
Molecular Cloning and Characteristics of a Lectin from the Bivalve Glycymeris yessoensis
 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
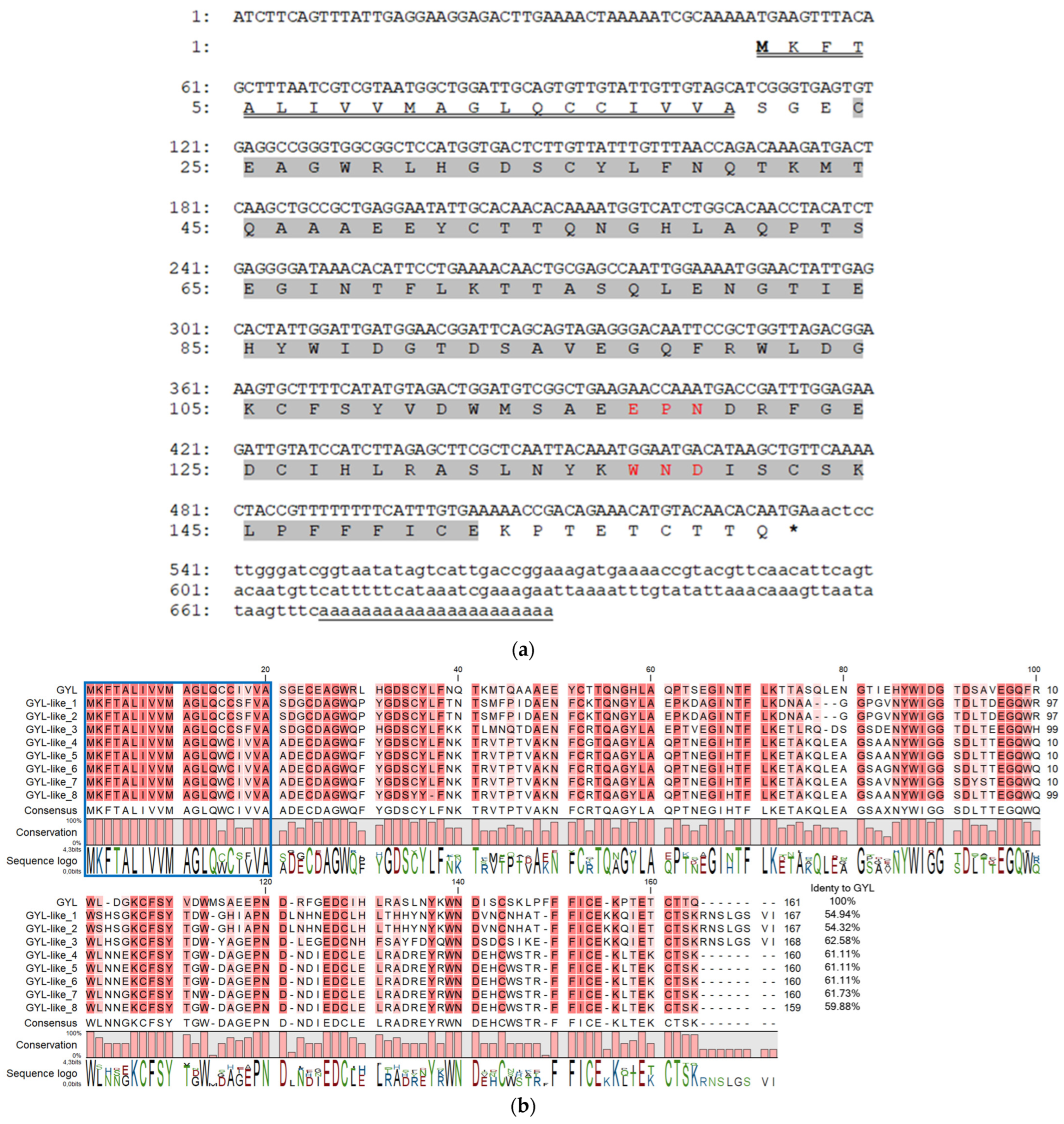
2.1. Determination of a Full-Length Sequence of GYL and Analysis of Its Isoforms in the Bivalve G. yessoensis
2.2. Characteristics of GYL
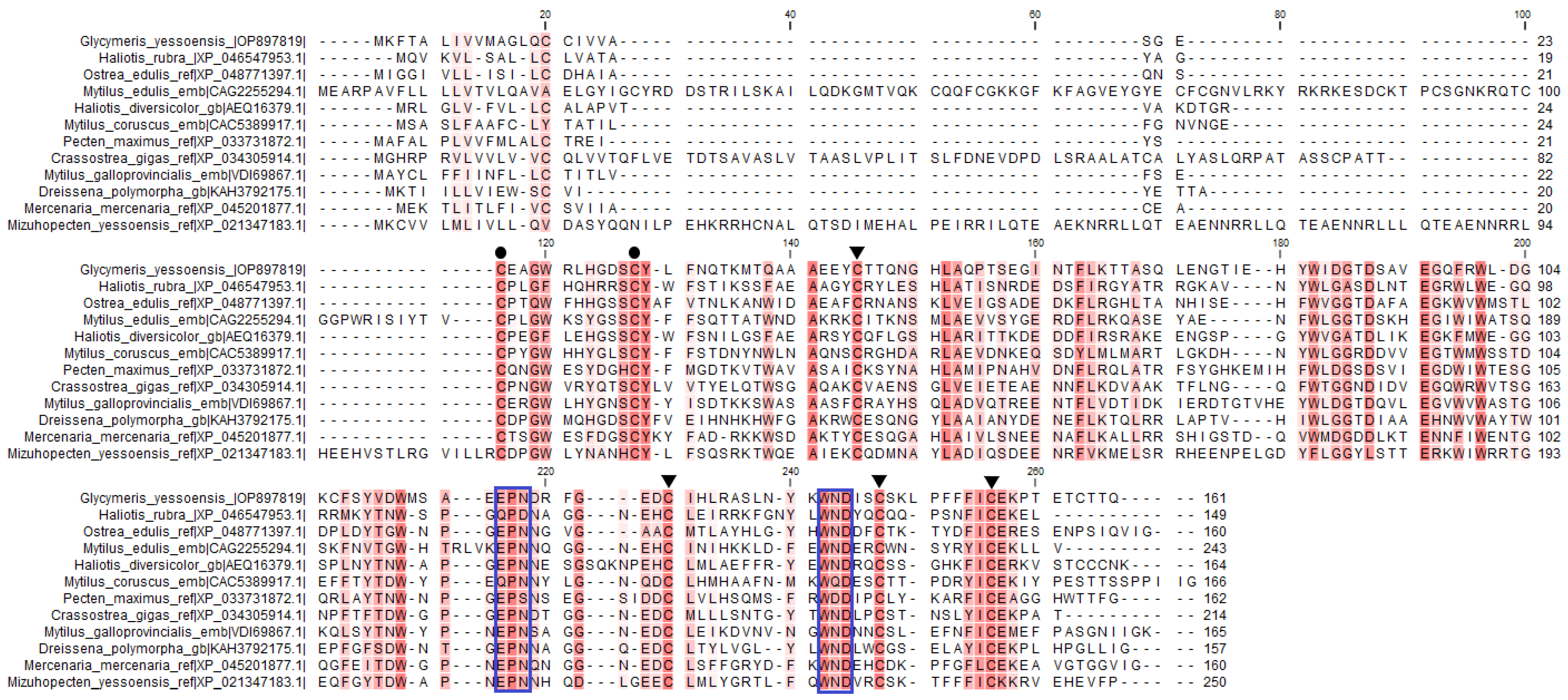
2.2.1. Homologous Analysis of GYL
2.2.2. Signal Peptide of the GYL Sequence
2.2.3. N-Glycosylation of GYL
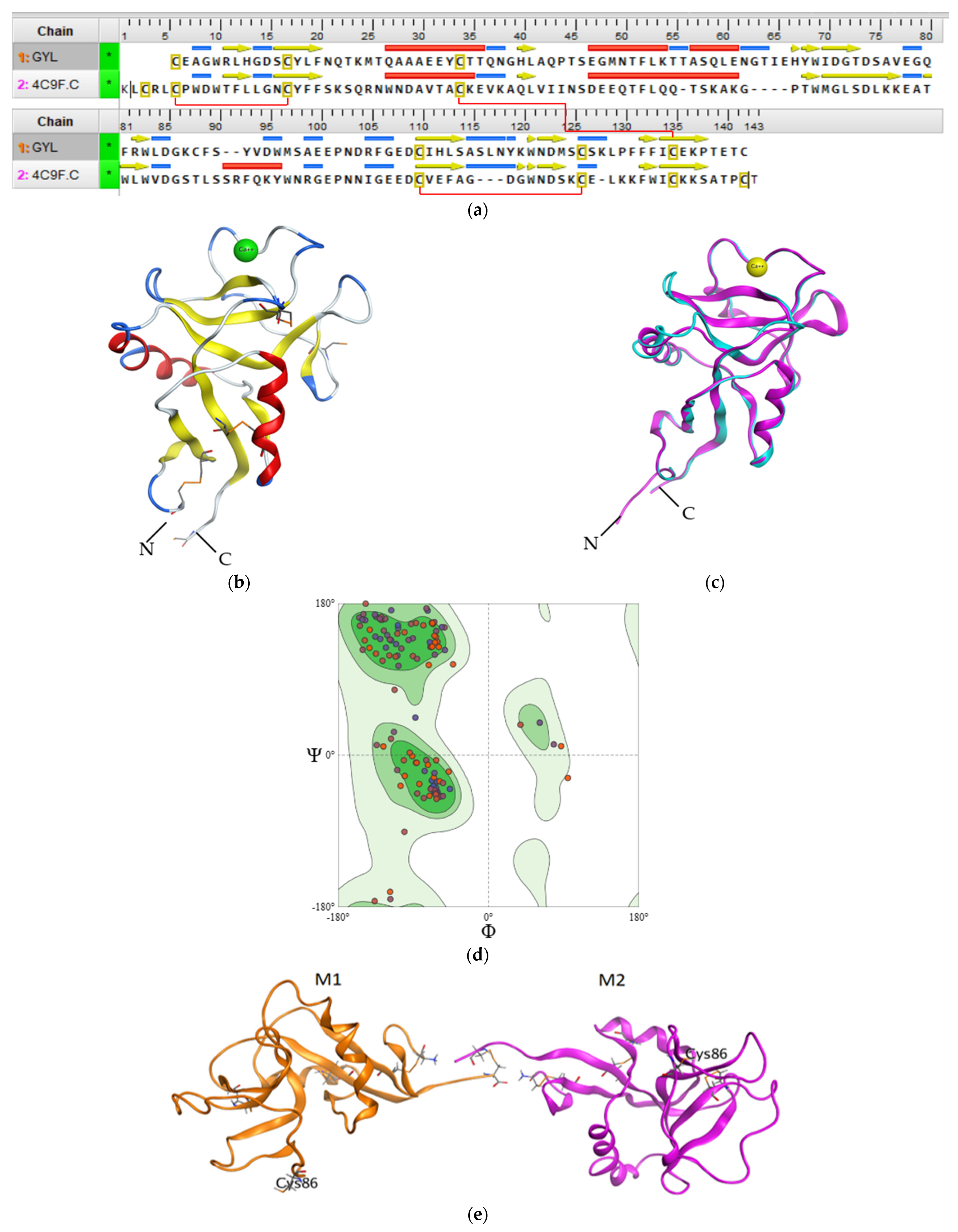
2.2.4. The Potential Tertiary Structure of GYL
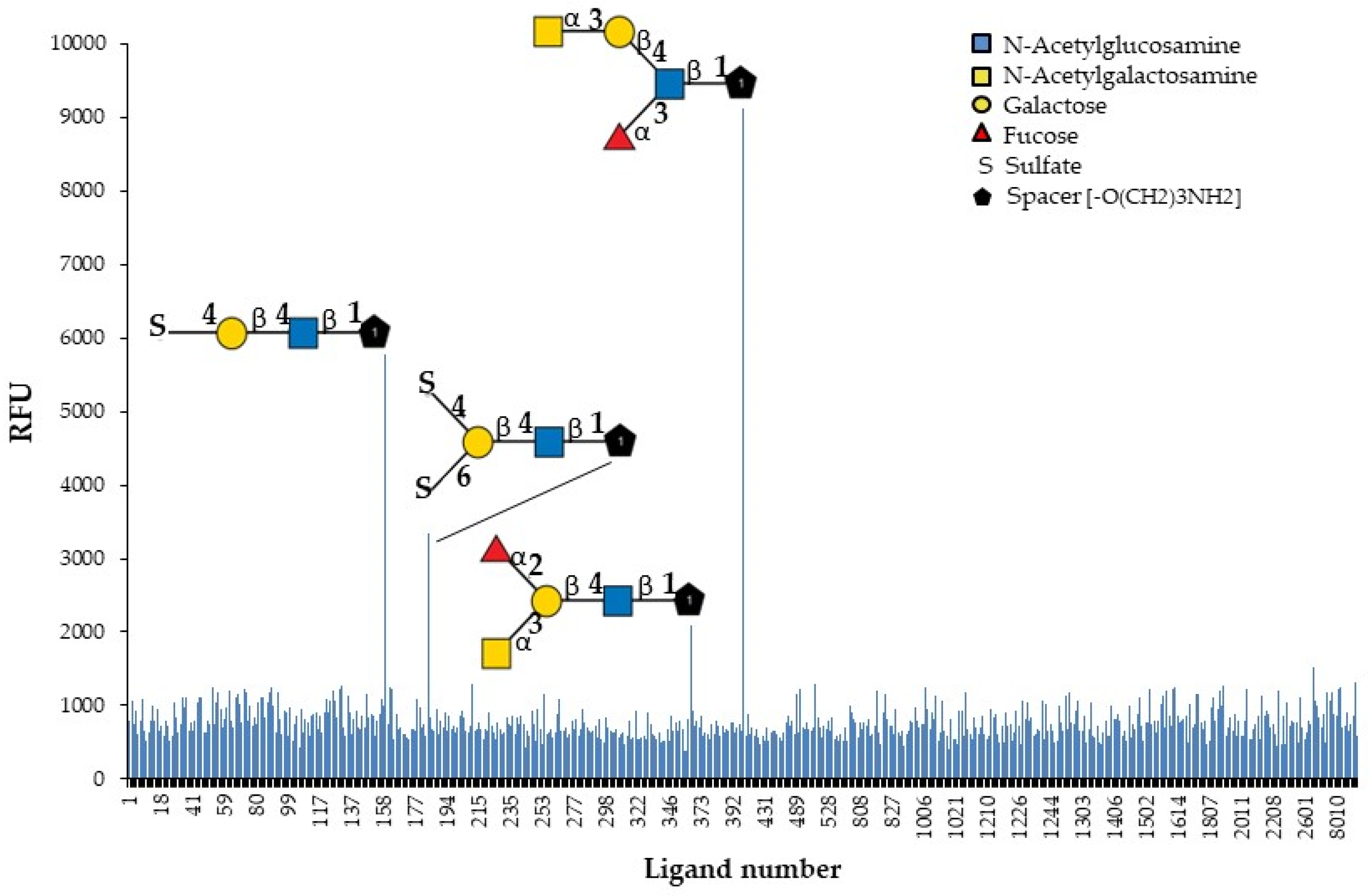
2.2.5. Affinity and Specificity for Oligosaccharides
3. Materials and Methods
3.1. cDNA Sequences Determination and Phylogenetic Analysis
3.2. Bioinformatics Analysis
3.3. N-Terminal Amino Acid Sequencing
3.4. Sugar Analysis
3.5. Determination of Fine Carbohydrate Specificity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buckley, K.; Rast, J.P. Dynamic Evolution of Toll-Like Receptor Multigene Families in Echinoderms. Front. Immunol. 2012, 3, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leoni, G.; De Poli, A.; Mardirossian, M.; Gambato, S.; Florian, F.; Venier, P.; Wilson, D.N.; Tossi, A.; Pallavicini, A.; Gerdol, M. Myticalins: A Novel Multigenic Family of Linear, Cationic Antimicrobial Peptides from Marine Mussels (Mytilus spp.). Mar. Drugs 2017, 15, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwanaga, S.; Lee, B.-L. Recent Advances in the Innate Immunity of Invertebrate Animals. BMB Rep. 2005, 38, 128–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, M.G.; Salazar, K.A.; Joffe, N.R. The immune response of cephalopods from head to foot. Fish Shellfish Immunol. 2015, 46, 145–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, D.; Guang-Hua, W.; Yan-Li, S.; Min, Z.; Yong-Hua, H. Black rockfish C-type lectin, SsCTL4: A pattern recognition receptor that promotes bactericidal activity and virus escape from host immune defense. Fish Shellfish Immunol. 2018, 79, 340–350. [Google Scholar] [CrossRef]
- Catanzaro, E.; Calcabrini, C.; Bishayee, A.; Fimognari, C. Antitumor Potential of Marine and Freshwater Lectins. Mar. Drugs 2019, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Nikolakopoulou, C.; Willment, J.A.; Brown, G.D. C-Type Lectin Receptors in Antifungal Immunity. In Lectin in Host Defense Against Microbial Infections; Hsieh, S.L., Ed.; Springer: Singapore, 2020; pp. 1–30. [Google Scholar]
- Breitenbach Barroso Coelho, L.C.; Marcelino Dos Santos Silva, P.; Felix de Oliveira, W.; de Moura, M.C.; Viana Pontual, E.; Soares Gomes, F.; Guedes Paiva, P.M.; Napoleao, T.H.; Dos Santos Correia, M.T. Lectins as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 1238–1252. [Google Scholar] [CrossRef] [Green Version]
- Nabi-Afjadi, M.; Heydari, M.; Zalpoor, H.; Arman, I.; Sadoughi, A.; Sahami, P.; Aghazadeh, S. Lectins and lectibodies: Potential promising antiviral agents. Cell. Mol. Biol. Lett. 2022, 27, 37. [Google Scholar] [CrossRef]
- Mitchell, C.A.; Ramessar, K.; O’Keefe, B.R. Antiviral lectins: Selective inhibitors of viral entry. Antivir. Res. 2017, 142, 37–54. [Google Scholar] [CrossRef]
- da Silva, L.C.N.; Mendonça, J.S.P.; de Oliveira, W.F.; Batista, K.L.R.; Zagmignan, A.; Viana, I.F.T.; Correia, M.T.D.S. Exploring lectin–glycan interactions to combat COVID-19: Lessons acquired from other enveloped viruses. Glycobiology 2021, 31, 358–371. [Google Scholar] [CrossRef]
- Loris, R. Principles of structures of animal and plant lectins. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2002, 1572, 198–208. [Google Scholar] [CrossRef]
- Gupta, G.S. Lectins: An Overview. In Animal Lectins: Form, Function and Clinical Applications; Springer: Vienna, Austria, 2012; pp. 3–25. [Google Scholar]
- Vasta, G.R.; Ahmed, H.; Fink, N.E.; Elola, M.T.; Marsh, A.G.; Snowden, A.; Odom, E.W. Animal lectins as self/non-self recognition molecules. Biochemical and genetic approaches to understanding their biological roles and evolution. Ann. N. Y. Acad. Sci. 1994, 712, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef] [PubMed]
- Cummings, R.D.; Liu, F.-T.; Rabinovich, G.A.; Stowell, S.R.; Vasta, G.R. Galectins. In Essentials of Glycobiology [Internet]; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Debra, M., Taroh, K., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2022; ISBN 978-1-621824-22-0. [Google Scholar]
- Ogawa, T.; Watanabe, M.; Naganuma, T.; Muramoto, K. Diversified Carbohydrate-Binding Lectins from Marine Resources. J. Amino Acids 2011, 2011, 838914. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Huang, X.; Wang, Z.; Tan, J.-M.; Hui, K.-M.; Wang, W.; Ren, Q. Function of two novel single-CRD containing C-type lectins in innate immunity from Eriocheir sinensis. Fish Shellfish Immunol. 2014, 37, 313–321. [Google Scholar] [CrossRef]
- Qiu, W.; Chen, F.; Chen, R.; Li, S.; Zhu, X.; Xiong, M.; Wang, K.-J. A New C-Type Lectin Homolog SpCTL6 Exerting Immunoprotective Effect and Regulatory Role in Mud Crab Scylla paramamosain. Front. Immunol. 2021, 12, 661823. [Google Scholar] [CrossRef]
- Jin, X.-K.; Li, S.; Guo, X.-N.; Cheng, L.; Wu, M.-H.; Tan, S.-J.; Zhu, Y.-T.; Yu, A.-Q.; Li, W.-W.; Wang, Q. Two antibacterial C-type lectins from crustacean, Eriocheir sinensis, stimulated cellular encapsulation in vitro. Dev. Comp. Immunol. 2013, 41, 544–552. [Google Scholar] [CrossRef]
- Xu, X.; Liao, K.; Shi, P.; Xu, J.; Ran, Z.; Zhou, C.; Zhang, L.; Cao, J.; Yan, X. Involvement of a novel Ca2+-independent C-type lectin from Sinonovacula constricta in food recognition and innate immunity. Fish Shellfish Immunol. 2020, 104, 374–382. [Google Scholar] [CrossRef]
- He, J.; Shen, C.; Liang, H.; Fang, X.; Lu, J. Antimicrobial properties and immune-related gene expression of a C-type lectin isolated from Pinctada fucata martensii. Fish Shellfish Immunol. 2020, 105, 330–340. [Google Scholar] [CrossRef]
- Li, H.; Zhang, H.; Jiang, S.; Wang, W.; Xin, L.; Wang, H.; Wang, L.; Song, L. A single-CRD C-type lectin from oyster Crassostrea gigas mediates immune recognition and pathogen elimination with a potential role in the activation of complement system. Fish Shellfish Immunol. 2015, 44, 566–575. [Google Scholar] [CrossRef]
- Song, X.; Xin, X.; Wang, H.; Li, H.; Zhang, H.; Jia, Z.; Liu, C.; Jiang, S.; Wang, L.; Song, L. A single-CRD C-type lectin (CgCLec-3) with novel DIN motif exhibits versatile immune functions in Crassostrea gigas. Fish Shellfish Immunol. 2019, 92, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Zhang, H.; Jiang, S.; Wang, L.; Liu, R.; Yi, Q.; Song, L. An EPD/WSD motifs containing C-type lectin from Argopectens irradians recognizes and binds microbes with broad spectrum. Fish Shellfish Immunol. 2015, 43, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, L.; Yang, J.; Zhang, H.; Wang, L.; Song, L. A four-CRD C-type lectin from Chlamys farreri mediating nonself-recognition with broader spectrum and opsonization. Dev. Comp. Immunol. 2013, 39, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Mizgina, T.O.; Chikalovets, I.V.; Molchanova, V.I.; Ziganshin, R.H.; Chernikov, O.V. Identification and Characterization of a Novel Lectin from the Clam Glycymeris yessoensis and Its Functional Characterization under Microbial Stimulation and Environmental Stress. Mar. Drugs 2021, 19, 474. [Google Scholar] [CrossRef] [PubMed]
- Morrison, G.M.; Semple, C.A.M.; Kilanowski, F.M.; Hill, R.E.; Dorin, J.R. Signal sequence conservation and mature peptide divergence within subgroups of the murine beta-defensin gene family. Mol. Biol. Evol. 2003, 20, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaeva, M.P.; Chausova, V.E.; Zelepuga, E.A.; Guzev, K.V.; Tabakmakher, V.M.; Monastyrnaya, M.M.; Kozlovskaya, E.P. A new multigene superfamily of Kunitz-type protease inhibitors from sea anemone Heteractis crispa. Peptides 2012, 34, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Lu, G. Vector NTI, a balanced all-in-one sequence analysis suite. Brief. Bioinform. 2004, 5, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, N.G.; Surolia, A. Role of glycosylation in nucleating protein folding and stability. Biochem. J. 2017, 474, 2333–2347. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, H.; Taylor, M.; Weis, W. Scavenger Receptor C-type Lectin Binds to the Leukocyte Cell Surface Glycan Lewisx by a Novel Mechanism. J. Biol. Chem. 2007, 282, 17250–17258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, H.; Rambaruth, N.D.; Jégouzo, S.A.; Jacobsen, K.M.; Djurhuus, R.; Poulsen, T.B.; Weis, W.I.; Taylor, M.E.; Drickamer, K. Binding Sites for Acylated Trehalose Analogs of Glycolipid Ligands on an Extended Carbohydrate Recognition Domain of the Macrophage Receptor Mincle. J. Biol. Chem. 2016, 291, 21222–21233. [Google Scholar] [CrossRef] [PubMed]
- Silva-Martín, N.; Bartual, S.G.; Ramírez-Aportela, E.; Chacón, P.; Park, C.G.; Hermoso, J.A. Structural Basis for Selective Recognition of Endogenous and Microbial Polysaccharides by Macrophage Receptor SIGN-R1. Structure 2014, 22, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourdine, J.-P.; Cioci, G.; Miguet, L.; Unverzagt, C.; Silva, D.V.; Varrot, A.; Gautier, C.; Smith-Ravin, E.J.; Imberty, A. High Affinity Interaction between a Bivalve C-type Lectin and a Biantennary Complex-type N-Glycan Revealed by Crystallography and Microcalorimetry. J. Biol. Chem. 2008, 283, 30112–30120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Musik, J.E.; Zalucki, Y.M.; Beacham, I.R.; Jennings, M.P. The role of signal sequence proximal residues in the mature region of bacterial secreted proteins in E. coli. Biochim. et Biophys. Acta (BBA)-Biomembr. 2022, 1864, 184000. [Google Scholar] [CrossRef]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef]
- Halder, S.; Surolia, A.; Mukhopadhyay, C. Impact of glycosylation on stability, structure and unfolding of soybean agglutinin (SBA): An insight from thermal perturbation molecular dynamics simulations. Glycoconj. J. 2015, 32, 371–384. [Google Scholar] [CrossRef]
- Staudacher, E. Mollusc N-glycosylation: Structures, Functions and Perspectives. Biomolecules 2021, 11, 1820. [Google Scholar] [CrossRef]
- Zhu, F.; Li, D.; Chen, K. Structures and functions of invertebrate glycosylation. Open Biol. 2019, 9, 180232. [Google Scholar] [CrossRef] [Green Version]
- Gutternigg, M.; Bürgmayr, S.; Pöltl, G.; Rudolf, J.; Staudacher, E. Neutral N-glycan patterns of the gastropods Limax maximus, Cepaea hortensis, Planorbarius corneus, Arianta arbustorum and Achatina fulica. Glycoconj. J. 2007, 24, 475–489. [Google Scholar] [CrossRef]
- García-Maldonado, E.; Cano-Sánchez, P.; Hernández-Santoyo, A. Molecular and functional characterization of a glycosylated Galactose-Binding lectin from Mytilus californianus. Fish Shellfish Immunol. 2017, 66, 564–574. [Google Scholar] [CrossRef]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo—Distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poget, S.F.; Legge, G.B.; Proctor, M.R.; Butler, P.G.; Bycroft, M.; Williams, R.L. The structure of a tunicate C-type lectin from polyandrocarpa misakiensis complexed with d-galactose. J. Mol. Biol. 1999, 290, 867–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, H.; Kusunoki, M.; Kurisu, G.; Fujimoto, T.; Aoyagi, H.; Hatakeyama, T. Characteristic Recognition of N-Acetylgalactosamine by an Invertebrate C-type Lectin, CEL-I, Revealed by X-ray Crystallographic Analysis. J. Biol. Chem. 2004, 279, 45219–45225. [Google Scholar] [CrossRef] [Green Version]
- Oinam, L.; Tateno, H. Evaluation of Glycan-Binding Specificity by Glycoconjugate Microarray with an Evanescent-Field Fluorescence Detection System. Methods Mol. Biol. 2022, 2460, 25–32. [Google Scholar] [CrossRef]
- Iiyama, C.; Yoneda, F.; Tsutsumi, M.; Tsutsui, S.; Nakamura, O. Mannose-binding C-type lectins as defense molecules on the body surface of the sea urchin Pseudocentrotus depressus. Dev. Comp. Immunol. 2021, 116, 103915. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Kamiya, T.; Kusunoki, M.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Goda, S.; Unno, H. Galactose Recognition by a Tetrameric C-type Lectin, CEL-IV, Containing the EPN Carbohydrate Recognition Motif. J. Biol. Chem. 2011, 286, 10305–10315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef]
- Moriuchi, H.; Unno, H.; Goda, S.; Tateno, H.; Hirabayashi, J.; Hatakeyama, T. Mannose-recognition mutant of the galactose/N-acetylgalactosamine-specific C-type lectin CEL-I engineered by site-directed mutagenesis. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2015, 1850, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Kokoulin, M.S.; Sigida, E.N.; Kuzmich, A.S.; Ibrahim, I.M.; Fedonenko, Y.P.; Konnova, S.A. Structure and antiproliferative activity of the polysaccharide from Halomonas aquamarina related to Cobetia pacifica. Carbohydr. Polym. 2022, 298, 120125. [Google Scholar] [CrossRef]
- Kokoulin, M.S.; Kuzmich, A.S.; Romanenko, L.A.; Chikalovets, I.V. Sulfated capsular polysaccharide from the marine bacterium Kangiella japonica inhibits T-47D cells growth in vitro. Carbohydr. Polym. 2022, 290, 119477. [Google Scholar] [CrossRef]
- Kokoulin, M.S.; Kalinovsky, A.I.; Komandrova, N.A.; Tomshich, S.V.; Romanenko, L.A.; Vaskovsky, V.E. The sulfated O-specific polysaccharide from the marine bacterium Cobetia pacifica KMM 3879T. Carbohydr. Res. 2014, 387, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Kokoulin, M.S.; Filshtein, A.P.; Romanenko, L.A.; Chikalovets, I.V.; Chernikov, O.V. Structure and bioactivity of sulfated α-D-mannan from marine bacterium Halomonas halocynthiae KMM 1376T. Carbohydr. Polym. 2020, 229, 115556. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The GenBank Accession Numbers | Name | Molecular Weight Mature Proteins | pI | Components of the Secondary Structure of GYL-like CTL | ||
|---|---|---|---|---|---|---|
| α-Helix, % | β-Strand, % | Coil, % | ||||
| OP897819 | GYL | 16,046.59 | 4.6 | 17.02 | 23.40 | 59.57 |
| OP897820 | GYL-like 1 | 16,482.77 | 5.15 | 15.07 | 26.71 | 58.22 |
| OP897821 | GYL-like 2 | 16,424.74 | 5.32 | 15.75 | 26.71 | 57.53 |
| OP897822 | GYL-like 3 | 16,862.01 | 4.39 | 17.01 | 22.45 | 60.54 |
| OP897823 | GYL-like 4 | 15,996.31 | 4.56 | 17.27 | 26.62 | 56.12 |
| OP897824 | GYL-like 5 | 16,095.44 | 4.65 | 17.27 | 25.90 | 56.83 |
| OP897825 | GYL-like 6 | 16,081.42 | 4.65 | 17.27 | 25.90 | 56.83 |
| OP897826 | GYL-like 7 | 16,144.37 | 4.71 | 17.27 | 28.78 | 53.96 |
| OP897827 | GYL-like 8 | 16,042.21 | 4.65 | 17.39 | 27.54 | 55.07 |
| # | Glycan Structure | Short or Trivial Name | Average RFU |
|---|---|---|---|
| 404 | GalNAcα1-3Galβ1-4(Fucα1-3)GlcNAcβ-sp3 | GalNAcα3′Lex-C3 | 9114 |
| 159 | 4-O-Su-Galβ1-4GlcNAcβ-sp3 | 4′-suLN-C3 | 5774 |
| 183 | 4,6-O-Su2-Galβ1-4GlcNAcβ-sp2 | 4′,6′-su2LN-C2 | 3338 |
| 368 | GalNAcα1-3(Fucα1-2)Galβ1-4GlcNAcβ-sp3 | A (type 2)-C3 | 2080 |
| 97 | Galβ1-4GlcNAcβ-sp3 | LN-C3 | 929 |
| 234 | Galβ1-4(Fucα1-3)GlcNAcβ-sp3 | Lex-C3 | 742 |
| 275 | GalNAcβ1-3Galβ1-4GlcNAcβ-sp3 | GalNAcβ3′LN-C3 | 615 |
| Selective GYL (50 µg/mL) binding entities, including a serial number of each glycan (#), glycan structure, and binding signals in relative fluorescence units (average RFU) in decreasing order. | |||
| Primers Name | Sequence (5′–3′) | PCR Objective |
|---|---|---|
| Lectin_PSTE_For | AGCNCARCCNCANWSNGARGG | 3’-RACE |
| Lectin_MMD_For | ATGATGGAYTAYGTNRAYTGGATG | |
| GYL_Rev1_Cap | ACATCTTTCCGGTCAATGACTAT | 5’ RACE |
| GYL_Rev2-T7 | ACTATATTACCGATCCCAAGGAGT | |
| GYL1_start | CAGTTTATTGAGGAAGGAGACTTG | PCR of full-length cDNA |
| GYL1_stop1 | CAATGACTATATTACCGATCCCAA | |
| M13 F | GTAAAACGACGGCCAGT | Sanger sequence of clones |
| M13 R | CAGGAAACAGCTATGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizgina, T.O.; Baldaev, S.N.; Likhatskaya, G.N.; Molchanova, V.I.; Kokoulin, M.S.; Filshtein, A.P.; Rogozhin, E.A.; Chikalovets, I.V.; Isaeva, M.P.; Chernikov, O.V. Molecular Cloning and Characteristics of a Lectin from the Bivalve Glycymeris yessoensis. Mar. Drugs 2023, 21, 55. https://doi.org/10.3390/md21020055
Mizgina TO, Baldaev SN, Likhatskaya GN, Molchanova VI, Kokoulin MS, Filshtein AP, Rogozhin EA, Chikalovets IV, Isaeva MP, Chernikov OV. Molecular Cloning and Characteristics of a Lectin from the Bivalve Glycymeris yessoensis. Marine Drugs. 2023; 21(2):55. https://doi.org/10.3390/md21020055
Chicago/Turabian StyleMizgina, Tatyana O., Sergey N. Baldaev, Galina N. Likhatskaya, Valentina I. Molchanova, Maxim S. Kokoulin, Alina P. Filshtein, Eugene A. Rogozhin, Irina V. Chikalovets, Marina P. Isaeva, and Oleg V. Chernikov. 2023. "Molecular Cloning and Characteristics of a Lectin from the Bivalve Glycymeris yessoensis" Marine Drugs 21, no. 2: 55. https://doi.org/10.3390/md21020055