culture of the mushroom Stereum hirsutum. Planta Med. 2014, 80, 918–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, H.; Iitaka, Y.; Kurasawa, S.; Takeuchi, T.; Umezawa, H. The crystal structure of a dibromo-derivative of MS-3: A glyoxalase I inhibitor produced by a mushroom, Stereum hirsutum. Agric. Biol. Chem. 1976, 40, 1781–1784. [Google Scholar] [CrossRef]
- Kurasawa, S.; Naganawa, H.; Takeuchi, T.; Umezawa, H. The structure of MS-3: A glyoxalase I inhibitor produced by a mushroom. Agric. Biol. Chem. 1975, 39, 2009–2014. [Google Scholar]
- Yasuzawa, T.; Saitoh, Y.; Sano, H. Structures of KS-501 and KS-502, the new inhibitors of Ca2+ and calmodulin-dependent cyclic nucleotide phosphodiesterase. J. Antibiot. 1990, 43, 336–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqueveque, P.; Céspedes, C.L.; Becerra, J.; Dávila, M.; Sterner, O. Bioactive compounds isolated from submerged fermentations of the Chilean fungus Stereum rameale. Z. Naturforsch. C. J. Biosci. 2015, 70, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Aqueveque, P.; Céspedes, C.L.; Becerra, J.; Aranda, M.; Sterner, O. Antifungal activities of secondary metabolites isolated from liquid fermentations of Stereum hirsutum (Sh134-11) against Botrytis cinerea (grey mould agent). Food Chem. Toxicol. 2017, 109, 1048–1054. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Q.; Dou, Q.P.; Yang, H. Discovery of natural proteasome inhibitors as novel anticancer therapeutics: Current status and perspectives. Curr. Protein Pept. Sci. 2018, 19, 358–367. [Google Scholar] [CrossRef]
- Seo, C.; Sohn, J.H.; Oh, H.; Kim, B.Y.; Ahn, J.S. Isolation of the protein tyrosine phosphatase 1B inhibitory metabolite from the marine-derived fungus Cosmospora sp. SF-5060. Bioorg. Med. Chem. Lett. 2009, 19, 6095–6097. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Fang, Y.; Xu, G.H.; Kim, W.G. Aquastatin A, a new inhibitor of enoyl-acyl carrier protein reductase from Sporothrix sp. FN611. Biol. Pharm. Bull. 2009, 32, 2061–2064. [Google Scholar] [CrossRef] [Green Version]
- Sakemi, S.; Hirai, H.; Ichiba, T.; Inagaki, T.; Kato, Y.; Kojima, N.; Nishida, H.; Parker, J.C.; Saito, T.; Tonai-Kachi, H. Thielavins as glucose-6-phosphatase (G6Pase) inhibitors: Producing strain, fermentation, isolation, structural elucidation and biological activities. J. Antibiot. 2002, 55, 941–951. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, S.; Ando, K.; Kawamoto, I.; Kase, H. KS-501 and KS-502, new inhibitors of Ca2+ and calmodulin-dependent cyclic-nucleotide phosphodiesterase from Sporothrix sp. J. Antibiot. 1989, 42, 1049–1055. [Google Scholar] [CrossRef]
- Ito-Kobayashi, M.; Aoyagi, A.; Tanaka, I.; Muramatsu, Y.; Umetani, M.; Takatsu, T. Sterenin A, B, C and D, novel 11beta-hydroxysteroid dehydrogenase type 1 inhibitors from Stereum sp. SANK 21205. J. Antibiot. 2008, 61, 128–135. [Google Scholar] [CrossRef]
- Zou, W.X.; Meng, J.C.; Lu, H.; Chen, G.X.; Shi, G.X.; Zhang, T.Y.; Tan, R.X. Metabolites of Colletotrichum gloeosporioides, an endophytic fungus in Artemisia mongolica. J. Nat. Prod. 2000, 63, 1529–1530. [Google Scholar] [CrossRef] [PubMed]
- Toki, S.; Ando, K.; Yoshida, M.; Matsuda, Y. PS-990, a novel neuritogenic compound from Acremonium sp. J. Antibiot. 1994, 47, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Chávez, J.; González-Andrade, M.; González, M.; Glenn, A.E.; Mata, R. Thielavins A, J and K: α-Glucosidase inhibitors from MEXU 27095, an endophytic fungus from Hintonia latiflora. Phytochemistry 2013, 94, 198–205. [Google Scholar] [CrossRef]
- Bejček, J.; Spiwok, V.; Kmoníčková, E.; Rimpelová, S. Na+/K+-ATPase Revisited: On Its Mechanism of Action, Role in Cancer, and Activity Modulation. Molecules 2021, 26, 1905. [Google Scholar]
- Matsumoto, K.; Tanaka, K.; Matsutani, S.; Sakazaki, R.; Hinoo, H.; Uotani, N.; Tanimoto, T.; Kawamura, Y.; Nakamoto, S.; Yoshida, T. Isolation and biological activity of thielocins: Novel phospholipase A2 inhibitors produced by Thielavia terricola RF-143. J. Antibiot. 1995, 48, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Huang, J.; Zhou, X.; Lin, X.; Liu, J.; Liao, S.; Wang, J.; Liu, F.A.; Tao, H.; Liu, Y. The fungal metabolites with potential antiplasmodial activity. Curr. Med. Chem. 2018, 25, 3796–3825. [Google Scholar] [CrossRef] [PubMed]
- Inokoshi, J.; Takagi, Y.; Uchida, R.; Masuma, R.; Omura, S.; Tomoda, H. Production of a new type of amidepsine with a sugar moiety by static fermentation of Humicola sp. FO-2942. J. Antibiot. 2010, 63, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Tomoda, H.; Tabata, N.; Ito, M.; Omura, S. Amidepsines, inhibitors of diacylglycerol acyltransferase produced by Humicola sp. FO-2942. II. Structure elucidation of amidepsines A, B and C. J. Antibiot. 1995, 48, 942–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M.; Nazir, M.; Hussain, H.; Tousif, M.I.; Elsebai, M.F.; Riaz, N.; Akhtar, N. Natural phenolics as inhibitors of the human neutrophil elastase (HNE) release: An overview of natural anti-inflammatory discoveries during recent years. Antiinflamm. Antiallergy Agents Med. Chem. 2018, 17, 70–94. [Google Scholar] [CrossRef]
- Tomoda, H.; Ito, M.; Tabata, N.; Masuma, R.; Yamaguchi, Y.; Omura, S. Amidepsines, inhibitors of diacylglycerol acyltransferase produced by Humicola sp. FO-2942. I. Production, isolation and biological properties. J. Antibiot. 1995, 48, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Matsutani, S.; Sakazaki, R.; Hinoo, H.; Terui, Y.; Tanaka, K.; Matsumoto, K.; Yoshida, T. 19 Structural elucidation of thielocins: Novel potent phospholipase A-2 inhibitors from a fungus. Tennen Yuki Kagobutsu Toronkai Koen Yoshishu 1992, 34, 142–149. [Google Scholar]
- Tomoda, H.; Yamaguchi, Y.; Tabata, N.; Kobayashi, T.; Masuma, R.; Tanaka, H.; Omura, S. Amidepsine E, an inhibitor of diacylglycerol acyltransferase produced by Humicola sp. FO-5969. J. Antibiot. 1996, 49, 929–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Nishida, H.; Pang, C.H.; Saito, T.; Sakemi, S.; Tonai-Kachi, H.; Yoshikawa, N.; Vanvolkenburg, M.A.; Parker, J.C.; Kojima, Y. CJ-21,164, a new D-glucose-6-phosphate phosphohydrolase inhibitor produced by a fungus Chloridium sp. J. Antibiot. 2002, 55, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Matsutani, S.; Matsumoto, K.; Yoshida, T. Effect of thielocin A1 beta on bee venom phospholipase A2-induced edema in mouse paw. Eur. J. Pharmacol. 1995, 279, 143–148. [Google Scholar] [CrossRef]
- Nagamitsu, T.; Marumoto, K.; Nagayasu, A.; Fukuda, T.; Arima, S.; Uchida, R.; Ohshiro, T.; Harigaya, Y.; Tomoda, H.; Omura, S. Total synthesis of amidepsine B and revision of its absolute configuration. J. Antibiot. 2009, 62, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Matsutani, S.; Matsumoto, K.; Yoshida, T. A novel type of phospholipase A2 inhibitor, thielocin A1 beta, and mechanism of action. J. Antibiot. 1992, 45, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Kitahara, N.; Haruyama, H.; Hata, T.; Takahashi, S. The structures of thielavins A., B and C. Prostaglandin synthetase inhibitors from fungi. J. Antibiot. 1983, 36, 599–600. [Google Scholar] [CrossRef] [Green Version]
- Inokoshi, J.; Kawamoto, K.; Takagi, Y.; Matsuhama, M.; Omura, S.; Tomoda, H. Expression of two human acyl-CoA:diacylglycerol acyltransferase isozymes in yeast and selectivity of microbial inhibitors toward the isozymes. J. Antibiot. 2009, 62, 51–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Nakamoto, S.; Sakazaki, R.; Matsumoto, K.; Terui, Y.; Sato, T.; Arita, H.; Matsutani, S.; Inoue, K.; Kudo, I. Thielocins A1 alpha and A1 beta, novel phospholipase A2 inhibitors from ascomycetes. J. Antibiot. 1991, 44, 1467–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, M.; Kudo, I.; Nakamura, H.; Yokoyama, Y.; Mori, H.; Inoue, K. Exacerbation of rat adjuvant arthritis by intradermal injection of purified mammalian 14-kDa group II phospholipase A2. FEBS Lett. 1990, 268, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, D.; Dussart, F. Selected fungal natural products with antimicrobial properties. Molecules 2020, 25, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M.; White, S.W.; Rock, C.O. Inhibiting bacterial fatty acid synthesis. J. Biol. Chem. 2006, 281, 17541–17544. [Google Scholar] [CrossRef] [Green Version]
- Moir, D.T. Identification of inhibitors of bacterial enoyl-acyl carrier protein reductase. Curr. Drug Targets Infect. Disord. 2005, 5, 297–305. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Sohn, M.J.; Kim, W.G. Aquastatin C, a new glycoaromatic derivative from Sporothrix sp. FN611. J. Antibiot. 2011, 64, 213–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, N.; Sancheti, P.; Jiang, Z.D.; McNaney, C.; DeCenzo, M.; Knight, B.; Stankis, M.; Kuranda, M.; Rothstein, D.M. Screening systems for detecting inhibitors of cell wall transglycosylation in Enterococcus. Cell wall transglycosylation inhibitors in Enterococcus. J. Antibiot. 1998, 51, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2017, 8, 361–376. [Google Scholar] [CrossRef]
- Van Schaftingen, E.; Gerin, I. The glucose-6-phosphatase system. Biochem. J. 2002, 362, 513–532. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Zayed, M.F.; Ross, S.A. 8-Hydroxyirilone 5-methyl ether and 8-hydroxyirilone, new antioxidant and α-amylase inhibitors isoflavonoids from Iris germanica rhizomes. Bioorg. Chem. 2017, 70, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khayat, M.T.; Ahmed, S.; Abo-Haded, H.; Alshali, K.Z. Mangostanaxanthone VIIII, a new xanthone from Garcinia mangostana pericarps, α-amylase inhibitory activity, and molecular docking studies. Rev. Bras. Farmacogn. 2019, 29, 206–212. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khayat, M.T.; Ahmed, S.; Abo-Haded, H. Garcixanthone D, a new xanthone, and other xanthone derivatives from Garcinia mangostana Pericarps: Their α-amylase inhibitory potential and molecular docking studies. Starch-Stärke 2019, 71, 1800354. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khayat, M.T.; Ahmed, S.; Abo-Haded, H. α-Amylase inhibitors xanthones from Garcinia mangostana pericarps and its possible use for the treatment of diabetes with their molecular docking studies. J. Food Biochem. 2019, 43, e12844. [Google Scholar] [CrossRef]
- Kennedy, B.P. Role of protein tyrosine phosphatase-1B in diabetes and obesity. Biomed. Pharmacother. 1999, 53, 466–470. [Google Scholar] [CrossRef]
- King, A.J.; Judd, A.S.; Souers, A.J. Inhibitors of diacylglycerol acyltransferase: A review of 2008 patents. Expert Opin. Ther. Pat. 2010, 20, 19–29. [Google Scholar] [CrossRef]
- Ferris, H.A.; Kahn, C.R. New mechanisms of glucocorticoid-induced insulin resistance: Make no bones about it. J. Clin. Invest. 2012, 122, 3854–3857. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Wang, Y.; Li, W.; Zhang, H.; Wang, X.; Mu, Q.; He, Z.; Yao, H. Esculin exhibited anti-inflammatory activities in vivo and regulated TNF-α and IL-6 production in LPS-stimulated mouse peritoneal macrophages in vitro through MAPK pathway. Int. Immunopharmacol. 2015, 29, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Kitahara, N.; Endo, A.; Furuya, K.; Takahashi, S. Thielavin A and B, new inhibitors of prostaglandin biosynthesis produced by Thielavia terricola. J. Antibiot. 1981, 34, 1562–1568. [Google Scholar] [CrossRef] [Green Version]
- Nieves, K.; Prudhomme, J.; Le Roch, K.G.; Franzblau, S.G.; Rodríguez, A.D. Natural product-based synthesis of novel anti-infective isothiocyanate- and isoselenocyanate-functionalized amphilectane diterpenes. Bioorg. Med. Chem. Lett. 2016, 26, 854–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Luis, S.; Pérez-Vásquez, A.; Mata, R. Natural products with calmodulin inhibitor properties. Phytochemistry 2007, 68, 1882–1903. [Google Scholar] [CrossRef] [PubMed]
- Mata, R.; Figueroa, M.; González-Andrade, M.; Rivera-Chávez, J.A.; Madariaga-Mazón, A.; Del Valle, P. Calmodulin inhibitors from natural sources: An update. J. Nat. Prod. 2015, 78, 576–586. [Google Scholar] [CrossRef]
- Kakkar, R.; Raju, R.V.; Sharma, R.K. Calmodulin-dependent cyclic nucleotide phosphodiesterase (PDE1). Cell Mol. Life Sci. 1999, 55, 1164–1186. [Google Scholar] [CrossRef] [PubMed]
- Toki, S.; Yoshida, M.; Ando, K.; Matsuda, Y. PS-990, a novel microbial metabolite, reversibly induces neurite extension in neuroblastoma cells. Biosci. Biotechnol. Biochem. 1995, 59, 1281–1286. [Google Scholar] [CrossRef]
- Evers, D.L.; Chao, C.F.; Wang, X.; Zhang, Z.; Huong, S.M.; Huang, E.S. Human cytomegalovirus-inhibitory flavonoids: Studies on antiviral activity and mechanism of action. Antiviral Res. 2005, 68, 124–134. [Google Scholar] [CrossRef]
- Guo, B.; Dai, J.R.; Ng, S.; Huang, Y.; Leong, C.; Ong, W.; Carté, B.K. Cytonic acids A and B: Novel tridepside inhibitors of hCMV protease from the endophytic fungus Cytonaema species. J. Nat. Prod. 2000, 63, 602–604. [Google Scholar] [CrossRef]
- Jang, J.P.; Jang, J.H.; Oh, M.; Son, S.; Kim, S.M.; Kim, H.M.; Shin, K.S.; Oh, H.; Soung, N.K.; Hong, Y.S. Inhibition of indoleamine 2,3-dioxygenase by thielavin derivatives from a soil fungus, Coniochaeta sp. 10F058. J. Antibiot. 2014, 67, 331–333. [Google Scholar] [CrossRef] [Green Version]
- Tjandrawinata, R.R.; Nailufar, F.; Arifin, P.F. Hydrogen potassium adenosine triphosphatase activity inhibition and downregulation of its expression by bioactive fraction DLBS2411 from Cinnamomum burmannii in gastric parietal cells. Int. J. Gen. Med. 2013, 6, 807–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamano, K.; Kinoshita-Okami, M.; Minagawa, K.; Haruyama, H.; Kinoshita, T.; Hosoya, T.; Furuya, K.; Kinoshita, K.; Tabata, K.; Hemmi, A. Aquastatin A, an inhibitor of mammalian adenosine triphosphatases from Fusarium aquaeductuum. Taxonomy, fermentation, isolation, structure determination and biological properties. J. Antibiot. 1993, 46, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, K.; Yoshida, M.; Izumikawa, M.; Takagi, M.; Shin-Ya, K.; Goshima, N.; Hirokawa, T.; Natsume, T.; Doi, T. Synthesis and biological evaluation of thielocin B1 analogues as protein-protein interaction inhibitors of PAC3 homodimer. Bioorg. Med. Chem. 2018, 26, 6023–6034. [Google Scholar] [CrossRef] [PubMed]
- Folmer, F.; Jaspars, M.; Schumacher, M.; Dicato, M.; Diederich, M. Marine natural products targeting phospholipases A2. Biochem. Pharmacol. 2010, 80, 1793–1800. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Matsutani, S.; Kanda, A.; Kato, T.; Yoshida, T. Thielocin B3, a novel antiinflammatory human group II phospholipase A2 specific inhibitor from ascomycetes. J. Antibiot. 1994, 47, 631–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, M.; Kudo, I.; Suwa, Y.; Inoue, K. Release of 14-kDa group-II phospholipase A2 from activated mast cells and its possible involvement in the regulation of the degranulation process. Eur. J. Biochem. 1992, 209, 257–265. [Google Scholar] [CrossRef]
- Dubin, G.M.; Fkyerat, A.; Tabacchi, R. Acetylenic aromatic compounds from Stereum hirsutum. Phytochemistry 2000, 53, 571–574. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Fungus | Host (Part) | Source, Place | Ref. |
|---|---|---|---|---|
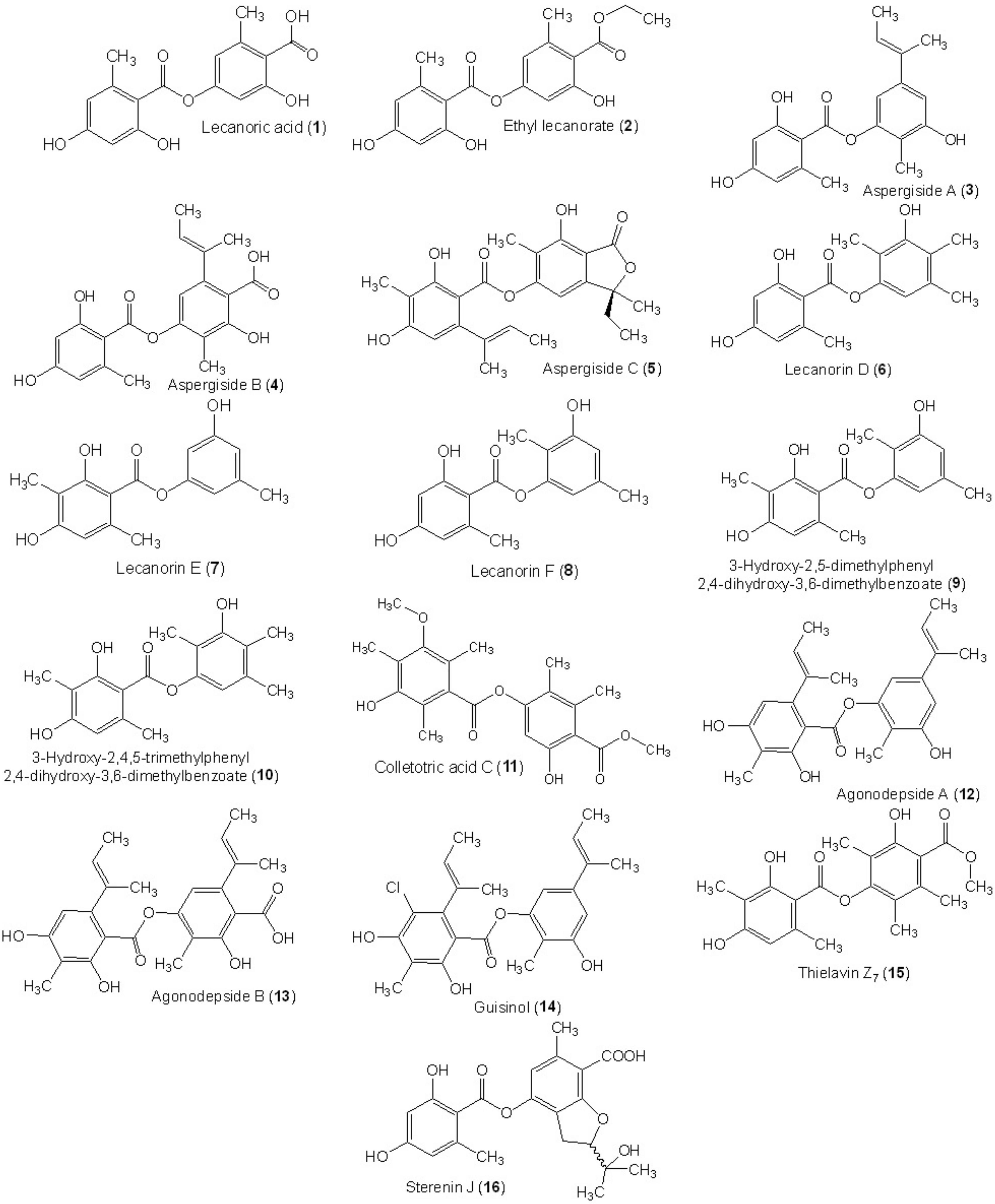
| Lecanoric acid (1) | Claviceps purpurea (PKS7) | - | Culture | [37] |
| Ethyl lecanorate (2) | Claviceps purpurea (PKS7) | - | Culture | [37] |
| Aspergiside A (3) | Aspergillus unguis PSU-RSPG204 (BCC56860) | - | Soil, Surat Thani Province, Thailand | [38] |
| Aspergiside B (4) | Aspergillus unguis PSU-RSPG204 (BCC56860) | - | Soil, Surat Thani Province, Thailand | [38] |
| Aspergiside C (5) | Aspergillus unguis PSU-RSPG204 (BCC56860) | - | Soil, Surat Thani Province, Thailand | [38] |
| Lecanorin D (6) | Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| Lecanorin E (7) | Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| Lecanorin F (8) | Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| 3-Hydroxy-2,5-dimethylphenyl 2,4-dihydroxy-3,6-dimethylbenzoate (9) | Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [49] |
| 3-Hydroxy-2,4,5-trimethylphenyl 2,4-dihydroxy-3,6-dimethylbenzoate (10) | Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [49] |
| Colletotric acid C (11) | Phoma sp. (SYSU-SK-7) | Healthy branch of Kandelia candel | Shankou Mangrove Nature Reserve, Guangxi Province, China | [46] |
| Agonodepside A (12) | Fungal strain (F7524) | Leaves of Derris thyrsiflora | Singapore | [50] |
| Agonodepside B (13) | Fungal strain (F7524) | Leaves of Derris thyrsiflora | Singapore | [50] |
| Guisinol (14) | Emericella unguis (isolate 1 (M87-2) | Stomolopus meleagris | Paria Bay, Venezuela | [51] |
| Emericella unguis (M90B-10) | Soft part of an unidentified mollusc | Paria Bay, Venezuela | [51] | |
| Thielavin Z7 (15) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Sterenin J (16) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
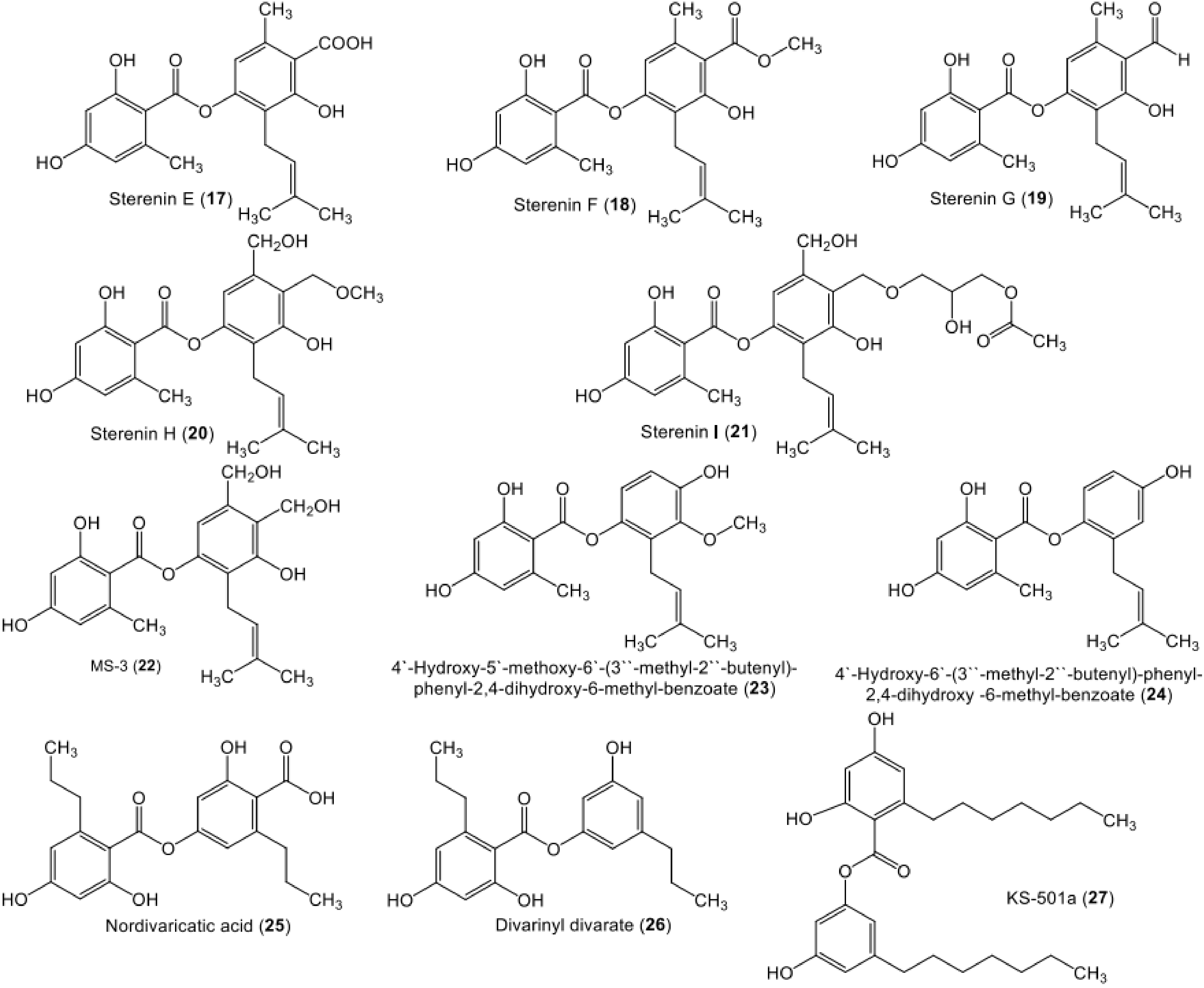
| Sterenin E (17) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| Sterenin F (18) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| Sterenin G (19) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| Sterenin H (20) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| Sterenin I (21) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| MS-3 (22) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [39,53,54,55,56] |
| Stereum rameale (strain 2511) | The bark of a dead tree | Native forest of Nothofagus species (Nothofagaceae), near Ñuble National Reserve, Ñuble Province, Chile | [57,58] | |
| 4′-Hydroxy-5′-methoxy-6′-(3″-methyl-2″-butenyl)-phenyl-2,4-dihydroxy-6-methyl-benzoate/ 4-Hydroxy-3-methoxy-2-(3-methylbut-2-en-1-yl)phenyl 2,4-dihydroxy-6-methylbenzoate (23) | Stereum hirsutum (AB733150.1) | Tibetan Plateau, China | [40] | |
| Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] | |
| 4′-Hydroxy-6′-(3″-methyl-2″-butenyl)-phenyl-2,4-dihydroxy-6-methyl-benzoate (24) | Stereum hirsutum (AB733150.1) | - | Tibetan Plateau, China | [40] |
| Nordivaricatic acid (25) | Fungal strain | - | Soil, Yunnan Province, China | [25] |
| 65-Divarinyl divarate (26) | Fungal strain | - | Soil, Yunnan Province, China | [25] |
| KS-501a (3-Heptyl-5-hydroxyphenyl 2-heptyl-4,6-dihydroxybenzoate) (27) | Acremonium sp. (BCC 14080) | Palm leaf | Khao Yai National Park, Nakhon Ratchasima Province, Thailand | [41,56] |
| KS-501a-2-O-β-D-galactopyranose (28) | Acremonium sp. (BCC 14080) | Palm leaf | Khao Yai National Park, Nakhon Ratchasima Province, Thailand | [41] |
| KS-501a-2-O-β-D-digalactopyranose (29) | Acremonium sp. (BCC 14080) | Palm leaf | Khao Yai National Park, Nakhon Ratchasima Province, Thailand | [41] |
| Arenicolin A (30) | Penicillium arenicola (NRRL 8095) | - | Soil, British Columbia, Canada | [42] |
| Penicillium arenicola (3392) | - | Soil, pine forest sample, near Kiev, Ukraine | [42] | |
| Penicillium arenicola (31507) | - | Mineral soil, under Pinus resinosa, Ontario, Canada | [42] | |
| Penicillium arenicola (31509) | - | Oil-soaked soil sample, Norman Wells, NW Territories, Canada | [42] | |
| Arenicolin B (31) | Penicillium arenicola (NRRL 8095) | - | Soil, British Columbia, Canada | [42] |
| Penicillium arenicola (3392) | - | Soil, pine forest sample, near Kiev, Ukraine | [42] | |
| Penicillium arenicola (31507) | - | Mineral soil, under Pinus resinosa, Ontario, Canada | [42] | |
| Penicillium arenicola (31509) | - | Oil-soaked soil sample, Norman Wells, NW Territories, Canada. | [42] | |
| Aquastatin A (32) | Fusarium aquaeductuum (SANK 11089) | - | Slime fluxes, Karuizawa, Nagano Prefecture, Japan | [59] |
| Cosmospora sp. SF-5060 | - | Inter-tidal sediment, Gejae Island, Korea | [60] | |
| Sporothrix sp. (FN611) | - | Soil, Jeonju City, Jeollabuk-do, Korea | [61] | |
| Depsitinuside (33) | Endophytic fungus, internal strain (8984) | Leaves of Viburnum tinus | - | [62] |
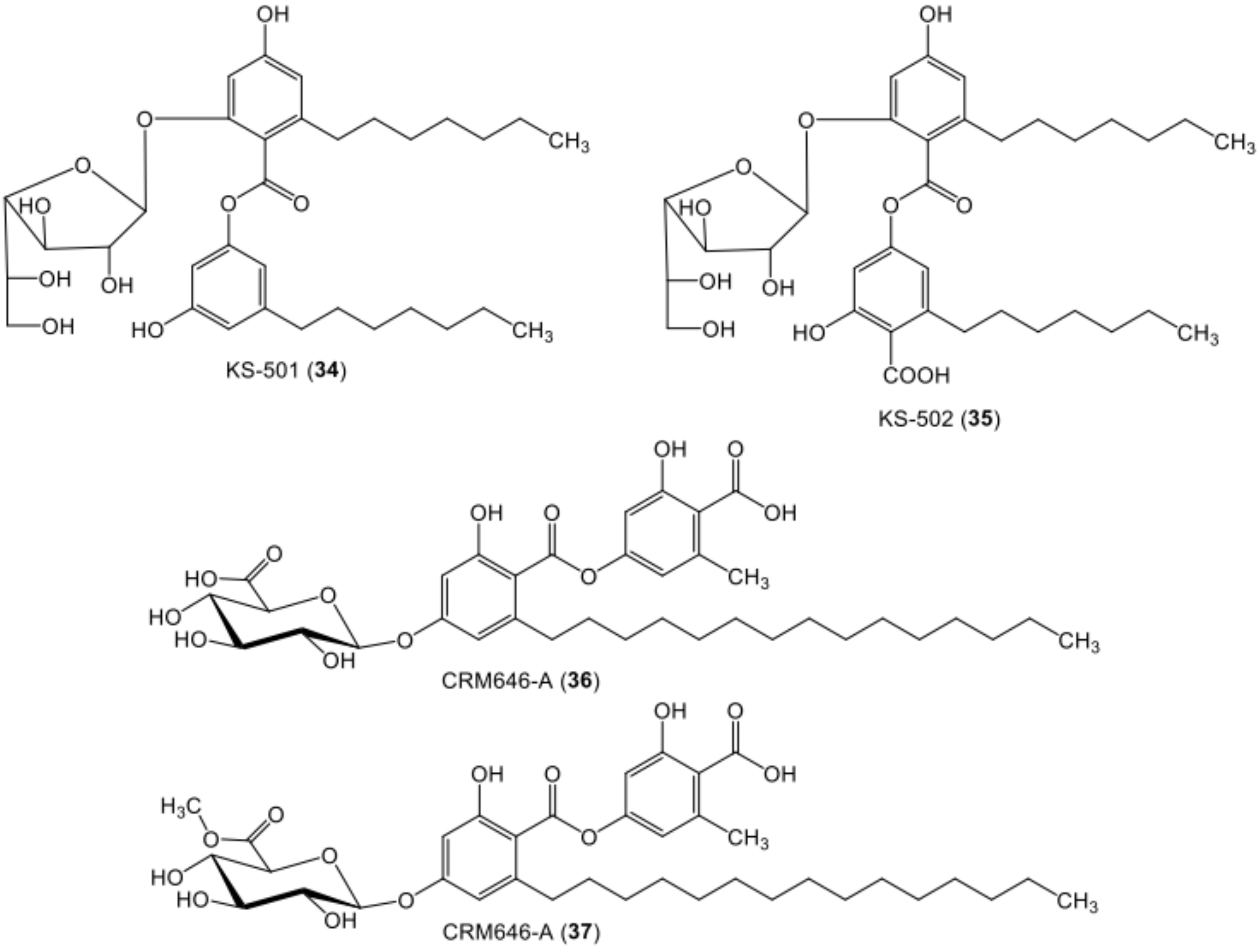
| KS-501 (2-(β-D-galactofuranosyloxy)-6-heptyl-4-hydroxybenzoic acid 3-heptyl-5-hydroxyphenyl ester) (34) | Sporothrix sp. (KAC-1985) | Fallen leaf | Yamakita-cho, Ashgarakami-gun, Kanagawa Prefecture, Japan | [56,63] |
| KS-502 (2-(β-D-galactofuranosyloxy)6-heptyl-4-hydroxybenzoic acid 4-carboxy-3-heptyl-5-hydroxyphenyl ester) (35) | Sporothrix sp. (KAC-1985) | Fallen leaf | Yamakita-cho, Ashgarakami-gun, Kanagawa Prefecture, Japan | [56,63] |
| CRM646-A (36) | Acremonium sp. (MT70646) | - | Soil, Geryong, Kongju, Korea | [43,44,47] |
| CRM646-B (37) | Acremonium sp. (MT70646) | - | Soil, Geryong, Kongju, Korea | [43,44,47] |
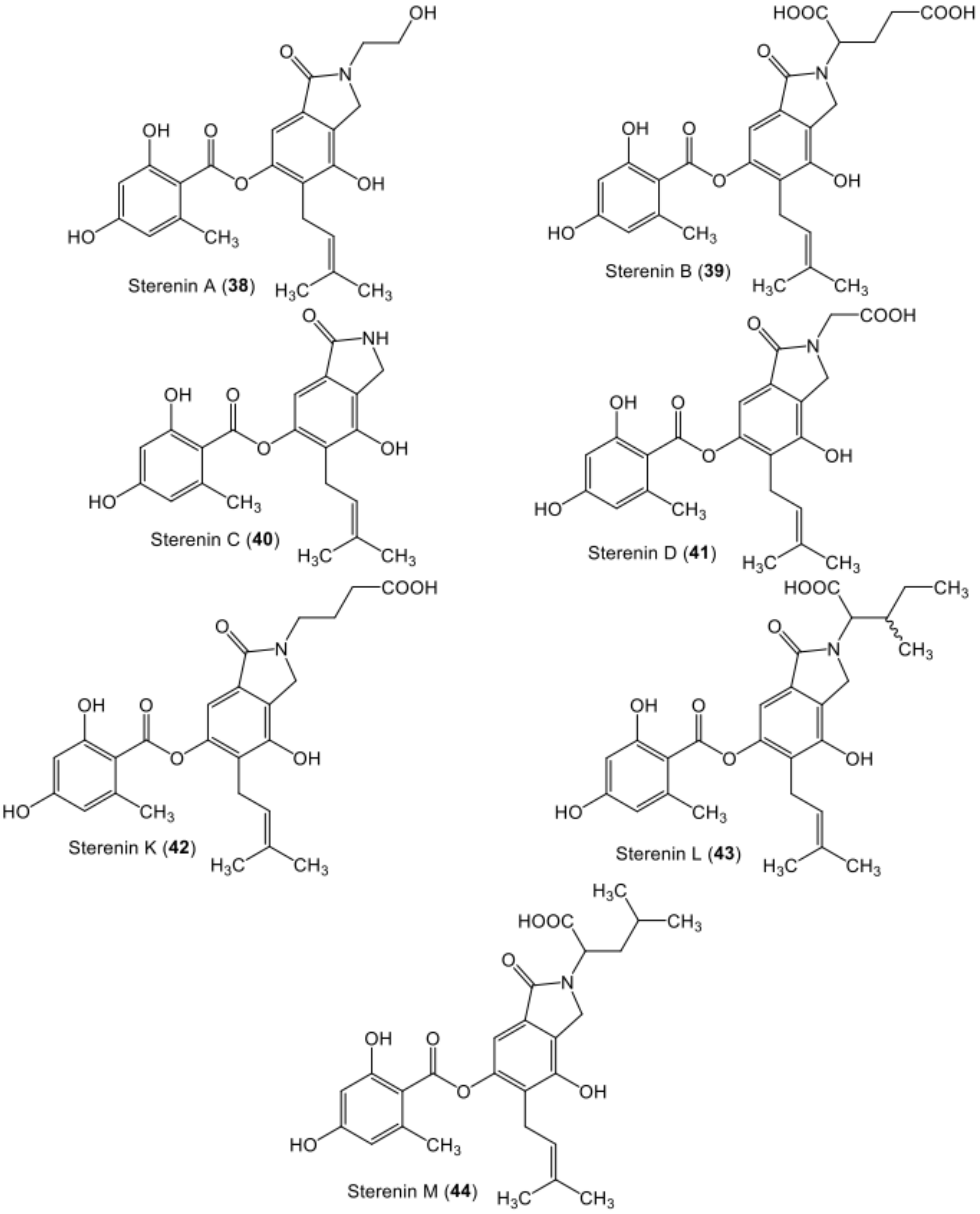
| Sterenin A (38) | Stereum sp. (SANK 21205) | - | Spore print of fresh basidiocarps, Gunma Prefecture, Japan | [64] |
| Sterenin B (39) | Stereum sp. (SANK 21205) | - | Spore print of fresh basidiocarps, Gunma Prefecture, Japan | [64] |
| Sterenin C (40) | Stereum sp. (SANK 21205) | - | Spore print of fresh basidiocarps, Gunma Prefecture, Japan | [64] |
| Sterenin D (41) | Stereum sp. (SANK 21205) | - | Spore print of fresh basidiocarps, Gunma Prefecture, Japan | [64] |
| Sterenin K (42) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| Sterenin L (43) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
| Sterenin M (44) | Stereum hirsutum (EU851110) | - | Tibetan mountain | [53] |
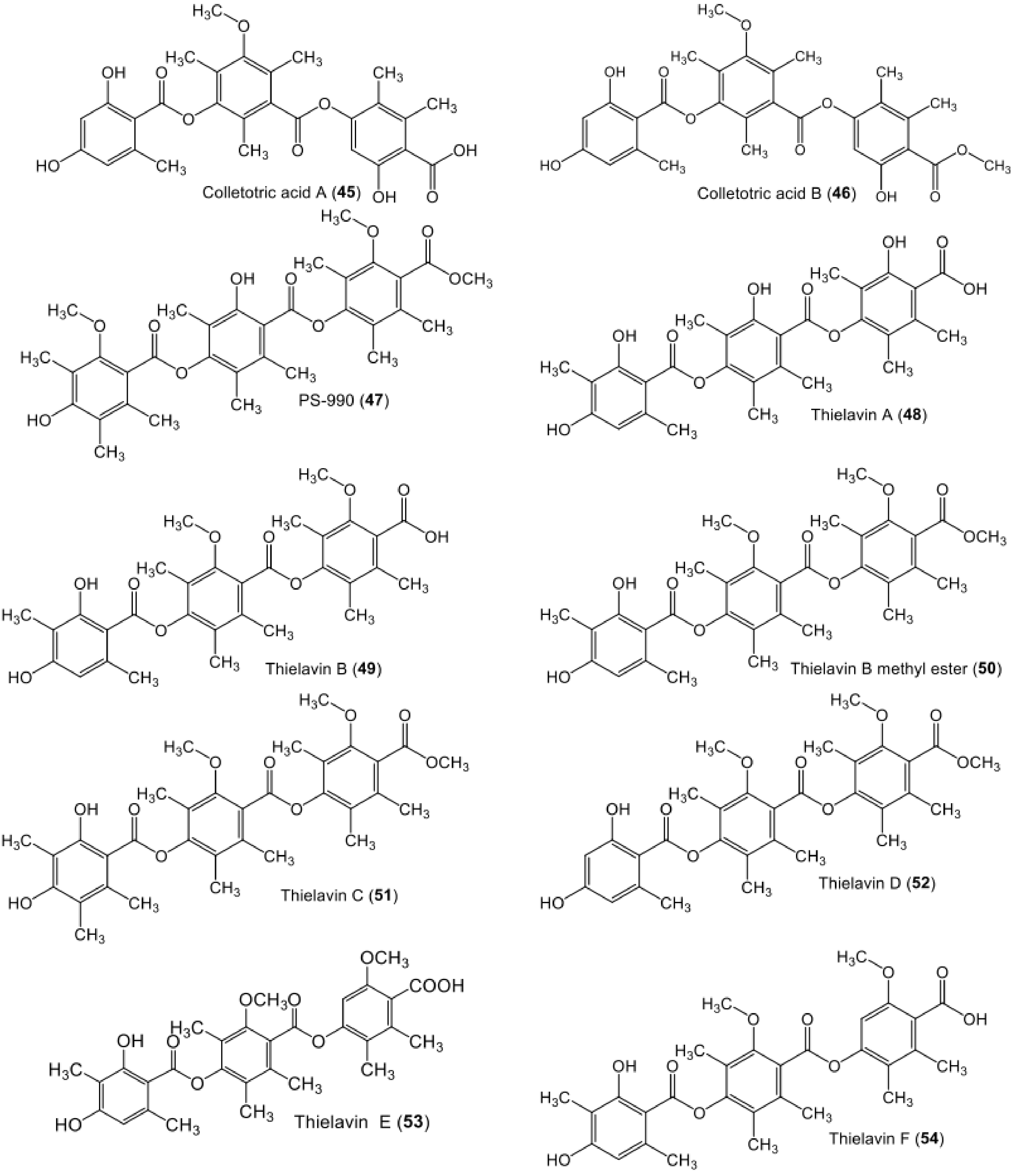
| Colletotric acid A (45) | Phoma sp. (SYSU-SK-7) | Healthy branch of Kandelia candel | Shankou Mangrove Nature Reserve, Guangxi Province, China | [46] |
| Colletotrichum gloeosporioides | Stem of Artemisia mongolica | Hillsides of the Zijin Mountain, suburb of Nanjing, China | [65] | |
| Colletotric acid B (46) | Phoma sp. (SYSU-SK-7) | Healthy branch of Kandelia candel | Shankou Mangrove Nature Reserve, Guangxi Province, China | [46] |
| PS-990 (47) | Acremonium sp. (KY12702) | - | Soil, Tokyo, Japan | [66] |
| Thielavin A (48) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavia sp. (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] | |
| Endophytic fungus MEXU (27905) | Healthy leaves of Hintonia latiflora | México | [67] | |
| Thielavin B (49) | Coniochaeta sp. (10F058a) | - | Soil, Korea | [68] |
| Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] | |
| Coniochaeta sp. (10F058a) | - | Soil, Ochang, Korea. | [68] | |
| Thielavin B methyl ester (50) | Mycosynthetix fungal strain 55526 | - | Leaf litter, North Carolina Smoky Mountains, United States. | [48] |
| Thielavin C (51) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavia terricola (SANK 10475) | - | [69] | ||
| Thielavin D (52) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin E (53) | Thielavia terricola RF-143 | - | [70] | |
| Thielavin F (54) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Coniochaeta sp. (10F058a) | - | Soil, Ochang, Korea | [68] | |
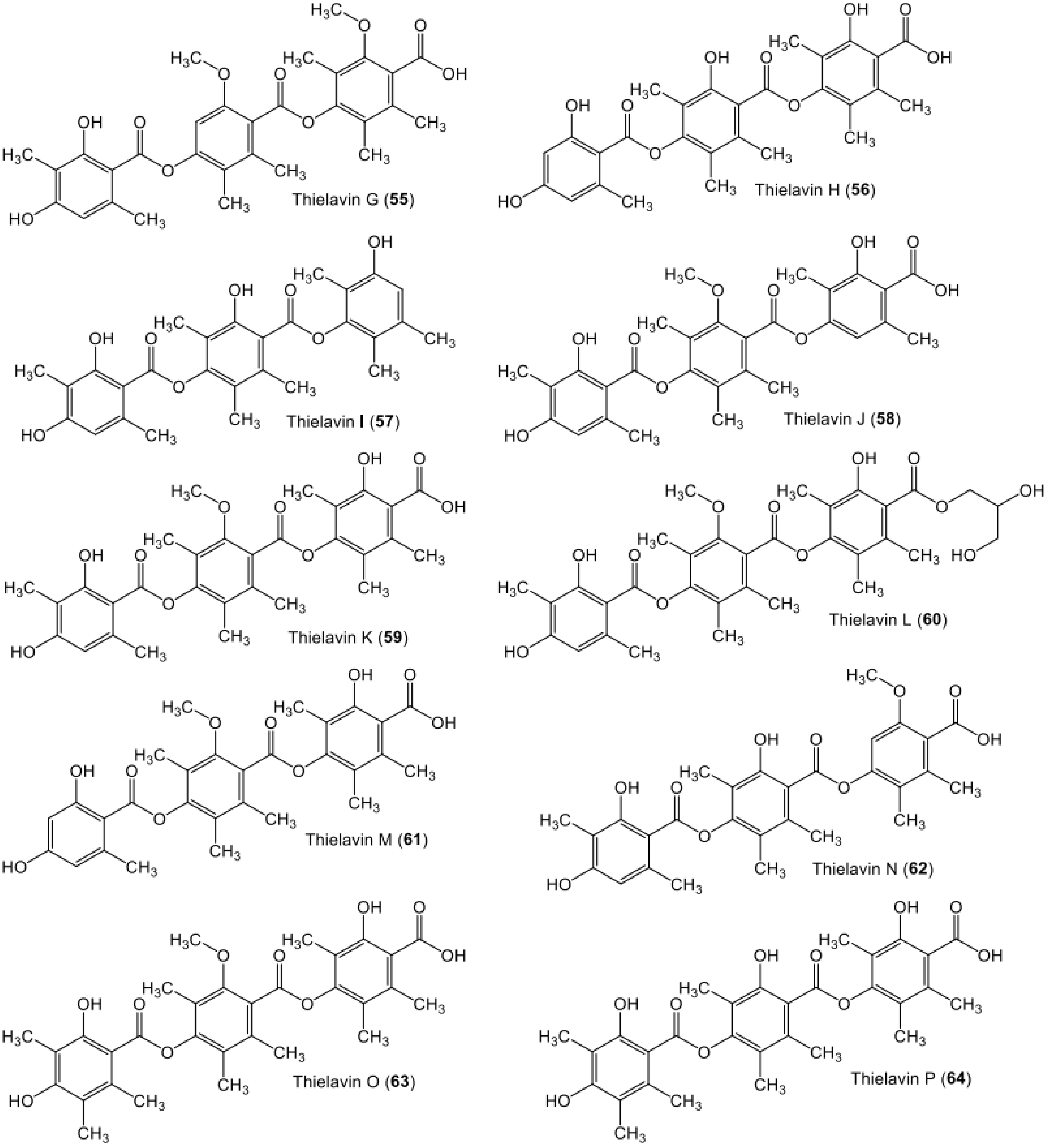
| Thielavin G (55) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin H (56) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavia sp UST030930-004 | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] | |
| Thielavin I (57) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin J (58) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavia sp UST030930-004 | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] | |
| Endophytic fungus MEXU 27905 | Healthy leaves of Hintonia latiflora | México | [67] | |
| Thielavin K (59) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavia sp UST030930-004 | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] | |
| Endophytic fungus MEXU 27905 | Healthy leaves of Hintonia latiflora | México | [67] | |
| Thielavin L (60) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin M (61) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin N (62) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin O (63) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
| Thielavin P (64) | Chaetomium carinthiacum (ATCC 46463) | - | American Type Culture Collection | [62] |
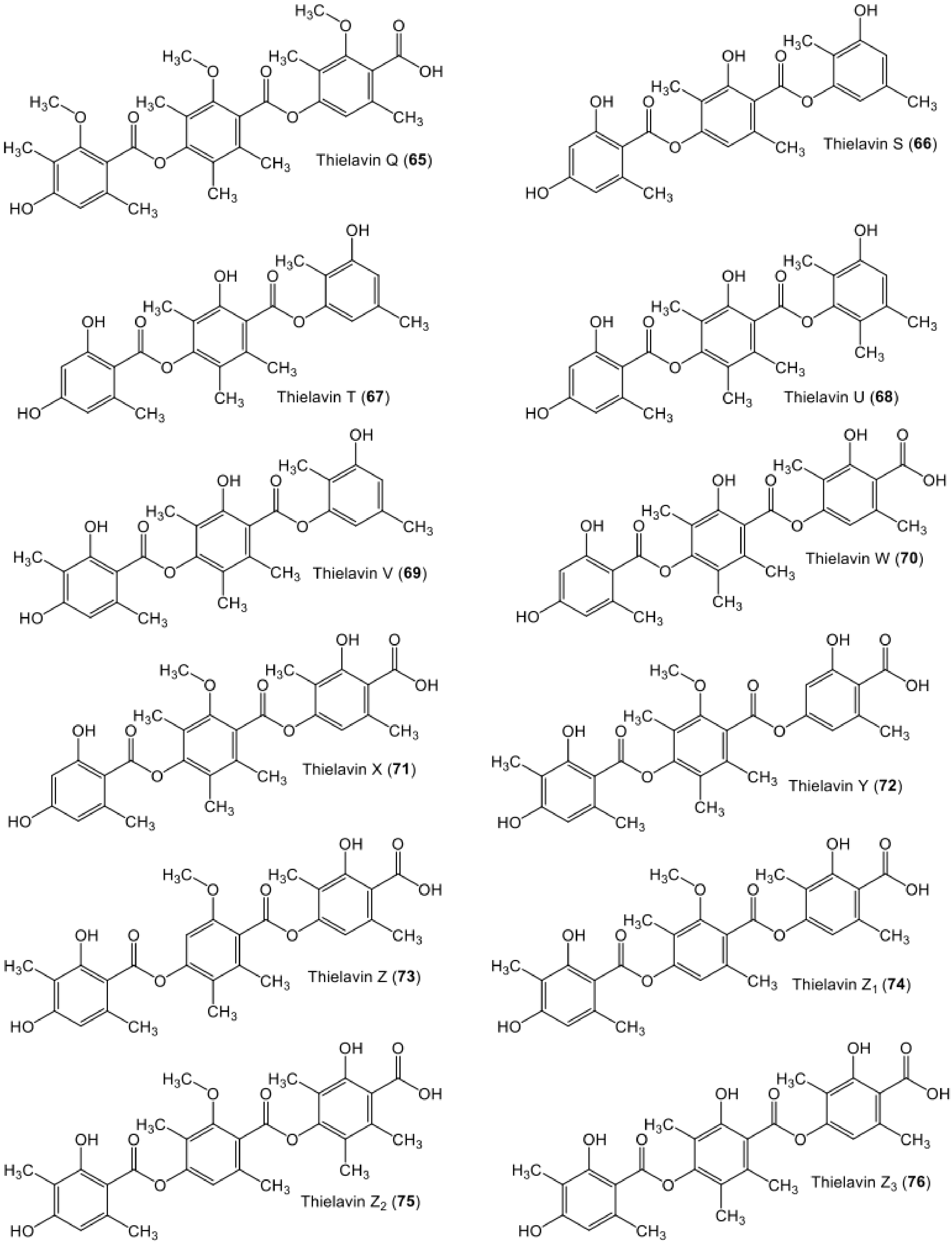
| Thielavin Q (65) | Coniochaeta sp. (10F058a) | - | Soil, Ochang, Korea | [68] |
| Thielavin S (66) | The endophyte Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| Thielavin T (67) | The endophyte Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| Thielavin U (68) | The endophyte Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| Thielavin V (69) | The endophyte Setophoma sp. (KM288713) | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [19] |
| Thielavin W (70 | Thielavia sp UST030930-004 | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin X (71) | Thielavia sp UST030930-004 | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Y (72) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z (73) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z1 (74) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z2 (75) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z3 (76) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z4 (77) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z5 (78) | Thielavia sp (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
| Thielavin Z6 (79) | Thielavia sp. (UST030930-004) | - | 12-d Biofilms developed at the pier of the Hong Kong University of Science and Technology in Port Shelter, Hong Kong, China | [52] |
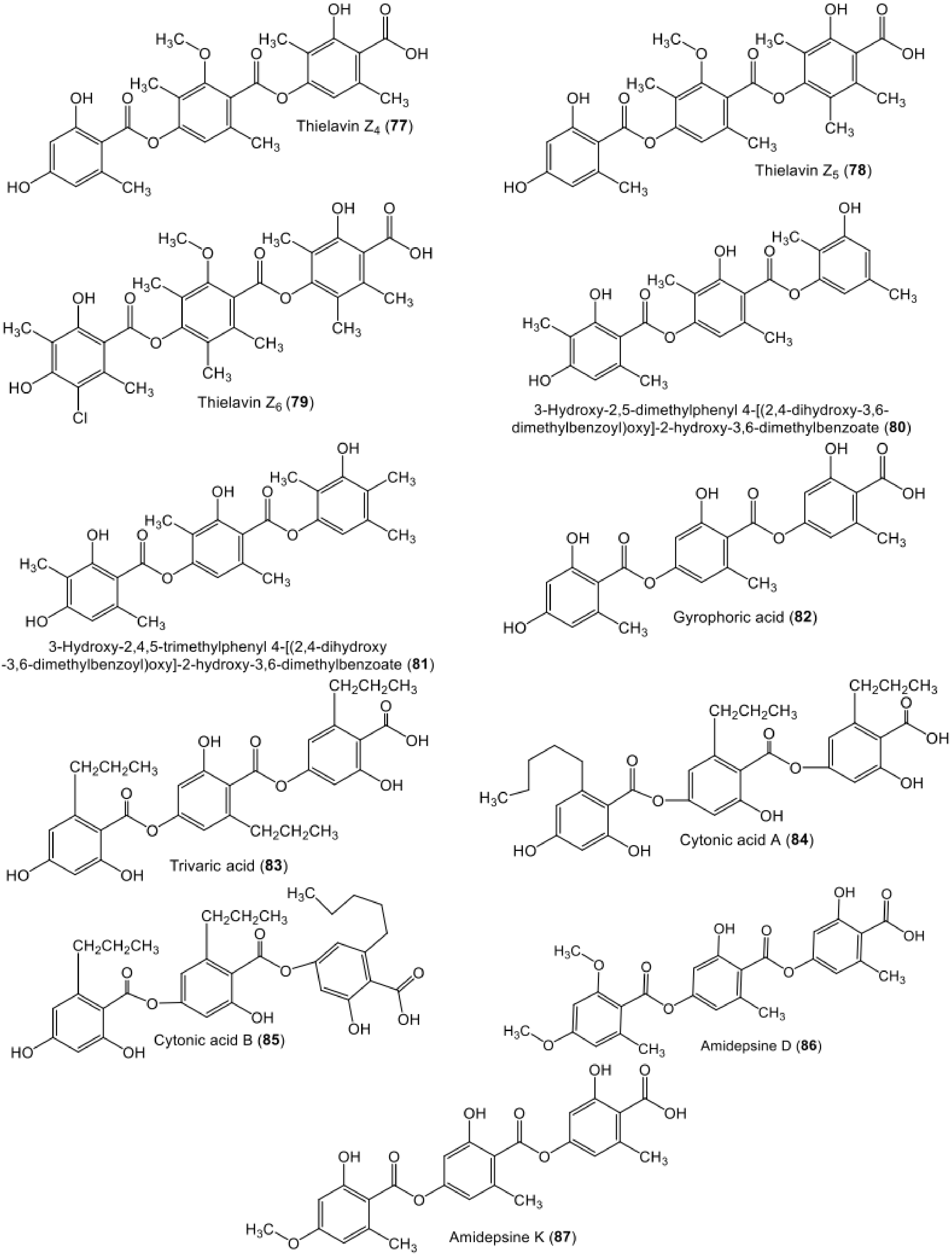
| Hydroxy-2,5-dimethylphenyl 4-[(2,4-dihydroxy-3,6-dimethylbenzoyl)oxy]-2-hydroxy-3,6-dimethylbenzoate (80) | Cladosporium uredinicola | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [49] |
| Hydroxy-2,4,5-trimethylphenyl 4-[(2,4-dihydroxy-3,6-dimethylbenzoyl)oxy]-2-hydroxy-3,6-dimethylbenzoate (81) | Cladosporium uredinicola | Fruits of Psidium guajava | Sӓo Carlos local trade, São Paulo state, Brazil | [49] |
| Gyrophoric acid (82) | Humicola sp. FO-2942 | - | Soil, Nagasaki, Japan | [71,72] |
| Trivaric acid (83) | Fungal strain | - | Soil, Yunnan Province, China | [25] |
| Cytonic acid A (84) | Cytonaema sp. (F32027) | Quercus sp. | United Kingdom | [73] |
| Cytonic acid B (85) | Cytonaema sp. (F32027) | Quercus sp. | United Kingdom | [73] |
| Amidepsine D (86) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [74,75] |
| Amidepsine K (87) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [71] |
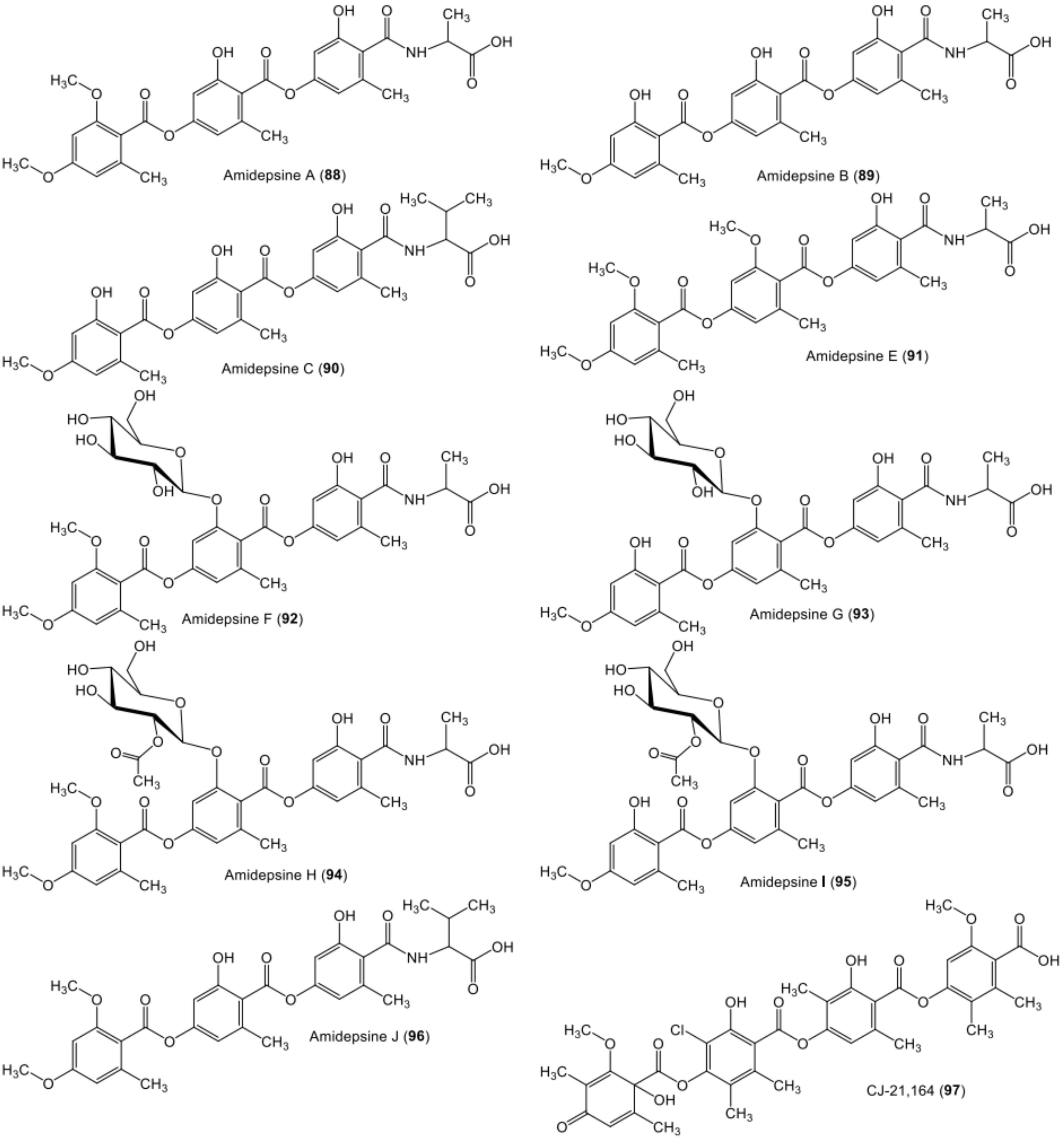
| Amidepsine A (88) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [74,75] |
| Amidepsine B (89) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [74,75] |
| Amidepsine C (90) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [74,75] |
| Amidepsine E (91) | Humicola sp. (FO-5969) | - | Soil, Asaka, Saitama, Japan | [76] |
| Amidepsine F (92) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [71] |
| Amidepsine G (93) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [71] |
| Amidepsine H (94) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [71] |
| Amidepsine I (95) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [71] |
| Amidepsine J (96) | Humicola sp. (FO-2942) | - | Soil, Nagasaki, Japan | [71] |
| 44-CJ-21,164 (97) | Chloridium sp. (CL48903). | - | China | [77] |
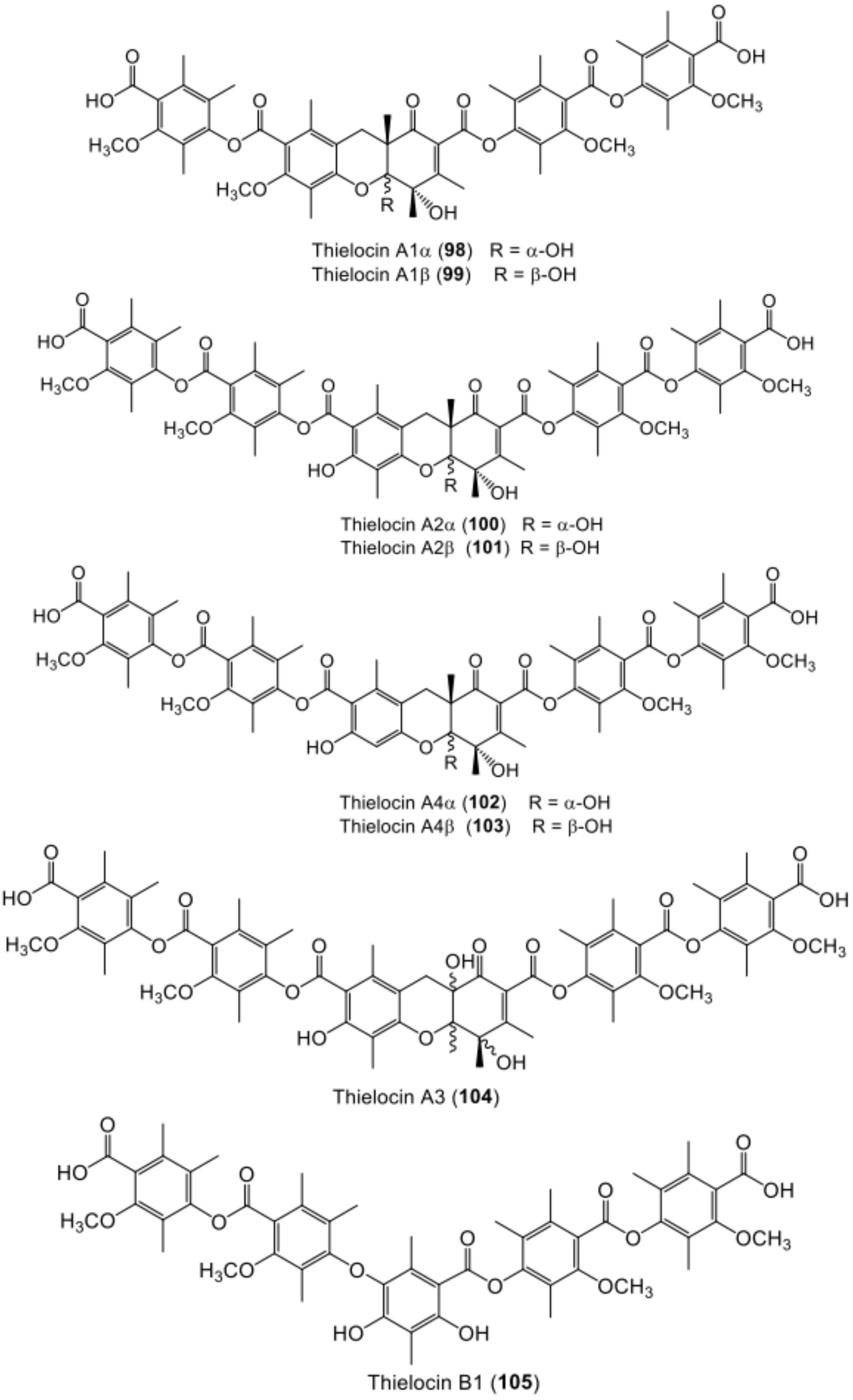
| Thielocin A1α (98) | Thielavia terricola (RF-143) | - | Culture | [78] |
| Thielocin A1β (99) | Thielavia terricola (RF-143) | - | Culture | [78] |
| Thielocin A2α (100) | Thielavia terricola (RF-143) | - | Culture | [70] |
| Thielocin A2β (101) | Thielavia terricola (RF-143) | - | Culture | [70] |
| Thielocin A4α (102) | Thielavia terricola (RF-143) | - | Culture | [79] |
| Thielocin A4β (103) | Thielavia terricola (RF-143) | - | Culture | [79] |
| Thielocin A3 (104) | Thielavia terricola (RF-143) | - | Culture | [70] |
| Thielocin B1 (105) | Thielavia terricola (RF-143) | - | Culture | [70] |
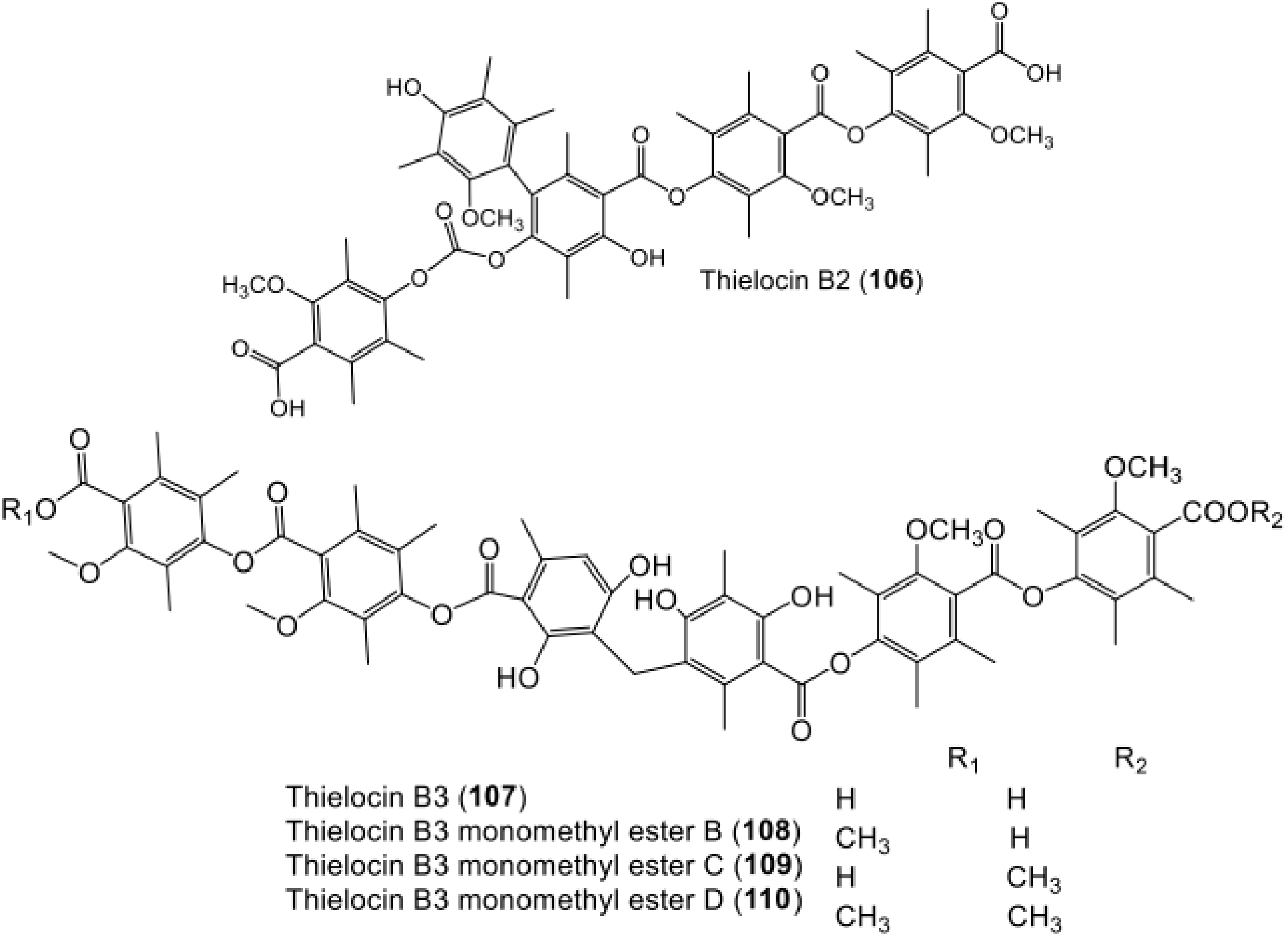
| Thielocin B2 (106) | Thielavia terricola (RF-143) | - | Culture | [70] |
| Thielocin B3 (107) | Thielavia terricola (RF-143) | - | Culture | [80] |
| Thielocin B3 monomethyl ester B (108) | Thielavia terricola (RF-143) | - | Culture | [80] |
| Thielocin B3 monomethyl ester C (109) | Thielavia terricola (RF-143) | - | Culture | [80] |
| Thielocin B3 monomethyl ester D (110) | Thielavia terricola (RF-143) | - | Culture | [80] |
| Compound Name | Activity | Assay/Microorganism/Model/Enzyme | Results | Positive Control | Ref. |
|---|---|---|---|---|---|
| Lecanoric acid (1) | Antitumor | Colorimetric CTC/HepG2 | 40.0 µM (IC50) | T-2 toxin 10.0 µM (IC50) | [37] |
| Ethyl lecanorate (2) | Antitumor | Colorimetric CTC/HepG2 | 40.0 µM (IC50) | T-2 toxin 10.0 µM (IC50) | [37] |
| Antitumor | Colorimetric CTC/CCF-STTG1 | 54.0 µM (IC50) | T-2 toxin 10.0 µM (IC50) | [37] | |
| Aspergiside A (3) | Antibacterial | Agar diffusion assay/S. aureus ATCC25923 | 8 µg/mL (MIC) | Vancomycin 0.25 µg/mL (MIC) | [38] |
| Antibacterial | Agar diffusion assay/MRSA | 8 µg/mL (MIC) | Vancomycin 0.50 µg/mL (MIC) | [38] | |
| Antifungal | Agar diffusion assay/C. albicans NCPF3153 | 200 µg/mL (MIC) | Amphotericin B 0.25 µg/mL (MIC) | [38] | |
| Antibacterial | Agar diffusion assay/C. neoformans ATCC 9013 | 64 µg/mL (MIC) | Amphotericin B 0.25 µg/mL (MIC) | [38] | |
| Antifungal | Agar diffusion assay/M. gypseum SH-MU-4 | 128 µg/mL (MIC) | Miconazole 1.0 µg/mL (MIC) | [38] | |
| Antitumor | MTT/KB | 53.69 µM (IC50) | Ellipticine 10.56 µM (IC50) Doxorubicin 0.65 µM (IC50) | [38] | |
| Antitumor | MTT/MCF-7 | 54.42 µM (IC50) | Doxorubicin 16.83 µM (IC50) Tamoxifen 20.78 µM (IC50) | [38] | |
| Antitumor | MTT/Vero | 83.75 µM (IC50) | Ellipticine 4.55 µM (IC50) | [38] | |
| Aspergiside B (4) | Antibacterial | Agar diffusion assay/S. aureus ATCC25923 | 128 µg/mL (MIC) | Vancomycin 0.25 µg/mL (MIC) | [38] |
| Antibacterial | Agar diffusion assay/MRSA | 128 µg/mL (MIC) | Vancomycin 0.50 µg/mL (MIC) | [38] | |
| Antibacterial | Agar diffusion assay/C. neoformans ATCC 9013 | 32 µg/mL (MIC) | Amphotericin B 0.25 µg/mL (MIC) | [38] | |
| Antitumor | MTT/Vero cells | 114.40 µM (IC50) | Ellipticine 4.55 µM (IC50) | [38] | |
| Aspergiside C (5) | Antibacterial | Agar diffusion assay/S. aureus ATCC25923 | 200 µg/mL (MIC) | Vancomycin 0.25 µg/mL (MIC) | [38] |
| Antibacterial | Agar diffusion assay/MRSA | 200 µg/mL (MIC) | Vancomycin 0.50 µg/mL (MIC) | [38] | |
| Antibacterial | Agar diffusion assay/C. neoformans ATCC 9013 | 200 µg/mL (MIC) | Amphotericin B 0.25 µg/mL (MIC) | [38] | |
| Antitumor | MTT/Vero cells | 45.07 µM (IC50) | Ellipticine 4.55 µM (IC50) | [38] | |
| 3-Hydroxy-2,5-dimethylphenyl 2,4-dihydroxy-3,6-dimethylbenzoate (9) | Antibacterial | Microbroth dilution/S. aureus | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] |
| Antibacterial | Microbroth dilution/B. subtilis | 25 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Antibacterial | Microbroth dilution/P. aeruginosa | 25 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Colletotric acid C (11) | Antibacterial | Agar diffusion assay/B. subtilis | 9.70 µg/mL (MIC) | Ampicillin 0.07 µg/mL (MIC) | [46] |
| Agonodepside A (12) | Enoyl-ACP reductase inhibition | Fluorometric InhA assay/M. tuberculosis InhA | 74 µM (IC50) | Triclosan 3 µM (IC50) | [50] |
| Thielavin Z7 (15) | Antifouling | Balanus Amphitrite(cyprid larvae) | 3.20 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Sterenin J (16) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 65.70 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin E (17) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 7.62 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin F (18) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 3.06 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin G (19) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 6.03 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin H (20) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 22.70 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin I (21) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 72.50 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| MS-3 (22) | Antibacterial | Plate diffusion/B. cereus | 25 mm (IZD) | Streptomycin 28 mm (IZD) Penicillin G 28 mm (IZD) | [57] |
| Antibacterial | Plate diffusion/B. subtilis | 25 mm (IZD) | Streptomycin 32 mm (IZD) Penicillin G 28 mm (IZD) | [57] | |
| Antibacterial | Plate diffusion/S. aureus | 28 mm (IZD) | Streptomycin 30 mm (IZD) Penicillin G 27 mm (IZD) | [57] | |
| α-Glucosidase inhibitory | Colorimetric/α-Glucosidase inhibitor | 23.82 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] | |
| 4′-Hydroxy-5′-methoxy-6′-(3″-methyl-2″-butenyl)-phenyl-2,4-dihydroxy-6-methyl-benzoate/ 4-Hydroxy-3-methoxy-2-(3-methylbut-2-en-1-yl)phenyl 2,4-dihydroxy-6-methylbenzoate (23) | Anti-inflammatory NO inhibitory potential | LPS-induced macrophages RAW 264.7 | 19.17 µM (IC50) | Hydrocortisone 48.15 µM (IC50) | [40] |
| Antibacterial | Agar plate diffusion/MRSA | 25 µg/mL (MIC) | Ancomycin 1.0 µg/mL (MIC) | [40] | |
| Antibacterial | Agar plate diffusion/S. aureus | 25 µg/mL (MIC) | Ancomycin 1.0 µg/mL (MIC) | [40] | |
| Antibacterial | Agar plate diffusion/B. subtilis | 25 µg/mL (MIC) | Ancomycin 0.5 µg/mL (MIC) | [40] | |
| Antitumor | MTT/A549 | 13.14 µM (IC50) | Cisplatin 14.33 µM (IC50) | [40] | |
| Antitumor | MTT/HepG2 | 49.02 µM (IC50) | Cisplatin 18.74 µM (IC50) | [40] | |
| α-Glucosidase inhibitory | Colorimetric/α-Glucosidase inhibitor | 14.17 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] | |
| 4′-Hydroxy-6′-(3″-methyl-2″-butenyl)-phenyl-2,4-dihydroxy-6-methyl-benzoate (24) | Antibacterial | Agar plate diffusion/MRSA | 25 µg/mL (MIC) | Ancomycin 1.0 µg/mL (MIC) | [40] |
| Antibacterial | Agar plate diffusion/S. aureus | 25 µg/mL (MIC) | Ancomycin 1.0 µg/mL (MIC) | [40] | |
| Antibacterial | Agar plate diffusion/B. subtilis | 50 µg/mL (MIC) | Ancomycin 0.5 µg/mL (MIC) | [40] | |
| KS-501a (27) | Antimalarial activity | Microculture radioisotope technique/Plasmodium falciparum K1 | 9.9 µM (IC50) | Dihydroartemisinin 0.0039 µM (IC50) | [41] |
| Antitumor | MTT/KB | 13.0 µM (IC50) | Ellipticine 1.99 µM (IC50) | [41] | |
| Antitumor | MTT/BC | 8.8 µM (IC50) | Ellipticine 0.49 µM (IC50) | [41] | |
| Antitumor | MTT/NC1-H187 | 13.6 µM (IC50) | Ellipticine 1.77 µM (IC50) | [41] | |
| Antitumor | MTT/Vero cells | 34.3 µM (IC50) | Ellipticine 1.94 µM (IC50) | [41] | |
| KS-501a-2-O-β-D- galactopyranose (28) | Antitumor | MTT/KB | >25 µM (IC50) | Ellipticine 1.99 µM (IC50) | [41] |
| Antitumor | MTT/BC | 4.4 µM (IC50) | Ellipticine 0.49 µM (IC50) | [41] | |
| Antitumor | MTT/NC1-H187 | 13.9 µM (IC50) | Ellipticine 1.77 µM (IC50) | [41] | |
| KS-501a-2-O-β-D- digalactopyranose (29) | Antitumor | MTT/KB | >25 µM (IC50) | Ellipticine 1.99 µM (IC50) | [41] |
| Antitumor | MTT/BC | 20.2 µM (IC50) | Ellipticine 0.49 µM (IC50) | [41] | |
| Antitumor | MTT/NC1-H187 | >25 µM (IC50) | Ellipticine 1.77 µM (IC50) | [41] | |
| Antitumor | MTT/Vero cells | 32.1 µM (IC50) | Ellipticine 1.94 µM (IC50) | [41] | |
| Arenicolin A (30) | Antitumor | Immunocytochemistry ICC/HCT-116 | 7.3 µM (IC50) | 5-FU 6.5 µM (IC50) | [42] |
| Antitumor | Immunocytochemistry ICC/IMR-32 | 6.0 µM (IC50) | 5-FU 5.7 µM (IC50) | [42] | |
| Antitumor | Immunocytochemistry ICC/BT-474 | 9.7 µM (IC50) | - | [42] | |
| Sterenin A (38) | 11β-HSD inhibitory | Luminescence immunoassay/HTRF cortisol | 240 nM (IC50) | - | [64] |
| α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 25.10 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] | |
| Sterenin B (39) | 11β-HSD inhibitory | Luminescence immunoassay/HTRF cortisol | 6600 nM (IC50) | - | [64] |
| α-Glucosidase inhibitory | Colorimetric/α-Glucosidase inhibitor | 12.23 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] | |
| Sterenin C (40) | 11β-HSD inhibitory | Luminescence immunoassay/HTRF cortisol | 230 nM (IC50) | - | [64] |
| α-Glucosidase inhibitory | Colorimetric/α-Glucosidase inhibitor | 3.31 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] | |
| Sterenin D (41) | 11β-HSD inhibitory | Luminescence immunoassay/HTRF cortisol | 2600 nM (IC50) | - | [64] |
| Antifungal | Plate diffusion/B. cinerea | 50 µg/mL (MFC) | Rovral 10 µg/mL (MFC) | [58] | |
| Antifungal | Plate diffusion/B. cinerea | 10 µg/mL (MIC) | Rovral 1 µg/mL (MFC) | [58] | |
| Sterenin K (42) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 36.64 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin L (43) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 13.09 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Sterenin M (44) | α-Glucosidase inhibitory | Colorimetric/α-Glucosidase | 27.52 µM (IC50) | Acarbose 640.88 µM (IC50) | [53] |
| Colletotric acid A (45) | Antibacterial | Agar diffusion assay/B. subtilis | 25 µg/mL (MIC) | Ampicillin 0.05 µg/mL (MIC) | [65] |
| Antibacterial | Agar diffusion assay/B. subtilis | 6.55 µg/mL (MIC) | Ampicillin 0.07 µg/mL (MIC) | [46] | |
| Antibacterial | Agar diffusion assay/S. aureus | 50 µg/mL (MIC) | Ampicillin 0.5 µg/mL (MIC) | [65] | |
| Antibacterial | Agar diffusion assay/S. lutea | 50 µg/mL (MIC) | Ampicillin 0.01 µg/mL (MIC) | [65] | |
| Antibacterial | Agar diffusion assay/H. lutea | 50 µg/mL (MIC) | Triadimefon 20 µg/mL (MIC) | [65] | |
| Antibacterial | Agar diffusion assay/P. aeruginosa | 3.27 µg/mL (MIC) | Ampicillin 0.15 µg/mL (MIC) | [46] | |
| Antibacterial | Agar diffusion assay/MRSA | 6.28 µg/mL (MIC) | Ampicillin 0.15 µg/mL (MIC) | [46] | |
| Antibacterial | Agar diffusion assay/S. typhimurium | 26.20 µg/mL (MIC) | Ampicillin 0.31 µg/mL (MIC) | [46] | |
| Antifungal | Agar diffusion assay/C. albicans | 3.27 µg/mL (MIC) | Ketoconazole 0.10 µg/mL (MIC) | [46] | |
| Antitumor | MTT/MDA-MB-435 | 37.01 µM (IC50) | Epirubicin 0.26 µM (IC50) | [46] | |
| Antitumor | MTT/A549 | 37.73 µM (IC50) | Epirubicin 5.60 µM (IC50) | [46] | |
| Colletotric acid B (46) | Antibacterial | Agar diffusion assay/P. aeruginosa | 1.67 µg/mL (MIC) | Ampicillin 0.15 µg/mL (MIC) | [46] |
| Antibacterial | Agar diffusion assay/MRSA | 3.36 µg/mL (MIC) | Ampicillin 0.15 µg/mL (MIC) | [46] | |
| Antitumor | MTT/MDA-MB-435 | 16.82 µM (IC50) | Epirubicin 0.26 µM (IC50) | [46] | |
| Antitumor | MTT/A549 | 20.75 µM (IC50) | Epirubicin 5.60 µM (IC50) | [46] | |
| Antibacterial | Agar diffusion assay/B. subtilis | 26.90 µg/mL (MIC) | Ampicillin 0.07 µg/mL (MIC) | [46] | |
| Thielavin A (48) | Phospholipase A (PLA) inhibition | Rat PLA2-II | 43 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 29 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Antifouling | Balanus amphitrite (cyprid larvae) | 54.99 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] | |
| Anti-diabetic | Colorimetric/α-Glucosidase inhibitor (αGHY) | 23.8 µM (IC50) | Acarbose 545 µM (IC50 | [67] | |
| Prostaglandin synthesis inhibition | Conversion of AA into PGH2 | 10 µM (IC50) | Indomethacin 30 µM (IC50) | [69,81] | |
| Conversion of PGH2 into PGE2 | 40 µM (IC50) | Indomethacin 130 µM (IC50) | |||
| Conversion of PGE2 into TXA2 | 150 µM (IC50) | Imidazole 200 µM (IC50) | |||
| Thielavin B (49) | Phospolipase A (PLA) inhibition | Rat PLA2-II | 1.3 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 2.4 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Prostaglandin synthesis inhibition | Conversion of AA into PGH2 | 40 µM (IC50) | Indomethacin 30 µM (IC50) | [69,81] | |
| Conversion of PGH2 into PGE2 | 9 µM (IC50) | Indomethacin 130 µM (IC50) | |||
| Conversion of PGE2 into TXA2 | 350 µM (IC50) | Imidazole 200 µM (IC50) | |||
| Thielavin B methyl ester (50) | Antitumor | SRB/MCF-7 | 7.3 µM (IC50) | 0.07 µM (IC50) | [48] |
| Antitumor | SRB/H460 | 6.6 µM (IC50) | < 0.01 µM (IC50) | [48] | |
| Antitumor | SRB/SF268 | 8.1 µM (IC50) | 0.04 µM (IC50) | [48] | |
| Thielavin C (51) | PLA inhibition | Rat PLA2-II | 0.46 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 2.1 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielavin D (52) | PLA inhibition | Rat PLA2-II | 1.1 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 6.2 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielavin E (53) | PLA inhibition | Rat PLA2-II | 4.5 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 9.3 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielavin H (56) | Antifouling | Balanus amphitrite | 12.64 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin J (58) | Anti-diabetic | Colorimetric/α-Glucosidase inhibitor (αGHY) | 15.8 µM (IC50) | Acarbose 545 µM (IC50) | [67] |
| Thielavin K (59) | Anti-diabetic | Colorimetric/α-Glucosidase inhibitor (αGHY) | 22.1 µM (IC50) | Acarbose 545 µM (IC50) | [67] |
| Thielavin S (66) | Antibacterial | Microbroth dilution/S. aureus ATCC 25923 | 100 µg/mL (MIC) | Tetracycline 3.12 µg/mL (MIC) | [19] |
| Thielavin T (67) | Antimicrobial | Microbroth dilution assay/S. aureus ATCC 25923 | 6.25 µg/mL (MIC) | Tetracycline 3.12 µg/mL (MIC) | [19] |
| Thielavin U (68) | Antibacterial | Microbroth dilution assay/S. aureus ATCC 25923 | 50 µg/mL (MIC) | Tetracycline 3.12 µg/mL (MIC) | [19] |
| Thielavin V (69) | Antibacterial | Microbroth dilution assay/S. aureus ATCC 25923 | 25 µg/mL (MIC) | Tetracycline 3.12 µg/mL (MIC) | [19] |
| Thielavin W (70) | Antifouling | Balanus Amphitrite(cyprid larvae) | 2.95 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin X (71) | Antifouling | Balanus Amphitrite(cyprid larvae) | 3.13 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin Y (72) | Antifouling | Balanus Amphitrite(cyprid larvae) | 5.78 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin Z2 (75) | Antifouling | Balanus Amphitrite(cyprid larvae) | 69.19 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin Z3 (76) | Antifouling | Balanus Amphitrite (cyprid larvae) | 4.23 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin Z4 (77) | Antifouling | Balanus Amphitrite(cyprid larvae) | 50.50 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin Z5 (78) | Antifouling | Balanus Amphitrite(cyprid larvae) | 25.86 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| Thielavin Z6 (79) | Antifouling | Balanus Amphitrite(cyprid larvae) | 17.86 µM (EC50) | Butenolide 4.62 µM (EC50) | [52] |
| 3-Hydroxy-2,5-dimethylphenyl 4-[(2,4-dihydroxy-3,6-dimethylbenzoyl)oxy]-2-hydroxy-3,6- dimethylbenzoate (80) | Antibacterial | Microbroth dilution/S. aureus | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] |
| Antibacterial | Microbroth dilution/B. subtilis | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Antibacterial | Microbroth dilution/P. aeruginosa | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Antibacterial | Microbroth dilution/E. coli | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| 3-Hydroxy-2,4,5-trimethylphenyl 4-[(2,4-dihydroxy-3,6-dimethylbenzoyl)oxy]-2-hydroxy-3,6- dimethylbenzoate (81) | Antibacterial | Microbroth dilution/S. aureus | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] |
| Antibacterial | Microbroth dilution/B. subtilis | 25 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Antibacterial | Microbroth dilution/P. aeruginosa | 25 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Antibacterial | Microbroth dilution/E. coli | 250 µg/mL (MIC) | Vancomycin and tetracycline | [49] | |
| Amidepsine D (86) | DGAT1 inhibition | Rat liver microsomes | 17.5 µM (IC50) | - | [74] |
| DGAT2 inhibition | Rat liver microsomes | 30 µM (IC50) | [71,82] | ||
| Triacylglycerol inhibition | Raji cells | 2.82 µM (IC50) | - | [74] | |
| Antibacterial | Disk diffusion/B. subtilis ATCC6633 | 8.0 mm (IZD) | - | [74] | |
| Amidepsine A (88) | DGAT1 inhibition | Rat liver microsomes | 10.2 µM (IC50) | - | [74] |
| DGAT2 inhibition | Rat liver microsomes | 70 µM (IC50) | - | [71,82] | |
| Triacylglycerol inhibition | Raji cells | 15.5 µM (IC50) | - | [74] | |
| Antibacterial | Disk diffusion/B. subtilis ATCC6633 | 11.0 mm (IZD) | - | [74] | |
| Amidepsine B (89) | DGAT1 inhibition | Rat liver microsomes | 19.2 µM (IC50) | - | [74] |
| DGAT2 inhibition | Rat liver microsomes | 60 µM (IC50) | - | [71,82] | |
| Triacylglycerol inhibition | Raji cells | 3.35 µM (IC50) | - | [74] | |
| Antibacterial | Disk diffusion/B. subtilis ATCC6633 | 7.0 mm (IZD) | - | [74] | |
| Amidepsine C (90) | DGAT1 inhibition | Rat liver microsomes | 51.6 µM (IC50) | - | [74] |
| DGAT2 inhibition | Rat liver microsomes | 100 µM (IC50) | - | [71,82] | |
| Triacylglycerol inhibition | Raji cells | 17.2 µM (IC50) | - | [74] | |
| Antibacterial | Disk diffusion/B. subtilis ATCC6633 | 9.0 mm (IZD) | - | [74] | |
| Amidepsine E (91) | Triacylglycerol inhibition | Raji cells | 91 µM (IC50) | - | [76] |
| DGAT1 inhibition | Rat liver microsomes | 124 µM (IC50) | - | [76] | |
| Amidepsine J (96) | DGAT1 inhibition | Rat liver microsomes | 40 µM (IC50) | - | [71] |
| DGAT2 inhibition | Rat liver microsomes | 40 µM (IC50) | [71] | ||
| CJ-21,164 (97) | G6Pase inhibition | Colorimetric assay | 102% (% inhibition) | [77] | |
| Glucose output inhibition | Colorimetric assay | 81% (% inhibition) | - | [77] | |
| Thielocin A1i (99) | PLA inhibition | Human PLA2-II | 6.2 µM (IC50) | - | [78,80,83,84] |
| Human PLA2-I | 140 µM (IC50) | - | |||
| Ki value human PLA2-II | 12 µM | - | |||
| Rat PLA2-II | 0.0033 µM (IC50) | Mepacrine 240 µM (IC50) | |||
| Vipera russelli venom PLA2-II | 17 µM (IC50) | ||||
| Crotalus adamanteus venom PLA2-II | 17 µM (IC50) | ||||
| Porcine pancreas PLA2-I | 63 µM (IC50) | ||||
| Rat PLA2-I | 21 µM (IC50) | Mepacrine 135 µM (IC50) | |||
| Bee venom PLA2-I | 2 µM (IC50) | ||||
| Naja naja venom PLA2-I | 7.1 µM (IC50) | ||||
| N. mocambique venom PLA2-I | 9.3 µM (IC50) | ||||
| Bee venom PLA2 | 1.4 µM (IC50) | p-Bromophenacyl bromide 80 µM (IC50) | [83,84] | ||
| Bee venom PLA2-induced paw edema | 42.4 (mg) | ||||
| Ki value Bee venom PLA2 | 0.57 µM | ||||
| Thielocin A2α (100) | PLA inhibition | Rat PLA2-II | 0.051 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 0.31 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielocin A2β (101) | PLA inhibition | Rat PLA2-II | 0.038 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 0.24 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | [70] | ||
| Rat PLA2-II | 0.032 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] | ||
| Human PLA2-II | 0.39 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielocin B1 (105) | PLA inhibition | Rat PLA2-II | 0.0078 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 0.17 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielocin B2 (106) | PLA inhibition | Rat PLA2-II | 0.070 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | [70] |
| Human PLA2-II | 2.7 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Thielocin B3 (107) | PLA inhibition | Human PLA2-I | 18 µM (IC50) | - | [70,80] |
| Rat PLA2-II | 0.012 µM (IC50) | Mepacrine 320 µM (IC50) p-Bromophenacyl bromide 6.7 µM (IC50) Manoalide 2.0 µM (IC50) | |||
| Human PLA2-II | 0.076 µM (IC50) | Mepacrine 76 µM (IC50) p-Bromophenacyl bromide 34 µM (IC50) Manoalide 1.5 µM (IC50) | |||
| Rat PLA2-I | 2.8 µM (IC50) | - | |||
| Mean Ki | 0.98 µM | - | |||
| Snake venom PLA2 | 0.0045 µM (IC50) 1.6 µM (ED50) | - | |||
| Exudate volume after carrageenan | 1.60 mL (conc. 1 mg/kg) 1.15 mL (conc. 3 mg/kg) | Indomethacin 1.08 mL (conc. 1 mg/kg) Dexamethasone 0.60 mL (conc. 0.1 mg/kg) | |||
| PLA2 activityin pleural exudate after carrageenan | 2.22 pmol/minute/mL (conc. 1 mg/kg) 0.76 pmol/minute/mL (conc. 3 mg/kg) | Indomethacin 7.36 pmol/minute/mL (conc. 1 mg/kg) Dexamethasone 7.94 pmol/minute/mL (conc. 0.1 mg/kg) | |||
| Thielocin B3 monomethyl ester B (108) | PLA inhibition | Human PLA2-II | 0.20 µM (IC50) | - | [80] |
| Snake venom PLA2 | 0.032 µM (IC50) 5.2 µM (ED50) | - | |||
| Thielocin B3 monomethyl ester C (109) | PLA inhibition | Human PLA2-II | 0.28 µM (IC50) | - | [80] |
| Snake venom PLA2 | 0.31 µM (IC50) 5.2 µM (ED50) | - | |||
| Thielocin B3 monomethyl ester D (110) | PLA inhibition | Human PLA2-II | 51 µM (IC50) | - | [80] |
| Snake venom PLA2 | >100 µM (IC50) 7.6 µM (ED50) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Fungal Depsides—Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities. Metabolites 2021, 11, 683. https://doi.org/10.3390/metabo11100683
Ibrahim SRM, Sirwi A, Eid BG, Mohamed SGA, Mohamed GA. Fungal Depsides—Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities. Metabolites. 2021; 11(10):683. https://doi.org/10.3390/metabo11100683
Chicago/Turabian StyleIbrahim, Sabrin R. M., Alaa Sirwi, Basma G. Eid, Shaimaa G. A. Mohamed, and Gamal A. Mohamed. 2021. "Fungal Depsides—Naturally Inspiring Molecules: Biosynthesis, Structural Characterization, and Biological Activities" Metabolites 11, no. 10: 683. https://doi.org/10.3390/metabo11100683