
SIRT1/Nrf2/NF-κB Signaling Mediates Anti-Inflammatory and Anti-Apoptotic Activities of Oleanolic Acid in a Mouse Model of Acute Hepatorenal Damage
 ,
,  ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedures
2.2. Plant Materials
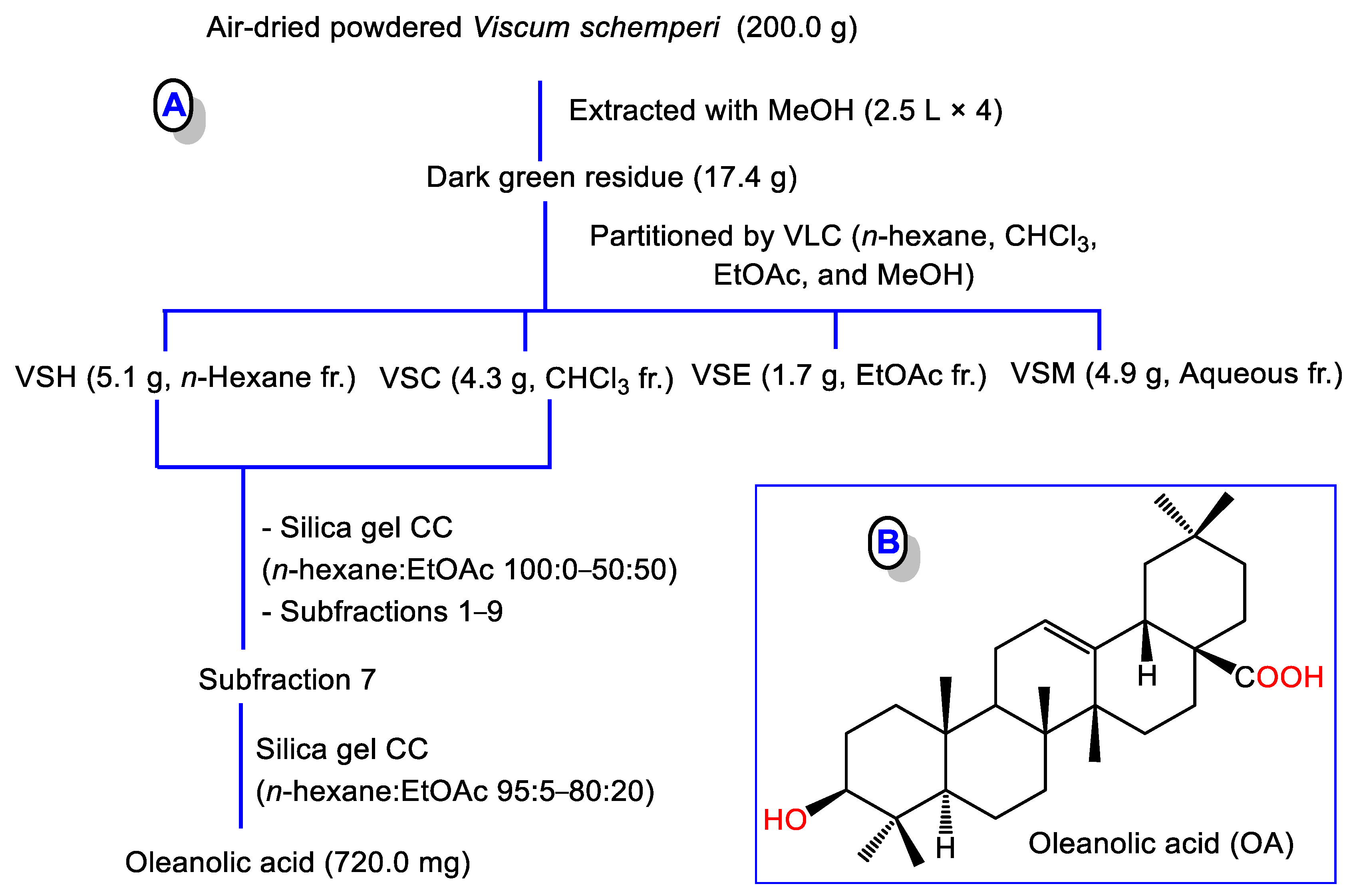
2.3. Extraction and Isolation of OA
2.4. Materials and Chemicals
2.5. Animals
2.6. Experimental Procedures
2.6.1. Serum Markers of Hepatorenal Damage
2.6.2. Oxidative and Antioxidant Markers
MDA
4-HNE
GSH
SOD
TAC
NOx
2.6.3. Histology and Immunohistochemistry (IHC)
2.6.4. ELISA
2.6.5. RT-PCR
2.7. Data Analysis
3. Results
3.1. Purification and Identification of OA
3.2. Functional Status and Histopathology of Liver and Kidney
3.3. Antioxidants and Oxidative Stress in the Kidney and Liver
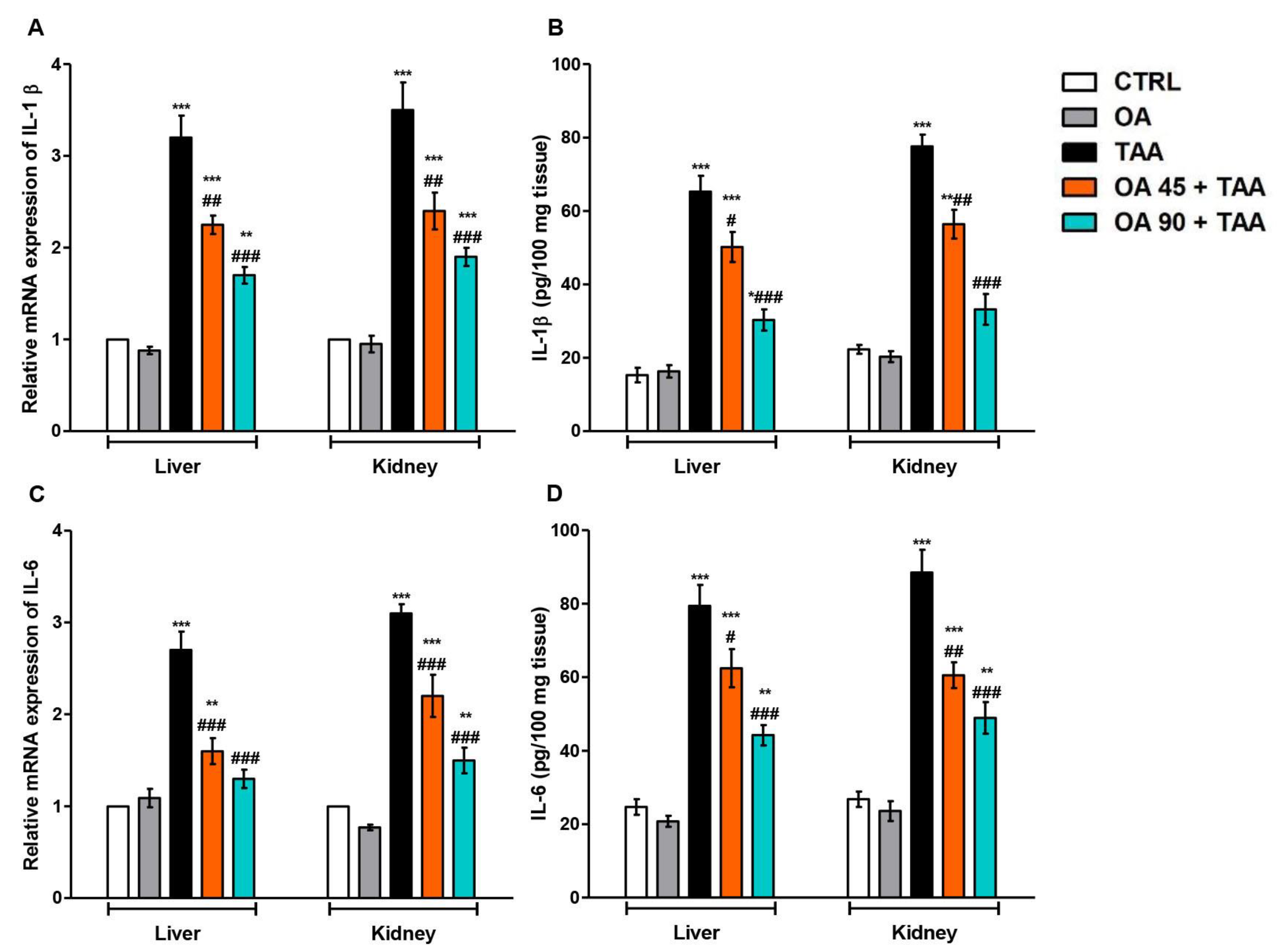
3.4. Inflammatory Response in the Liver and Kidney
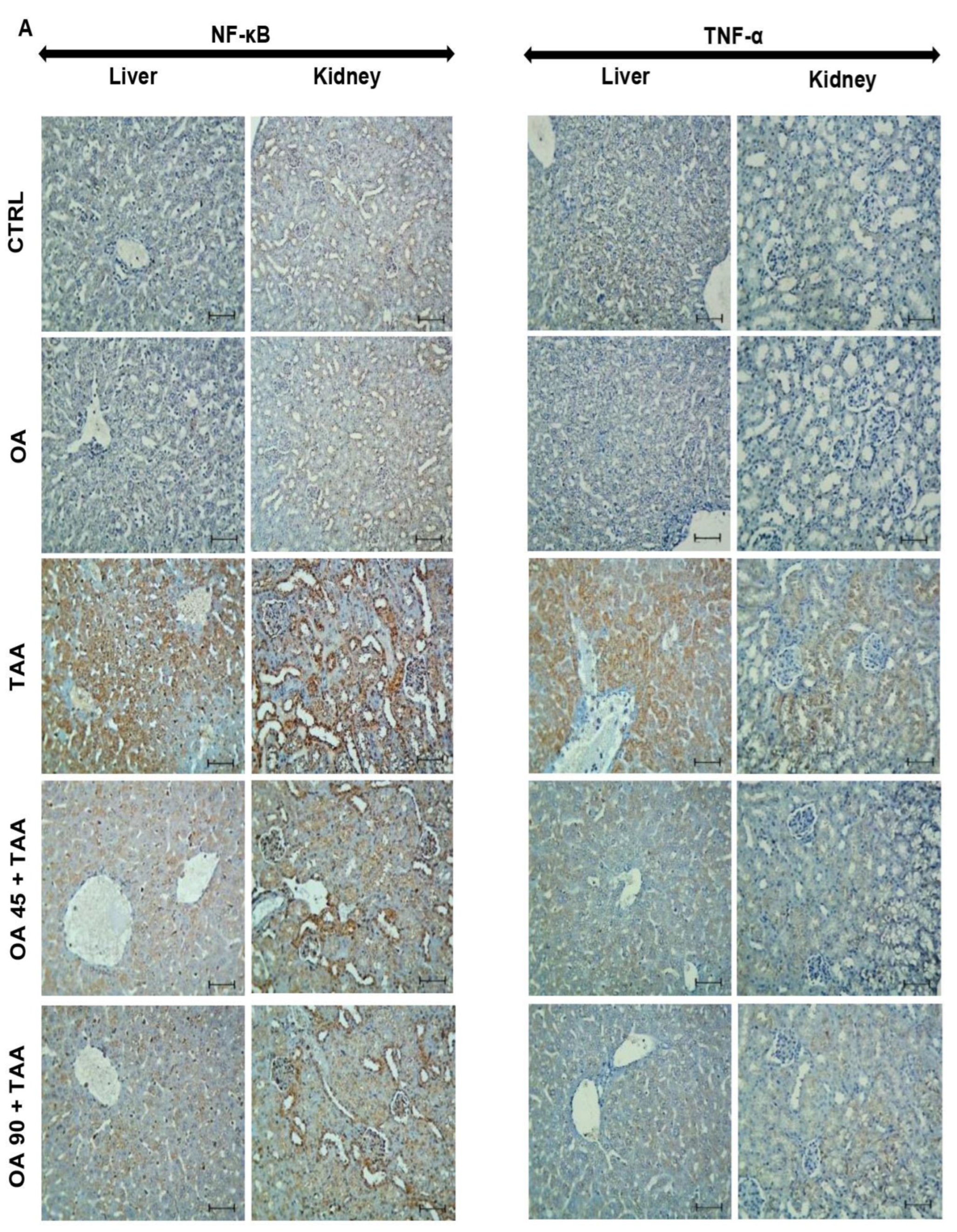
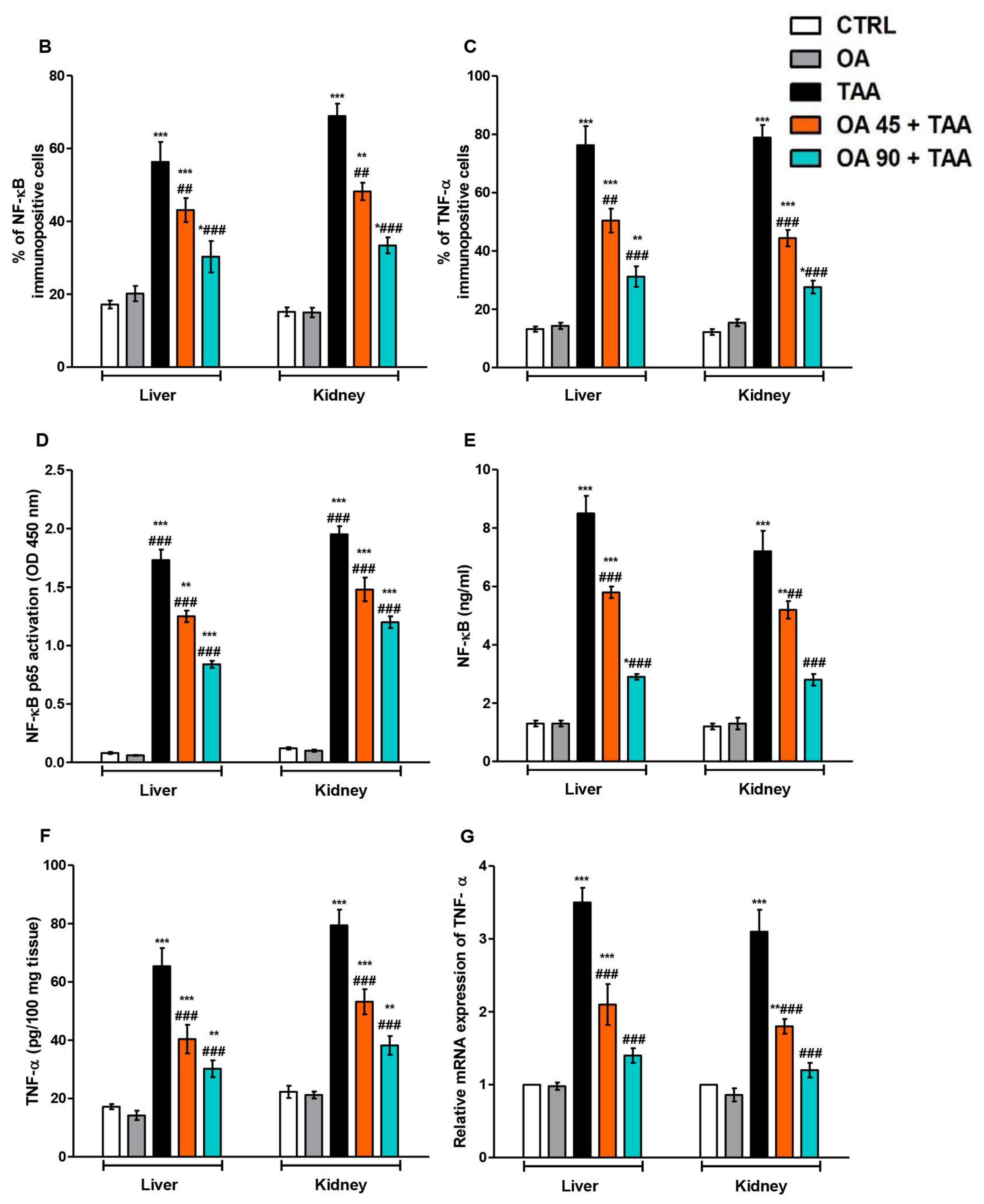
3.5. NF-κB/TNF-α Inflammatory Signaling Pathway in the Kidney and Liver
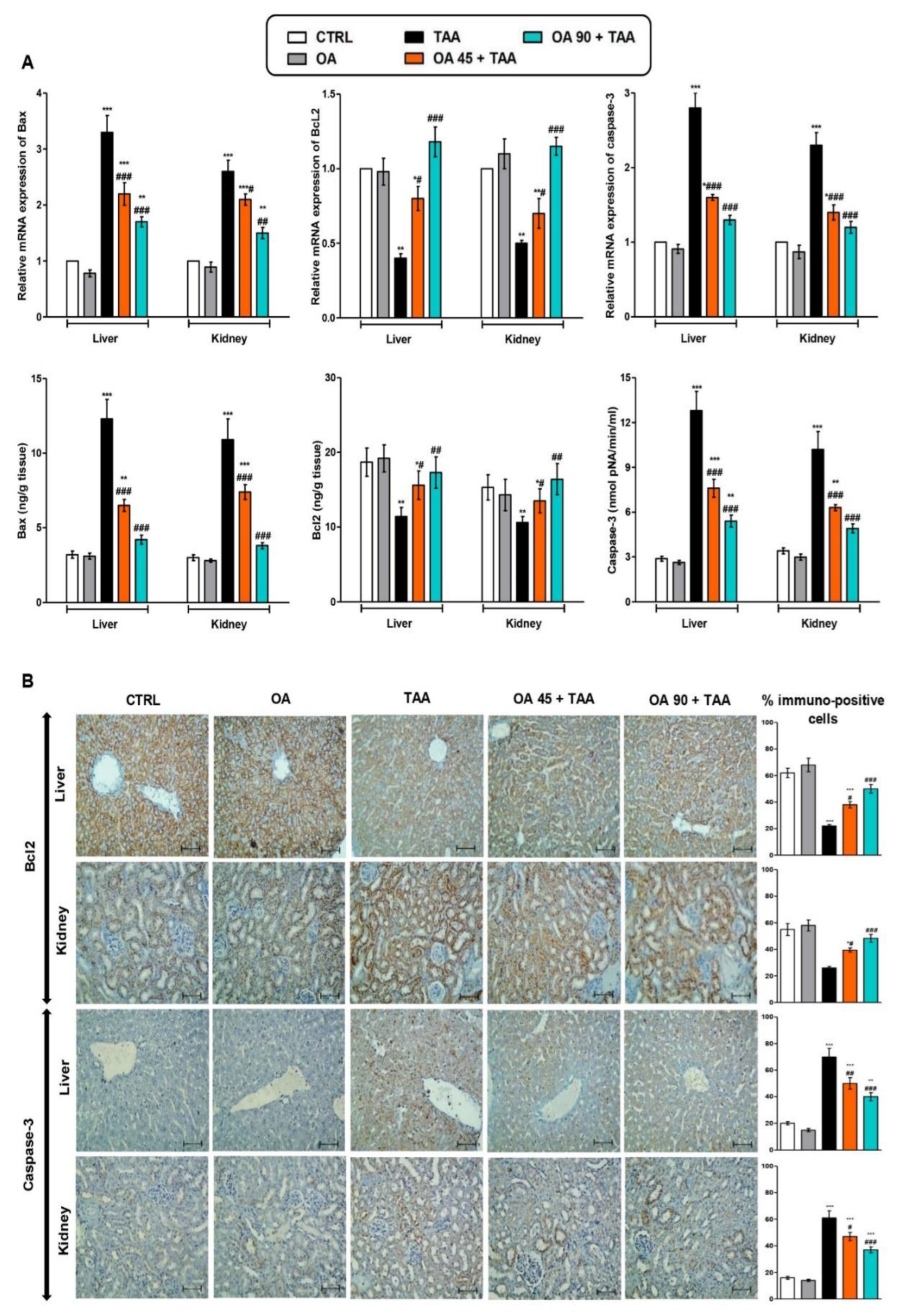
3.6. Apoptotic Changes in the Liver and Kidney
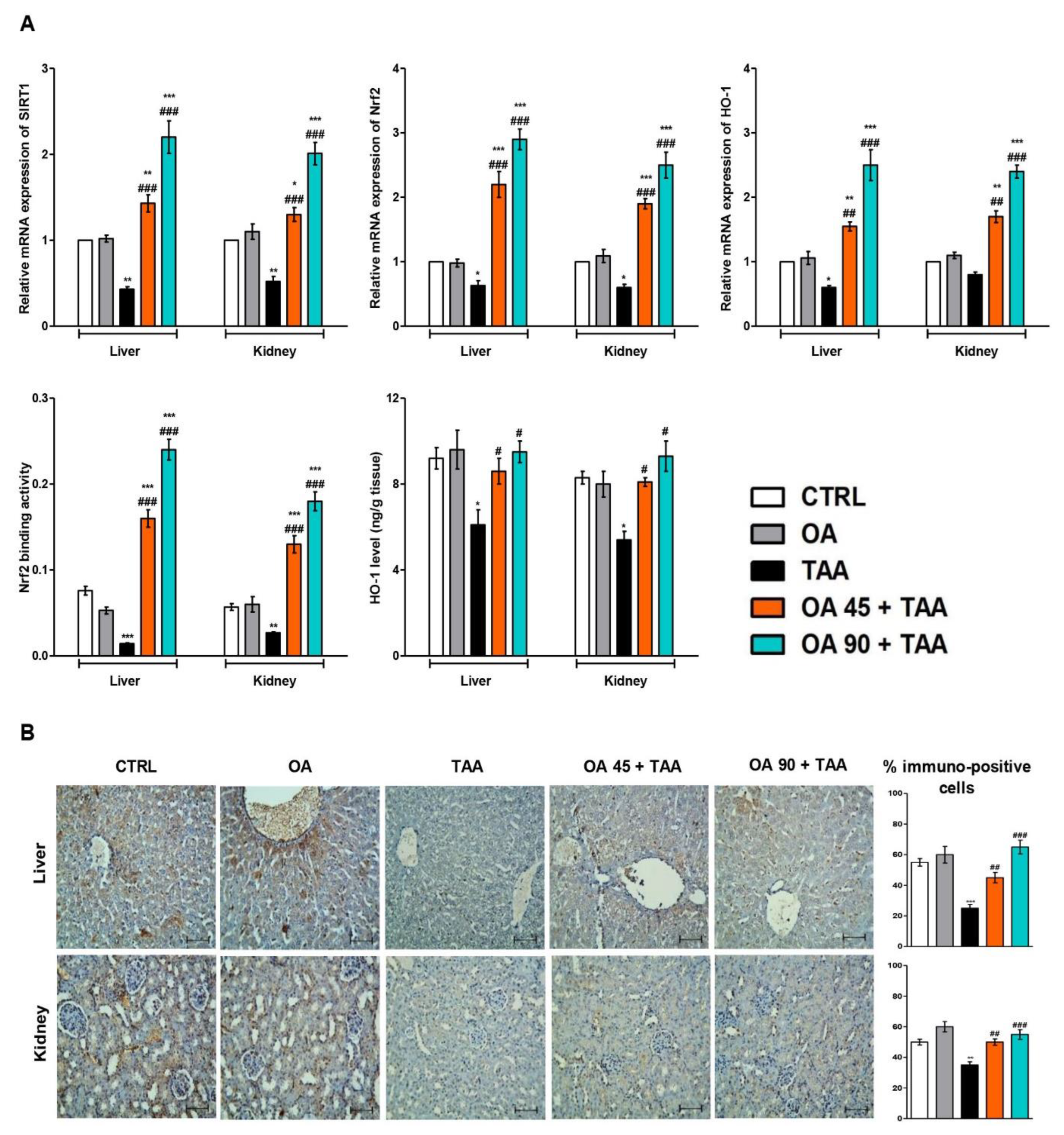
3.7. SIRT1/Nrf2/HO-1 Signaling in Liver and Kidney
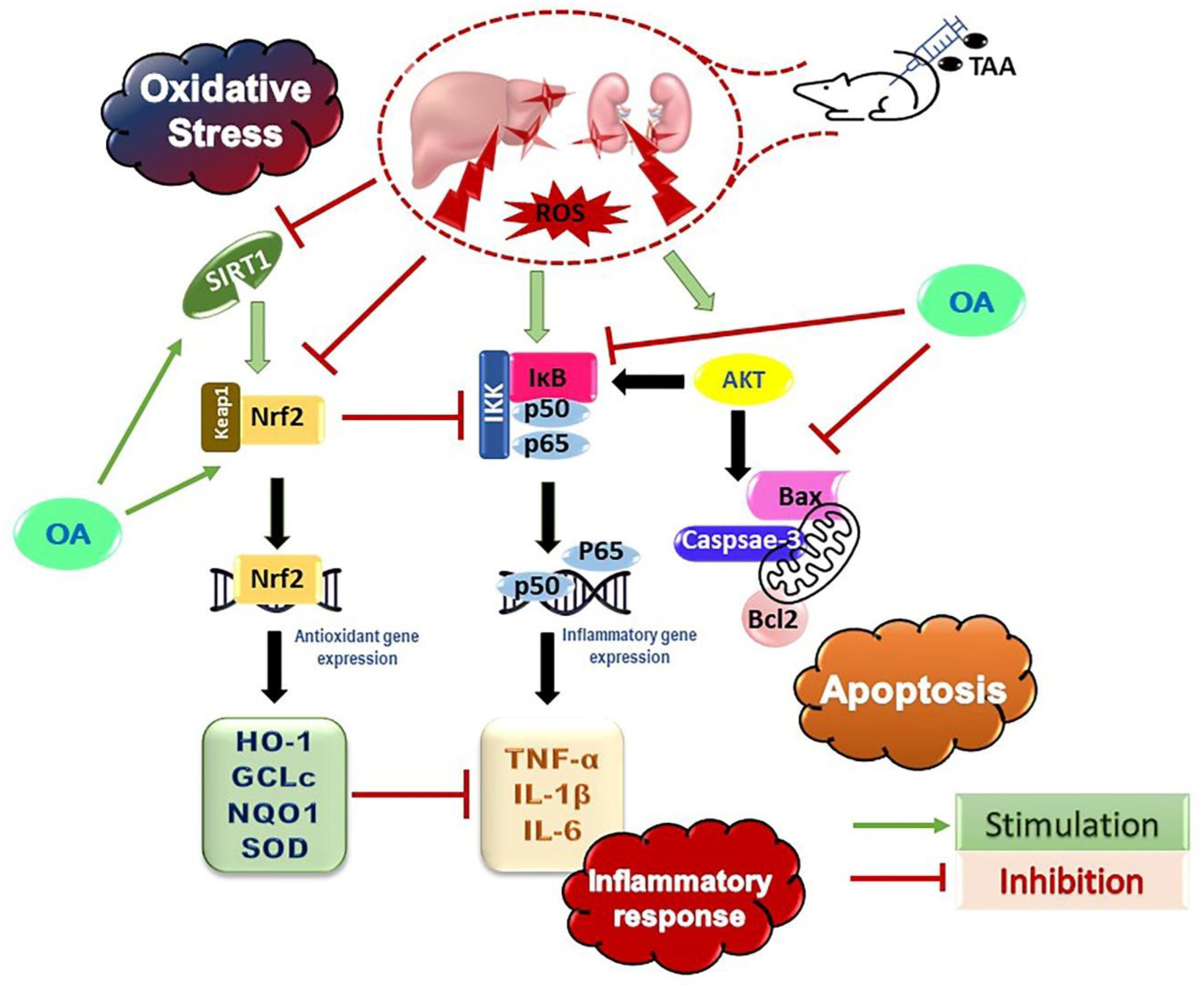
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, D.Z.; Chu, P.Y.; Li, Y.H.; Chandrasekaran, V.R.M.; Liu, M.Y. Role of flavin-containing-monooxygenase-dependent neutrophil activation in thioacetamide-induced hepatic inflammation in rats. Toxicology 2012, 298, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Türkmen, N.B.; Hande, Y.Ü.C.E.; Taşlidere, A.; Şahin, Y.; Çiftçi, O. The Ameliorate effects of nerolidol on thioacetamide-induced oxidative damage in heart and kidney tissue. Turk. J. Pharm. Sci. 2022, 19, 1–8. [Google Scholar] [CrossRef]
- Wang, K.; Deng, Y.; Zhang, J.; Cheng, B.; Huang, Y.; Meng, Y.; Zhong, K.; Xiong, G.; Guo, J.; Liu, Y.; et al. Toxicity of thioacetamide and protective effects of quercetin in zebrafish (Danio rerio) larvae. Environ. Toxicol. 2021, 36, 2062–2072. [Google Scholar] [CrossRef]
- Alamery, S.; Zargar, S.; Yaseen, F.; Wani, T.A.; Siyal, A. Evaluation of the effect of Wheat Germ oil and Olmutinib on the thioacetamide-induced liver and kidney toxicity in mice. Life 2022, 12, 900. [Google Scholar] [CrossRef]
- Vázquez-Atanacio, M.J.; Bautista, M.; González-Cortazar, M.; Romero-Estrada, A.; De la O.-Arciniega, M.; Castañeda-Ovando, A.; Sosa-Gutiérrez, C.G.; Ojeda-Ramírez, D. Nephroprotective activity of Papaloquelite (Porophyllum ruderale) in Thioacetamide-Induced Injury Model. Plants 2022, 11, 3460. [Google Scholar] [CrossRef]
- Zargar, S.; Alonazi, M.; Rizwana, H.; Wani, T.A. Resveratrol reverses thioacetamide-induced renal assault with respect to oxidative stress, renal function, DNA damage, and cytokine release in Wistar rats. Oxid. Med. Cell. Longev. 2019, 2019, 1702959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, H.M.; Oda, S.S.; Sedeek, E.K. Protective effect of Lactéol® forte against thioacetamide-induced hepatic injury in male albino rats. Alex. J. Vet. Sci. 2020, 67, 92–98. [Google Scholar] [CrossRef]
- Ejiofor, E.U.; Oyedemi, S.O.; Onoja, S.O.; Omeh, N.Y. Amaranthus hybridus Linn leaf extract ameliorates oxidative stress and hepatic damage abnormalities induced by thioacetamide in rats. S. Afr. J. Bot. 2022, 146, 213–221. [Google Scholar] [CrossRef]
- El-Kashef, D.H.; Serrya, M.S. Sitagliptin ameliorates thioacetamide-induced acute liver injury via modulating TLR4/NF-KB signaling pathway in mice. Life Sci. 2019, 228, 266–273. [Google Scholar] [CrossRef]
- Ali, M.; Khan, T.; Fatima, K.; Ali, Q.U.; Ovais, M.; Khalil, A.T.; Ullah, I.; Raza, A.; Shinwari, Z.K.; Idrees, M. Selected hepatoprotective herbal medicines: Evidence from ethnomedicinal applications, animal models, and possible mechanism of actions. Phytother. Res. 2018, 32, 199–215. [Google Scholar] [CrossRef] [Green Version]
- Anaeigoudari, A. Hepato-and reno-protective effects of thymoquinone, crocin, and carvacrol: A comprehensive review. Asian Pac. J. Trop. Biomed. 2022, 12, 185–196. [Google Scholar] [CrossRef]
- Chen, D.Q.; Hu, H.H.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Zhao, Y.Y. Natural products for the prevention and treatment of kidney disease. Phytomedicine 2018, 50, 50–60. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Simental-Mendía, L.E.; Butler, A.E.; Sahebkar, A. Protective effects of plant-derived natural products on renal complications. J. Cell. Physiol. 2019, 234, 12161–12172. [Google Scholar] [CrossRef]
- Rahman, M.A.; Akter, S.; Dorotea, D.; Mazumder, A.; Uddin, M.N.; Hannan, M.A.; Hossen, M.J.; Ahmed, M.S.; Kim, W.; Kim, B.; et al. Renoprotective potentials of small molecule natural products targeting mitochondrial dysfunction. Front. Pharmacol. 2022, 13, 925993. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; El-Agamy, D.S.; Abdallah, H.M.; Ahmed, N.; Elkablawy, M.A.; Mohamed, G.A. Protective activity of tovophyllin A, a xanthone isolated from Garcinia mangostana pericarps, against acetaminophen-induced liver damage: Role of Nrf2 activation. Food Funct. 2018, 9, 3291–3300. [Google Scholar] [CrossRef]
- Sirwi, A.; Shaik, R.A.; Alamoudi, A.J.; Eid, B.G.; Kammoun, A.K.; Ibrahim, S.R.M.; Mohamed, G.A.; Abdallah, H.M.; Abdel-Naim, A.B. Mokko lactone attenuates doxorubicin-induced hepatotoxicity in rats: Emphasis on Sirt-1/FOXO1/NF-κB axis. Nutrients 2021, 13, 4142. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.A.; Ibrahim, S.R.M.; El-Agamy, D.S.; Elsaed, W.M.; Sirwi, A.; Asfour, H.Z.; Koshak, A.E.; Elhady, S.S. Cucurbitacin E glucoside alleviates concanavalin A-induced hepatitis through enhancing SIRT1/Nrf2/HO-1 and inhibiting NF-ĸB/NLRP3 signaling pathways. J. Ethnopharmacol. 2022, 292, 115223. [Google Scholar] [CrossRef] [PubMed]
- Khayat, M.T.; Mohammad, K.A.; Mohamed, G.A.; El-Agamy, D.S.; Elsaed, W.M.; Ibrahim, S.R.M. γ-Mangostin abrogates AINT-induced cholestatic liver injury: Impact on Nrf2/NF-κB/NLRP3/Caspase-1/IL-1β/GSDMD signalling. Life Sci. 2023, 322, 121663. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Wei, X.Y.; Qiu, Z.D.; Gong, L.; Chen, Z.Y.; Ma, Y.; Shen, Y.; Zhao, Y.J.; Wang, W.H.; Yang, B. Exploring the resources of the genus Viscum for potential therapeutic applications. J. Ethnopharmacol. 2021, 277, 114233. [Google Scholar] [CrossRef]
- Hawu, O.; Ravhuhali, K.E.; Musekwa, M.G.; Sipango, N.; Mudau, H.S.; Mokoboki, K.H.; Moyo, B. Utilization of the Viscum species for diet and medicinal purposes in ruminants: A Review. Animals 2022, 12, 2569. [Google Scholar] [CrossRef]
- Skrypnik, L.; Maslennikov, P.; Feduraev, P.; Pungin, A.; Belov, N. Ecological and landscape factors affecting the spread of European mistletoe (Viscum album L.) in urban areas (A case study of the kaliningrad city, Russia). Plants 2020, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.N.; Saha, C.; Galun, D.; Upreti, D.K.; Bayry, J.; Kaveri, S.V. European Viscum album: A potent phytotherapeutic agent with multifarious phytochemicals, pharmacological properties and clinical evidence. RSC Adv. 2016, 6, 23837–23857. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, H.M.; Farag, M.A.; Abdel-Naim, A.B.; Ghareib, S.A.; Abdel-Sattar, E.A. Mechanistic evidence of Viscum schimperi (Viscaceae) antihyperglycemic activity: From a bioactivity-guided approach to comprehensive metabolite profiling. Phytother. Res. 2015, 29, 1737–1743. [Google Scholar] [CrossRef]
- Abdallah, H.M.; ALGhamdi, D.O.; Al-Salem, M.S.; Alattas, M.; El-Bassossy, H.M.; Alahdal, A.M.; Shehata, I.A.; Abdel-Sattar, E.A. Effect of Viscum schimperi on advanced glycation endproducts formation. Pak. J. Pharm. Sci. 2016, 29 (Suppl. 6), 2307–2316. [Google Scholar] [PubMed]
- Abdel-Sattar, E.A.; Elberry, A.A.; Harraz, F.M.; Ghareib, S.A.; Nagy, A.A.; Gabr, S.A. Antihyperglycemic and hypolipidaemic effects of the methanolic extract of Saudi mistletoe (Viscum schimperi Engl.). J. Adv. Res. 2011, 2, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Potočnjak, I.; Šimić, L.; Vukelić, I.; Domitrović, R. Oleanolic acid attenuates cisplatin-induced nephrotoxicity in mice and chemosensitizes human cervical cancer cells to cisplatin cytotoxicity. Food Chem. Toxicol. 2019, 132, 110676. [Google Scholar] [CrossRef]
- Reisman, S.A.; Aleksunes, L.M.; Klaassen, C.D. Oleanolic acid activates Nrf2 and protects from acetaminophen hepatotoxicity via Nrf2-dependent and Nrf2-independent processes. Biochem. Pharmacol. 2009, 77, 1273–1282. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.L.; Lu, Y.F.; Xu, S.F.; Wu, Q.; Liu, J. Oeanolic acid protects against the hepatotoxicity of D-galactosame plus endotoxin in mice. Biomed Pharmacother. 2017, 93, 1040–1046. [Google Scholar] [CrossRef]
- Stefanello, S.T.; Hartmann, D.D.; Amaral, G.P.; Courtes, A.A.; Leite, M.T.B.; da Silva, T.C.; Gonçalves, D.F.; Souza, M.B.; da Rosa, P.; Dornelles, C.L.; et al. Antioxidant protection by β-selenoamines against thioacetamide-induced oxidative stress and hepatotoxicity in mice. J. Biochem. Mol. Toxicol. 2017, 31, e21974. [Google Scholar] [CrossRef]
- El-Agamy, D.S. Pirfenidone ameliorates concanavalin A-induced hepatitis in mice via modulation of reactive oxygen species/nuclear factor kappa B signalling pathways. J. Pharm. Pharmacol. 2016, 68, 1559–1566. [Google Scholar] [CrossRef]
- Lam, H.Y.P.; Hung, M.Y.; Liang, T.R.; Peng, S.Y. An in-vivo study into the effects of schisandrin b in the liver, spleen, kidney, and brain of acute thioacetamide-intoxicated mice. Iran J. Pharm. Res. 2021, 20, 300–314. [Google Scholar]
- Toprak, T.; Sekerci, C.A.; Aydın, H.R.; Ramazanoglu, M.A.; Arslan, F.D.; Basok, B.I.; Kucuk, H.; Kocakgol, H.; Aksoy, H.Z.; Asci, S.S.; et al. Protective effect of chlorogenic acid on renal ischemia/reperfusion injury in rats. Arch. Ital. Urol. Androl. 2020, 92, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.M.; Elbadawy, H.M.; Ibrahim, S.R.M.; Mohamed, G.A.; Elsaed, W.M.; Alhaddad, A.A.; Ahmed, N.; Abo-Haded, H.; El-Agamy, D.S. Alpha-mangostin as a new therapeutic candidate for concanavalin A-induced autoimmune hepatitis: Impact on the SIRT1/Nrf2 and NF-κB crosstalk. Plants 2022, 11, 2441. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, W.; Simic, N.; Weis, R.; Saf, R.; Kunert, O. Complete assignments of 1H and 13C NMR resonances of oleanolic acid, 18α-oleanolic acid, ursolic acid and their 11-oxo derivatives. Magn. Reason. Chem. 2003, 41, 636–638. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants-rich sources for a new group of multi-potent plant extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [Green Version]
- Ayoub, I.M.; El-Baset, M.A.; Elghonemy, M.M.; Bashandy, S.A.E.; Ibrahim, F.A.A.; Ahmed-Farid, O.A.H.; El Gendy, A.E.G.; Afifi, S.M.; Esatbeyoglu, T.; Farrag, A.R.H.; et al. Chemical profile of Cyperus laevigatus and its protective effects against thioacetamide-induced hepatorenal toxicity in rats. Molecules 2022, 27, 6470. [Google Scholar] [CrossRef] [PubMed]
- Obert, L.A.; Elmore, S.A.; Ennulat, D.; Frazier, K.S. A Review of specific biomarkers of chronic renal injury and their potential application in nonclinical safety assessment studies. Toxicol. Pathol. 2021, 49, 996–1023. [Google Scholar] [CrossRef] [PubMed]
- Al-Attar, A.M.; Alrobai, A.A.; Almalki, D.A. Protective effect of olive and juniper leaves extracts on nephrotoxicity induced by thioacetamide in male mice. Saudi J. Biol. Sci. 2017, 24, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Bashandy, S.A.; El Awdan, S.A.; Mohamed, S.M.; Omara, E.A.A. Allium porrum and Bauhinia variegata mitigate acute liver failure and nephrotoxicity induced by thioacetamide in male rats. Indian J. Clin. Biochem. 2020, 35, 147–157. [Google Scholar] [CrossRef]
- Omar, A.M.S. The potential protective influence of flaxseed oil against renal toxicity induced by thioacetamide in rats. Saudi J. Biol. Sci. 2018, 25, 1696–1702. [Google Scholar] [CrossRef] [Green Version]
- Koen, Y.M.; Sarma, D.; Hajovsky, H.; Galeva, N.A.; Williams, T.D.; Staudinger, J.L.; Hanzlik, R.P. Protein targets of thioacetamide metabolites in rat hepatocytes. Chem. Res. Toxicol. 2013, 26, 64–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, H.N.; El Awdan, S.A.; Hegazy, G.A. Protective role of antioxidants on thioacetamideinduced acute hepatic encephalopathy: Biochemical and ultrastructural study. Tissue Cell. 2013, 45, 350–362. [Google Scholar] [CrossRef]
- de David, C.; Rodrigues, G.; Bona, S.; Meurer, L.; Gonzalez-Gallego, J.; Tunon, M.J.; Marroni, N.P. Role of quercetin in preventing thioacetamide-induced liver injury in rats. Toxicol. Pathol. 2011, 39, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Wang, C.; Li, X.; Liang, G. Oleanolic acid reduces oxidative stress and neuronal apoptosis after experimental subarachnoid hemorrhage by regulating Nrf2/HO-1 pathway. Drug Dev. Res. 2022, 83, 680–687. [Google Scholar] [CrossRef]
- Nyakudya, T.T.; Isaiah, S.; Ayeleso, A.; Ndhlala, A.R.; Mukwevho, E.; Erlwanger, K.H. Short-term neonatal oral administration of oleanolic acid protects against fructose-induced oxidative stress in the skeletal muscles of suckling rats. Molecules 2019, 24, 661. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.B.; Wang, R.X.; Deng, H.J.; Wang, Y.H.; Tang, J.D.; Cao, F.Y.; Wang, J.H. Protective effects of oleanolic acid on oxidative stress and the expression of cytokines and collagen by the AKT/NF-κB pathway in silicotic rats. Mol. Med. Rep. 2017, 15, 3121–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amer, M.G.; Mazen, N.F.; Mohamed, A.M. Caffeine intake decreases oxidative stress and inflammatory biomarkers in experimental liver diseases induced by thioacetamide: Biochemical and histological study. Int. J. Immunopathol Pharmacol. 2017, 30, 13–24. [Google Scholar] [CrossRef]
- Amanzada, A.; Moriconi, F.; Mansuroglu, T.; Cameron, S.; Ramadori, G.; Malik, I.A. Induction of chemokines and cytokines before neutrophils and macrophage recruitment in different regions of rat liver after TAA administration. Lab. Investig. 2014, 94, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Marchyshak, T.; Yakovenko, T.; Shmarakov, I.; Tkachuk, Z. The potential protective effect of oligoribonucleotides-D-mannitol complexes against thioacetamide-induced hepatotoxicity in mice. Pharmaceuticals 2018, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.R.; Lee, J.A.; Kim, M.; Lee, S.; Oh, M.; Moon, J.; Nam, J.-W.; Choi, H.; Mun, Y.-J.; Roh, S.-S. Gardeniae fructus attenuates thioacetamide-induced liver fibrosis in mice via both AMPK/SIRT1/NF-B pathway and Nrf2 signaling. Antioxidants 2021, 10, 1837. [Google Scholar] [CrossRef]
- Bao, J.; Yan, W.; Xu, K.; Chen, M.; Chen, Z.; Ran, J.; Xiong, Y.; Wu, L. Oleanolic acid decreases IL-1β-induced activation of fibroblast-like synoviocytes via the SIRT3-NF-κB Axis in osteoarthritis. Oxid. Med. Cell. Longev. 2020, 2020, 7517219. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.M.; Kim, H.M.; Lee, M.; An, H.J. Oleanolic acid alleviates atopic dermatitis-like responses in vivo and in vitro. Int. J. Mol. Sci. 2021, 22, 12000. [Google Scholar] [CrossRef]
- Peng, X.P.; Li, X.H.; Li, Y.; Huang, X.T.; Luo, Z.Q. The protective effect of oleanolic acid on NMDA-induced MLE-12 cells apoptosis and lung injury in mice by activating SIRT1 and reducing NF-κB acetylation. Int. Immunopharmacol. 2019, 70, 520–529. [Google Scholar] [CrossRef]
- Ghosh, S.; Sarkar, A.; Bhattacharyya, S.; Sil, P.C. Silymarin Protects Mouse Liver and Kidney from Thioacetamide Induced Toxicity by Scavenging Reactive Oxygen Species and Activating PI3K-Akt Pathway. Front. Pharmacol. 2016, 7, 481. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liu, X.; Ding, C.; Gu, Y.; Liu, W. Dihydromyricetin reverses thioacetamide-induced liver fibrosis through inhibiting NF-κB-mediated inflammation and TGF-β1-regulated of PI3K/Akt signaling pathway. Front. Pharmacol. 2021, 12, 783886. [Google Scholar] [CrossRef]
- Shin, M.R.; Lee, S.H.; Roh, S.-S. The Potential hepatoprotective effect of Paeoniae Radix Alba in thioacetamide-induced acute liver injury in rats. Evid. Based Complement. Altern. Med. 2022, 2022, 7904845. [Google Scholar] [CrossRef] [PubMed]
- ElBaset, M.A.; Salem, R.S.; Ayman, F.; Ayman, N.; Shaban, N.; Afifi, S.M.; Esatbeyoglu, T.; Abdelaziz, M.; Elalfy, Z.S. Effect of empagliflozin on thioacetamide-induced liver injury in rats: Role of AMPK/SIRT-1/HIF-1α pathway in halting liver fibrosis. Antioxidants 2022, 11, 2152. [Google Scholar] [CrossRef]
- Hong, Y.A.; Lim, J.H.; Kim, M.Y.; Kim, E.N.; Koh, E.S.; Shin, S.J.; Choi, B.S.; Park, C.W.; Chang, Y.S.; Chung, S. Delayed treatment with oleanolic acid attenuates tubulointerstitial fibrosis in chronic cyclosporine nephropathy through Nrf2/HO-1 signaling. J. Transl. Med. 2014, 12, 50. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, D.K.; Jena, G.; Kumar, V. Dimethyl fumarate protects thioacetamide-induced liver damage in rats: Studies on Nrf2, NLRP3, and NF-κB. J. Biochem. Mol. Toxicol. 2020, 34, e22476. [Google Scholar] [CrossRef]
- Hafez, H.M.; Ibrahim, M.A.; Zedan, M.Z.; Hassan, M.; Hassanein, H. Nephroprotective effect of cilostazol and verapamil against thioacetamide-induced toxicity in rats may involve Nrf2/HO-1/NQO-1 signaling pathway. Toxicol. Mech. Methods. 2019, 29, 146–152. [Google Scholar] [CrossRef]
- Hussein, R.M.; Sawy, D.M.; Kandeil, M.A.; Farghaly, H.S. Chlorogenic acid, quercetin, coenzyme Q10 and silymarin modulate Keap1-Nrf2/heme oxygenase-1 signaling in thioacetamide-induced acute liver toxicity. Life Sci. 2021, 277, 119460. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Yoon, H.E.; Kim, S.J.; Kim, S.J.; Koh, E.S.; Hong, Y.A.; Park, C.W.; Chang, Y.S.; Shin, S.J. Oleanolic acid attenuates renal fibrosis in mice with unilateral ureteral obstruction via facilitating nuclear translocation of Nrf2. Nutr. Metab. 2014, 11, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Mouse) | Sequence (5′-3′) | Product (bp) |
|---|---|---|
| IL-1β | F: GCAACTGTTCCTGAACTCAACT | 81 |
| R: GGGTCCGTCAACTTCAAAGA | ||
| IL-6 | F: AGTCCTTCCTACCCCAATTTCC | 79 |
| R: GGTCTTGGTCCTTAGCCACT | ||
| TNF-α | F: TGAACTTCGGGGTGATCGGT | 99 |
| R: GGTGGTTTGTGAGTGTGAGGG | ||
| Bax | F: TGAAGACAGGGGCCTTTTTG | 140 |
| R: AATTCGCCGGAGACACTCG | ||
| Bcl2 | F: CCTGTGGATGACTGAGTACCTG | 123 |
| R: AGCCAGGAGAAATCAAACAGAGG | ||
| Caspase-3 | F: ATGGAGAACAACAAAACCTCAGT | 74 |
| R: TTGCTCCCATGTATGGTCTTTAC | ||
| SIRT1 | F: CGATGACAGAACGTCACACG | 111 |
| R: ATTGTTCGAGGATCGGTGCC | ||
| Nrf2 | F: AAGAATAAAGTCGCCGCCCA | 170 |
| R: AGATACAAGGTGCTGAGCCG | ||
| HO-1 | F: GAAATCATCCCTTGCACGCC | 122 |
| R: CCTGAGAGGTCACCCAGGTA | ||
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | F: AGGTCGGTGTGAACGGATTTG | 123 |
| R: TGTAGACCATGTAGTTGAGGTCA |
| Serum Parameters | Groups | ||||
|---|---|---|---|---|---|
| Control | OA | TAA | OA 45 + TAA | OA 90 + TAA | |
| ALT (IU/L) | 38 ± 4.8 | 33.6 ± 3.3 | 456 ± 26.1 * | 290 ± 25 *## | 159.3 ± 8.3 *## |
| AST (IU/L) | 49.2 ± 3.8 | 51 ± 6 | 699 ± 33.1 * | 419.8 ± 19.4 *## | 260 ± 27.6 *## |
| ALP (IU/L) | 37.2 ± 2.9 | 33.4 ± 3.1 | 535.7 ± 51.6 * | 336 ± 40.3 *## | 252.8 ± 25.1 *## |
| LDH (IU/L) | 65.1 ± 10.6 | 73.8 ± 9.7 | 945.8 ± 70.5 * | 500.8 ± 44.7 *## | 350.5 ± 23.9 *## |
| BUN (mg/dL) | 20.7 ± 1.5 | 24.2 ± 1.9 | 48.6 ± 3 * | 37.8 ± 3.3 *# | 28.2 ± 2.4 *## |
| Creatinine (mg/dL) | 0.9 ± 0.07 | 0.9 ± 0.07 | 2.5 ± 0.1 * | 1.8 ± 0.1 *## | 1.5 ± 0.09 *## |
| Parameters | Groups | ||||
|---|---|---|---|---|---|
| Control | OA | TAA | OA 45 + TAA | OA 90 + TAA | |
| MDA (nmol/g tissue) | |||||
| Liver | 23.4 ± 1.9 | 20.9 ± 1.4 | 67.4 ± 5.2 *** | 43.8 ± 2.7 ***## | 28.5 ± 2.2 ### |
| Kidney | 20.5 ± 2.5 | 18.7 ± 1.3 | 72.3 ± 5.9 *** | 50.9 ± 3.7 ***## | 33.6 ± 4.1 *### |
| 4-HNE (µg/mL) | |||||
| Liver | 0.27 ± 0.02 | 0.3 ± 0.03 | 1.2 ± 0.05 *** | 0.8 ± 0.04 ***## | 0.4 ± 0.03 ### |
| Kidney | 0.36 ± 0.04 | 0.04 ± 0.04 | 1.3 ± 0.1 *** | 0.9 ± 0.07 ***## | 0.58 ± 0.05 *### |
| NOx (µM/L) | |||||
| Liver | 35.8 ± 4.2 | 30.2 ± 2.8 | 237.2 ± 11.6 *** | 120.2 ± 9.8 ***### | 97.5 ± 4.3 **### |
| Kidney | 48.9 ± 3.1 | 39.7 ± 3.0 | 256.3 ± 16.2 *** | 142.1 ± 11.5 ***### | 119.2 ± 7.2 **### |
| GSH (µmol/g tissue) | |||||
| Liver | 16.7 ± 1.0 | 17.5 ± 1.5 | 5.3 ± 0.6 *** | 7.8 ± 0.4 ***# | 13.9 ± 1.2 ### |
| Kidney | 15.4 ± 1.2 | 17.2 ± 1.3 | 3.2 ± 0.2 *** | 8.5 ± 0.6 ***### | 14.3 ± 0.9 ### |
| SOD (Unit/g tissue) | |||||
| Liver | 26.5 ± 2.1 | 27.55 ± 1.4 | 12.4 ± 1.2 *** | 18.6 ± 1.2 ***## | 20.6 ± 1.6 ### |
| Kidney | 24.2 ± 1.4 | 28.2 ± 1.2 | 11.8 ± 1.1 *** | 17.2 ± 0.9 ***## | 22.4 ± 2.1 ### |
| TAC (mmol/g tissue) | |||||
| Liver | 0.94 ± 0.03 | 0.9 ± 0.07 | 0.4 ± 0.03 *** | 0.55 ± 0.04 ***# | 0.75 ± 0.05 ### |
| Kidney | 0.88 ± 0.05 | 0.8 ± 0.04 | 0.3 ± 0.02 *** | 0.43 ± 0.02 ***# | 0.66 ± 0.04 ### |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqrad, M.A.I.; El-Agamy, D.S.; Ibrahim, S.R.M.; Sirwi, A.; Abdallah, H.M.; Abdel-Sattar, E.; El-Halawany, A.M.; Elsaed, W.M.; Mohamed, G.A. SIRT1/Nrf2/NF-κB Signaling Mediates Anti-Inflammatory and Anti-Apoptotic Activities of Oleanolic Acid in a Mouse Model of Acute Hepatorenal Damage. Medicina 2023, 59, 1351. https://doi.org/10.3390/medicina59071351
Alqrad MAI, El-Agamy DS, Ibrahim SRM, Sirwi A, Abdallah HM, Abdel-Sattar E, El-Halawany AM, Elsaed WM, Mohamed GA. SIRT1/Nrf2/NF-κB Signaling Mediates Anti-Inflammatory and Anti-Apoptotic Activities of Oleanolic Acid in a Mouse Model of Acute Hepatorenal Damage. Medicina. 2023; 59(7):1351. https://doi.org/10.3390/medicina59071351
Chicago/Turabian StyleAlqrad, Manea A. I., Dina S. El-Agamy, Sabrin R. M. Ibrahim, Alaa Sirwi, Hossam M. Abdallah, Essam Abdel-Sattar, Ali M. El-Halawany, Wael M. Elsaed, and Gamal A. Mohamed. 2023. "SIRT1/Nrf2/NF-κB Signaling Mediates Anti-Inflammatory and Anti-Apoptotic Activities of Oleanolic Acid in a Mouse Model of Acute Hepatorenal Damage" Medicina 59, no. 7: 1351. https://doi.org/10.3390/medicina59071351