Current Research on the Safety of Pyrethroids Used as Insecticides

 , ,
, ,
Abstract
:1. Introduction
2. Permethrin
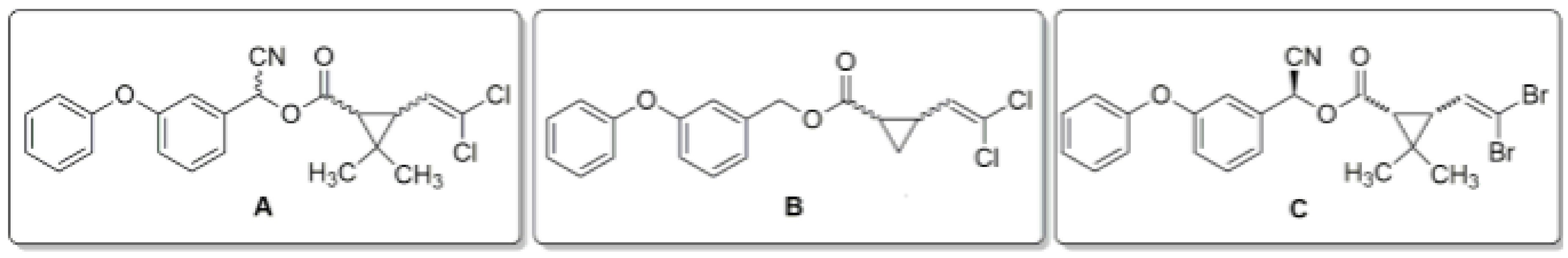
2.1. Chemical Properties and Application
2.2. Mechanism of Action
2.3. Animals Studies
2.4. Human Studies
3. Deltamethrin
3.1. Chemical Properties and Application
3.2. Mechanism of Action
3.3. Animal Studies
3.4. Human Studies
4. Alpha-Cypermethrin
4.1. Chemical Properties and Application
4.2. Mechanism of Action
4.3. Animal Studies
4.4. Human Studies
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Bradberry, S.M.; Cage, S.A.; Proudfoot, A.T.; Vale, J.A. Poisoning due to pyrethroids. Toxicol. Rev. 2005, 24, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G. The neurotoxicity of organochlorine and pyrethroid pesticides. Handb. Clin. Neurol. 2015, 131, 135–148. [Google Scholar] [PubMed]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity. Recent Adv. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Cárcamo, J.G.; Aguilar, M.N.; Carreño, C.F.; Vera, T.; Arias-Darraz, L.; Figueroa, J.E.; Romero, A.P.; Alvarez, M.; Yañez, A.J. Consecutive emamectin benzoate and deltamethrin treatments affect the expressions and activities of detoxification enzymes in the rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 191, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Toynton, K.; Luukinen, B.; Buhl, K.; Stone, D. Permethrin Technical Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services: Baker City, OR, USA, 2009. [Google Scholar]
- Skolarczyk, J.; Pekar, J.; Nieradko-Iwanicka, B. Immune disorders induced by exposure to pyrethroid insecticides. Postepy Hig. Med. Dośw. 2017, 71, 446–453. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Pesticide Evaluation Scheme, Vector Ecology and Management; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Singh, A.K.; Tiwari, M.N.; Prakash, O.; Singh, M.P. A current review of cypermethrin-induced neurotoxicity and nigrostriatal dopaminergic neurodegeneration. Curr. Neuropharmacol. 2012, 10, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Burridge, L.E.; Haya, K. Lethality of pyrethrins to larvae and postlarvae of the American lobster (Homarus americanus). Ecotoxicol. Environ. Saf. 1997, 38, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Lidova, J.; Stara, A.; Kouba, A.; Velisek, J. The effects of cypermethrin on oxidative stress and antioxidant biomarkers in marbled crayfish (Procambarus fallax f. virginalis). Neuro Endocrinol. Lett. 2016, 37 (Suppl. 1), 53–59. [Google Scholar] [PubMed]
- Lutnicka, H.; Kozińska, A. Pyrethroids as a predisposing factor in fish diseases. Ochr. Środ. Zasobów Nat. 2009, 41, 285–292. [Google Scholar]
- Wang, Y.; Lv, L.; Yu, Y.; Yang, G.; Xu, Z.; Wang, Q.; Cai, L. Single and joint toxic effects of five selected pesticides on the early life stages of zebrafish (Denio renio). Chemosphere 2017, 170, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Orsborne, J.; DeRaedt, B.S.; Hendy, A.; Gezan, S.; Kaur, H.; Wilder-Smith, A.; Lindsay, S.W.; Logan, J. Personal protection of permethrin-treated clothing against Aedes aegypti, the vector of Dengue and Zika virus, in the Laboratory. PLoS ONE 2016, 11, e0152805. [Google Scholar] [CrossRef] [PubMed]
- Ranjkesh, M.R.; Naghili, B.; Goldust, M.; Rezaee, E. The efficacy of permethrin 5% vs. oral ivermectin for the treatment of scabies. Ann. Parasitol. 2013, 59, 189–194. [Google Scholar] [PubMed]
- Hughes, E.A.; Flores, A.P.; Romero, L.M.; Zaits, A.; Glass, C.R.; Montserrat, J.M. Potential dermal exposure to deltamethrin and risk assessment for manual sprayers: Influence of crop type. Sci. Total Environ. 2008, 391, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Singleton, S.T.; Lein, P.J.; Farahat, F.M.; Farahat, T.; Bonner, M.R.; Knaak, J.B.; Olson, J.R. Characterization of α-cypermethrin exposure in Egyptian agricultural workers. Int. J. Hyg. Environ. Health 2014, 217, 538–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, M.F.; Edwards, B.C. In vitro dermal absorption of pyrethroid pesticides in human and rat skin. Toxicol. Appl. Pharmacol. 2010, 246, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.F.; Edwards, B.C. In vivo dermal absorption of pyrethroid pesticides in the rat. J. Toxicol. Environ. Health 2016, 79, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Glorennec, P.; Serrano, T.; Fravallo, M.; Warembourg, C.; Monfort, C.; Cordier, S.; Viel, J.; Le Gléau, F.; Le Bot, B.; Chevrier, C. Determinants of children’s exposure to pyrethroid insecticides in western France. Environ. Int. 2017, 104, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Del Prado-Lu, J.L. Insecticide residues in soil, water, and eggplant fruits and farmers’ health effects due to exposure to pesticides. Environ. Health Prev. Med. 2015, 20, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Nasuti, C.; Fattoretti, P.; Carloni, M.; Fedeli, D.; Ubaldi, M.; Ciccocioppo, R.; Gabbianelli, R. Neonatal exposure to permethrin pesticide causes lifelong fear and spatial learning deficits and alters hippocampal morphology of synapses. J. Neurodev. Dis. 2014, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasuti, C.; Carloni, M.; Fedeli, D.; Gabbianelli, R.; Di Stefano, A.; Serafina, C.; Silva, I.; Dominques, V.; Ciccocioppo, R. Effects of early life permethrin exposure on spatial working memory and on monoamine levels in different brain areas of pre-senescent rats. Toxicology 2013, 303, 162–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, B.J.; Hauptman, M.; Woolf, A.D.; Goldman, R.H. Insect Repellants during Pregnancy in the Era of the Zika Virus. Obstet. Gynecol. 2016, 128, 1111–1115. [Google Scholar] [CrossRef] [PubMed]
- Central Disease Control. CDC Zika Interim Response Plan; Central Disease Control: Atlanta, GA, USA, 2017.
- Sundaramoorthy, R.; Velusamy, Y.; Balaji, A.P.; Mukherjee, A.; Chandrasekaran, N. Comparative cytotoxic and genotoxic effects of permethrin and its nanometric form on human erythrocytes and lymphocytes in vitro. Chem. Biol. Interact. 2016, 257, 119–124. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Specifications and Evaluations for Public Health Pesticides. Permethrin (25:75 Cis:Trans Isomer Ratio, Nonracemic) 3-Phenoxybenzyl (1RS,3RS;1RS,3SR)-3-(2,2 dichlorovinyl)2,2-dimethyl-cyclopropane Carboxylate; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization (WHO). Deltamethrin Long-Lasting (Coated onto Filaments) Insecticidal Net. (s)α-Cyano-3-phenoxybenzyl (1r,3r)-3-(2,2dibromovinyl)-2,2-dimethylcyclopropane Carboxylate; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization (WHO). Specifications and Evaluations for Public Health Pesticides Alpha-Cypermethrin Long-Lasting (Incorporated into Filaments) Insecticidal Net. A Racemic Mixture of: (S)-α-Cyano-3-phenoxybenzyl-(1R,3R)-3-(2,2-dichlorovinyl)2,2-dimethylcyclopropane-carboxylate and (R)-α-Cyano-3-phenoxybenzyl-(1S,3S)-3-(2,2-dichlorovinyl)2,2-dimethylcyclopropane-carboxylate; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Hossain, M.M.; Liu, J.; Richardson, J.R. Pyrethroid insecticides directly activate microglia through interaction with voltage-gated sodium channels. Toxicol. Sci. 2017, 155, 112–123. [Google Scholar] [CrossRef] [PubMed]
- DeGroot, W.D. Intravenous lipid emulsion for treating permethrin toxicosis in a cat. Can. Vet. J. 2014, 55, 1253–1254. [Google Scholar] [PubMed]
- Shafer, T.J.; Rijal, S.O.; Gross, G.W. Complete inhibition of spontaneous activity in neuronal networks in vitro by deltamethrin and permethrin. NeuroToxicology 2008, 29, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Dohlman, T.M.; Phillips, P.E.; Madson, D.M.; Clark, C.A.; Gunn, P.J. Effects of label dose permethrin administration in yearling beef cattle: I. Bull reproductive function and testicular histopathology. Theriogenology 2016, 85, 1534–1539. [Google Scholar] [CrossRef] [PubMed]
- Omotoso, G.O.; Onanuga, I.O.; Ibrahim, R.B. Histological effects of permethrin insecticide on the testis of adult wistar rats. J. Med. Biomed. Sci. 2014, 6, 125–129. [Google Scholar] [CrossRef]
- Kotila, T.; Yön, N.D. The effects of permethrin on rat ovarian tissue morphology. Exp. Toxicol. Pathol. 2015, 67, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Vadhana, M.S.; Carloni, M.; Nasuti, C.; Fedeli, D.; Gabbianelli, R. Early life permethrin insecticide treatment leads to heart damage in adult rats. Exp. Gerontol. 2011, 46, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Vadhana, M.S.; Arumugam, S.S.; Carloni, M.; Nasuti, C.; Gabbianelli, R. Early life permethrin treatment leads to long-term cardiotoxicity. Chemosphere 2013, 93, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Fedeli, D.; Carloni, M.; Nasuti, C.; Gambini, A.; Scocco, V.; Gabbianelli, R. Early life permethrin exposure leads to hypervitaminosis D, nitric oxide and catecholamines impairment. Pestic. Biochem. Physiol. 2013, 107, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Pratera, M.R.; Gogal Jra, R.M.; Blaylockb, B.L.; Longstrethc, J.; Holladaya, S.D. Single-dose topical exposure to the pyrethroid insecticide, permethrin in C57BL/6N mice: Effects on thymus and spleen. Food Chem. Toxicol. 2002, 40, 1863–1873. [Google Scholar] [CrossRef]
- Gabbianelli, R.; Falcioni, M.L.; Nasuti, C.; Cantalamessa, F.; Imada, I.; Inoue, M. Effect of permethrin insecticide on rat polymorphonuclear neutrophils. Chem. Biol. Interact. 2009, 182, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Carloni, M.; Nasuti, C.; Fedeli, D.; Montani, M.; Amici, A.; Vadhana, M.S.; Gabbianelli, R. The impact of early life permethrin exposure on development of neurodegeneration in adulthood. Exp. Gerontol. 2012, 47, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, L.; Nasuti, C.; Mirto, M.; Caradonna, F.; Gabbianelli, R. Intergenerational Effect of early life exposure to Permethrin: Changes in global DNA methylation and in Nurr1 Gene Expression. Toxics 2015, 3, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goksugur, S.B.; Karatas, Z.; Goksugur, N.; Bekdas, M.; Demircioglu, F. Metabolic acidosis in an infant associated with permethrin toxicity. Pediatr. Dermatol. 2015, 32, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Corcellas, C.; Feo, M.L.; Torres, J.P.; Malm, O.; Ocampo-Duque, W.; Eljarrat, E.; Barceló, D. Pyrethroids in human breast milk: Occurrence and nursing daily intake estimation. Environ. Int. 2014, 47, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Outhlote, Y.; Bouchard, M. Urinary metabolities of organophosphate and pyrethroid pesticides and behavioral problems in Canadian children. Environ. Health Perspect. 2013, 121, 1378–1384. [Google Scholar]
- Stein, E.A.; Washburn, M.; Walczak, C.; Bloom, A.S. Effects of pyrethroid insecticides on operant responding maintained by food. Neurotoxicol. Teratol. 1987, 9, 27–31. [Google Scholar] [CrossRef]
- Johnson, M.; Luukinen, B.; Buhl, K.; Stone, D. Deltamethrin Technical Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services: Baker City, OR, USA, 2010. [Google Scholar]
- Ding, R.; Cao, Z.; Wang, Y.; Gao, X.; Luo, H.; Zhang, C.; Ma, S.; Jin, H.; Lu, C. The implication of p66shc in oxidative stress induced by deltamethrin. Chem. Biol. Interact. 2017, 278, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Thomas, A.; Pillai, M. Deltamethrin: Promising mosquito control agent against adult stage of Aedes aegypti L. Asian Pac. J. Trop. Med. 2011, 4, 430–435. [Google Scholar] [CrossRef]
- Hołyńska-Iwan, I.; Bogusiewicz, J.; Chajdas, D.; Szewczyk-Golec, K.; Lampka, M.; Olszewska-Słonina, D. The immediate influence of deltamethrin on ion transport through rabbit skin. An in vitro study. Pestic. Biochem. Physiol. 2018, 148, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Khalatbary, A.; Ghaffari, E.; Iranian, B. Protective role of oleuropein against acute deltamethrin-induced neurotoxicity in rat brain. Iran. Biomed. J. 2015, 19, 247–253. [Google Scholar] [PubMed]
- Kumar, A.; Sasmal, D.; Bhaskar, A.; Mukhopadhyay, K.; Thakur, A.; Sharma, N. Deltamethrin-induced oxidative stress and mitochondrial caspase-dependent signaling pathways in murine splenocytes. Environ. Toxicol. 2014, 31, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sharma, N. Comparative efficacy of piperine and curcumin in deltamethrin induced splenic apoptosis and altered immune functions. Pestic. Biochem. Physiol. 2015, 119, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sasmal, D.; Sharma, N. Mechanism of deltamethrin induced thymic and splenic toxicity in mice and its protection by piperine and curcumin: In vivo study. Drug Chem. Toxicol. 2018, 41, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Gündüz, E.; Ülger, B.V.; İbiloğlu, I.; Ekinci, A.; Dursun, R.; Zengin, Y.; İçe, M.; Uslukaya, Ö.; Ekinci, C.; Güloğlu, C. Glutamine provides effective protection against deltamethrin-induced acute hepatotoxicity in rats but not against nephrotoxicity. Med. Sci. Monit. 2015, 21, 1107–1114. [Google Scholar] [PubMed]
- Abdel-Daim, M.M.; Abuzead, S.M.; Halawa, S.M. Protective role of Spirulina platensis against acute deltamethrin-induced toxicity in rats. PLoS ONE 2013, 8, e72991. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; El-Ghoneimy, A. Synergistic protective effects of ceftriaxone and ascorbic acid against subacute deltamethrin induced nephrotoxicity in rats. Ren. Fail. 2015, 37, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Maalej, A.; Mahmoudi, A.; Bouallagui, Z.; Fki, I.; Marrekchi, R.; Sayadi, S. Olive phenolic compounds attenuate deltamethrin-induced liver and kidney toxicity through regulating oxidative stress, inflammation and apoptosis. Food Chem. Toxicol. 2017, 106, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Ogaly, H.A.; Khalaf, A.A.; Ibrahima, M.A.; Galal, M.K.; Abd-Elsalam, R.M. Influence of green tea extract on oxidative damage and apoptosis induced by deltamethrin in rat brain. Neurotoxicol. Teratol. 2015, 50, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Rjeibia, I.; Saada, A.B.; Hfaiedh, N. Oxidative damage and hepatotoxicity associated with deltamethrin in rats: The protective effects of Amaranthus spinosus seed extract. Biomed. Pharmacother. 2016, 84, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Oda, S.S.; El-Maddawy, Z. Protective effect of vitamin E and selenium combination on deltamethrin-induced reproductive toxicity in male rats. Exp. Toxicol. Pathol. 2012, 64, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Chou, C.; Liang, W.; Jan, C. Effect of the pesticide, deltamethrin, on Ca2+ signaling and apoptosis in OC2 human oral cancer cells. Drug Chem. Toxicol. 2014, 37, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, I.; Oose, K.; Otaki, Y.; Ihara, D.; Fukuchi, M.; Tabuchi, A.; Tsuneki, H.; Tabuchi, Y.; Kondo, T.; Saitoh, A.; et al. Type II pyrethroid deltamethrin produces antidepressant-like effects in mice. Behav. Brain Res. 2013, 257, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Shujie, W.; Qingland, Z.; Lan, Y.; Bohong, X.; Yurui, I. Health survey among farmers exposed to deltamethrin in the cotton fields. Ecotoxicol. Environ. Safe 1988, 15, 1–6. [Google Scholar] [CrossRef]
- Kim, K.B.; Anand, S.; Kim, H.; White, C.; Bruckner, J. Toxicokinetics and tissue distribution of deltamethrin in adult Sprague-Dawley rats. Toxicol. Sci. 2008, 101, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Viel, J.F.; Warembourg, C.; Le Mauer-Idrissi, G.; Lacroix, A.; Limon, G.; Rouget, F.; Monfort, C.; Durand, G.; Cordier, S.; Cherier, C. Pyrethroid insecticide exposure and cognitive developmental disabilities in children: The PELAGIE mother-child cohort. Environ. Int. 2015, 82, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, R.; Husain, R.; Adhami, V.M.; Seth, P.K. Behavioral, neurochemical, and neuromorphological effects of deltamethrin in adult rats. J. Toxicol. Environ. Health 1996, 48, 515–526. [Google Scholar] [PubMed]
- Elwan, M.A.; Richardson, J.R.; Guillot, T.S.; Caudle, W.M.; Miller, G.W. Pyrethroid pesticide-induced alterations in dopamine transporter function. Toxicol. Appl. Pharmacol. 2006, 211, 188–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Okda, E.; Abdel-Hamid, M.A.; Hamdy, A.M. Immunological and genotoxic effects of occupational exposure to α-cypermethrin pesticide. Int. J. Occup. Med. Environ. Health 2017, 30, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, J.; Subdys, J. Alfa-cypermetryna—Aerozole, mieszanina izomerów. Dokumentacja proponowanych wartości dopuszczalnych wielkości narażenia zawodowego. Podstawy i Metody Oceny Środowiska Pracy 2005, 3, 19–36. [Google Scholar]
- Costa, C.; Rapisarda, V.; Catania, S.; DiNola, C.; Ledda, C.; Fenga, C. Cytokine patterns in green house workers occupationally exposed to-cypermethrin: An observational study. Environ. Toxicol. Pharmacol. 2013, 36, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Hocine, L.; Merzouk, H.; Merzouk, S.A.; Ghorzi, H.; Youbi, M.; Narce, M. The effects of alpha-cypermethrin exposure on biochemical and redox parameters in pregnant rats and their newborns. Pestic. Biochem. Physiol. 2016, 134, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Laugeray, A.; Herzine, A.; Perche, O.; Richard, O.; Montecot-Dubourg, C.; Menuet, A.; Mazaud-Guittlot, S.; Lesne, L.; Jeguo, B.; Mortaud, S. In utero and lactational exposure to low-doses of the pyrethroid insecticide cypermethrin leads to neurodevelopmental defects in male mice—An ethological and transcriptomic study. PLoS ONE 2017, 12, e0184475. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Ramos, E.; Ares, I.; Castellano, V.; Martínez, M.; Rosa, M.; Larranaga, M.; Anadón, A.; Martínez, M.A. Oxidative stress and gene expression profiling of cell death pathways in α-cypermethrin treated SH SY5Y cells. Arch. Toxicol. 2017, 91, 2151–2164. [Google Scholar] [CrossRef] [PubMed]
- Motsoeneng, P.M.; Dalvie, M.A. Relationship between Urinary Pesticide Residue Levels and Neurotoxic Symptoms among Women on Farms in the Western Cape, South Africa. Int. J. Environ. Res. Public Health 2015, 12, 6281–6299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S.; Bhattacharyya, D.; Mandal, T.K.; Dey, S. Neuropharmacological effects of alpha-cypermethrin in rats. Indian J. Pharmacol. 2005, 37, 18–20. [Google Scholar] [CrossRef]
- Poulos, L.; Athanaselis, S.; Coutselinis, A. Acute intoxication with cypermethrin (NRDC 149). J. Toxicol. Clin. Toxicol. 1982, 19, 519–520. [Google Scholar] [CrossRef] [PubMed]
- European Union Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 Establishing a Framework for Community. 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32009L0128 (accessed on 27 August 2018).


{kind=link}
| Pyrethroid/Group | Half-Life in the Body | Excreted Substances | Toxic Dose for Humans | Quantity of Active Substance | Negative Effects in Humans |
|---|---|---|---|---|---|
| Permethrin I (T) | 12.3 h in plasma 9–23 in neuronal tissue | cis-DCCA trans- DCCA 3-PBA | <0.25 mg/kg/day (RfD) >50 mg/kg | 3.75–7.5% | Reproductive system Skeletal system Cardiovascular system Immune system Neuronal system |
| Deltamethrin II (CS) | 11.5 h in plasma | 3-PBA cis-DBCA | <0.01 mg/kg/day (RfD) >30 mg/kg | 0.25–0.5% | Immune system Reproductive system Neuronal system Skeletal muscles Inducing oxidative stress |
| α-cypermethrin II (CS) | 2.5 days in fat tissue | F-PBA CPA | <0.05 mg/kg/day (RfD) >50 mg/kg | 0.25–0.5% | Immune system Reproductive system Neuronal system Inducing apoptosis in SH-SY5Y line cells |
| Pyrethroid/Group | Routes of Exposure | Sources (Pyrethroid Concentration) |
|---|---|---|
| Permethrin I (T) | Skin | Topical creams for scabies (5%). Flea/tick medicines for pets (25–75%) |
| Respiratory tract | Mosquito nets (0.25–1%) | |
| Deltamethrin II (CS) | Skin | The highest accumulation should be expected in: cotton, coffee, hops, artichokes, corn, broccoli, apples, plums and grain (1–2.5%) Flea/tick medicines for pets (0.25–10%) |
| Gastrointestinal tract | The highest accumulation should be expected in hops, artichokes, broccoli and grain | |
| Respiratory tract | Mosquito nets (0.25–1%) | |
| α-cypermethrin II (CS) | Skin | Crop protection chemicals—cotton, rice, potatoes, citrus fruits, grapes and soy (1–2.5%) Flea/tick medicines for pets (3–5%) |
| Gastrointestinal tract | The highest accumulation should be expected in soy, citrus fruits and grapes | |
| Respiratory tract | Mosquito nets (6%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrustek, A.; Hołyńska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A.; Olszewska-Słonina, D. Current Research on the Safety of Pyrethroids Used as Insecticides. Medicina 2018, 54, 61. https://doi.org/10.3390/medicina54040061
Chrustek A, Hołyńska-Iwan I, Dziembowska I, Bogusiewicz J, Wróblewski M, Cwynar A, Olszewska-Słonina D. Current Research on the Safety of Pyrethroids Used as Insecticides. Medicina. 2018; 54(4):61. https://doi.org/10.3390/medicina54040061
Chicago/Turabian StyleChrustek, Agnieszka, Iga Hołyńska-Iwan, Inga Dziembowska, Joanna Bogusiewicz, Marcin Wróblewski, Anna Cwynar, and Dorota Olszewska-Słonina. 2018. "Current Research on the Safety of Pyrethroids Used as Insecticides" Medicina 54, no. 4: 61. https://doi.org/10.3390/medicina54040061