Oxylipin Profiles in Plasma of Patients with Wilson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics
2.2. Metabolomic Profiling
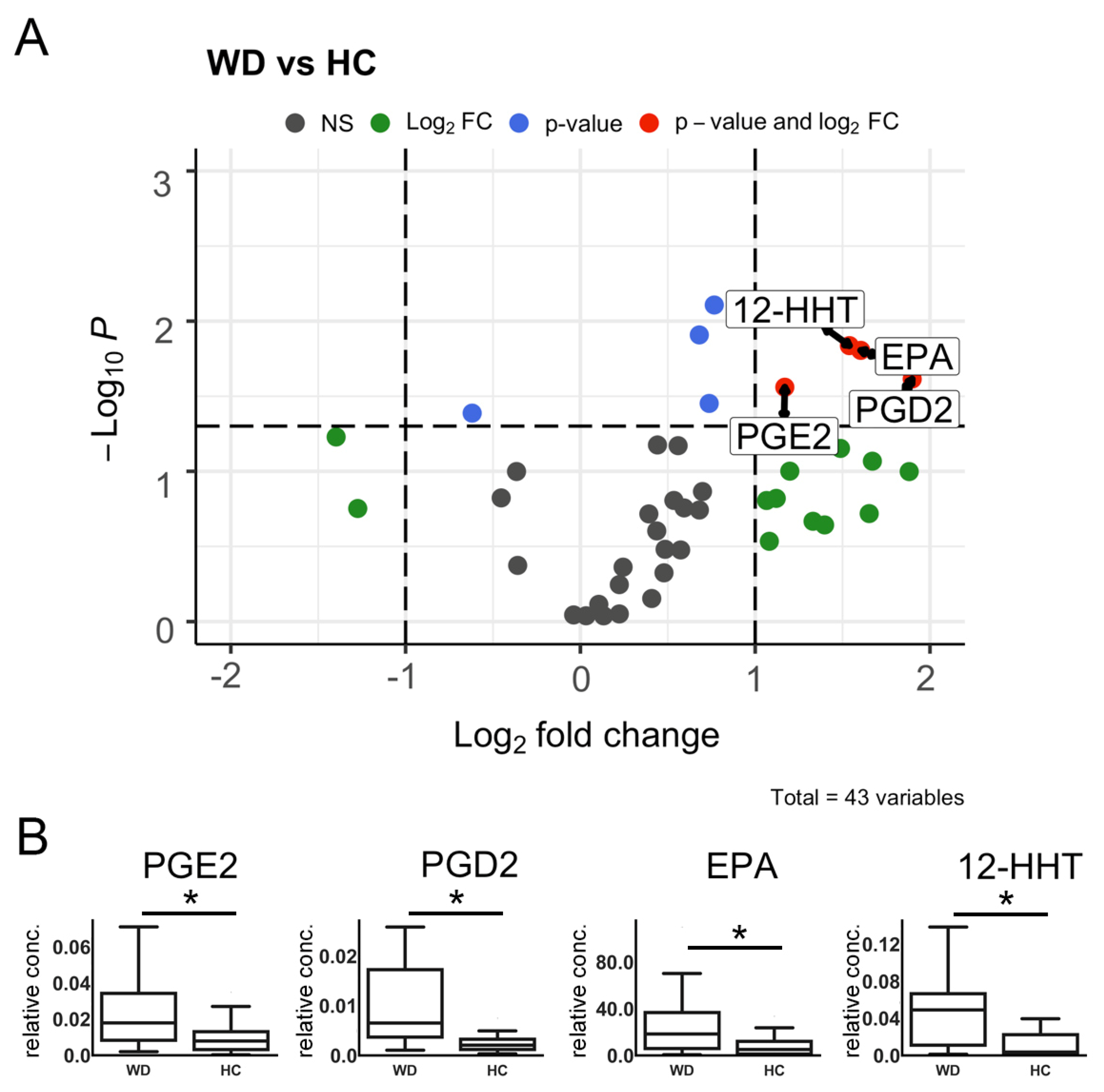
2.3. Volcano Plot Analysis
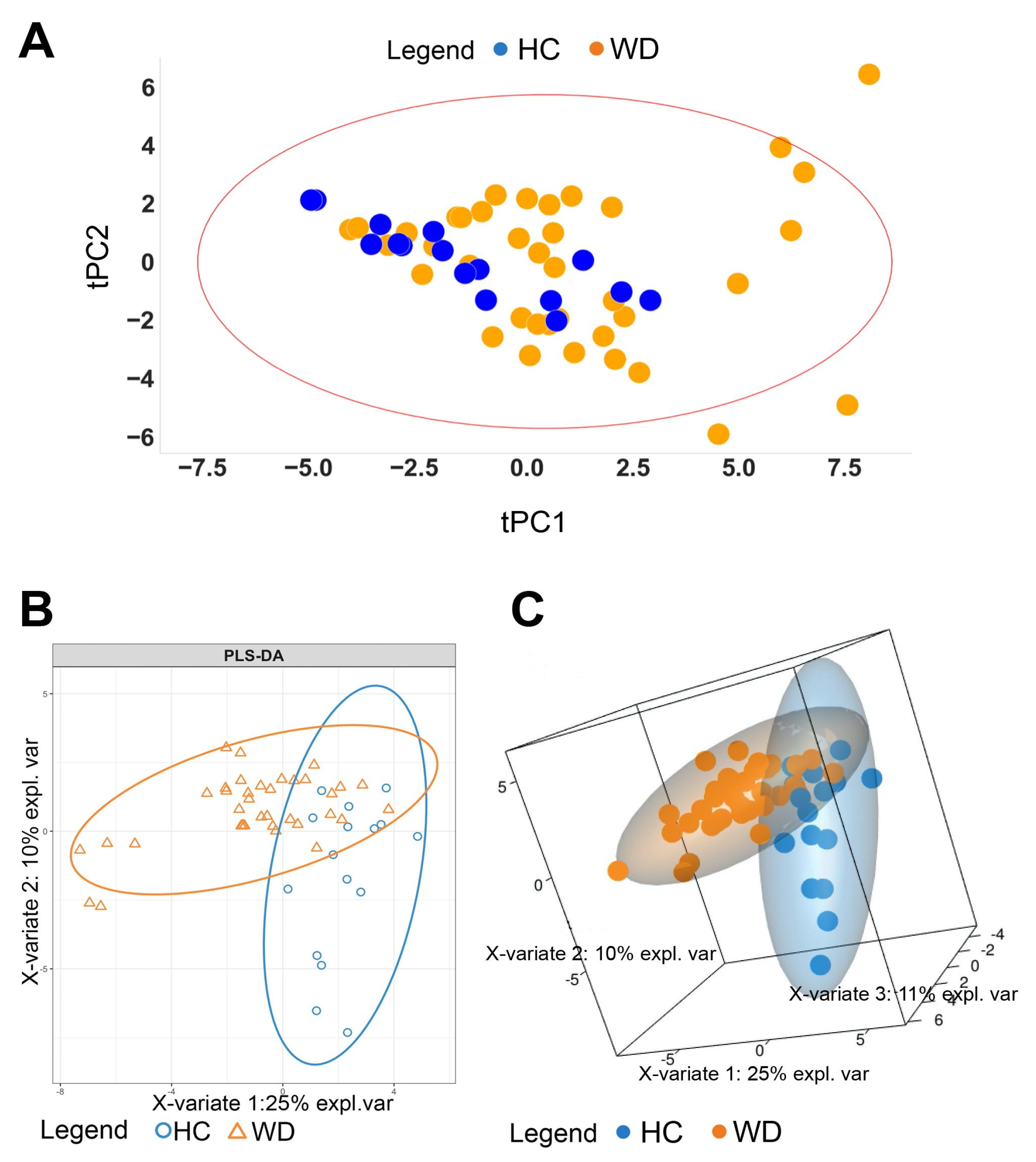
2.4. PLS-DA Model
2.5. Similarity Matrix
- (A)
- HC/WD patients;
- (B)
- Sex;
- (C)
- Nephropathy status;
- (D)
- Psychosomatic status; and
- (E)
- Form of the disease.
2.6. Pathway Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Population and Study Design
4.3. Clinical Evaluation
4.4. Blood Sample Collection
4.5. UPLC-MS/MS Conditions and Sample Preparation
4.6. Experimental Data Analysis and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| COX | Cyclooxygenase |
| CYP450 | Cytochrome P450 monooxygenase |
| DHA | Docosahexaenoic acid |
| DiHOME | Dihydroxyoctadecamonoenoic acid |
| HC | Healthy control |
| HDoHE | Hydroxydocosahexaenoic acid |
| HETE | Hydroxyeicosatetraenoic acid |
| HODE | Hydroxyoctadecadienoic acid |
| LA | Linoleic acid |
| LOX | Lipoxygenase |
| PG | Prostaglandin |
| PUFAs | Polyunsaturated fatty acids |
| WD | Wilson disease |
| UPLC-MS/MS | Ultra-performance liquid chromatography-tandem mass spectrometry |
References
- Bull, P.C.; Thomas, G.R.; Rommens, J.M.; Forbes, J.R.; Cox, D.W. The Wilson disease gene is a putative copper transporting P-type ATPase similar to the menkes gene. Nat. Genet. 1993, 5, 327–337. [Google Scholar] [CrossRef]
- Compston, A. Progressive lenticular degeneration: A familial nervous disease associated with cirrhosis of the liver, by S. A. Kinnier Wilson, (From the National Hospital, and the Laboratory of the National Hospital, Queen Square, London) Brain 1912: 34; 295–509. Brain 2009, 132, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Woimant, F.; Djebrani-Oussedik, N.; Poujois, A. New tools for Wilson’s disease diagnosis: Exchangeable copper fraction. Ann. Transl. Med. 2019, 7, S70. [Google Scholar] [CrossRef] [PubMed]
- Hermann, W. Classification and differential diagnosis of Wilson’s disease. Ann. Transl. Med. 2019, 7, S63. [Google Scholar] [CrossRef] [PubMed]
- Ala, A.; Walker, A.P.; Ashkan, K.; Dooley, J.S.; Schilsky, M.L. Wilson’s disease. Lancet 2007, 369, 397–408. [Google Scholar] [CrossRef]
- Huster, D. Wilson disease. Best Pract. Res. Clin. Gastroenterol. 2010, 24, 531–539. [Google Scholar] [CrossRef]
- Kieffer, D.A.; Medici, V. Wilson disease: At the crossroads between genetics and epigenetics—A review of the evidence. Liver Res. 2017, 1, 121–130. [Google Scholar] [CrossRef]
- Huster, D.; Khne, A.; Bhattacharjee, A.; Raines, L.; Jantsch, V.; Noe, J.; Schirrmeister, W.; Sommerer, I.; Sabri, O.; Berr, F.; et al. Diverse functional properties of wilson disease ATP7B variants. Gastroenterology 2012, 142, 947–956.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gromadzka, G.; Kruszyńska, M.; Wierzbicka, D.; Litwin, T.; Dziezyc, K.; Wierzchowska-Ciok, A.; Chabik, G.; Członkowska, A. Gene variants encoding proteins involved in antioxidant defense system and the clinical expression of Wilson disease. Liver Int. 2015, 35, 215–222. [Google Scholar] [CrossRef]
- Ferenci, P.; Stremmel, W.; Członkowska, A.; Szalay, F.; Viveiros, A.; Stättermayer, A.F.; Bruha, R.; Houwen, R.; Pop, T.L.; Stauber, R.; et al. Age and Sex but Not ATP7B Genotype Effectively Influence the Clinical Phenotype of Wilson Disease. Hepatology 2019, 69, 1464–1476. [Google Scholar] [CrossRef]
- Mordaunt, C.E.; Kieffer, D.A.; Shibata, N.M.; Członkowska, A.; Litwin, T.; Weiss, K.H.; Zhu, Y.; Bowlus, C.L.; Sarkar, S.; Cooper, S.; et al. Epigenomic signatures in liver and blood of Wilson disease patients include hypermethylation of liver-specific enhancers. Epiganetics Chromatin 2019, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Mazi, T.A.; Sarode, G.V.; Czlonkowska, A.; Litwin, T.; Kim, K.; Shibata, N.M.; Medici, V. Dysregulated choline, methionine, and aromatic amino acid metabolism in patients with wilson disease: Exploratory metabolomic profiling and implications for hepatic and neurologic phenotypes. Int. J. Mol. Sci. 2019, 20, 5937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarode, G.V.; Kim, K.; Kieffer, D.A.; Shibata, N.M.; Litwin, T.; Czlonkowska, A.; Medici, V. Metabolomics profiles of patients with Wilson disease reveal a distinct metabolic signature. Metabolomics 2019, 15, 43. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [Green Version]
- Layé, S.; Nadjar, A.; Joffre, C.; Bazinet, R.P. Anti-Inflammatory Effects of Omega-3 Fatty Acids in the Brain: Physiological Mechanisms and Relevance to Pharmacology. Pharmacol. Rev. 2018, 70, 12–38. [Google Scholar] [CrossRef]
- Chistyakov, D.V.; Astakhova, A.A.; Sergeeva, M.G. Resolution of inflammation and mood disorders. Exp. Mol. Pathol. 2018, 105, 190–201. [Google Scholar] [CrossRef]
- Burstein, S.H. The chemistry, biology and pharmacology of the cyclopentenone prostaglandins. Prostaglandins Other Lipid Mediat. 2020, 148, 106408. [Google Scholar] [CrossRef]
- Nagy, L.; Tontonoz, P.; Alvarez, J.G.A.; Chen, H.; Evans, R.M. Oxidized LDL regulates macrophage gene expression through ligand activation of PPARγ. Cell 1998, 93, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Moran, J.H.; Weise, R.; Schnellmann, R.G.; Freeman, J.P.; Grant, D.F. Cytotoxicity of linoleic acid diols to renal proximal tubular cells. Toxicol. Appl. Pharmacol. 1997, 146, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Huster, D.; Purnat, T.D.; Burkhead, J.L.; Ralle, M.; Fiehn, O.; Stuckert, F.; Olson, N.E.; Teupser, D.; Lutsenko, S. High copper selectively alters lipid metabolism and cell cycle machinery in the mouse model of Wilson disease. J. Biol. Chem. 2007, 282, 8343–8355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wooton-Kee, C.R.; Jain, A.K.; Wagner, M.; Grusak, M.A.; Finegold, M.J.; Lutsenko, S.; Moore, D.D. Elevated copper impairs hepatic nuclear receptor function in Wilson’s disease. J. Clin. Investig. 2015, 125, 3449–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.P.; Koganti, L.; Muchenditsi, A.; Pendyala, V.S.; Huso, D.; Hankin, J.; Murphy, R.C.; Huster, D.; Merle, U.; Mangels, C.; et al. Activation of liver X receptor/retinoid X receptor pathway ameliorates liver disease in Atp7B−/− (Wilson disease) mice. Hepatology 2016, 63, 1828–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.K.; Piccolo, B.D.; John, C.S.; Green, Z.D.; Thyfault, J.P.; Adams, S.H. Oxylipin profiling of alzheimer’s disease in nondiabetic and type 2 diabetic elderly. Metabolites 2019, 9, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Lang, S.; Duan, Y.; Wang, Y.; Shawcross, D.L.; Louvet, A.; Mathurin, P.; Ho, S.B.; Stärkel, P.; Schnabl, B. Serum and Fecal Oxylipins in Patients with Alcohol-Related Liver Disease. Dig. Dis. Sci. 2019, 64, 1878–1892. [Google Scholar] [CrossRef]
- Chocholoušková, M.; Jirásko, R.; Vrána, D.; Gatěk, J.; Melichar, B.; Holčapek, M. Reversed phase UHPLC/ESI-MS determination of oxylipins in human plasma: A case study of female breast cancer. Anal. Bioanal. Chem. 2019, 411, 1239–1251. [Google Scholar] [CrossRef]
- Ryan, A.; Nevitt, S.J.; Tuohy, O.; Cook, P. Biomarkers for diagnosis of Wilson’s disease. Cochrane Database Syst. Rev. 2019, 11, 1465–1858. [Google Scholar] [CrossRef]
- Rago, B.; Fu, C. Development of a high-throughput ultra performance liquid chromatography-mass spectrometry assay to profile 18 eicosanoids as exploratory biomarkers for atherosclerotic diseases. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 936, 25–32. [Google Scholar] [CrossRef]
- Shishehbor, M.H.; Zhang, R.; Medina, H.; Brennan, M.L.; Brennan, D.M.; Ellis, S.G.; Topol, E.J.; Hazen, S.L. Systemic elevations of free radical oxidation products of arachidonic acid are associated with angiographic evidence of coronary artery disease. Free Radic. Biol. Med. 2006, 41, 1678–1683. [Google Scholar] [CrossRef] [Green Version]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress part I: Mechanisms involved in metal-induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Gaetke, L.M.; Chow, C.K. Copper toxicity, oxidative stress, and antioxidant nutrients. Toxicology 2003, 189, 147–163. [Google Scholar] [CrossRef]
- Santoro, N.; Caprio, S.; Feldstein, A.E. Oxidized metabolites of linoleic acid as biomarkers of liver injury in nonalcoholic steatohepatitis. Clin. Lipidol. 2013, 8, 411–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, N.; Kumasaka, R.; Osawa, H.; Yamabe, H.; Shirato, K.I.; Fujita, T.; Murakami, R.I.; Shimada, M.; Nakamura, M.; Okumura, K.; et al. Effects of eicosapentaenoic acids on oxidative stress and plasma fatty acid composition in patients with lupus nephritis. In Vivo (Brooklyn) 2005, 19, 879–882. [Google Scholar]
- Payahoo, L.; Khajebishak, Y.; Jafarabadi, M.A.; Ostadrahimi, A. Oleoylethanolamide supplementation reduces inflammation and oxidative stress in obese people: A clinical trial. Adv. Pharm. Bull. 2018, 8, 479–487. [Google Scholar] [CrossRef]
- Du, C.; Fujii, Y.; Ito, M.; Harada, M.; Moriyama, E.; Shimada, R.; Ikemoto, A.; Okuyama, H. Dietary polyunsaturated fatty acids suppress acute hepatitis, alter gene expression and prolong survival of female Long-Evans Cinnamon rats, a model of Wilson disease. J. Nutr. Biochem. 2004, 15, 273–280. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids, inflammation, and immunity. Lipids 2001, 36, 1007–1024. [Google Scholar] [CrossRef]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Bensinger, S.J.; Tontonoz, P. Integration of metabolism and inflammation by lipid-activated nuclear receptors. Nature 2008, 454, 470–477. [Google Scholar] [CrossRef]
- Nagasaka, H.; Miida, T.; Inui, A.; Inoue, I.; Tsukahara, H.; Komatsu, H.; Hiejima, E.; Fujisawa, T.; Yorifuji, T.; Hiranao, K.; et al. Fatty liver and anti-oxidant enzyme activities along with peroxisome proliferator-activated receptors γ and α expressions in the liver of Wilson’s disease. Mol. Genet. Metab. 2012, 107, 542–547. [Google Scholar] [CrossRef]
- He, K.; Chen, Z.; Ma, Y.; Pan, Y. Identification of high-copper-responsive target pathways in Atp7b knockout mouse liver by GSEA on microarray data sets. Mamm. Genome 2011, 22, 703–713. [Google Scholar] [CrossRef]
- Fu, J.; Oveisi, F.; Gaetani, S.; Lin, E.; Piomelli, D. Oleoylethanolamide, an endogenous PPAR-α agonist, lowers body weight and hyperlipidemia in obese rats. Neuropharmacology 2005, 48, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Aleshin, S.; Strokin, M.; Sergeeva, M.; Reiser, G. Peroxisome proliferator-activated receptor (PPAR)β/δ, a possible nexus of PPARα- and PPARγ-dependent molecular pathways in neurodegenerative diseases: Review and novel hypotheses. Neurochem. Int. 2013, 63, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Hu, S.; Xu, B.; Snyder, G.D.; Harmon, S.; Yao, J.; Liu, Y.; Sangras, B.; Falck, J.R.; Weintraub, N.L.; et al. 14,15-Dihydroxyeicosatrienoic acid activates peroxisome proliferator-activated receptor-α. Am. J. Physiol. Hear. Circ. Physiol. 2006, 290, H55–H63. [Google Scholar] [CrossRef] [PubMed]
- Mangino, M.; Roederer, M.; Beddall, M.H.; Nestle, F.O.; Spector, T.D. Innate and adaptive immune traits are differentially affected by genetic and environmental factors. Nat. Commun. 2017, 8, 13850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostermann, A.I.; Greupner, T.; Kutzner, L.; Hartung, N.M.; Hahn, A.; Schuchardt, J.P.; Schebb, N.H. Intra-individual variance of the human plasma oxylipin pattern: Low inter-day variability in fasting blood samples: Versus high variability during the day. Anal. Methods 2018, 10, 4935–4944. [Google Scholar] [CrossRef] [Green Version]
- Chistyakov, D.V.; Azbukina, N.V.; Astakhova, A.A.; Goriainov, S.V.; Chistyakov, V.V.; Tiulina, V.V.; Baksheeva, V.E.; Kotelin, V.I.; Fedoseeva, E.V.; Zamyatnin, A.A.; et al. Comparative lipidomic analysis of inflammatory mediators in the aqueous humor and tear fluid of humans and rabbits. Metabolomics 2020, 16, 27. [Google Scholar] [CrossRef]
- FDA. Guidance for Industry Process Validation: General Principles and Practices. 2011. Available online: https://www.fda.gov/files/drugs/published/Process-Validation--General-Principles-and-Practices.pdf (accessed on 27 May 2020).
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A. mixOmics: An R package for omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wilson Disease Patients | ||||||
| Sex | F (n = 22) | M (n = 17) | ||||
| mean | sd | n | mean | sd | n | |
| Age | 35.68 | 13.17 | 32.18 | 12.36 | ||
| Serum Cu, mkM | 8.5 | 4.3 | 9.62 | 4.36 | ||
| Shvab scale, % | 80.95 | 18.41 | 67.5 | 22.36 | ||
| Leipzig score | 7.32 | 2.19 | 6.42 | 2.57 | ||
| Ceruloplasmin, mg/dL | 9.61 | 7.2 | 12.78 | 8.02 | ||
| Height, cm | 169.95 | 5.55 | 180.8 | 7.44 | ||
| Longevity illness, years | 13.86 | 11.34 | 9.81 | 9.32 | ||
| Longevity treatment, years | 12.86 | 11.25 | 8.21 | 8.75 | ||
| Weight, kg | 61.2 | 13.27 | 75.67 | 12.21 | ||
| Form (akinetic-rigid/trembling/others) | 2014/6/2 | 2011/4/2 | ||||
| Nephropathy | 5 | 8 | ||||
| Portal hypertension | 5 | 11 | ||||
| Psycoproductive somatic | 8 | 6 | ||||
| Healthy Donors | ||||||
| F (11) | M (5) | |||||
| mean | sd | n | mean | sd | n | |
| Age | 37.88 | 15.96 | 49.2 | 12.19 | ||
| Name | 12-HHT * | EPA * | 14,15-DHET | 9-HODE | OEA | PGE2 * | 9-KODE |
|---|---|---|---|---|---|---|---|
| VIP-scores | 1.899456 | 1.741633 | 1.739218 | 1.624940 | 1.617023 | 1.594828 | 1.837164 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azbukina, N.V.; Lopachev, A.V.; Chistyakov, D.V.; Goriainov, S.V.; Astakhova, A.A.; Poleshuk, V.V.; Kazanskaya, R.B.; Fedorova, T.N.; Sergeeva, M.G. Oxylipin Profiles in Plasma of Patients with Wilson’s Disease. Metabolites 2020, 10, 222. https://doi.org/10.3390/metabo10060222
Azbukina NV, Lopachev AV, Chistyakov DV, Goriainov SV, Astakhova AA, Poleshuk VV, Kazanskaya RB, Fedorova TN, Sergeeva MG. Oxylipin Profiles in Plasma of Patients with Wilson’s Disease. Metabolites. 2020; 10(6):222. https://doi.org/10.3390/metabo10060222
Chicago/Turabian StyleAzbukina, Nadezhda V., Alexander V. Lopachev, Dmitry V. Chistyakov, Sergei V. Goriainov, Alina A. Astakhova, Vsevolod V. Poleshuk, Rogneda B. Kazanskaya, Tatiana N. Fedorova, and Marina G. Sergeeva. 2020. "Oxylipin Profiles in Plasma of Patients with Wilson’s Disease" Metabolites 10, no. 6: 222. https://doi.org/10.3390/metabo10060222