
Secondary Metabolites from Marine-Derived Bacillus: A Comprehensive Review of Origins, Structures, and Bioactivities
 , , , , , and
, , , , , and
Abstract
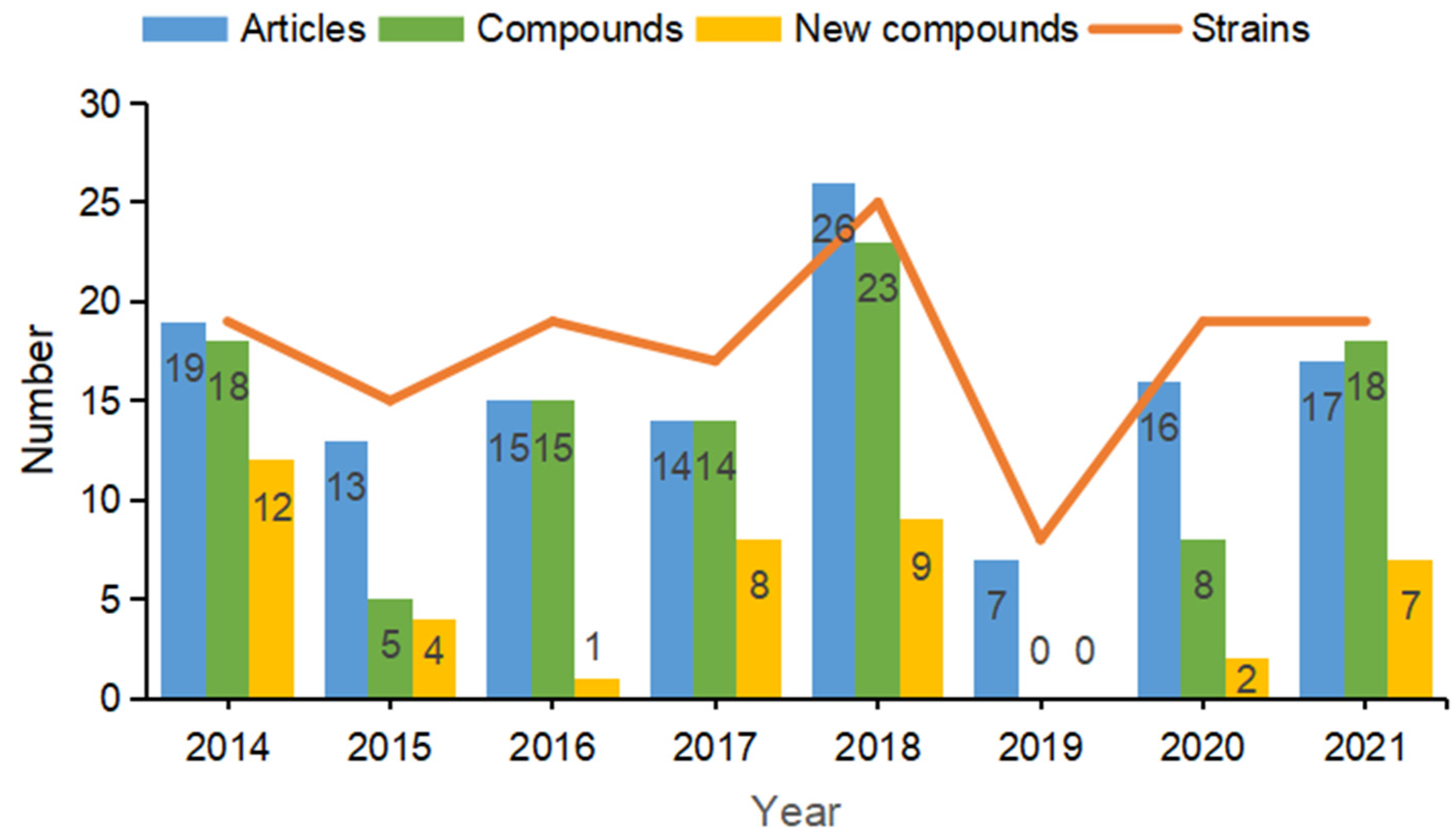
:1. Introduction
2. Structural Diversity
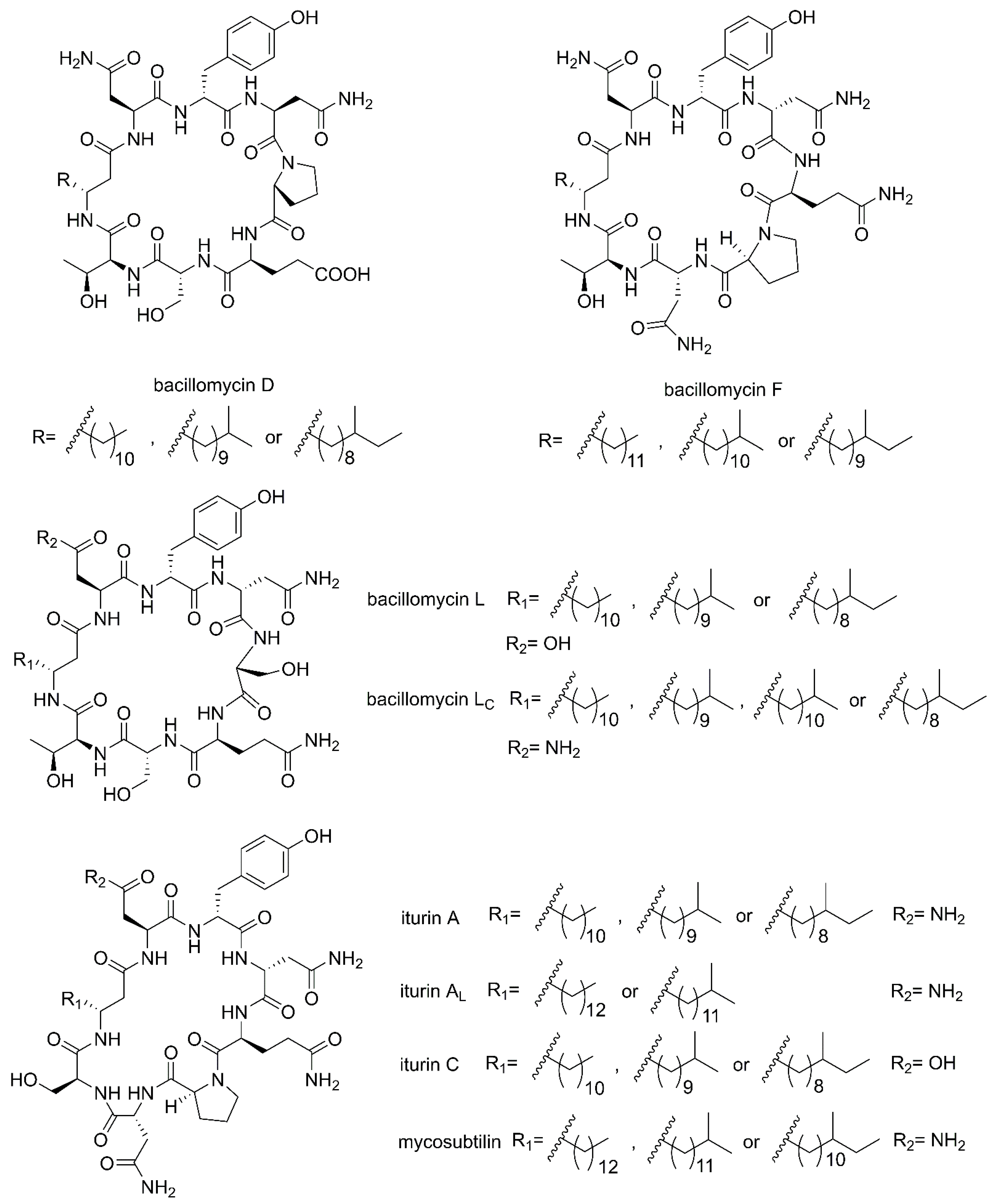
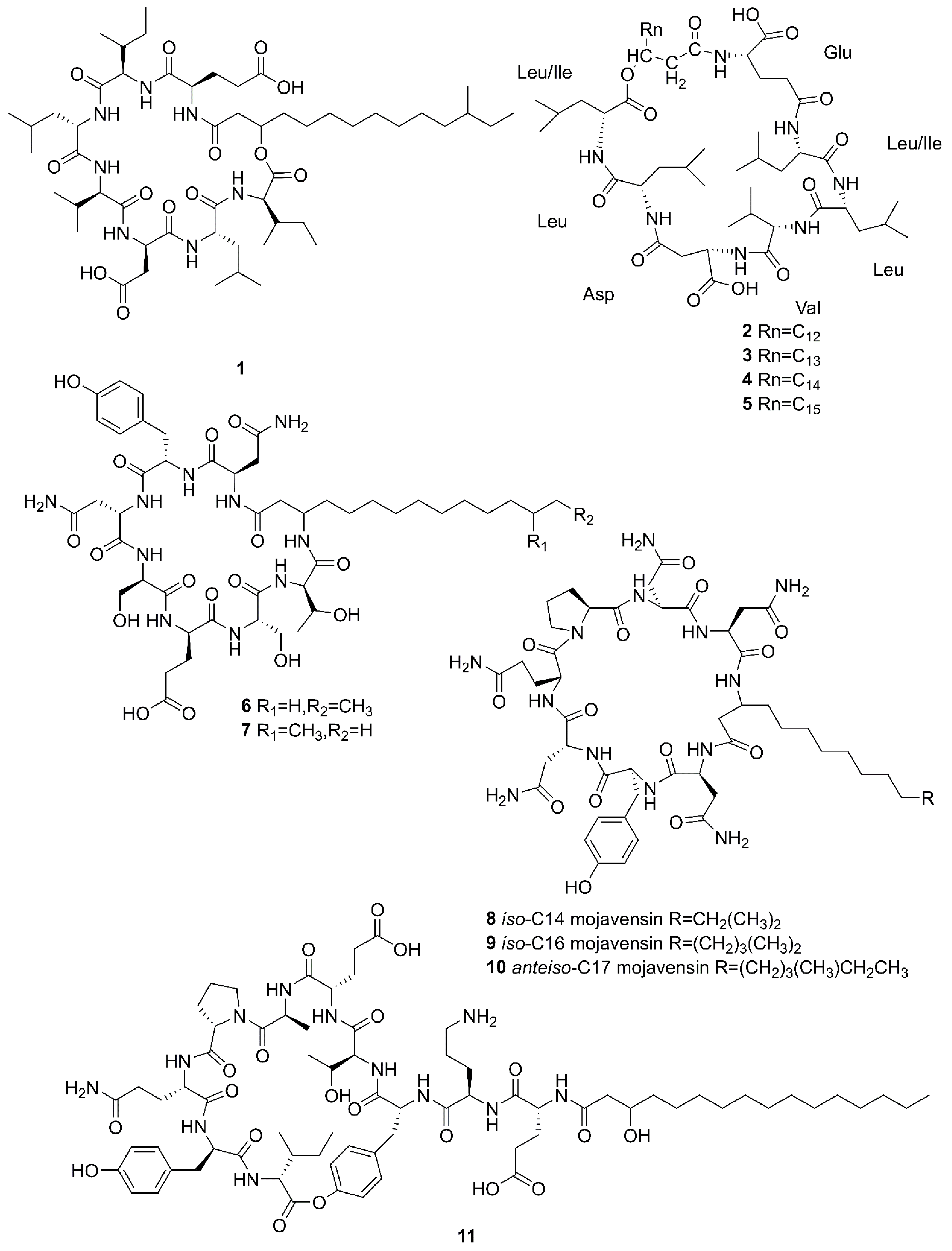
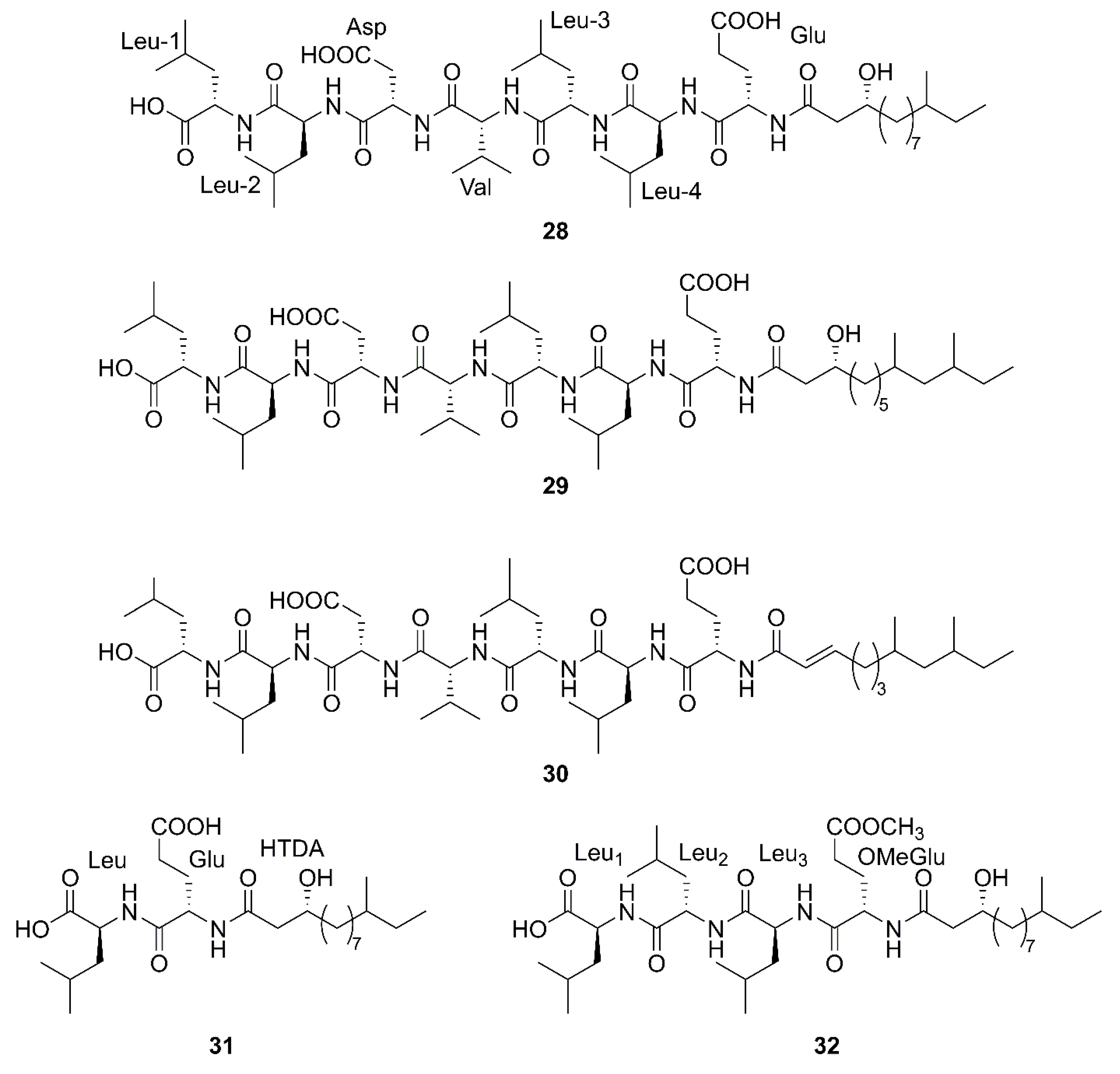
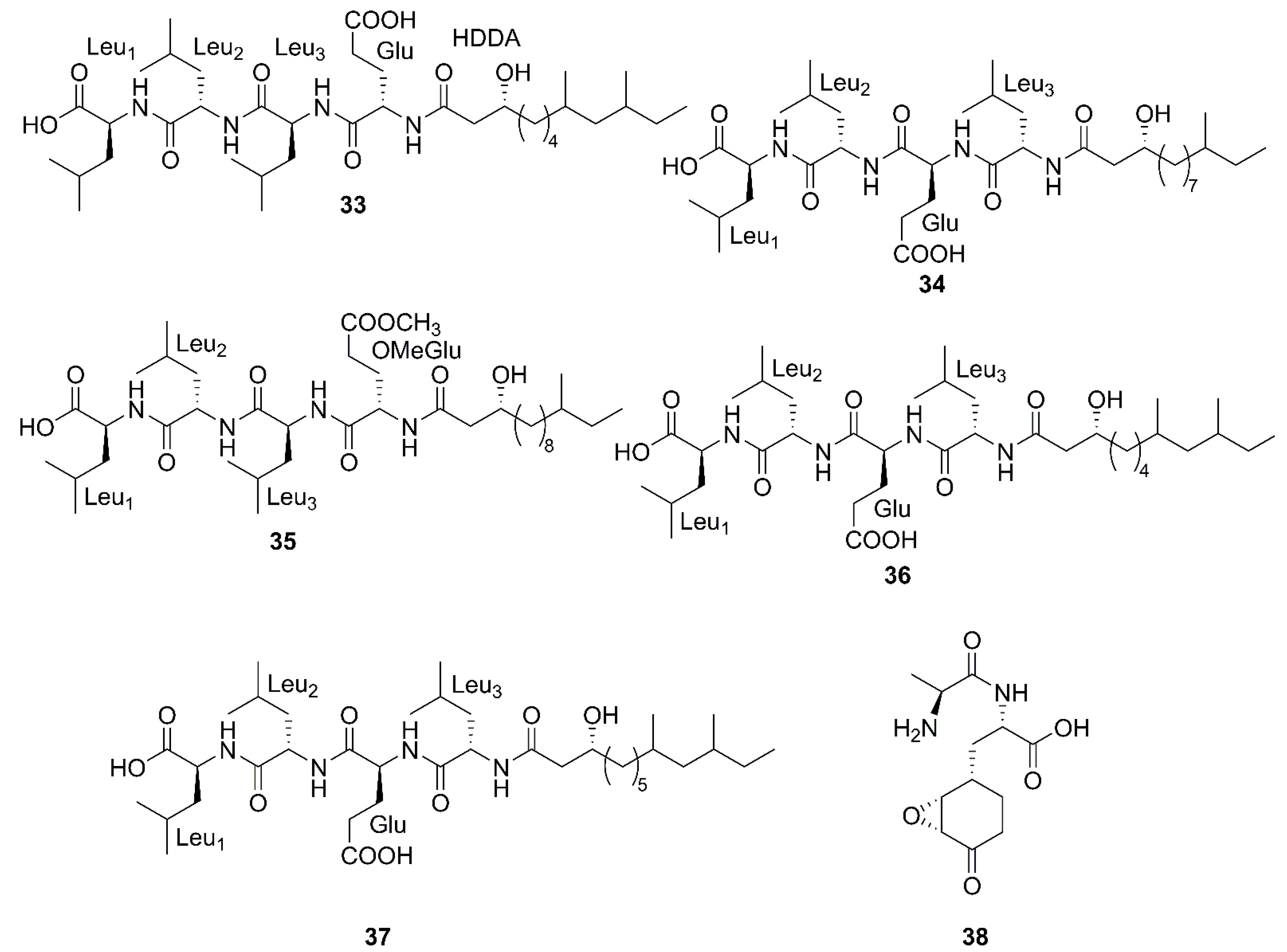
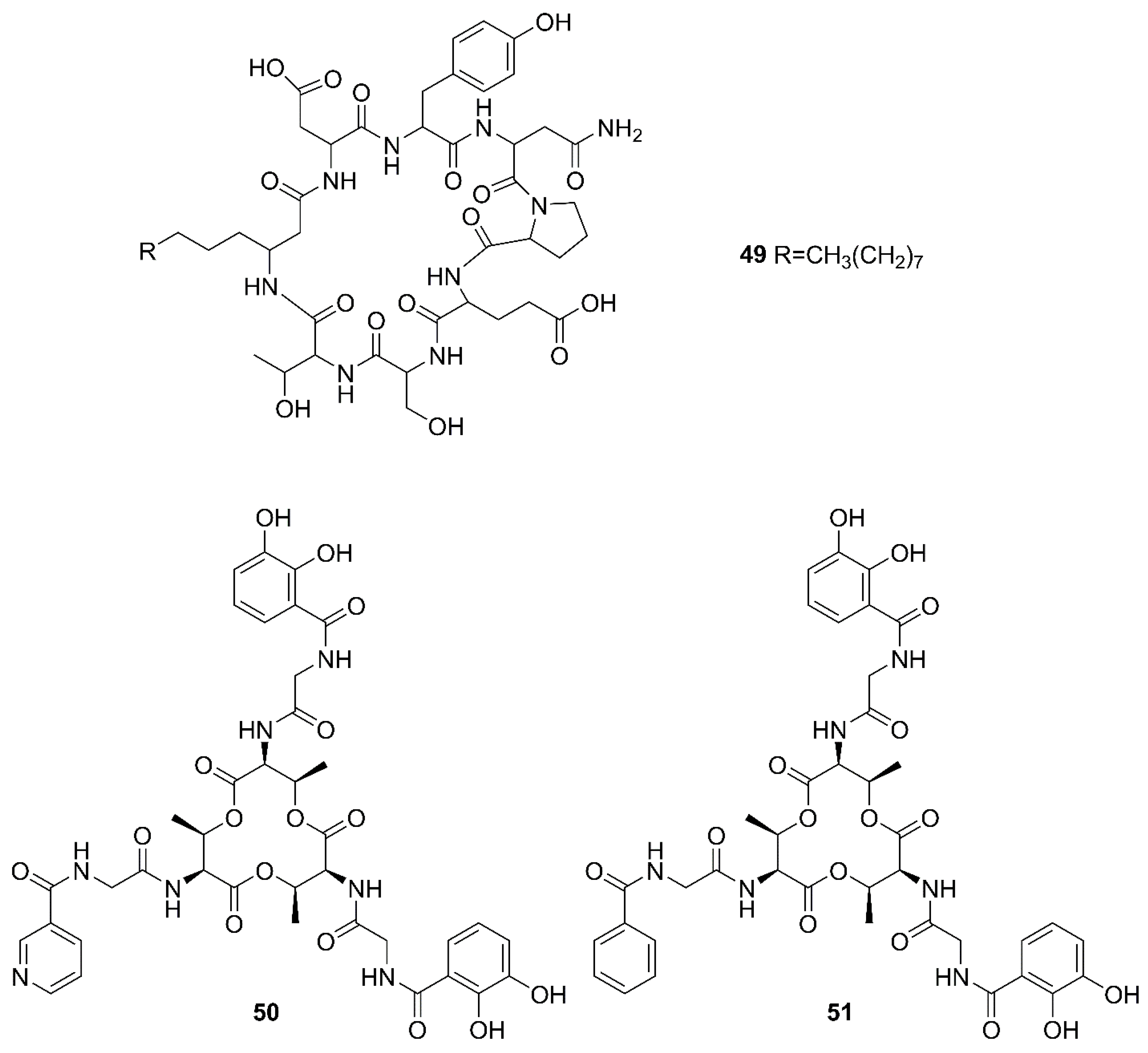
2.1. Cyclic Lipopeptides
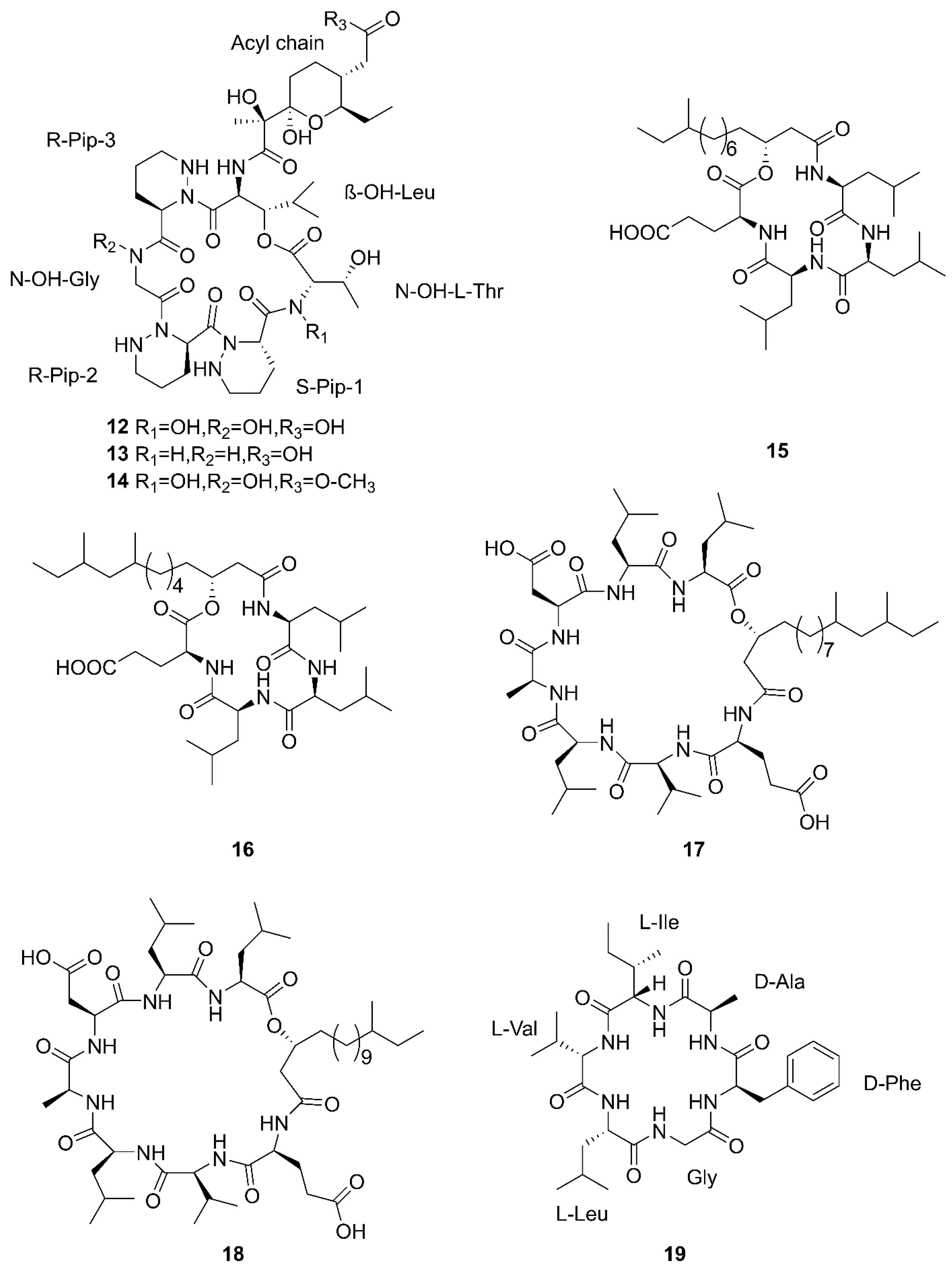
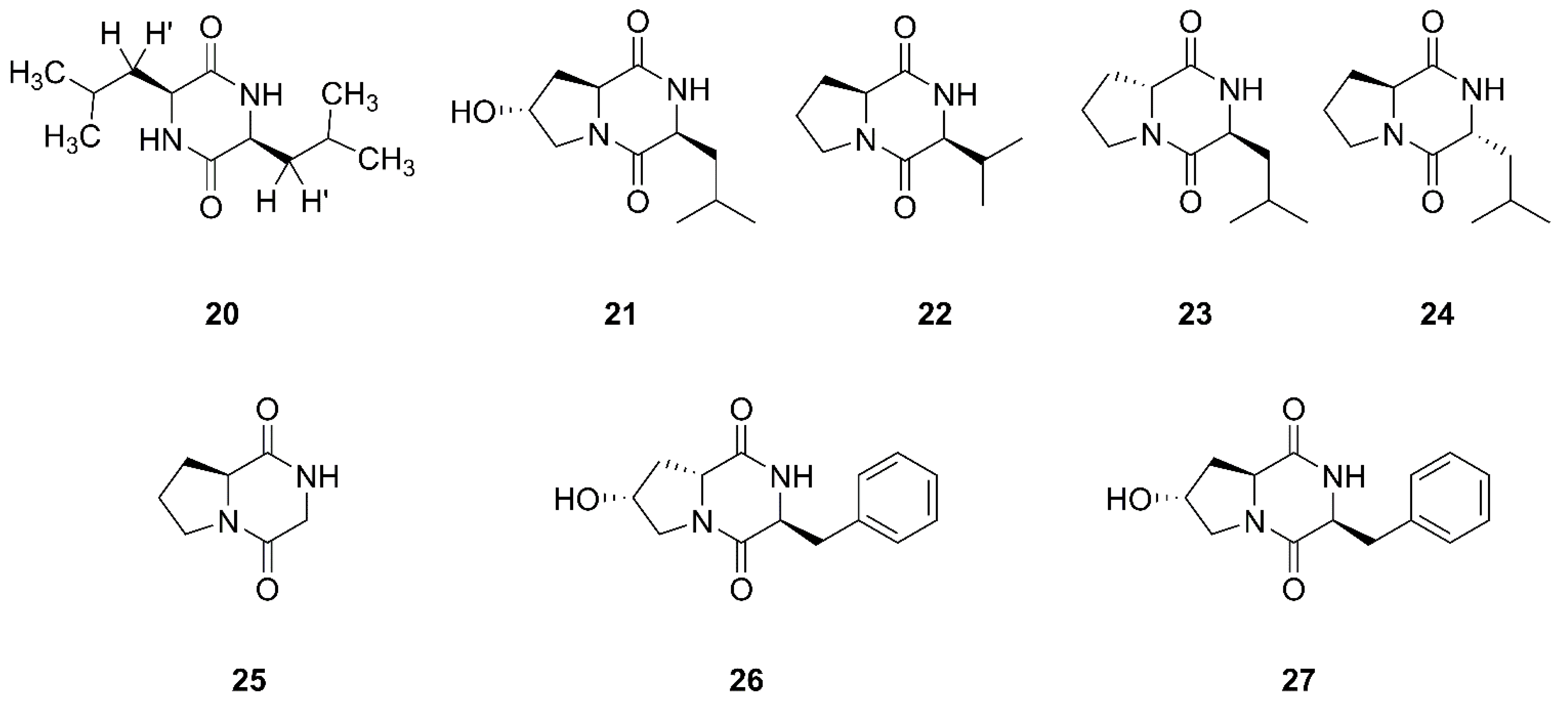
2.2. Diketopiperazines
2.3. Linear Lipopeptides
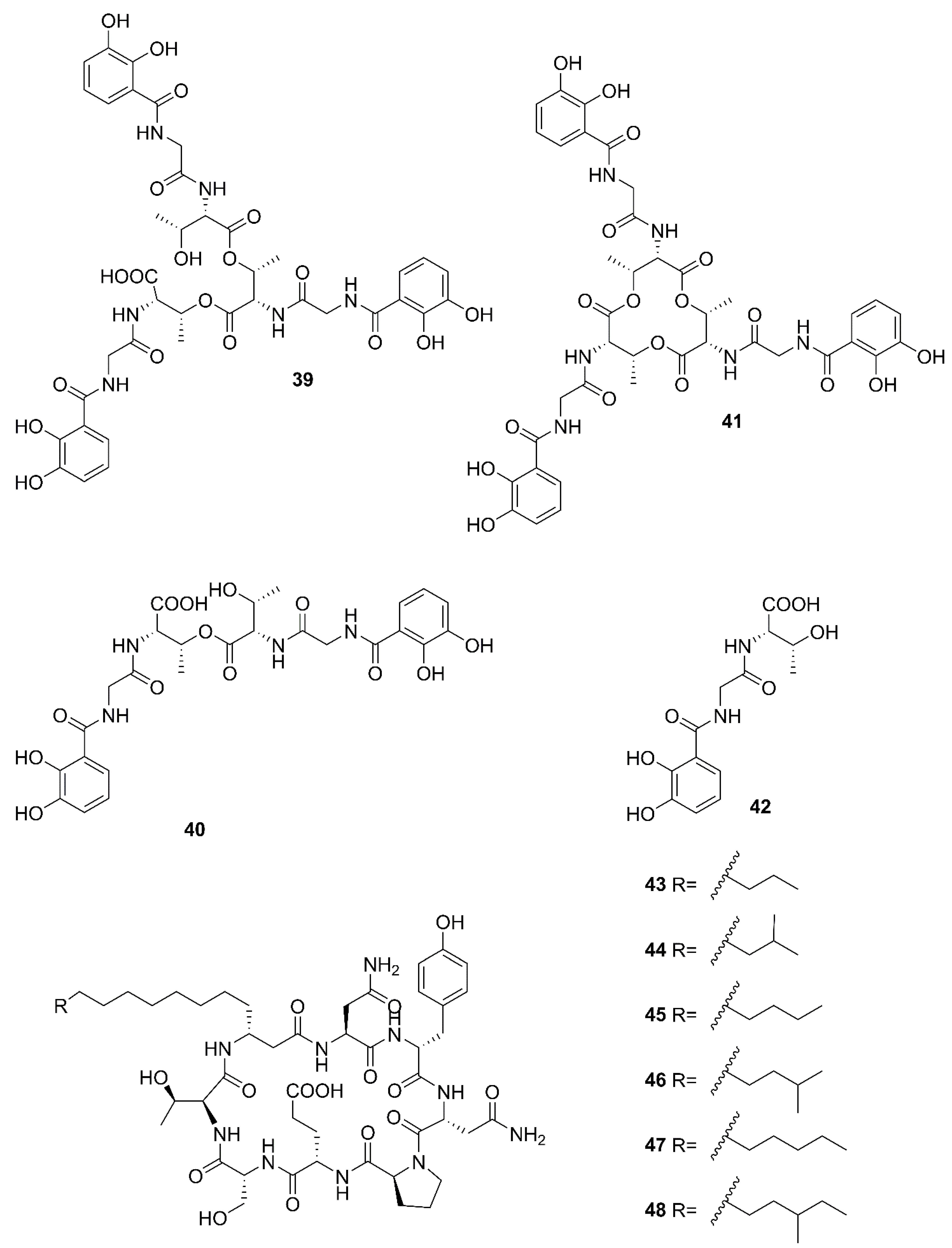
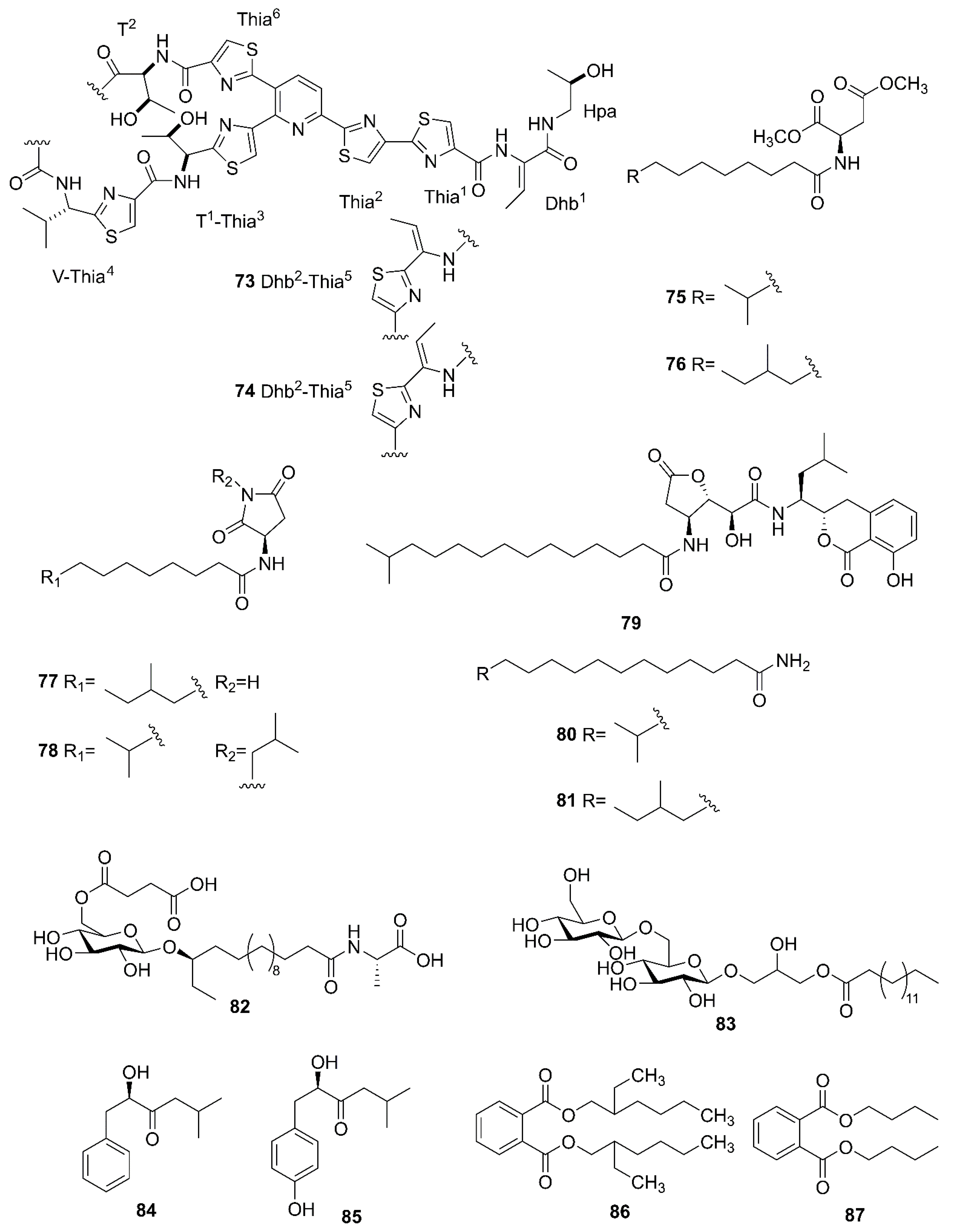
2.4. Nonribosomal Peptides
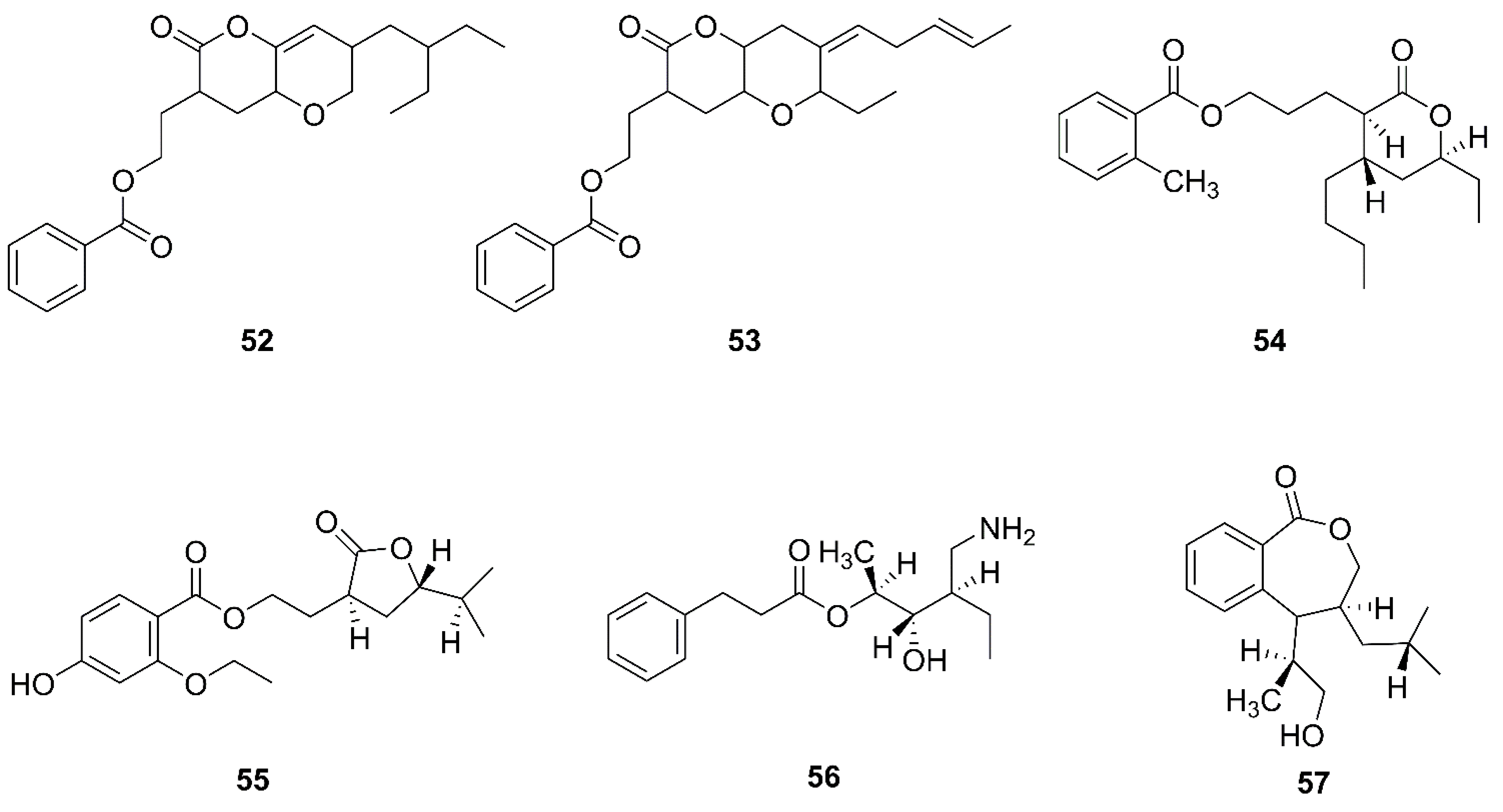
2.5. Polyketides
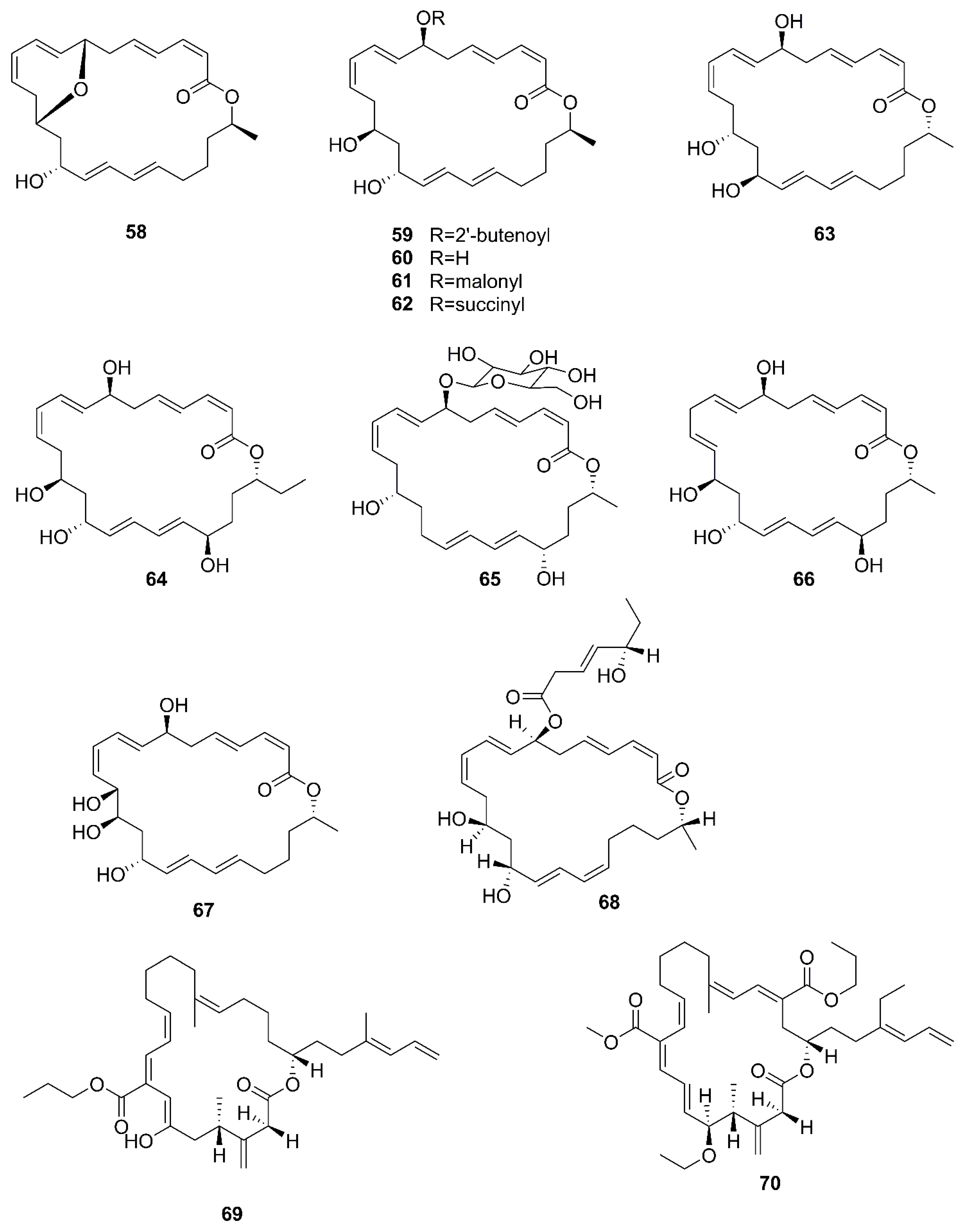
2.6. Macrolactins
2.7. Other Compounds
3. Biological Activities
3.1. Antimicrobial Activities
3.2. Antifungal Activities
3.3. Cytotoxic Activities
3.4. Other Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine Microorganisms as a Promising and Sustainable Source of Bioactive Molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef]
- Barzkar, N. Marine Microbial Alkaline Protease: An Efficient and Essential Tool for Various Industrial Applications. Int. J. Biol. Macromol. 2020, 161, 1216–1229. [Google Scholar] [CrossRef]
- Zheng, L.H.; Zhu, X.J.; Yang, K.L.; Zhu, M.H.; Farooqi, A.A.; Kang, D.L.; Sun, M.; Xu, Y.X.; Lin, X.K.; Feng, Y.G.; et al. PBN11-8, a Cytotoxic Polypeptide Purified from Marine Bacillus, Suppresses Invasion and Migration of Human Hepatocellular Carcinoma Cells by Targeting Focal Adhesion Kinase Pathways. Polymers 2018, 10, 1043. [Google Scholar] [CrossRef]
- De Rop, A.S.; Rombaut, J.; Willems, T.; De Graeve, M.; Vanhaecke, L.; Hulpiau, P.; De Maeseneire, S.L.; De Mol, M.L.; Soetaert, W.K. Novel Alkaloids from Marine Actinobacteria: Discovery and Characterization. Mar. Drugs 2021, 20, 6. [Google Scholar] [CrossRef]
- El-Sersy, N.A.; Abdelwahab, A.E.; Abouelkhiir, S.S.; Abou-Zeid, D.M.; Sabry, S.A. Antibacterial and Anticancer Activity of ε-Poly-L-Lysine (Ε-Pl) Produced by a Marine Bacillus subtilis sp. J. Basic Microbiol. 2012, 52, 513–522. [Google Scholar] [CrossRef]
- Habbu, P.; Warad, V.; Shastri, R.; Madagundi, S.; Kulkarni, V.H. Antimicrobial Metabolites from Marine Microorganisms. Chin. J. Nat. Med. 2016, 14, 101–116. [Google Scholar] [CrossRef]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine Microorganisms as an Untapped Source of Bioactive Compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- El-Hossary, E.M.; Cheng, C.; Hamed, M.M.; El-Sayed Hamed, A.N.; Ohlsen, K.; Hentschel, U.; Abdelmohsen, U.R. Antifungal Potential of Marine Natural Products. Eur. J. Med. Chem. 2017, 126, 631–651. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, S.S.; Baindara, P.; Sharma, S.; Khatri, N.; Grover, V.; Patil, P.B.; Korpole, S. Surfactin Like Broad Spectrum Antimicrobial Lipopeptide Co-Produced with Sublancin from Bacillus Subtilis Strain A52: Dual Reservoir of Bioactives. Front. Microbiol. 2020, 11, 1167. [Google Scholar] [CrossRef]
- Mawlankar, R.; Thorat, M.N.; Krishnamurthi, S.; Dastager, S.G. Bacillus cellulasensis sp. nov., Isolated from Marine Sediment. Arch. Microbiol. 2016, 198, 83–89. [Google Scholar] [CrossRef]
- Liu, R.; Huang, Z.B.; Dong, C.M.; Shao, Z.Z. Lottiidibacillus patelloidae Gen. Nov., Sp. Nov., Isolated from the Intestinal Tract of a Marine Limpet and Reclassification of Bacillus taeanensis as Maribacillus taeanensis Gen. Nov., Comb. Nov. Antonie Leeuwenhoek 2019, 112, 797–807. [Google Scholar] [CrossRef]
- Tareq, F.S.; Shin, H.J. Bacilotetrins A and B, Anti-Staphylococcal Cyclic-Lipotetrapeptides from a Marine-Derived Bacillus subtilis. J. Nat. Prod. 2017, 80, 2889–2892. [Google Scholar] [CrossRef]
- Chakraborty, K.; Kizhakkekalam, V.K.; Joy, M.; Dhara, S. Difficidin Class of Polyketide Antibiotics from Marine Macroalga-Associated Bacillus as Promising Antibacterial Agents. Appl. Microbiol. Biotechnol. 2021, 105, 6395–6408. [Google Scholar] [CrossRef]
- Harwood, C.R.; Mouillon, J.M.; Pohl, S.; Arnau, J. Secondary Metabolite Production and the Safety of Industrially Important members of the Bacillus subtilis group. FEMS. Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef]
- Dame, Z.T.; Rahman, M.; Islam, T. Bacilli as Sources of Agrobiotechnology: Recent Advances and Future Directions. Green Chem. Lett. Rev. 2021, 14, 245–270. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.H. Bacillus velezensis: A Valuable Member of Bioactive Molecules Within Plant Microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef]
- Bibi, F.; Naseer, M.I.; Azhar, E.I. Assessing the Diversity of Bacterial Communities from Marine Sponges and Their Bioactive Compounds. Saudi J. Biol. Sci. 2021, 28, 2747–2754. [Google Scholar] [CrossRef]
- Zhang, L.L.; Sun, C.M. Fengycins, Cyclic Lipopeptides from Marine Bacillus subtilis strains, Kill the Plant-Pathogenic Fungus Magnaporthe grisea by Inducing Reactive Oxygen Species Production and Chromatin Condensation. Appl. Environ. Microbiol. 2018, 84, e00418–e00445. [Google Scholar] [CrossRef]
- Kizhakkekalam, V.K.; Chakraborty, K.; Joy, M. Oxygenated Elansolid Type of Polyketide Spanned Macrolides from a Marine Heterotrophic Bacillus as Prospective Antimicrobial Agents Against Multidrug Resistant Pathogens. Int. J. Antimicrob. Agents 2020, 55, 105892. [Google Scholar] [CrossRef]
- Karthik, L.; Sun, W.; Wang, Y.K.; Mulati, N.; Gong, S.Q.; Zhang, F.L.; Li, Z.Y.; Li, Y.X. Biosynthesis In Vitro of Bacillamide Intermediate-Heterocyclic Alacysthiazole by Heterologous Expression of Nonribosomal Peptide Synthetase (NRPS). J. Biotechnol. 2019, 292, 5–11. [Google Scholar] [CrossRef]
- Yi, X.X.; Gan, Y.M.; Jiang, L.; Yu, L.; Liu, Y.H.; Gao, C.H. Rapid Improvement in the Macrolactins Production of Bacillus Sp. Combining Atmospheric Room Temperature Plasma with the Specific Growth Rate Index. J. Biosci. Bioeng. 2020, 130, 48–53. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, J.; Wang, X.; Wang, S. NMR Spectroscopic And MS/MS Spectrometric Characterization of a New Lipopeptide Antibiotic Bacillopeptin B1 Produced by a Marine Sediment-Derived Bacillus amyloliquefaciens SH-B74. J. Antibiot. 2014, 67, 175–178. [Google Scholar] [CrossRef]
- Ma, Y.X.; Kong, Q.; Qin, C.; Chen, Y.L.; Chen, Y.J.; Lv, R.H.; Zhou, G.H. Identification of Lipopeptides in Bacillus megaterium by Two-Step Ultrafiltration and LC-ESI-MS/MS. AMB Express 2016, 6, 79. [Google Scholar] [CrossRef]
- Wu, Q.H.; Throckmorton, K.; Maity, M.; Chevrette, M.G.; Braun, D.R.; Rajski, S.R.; Currie, C.R.; Thomas, M.G.; Bugni, T.S. Bacillibactins E and F from a Marine Sponge-Associated Bacillus sp. J. Nat. Prod. 2021, 84, 136–141. [Google Scholar] [CrossRef]
- Shin, D.; Byun, W.S.; Moon, K.; Kwon, Y.; Bae, M.; Um, S.; Lee, S.K.; Oh, D.C. Coculture of Marine Streptomyces sp. With Bacillus sp. Produces a New Piperazic Acid-Bearing Cyclic Peptide. Front. Chem. 2018, 6, 498. [Google Scholar] [CrossRef]
- Stincone, P.; Brandelli, A. Marine bacteria as source of antimicrobial compounds. Crit. Rev. Biotechnol. 2020, 40, 306–319. [Google Scholar] [CrossRef]
- Fu, X.T.; Gong, L.F.; Liu, Y.; Lai, Q.L.; Li, G.Y.; Shao, Z.Z. Bacillus pumilus Group Comparative Genomics: Toward Pangenome Features, Diversity, and Marine Environmental Adaptation. Front. Microbiol. 2021, 2, 571212. [Google Scholar] [CrossRef]
- Subramenium, G.A.; Swetha, T.K.; Iyer, P.M.; Balamurugan, K.; Pandian, S.K. 5-Hydroxymethyl-2-Furaldehyde from Marine Bacterium Bacillus Subtilis Inhibits Biofilm and Virulence of Candida albicans. Microbiol. Res. 2018, 207, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.H.; Yu, Q.H.; Yang, S.; Wen, J.; Guo, Z.H.; Wang, X.L.; Wang, N. Ultrafast Recovery of Uranium from Seawater by Bacillus velezensis Strain UUS-1 with Innate Anti-Biofouling Activity. Adv. Sci. (Weinh.) 2019, 6, 1900961. [Google Scholar] [CrossRef]
- Ibrahim, A.Y.; Youness, E.R.; Mahmoud, M.G.; Asker, M.S.; El-Newary, S.A. Acidic Exopolysaccharide Produced from Marine Bacillus Amyloliquefaciens 3MS 2017 For the Protection and Treatment of Breast Cancer. Breast Cancer 2020, 14, 1178223420902075. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Cong, B.L.; Tian, Y.Q.; He, Y.; Yang, H.H. Characterization of Novel Cyclic Lipopeptides Produced by Bacillus Sp. SY27F. Process Biochem. 2019, 83, 206–213. [Google Scholar] [CrossRef]
- Dan, A.K.; Manna, A.; Ghosh, S.; Sikdar, S.; Sahu, R.; Parhi, P.K.; Parida, S. Molecular Mechanisms of the Lipopeptides from Bacillus subtilis in the Apoptosis of Cancer Cells-A Review on Its Current Status in Different Cancer Cell Lines. Adv. Cancer Biol. Met. 2021, 3, 100019. [Google Scholar] [CrossRef]
- Routhu, S.R.; Nagarjuna Chary, R.; Shaik, A.B.; Prabhakar, S.; Kumar, C.G.; Kamal, A. Induction of Apoptosis in Lung Carcinoma Cells by Antiproliferative Cyclic Lipopeptides from Marine Algicolous Isolate Bacillus atrophaeus Strain AKLSR1. Process Biochem. 2019, 79, 142–154. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological Control of Plant Pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Tareq, F.S.; Lee, M.A.; Lee, H.-S.; Lee, J.-S.; Lee, Y.-J.; Shin, H.J. Gageostatins A–C, Antimicrobial Linear Lipopeptides from a Marine Bacillus subtilis. Mar. Drugs 2014, 12, 871–885. [Google Scholar] [CrossRef]
- Aleti, G.; Sessitsch, A.; Brader, G. Genome mining: Prediction of Lipopeptides and Polyketides from Bacillus and Related Firmicutes. Comput. Struct. Biotechnol. J. 2015, 13, 192–203. [Google Scholar] [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative Analysis of the Complete Genome Sequence of the Plant Growth-Promoting Bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef]
- Chen, X.H.; Scholz, R.; Borriss, M.; Junge, H.; Mögel, G.; Kunz, S.; Borriss, R. Difficidin and Bacilysin Produced by Plant-Associated Bacillus amyloliquefaciens Are Efficient in Controlling Fire Blight Disease. J. Biotechnol. 2009, 140, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Zhou, Q.N.; Luo, H.Y.; Xia, L.Q.; Li, L.; Sun, M.; Yu, Z.Q. Systemic Nematicidal Activity and Biocontrol Efficacy of Bacillus firmus Against the Root-Knot Nematode Meloidogyne incognita. World J. Microbiol. Biotechnol. 2015, 31, 661–667. [Google Scholar] [CrossRef]
- Xu, K.; Yuan, X.L.; Li, C.; Li, A.X. Recent Discovery of Heterocyclic Alkaloids from Marine-Derived Aspergillus Species. Mar. Drugs 2020, 18, 54. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wei, Q.; Yuan, X.; Xu, K. Newly Reported Alkaloids Produced by Marine-Derived Penicillium Species (covering 2014-2018). Bioorg. Chem. 2020, 99, 103840. [Google Scholar] [CrossRef]
- Mi, Y.; Zhang, J.R.; He, S.; Yan, X.J. New Peptides Isolated from Marine Cyanobacteria, an Overview over the Past Decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile Weapons for Plant Disease Biocontrol. Trends Microbiol. 2007, 16, 115–125. [Google Scholar] [CrossRef]
- Vairagkar, U.; Mirza, Y. Antagonistic Activity of Antimicrobial Metabolites Produced from Seaweed-Associated Bacillus amyloliquefaciens MTCC 10456 Against Malassezia Spp. Probiotics Antimicrob. Proteins 2021, 13, 1228–1237. [Google Scholar] [CrossRef]
- Nair, D.; Vanuopadath, M.; Nair, B.G.; Pai, J.G.; Nair, S.S. Identification and Characterization of a Library of Surfactins and Fengycins from a Marine Endophytic Bacillus Sp. J. Basic Microbiol. 2016, 56, 1159–1172. [Google Scholar] [CrossRef]
- Ma, Z.W.; Zhang, S.Y.; Zhang, S.H.; Wu, G.Y.; Shao, Y.; Mi, Q.F.; Liang, J.Y.; Hu, J.C. Isolation and Characterization of a New yclic Lipopeptide Surfactin from a Marine-Derived Bacillus velezensis SH-B74. J. Antibiot. 2020, 73, 863–867. [Google Scholar] [CrossRef]
- Chen, Y.L.; Liu, S.L.A.; Mou, H.J.; Ma, Y.X.; Li, M.; Hu, X.K. Characterization of Lipopeptide Biosurfactants Produced by Bacillus licheniformis MB01 from Marine Sediments. Front. Microbiol. 2017, 8, 871. [Google Scholar] [CrossRef]
- Ma, Z.W.; Hu, J.C. Production and Characterization of Iturinic Lipopeptides as Antifungal Agents and Biosurfactants Produced by a Marine Pinctada martensii-Derived Bacillus mojavensis B0621A. Appl. Biochem. Biotechnol. 2014, 173, 705–715. [Google Scholar] [CrossRef]
- Ma, Z.W.; Hu, J.C. Plipastatin A1 Produced by a Marine Sediment-Derived Bacillus amyloliquefaciens SH-B74 Contributes to the Control of Gray Mold Disease in Tomato. 3 Biotech 2018, 8, 125. [Google Scholar] [CrossRef]
- Tareq, F.S.; Hasan, C.M.; Lee, H.-S.; Lee, Y.-J.; Lee, J.S.; Surovy, M.Z.; Islam, M.T.; Shin, H.J. Gageopeptins A and B, New Inhibitors of Zoospore Motility of the Phytopathogen Phytophthora Capsici from a Marine-Derived Bacterium Bacillus Sp. 109GGC020. Bioorg. Med. Chem. Lett. 2015, 25, 3325–3329. [Google Scholar] [CrossRef]
- Wiese, J.; Abdelmohsen, U.R.; Motiei, A.; Humeida, U.H.; Imhoff, J.F. Bacicyclin, a New Antibacterial Cyclic Hexapeptide from Bacillus Sp. Strain BC028 Isolated from Mytilus edulis. Bioorg. Med. Chem. Lett. 2018, 28, 558–561. [Google Scholar] [CrossRef] [PubMed]
- Li, W.L.; Xia, J. Recent Advances in Diketopiperazines Biosynthesis. Microbiol. China 2012, 41, 111–121. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Lai, T.K.; Saha, A.; Selvin, J.; Mukherjee, J. Structural Elucidation and Antimicrobial Activity of a Diketopiperazine Isolated from A Bacillus Sp. Associated with the Marine Sponge Spongia officinalis. Nat. Prod. Res. 2021, 35, 2315–2323. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yu, S.Y.; Jiang, W.; Zhang, L.K.; Jin, C.L.; Zhou, X.J. Isolation and Identification of Antidiatom Attachment Active Constituents of a Strain of Epiphytic Bacillus Sponge. Mar. Sci. 2016, 40, 23–32. [Google Scholar] [CrossRef]
- Mohan, G.; Thipparamalai Thangappanpillai, A.K.; Ramasamy, B. Antimicrobial Activities of Secondary Metabolites and Phylogenetic Study of Sponge Endosymbiotic Bacteria, Bacillus Sp. at Agatti Island, Lakshadweep Archipelago. Biotechnol. Rep. 2016, 11, 44–52. [Google Scholar] [CrossRef]
- Yan, Y.H.; Li, Y.Z.; Zhang, Z.W.; Wang, X.H.; Niu, Y.Z.; Zhang, S.H.; Xu, W.L.; Ren, C.G. Advances of peptides for antibacterial applications. Colloid. Surf. B 2021, 202, 111682. [Google Scholar] [CrossRef]
- Tareq, F.S.; Lee, M.A.; Lee, H.-S.; Lee, Y.-J.; Lee, J.S.; Hasan, C.M.; Islam, M.T.; Shin, H.J. Gageotetrins A–C, Noncytotoxic Antimicrobial Linear Lipopeptides from a Marine Bacterium Bacillus subtilis. Org. Lett. 2014, 16, 928–931. [Google Scholar] [CrossRef]
- Tareq, F.S.; Lee, M.A.; Lee, H.-S.; Lee, Y.-J.; Lee, J.S.; Hasan, C.M.; Islam, M.T.; Shin, H.J. Non-Cytotoxic Antifungal Agents: Isolation and Structures of Gageopeptides A–D from a Bacillus Strain 109GGC020. J. Agric. Food Chem. 2014, 62, 5565–5572. [Google Scholar] [CrossRef]
- Gulick, A.M. Nonribosomal Peptide Synthetase Biosynthetic Clusters of ESKAPE Pathogens. Nat. Prod. Rep. 2017, 34, 981–1009. [Google Scholar] [CrossRef]
- Zhou, M.J.; Liu, F.W.; Yang, X.Y.; Jin, J.; Dong, X.; Zeng, K.-W.; Liu, D.; Zhang, Y.T.; Ma, M.; Yang, D.H. Bacillibactin and Bacillomycin Analogues with Cytotoxicities against Human Cancer Cell Lines from Marine Bacillus Sp. PKU-MA00093 and PKU-MA00092. Mar. Drugs 2018, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Tabbene, O.; Azaiez, S.; Di Grazia, A.; Karkouch, I.; Ben Slimene, I.; Elkahoui, S.; Alfeddy, M.N.; Casciaro, B.; Luca, V.; Limam, F.; et al. Bacillomycin D And Its Combination with Amphotericin B: Promising Antifungal Compounds with Powerful Antibiofilm Activity and Wound-Healing Potency. J. Appl. Microbiol. 2016, 120, 289–300. [Google Scholar] [CrossRef]
- Miyanaga, A. Structure and Function of Polyketide Biosynthetic Enzymes: Various Strategies for Production of Structurally Diverse Polyketides. Biosci. Biotechnol. Biochem. 2017, 81, 2227–2236. [Google Scholar] [CrossRef]
- Chakraborty, K.; Thilakan, B.; Raola, V.K. Previously Undescribed Antibacterial Polyketides from Heterotrophic Bacillus amyloliquefaciens Associated with Seaweed Padina gymnospora. Appl. Microbiol. Biotechnol. 2018, 184, 716–732. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Thilakan, B.; Chakraborty, R.D.; Raola, V.K.; Joy, M. O-Heterocyclic Derivatives with Antibacterial Properties from Marine Bacterium Bacillus Subtilis Associated with Seaweed, Sargassum myriocystum. Appl. Microbiol. Biotechnol. 2017, 101, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Mondol, M.A.; Shahidullah Tareq, F.; Kim, J.H.; Lee, M.A.; Lee, H.-S.; Lee, J.S.; Lee, Y.-J.; Shin, H.J. New Antimicrobial Compounds from a Marine-Derived Bacillus sp. J. Antibiot. 2013, 66, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhou, Y.-X.; Tang, X.-X.; Liu, X.-X.; Yi, Z.-W.; Fang, M.-J.; Wu, Z.; Jiang, F.-Q.; Qiu, Y.-K. Macrolactins from Marine-Derived Bacillus subtilis B5 Bacteria as Inhibitors of Inducible Nitric Oxide and Cytokines Expression. Mar. Drugs 2016, 14, 195. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.H.; Chen, X.Q.; Yu, L.; Jiang, L.; Pan, D.J.; Jiang, S.; Gan, Y.M.; Liu, Y.H.; Yi, X.X. New 24-Membered Macrolactins Isolated from Marine Bacteria Bacillus siamensis as Potent Fungal Inhibitors against Sugarcane Smut. J. Agric. Food Chem. 2021, 69, 4392–4401. [Google Scholar] [CrossRef]
- Chakraborty, K.; Thilakan, B.; Raola, V.K. Polyketide Family of Novel Antibacterial 7-O-Methyl-5’-Hydroxy-3’-Heptenoate-Macrolactin from Seaweed-Associated Bacillus subtilis MTCC 10403. J. Agric. Food Chem. 2014, 62, 12194–12208. [Google Scholar] [CrossRef]
- Wang, W.; Park, K.-H.; Lee, J.; Oh, E.; Park, C.; Kang, E.; Lee, J.; Kang, H. A New Thiopeptide Antibiotic, Micrococcin P3, from a Marine-Derived Strain of the Bacterium Bacillus stratosphericus. Molecules 2020, 25, 4383. [Google Scholar] [CrossRef]
- Zhou, S.-Y.; Hu, Y.-J.; Meng, F.-C.; Qu, S.-Y.; Wang, R.; Andersen, R.J.; Liao, Z.-H.; Chen, M. Bacillamidins A-G from a Marine-Derived Bacillus pumilus. Mar. Drugs 2018, 16, 326. [Google Scholar] [CrossRef] [Green Version]
- Tareq, F.S.; Lee, H.-S.; Lee, Y.-J.; Lee, J.S.; Shin, H.J. Ieodoglucomide C and Ieodoglycolipid, New Glycolipids from a Marine-Derived Bacterium Bacillus licheniformis 09IDYM23. Lipids 2015, 50, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Leutou, A.S.; Jeong, H.; Kim, D.; Seong, C.N.; Nam, S.-J.; Lim, K.-M. Anti-Pigmentary Effect of (-)-4-Hydroxysattabacin from the Marine-Derived Bacterium Bacillus Sp. Mar. Drugs 2017, 15, 138. [Google Scholar] [CrossRef] [PubMed]
- Lotfya, W.A.; Mostafab, S.W.; Adel, A.A.; Ghanem, K.M. Production of Di-(2-Ethylhexyl) Phthalate by Bacillus subtilis AD35: Isolation, Purifification, Characterization and Biological Activities. Microb. Pathog. 2018, 124, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.-Z.; Fu, H.-R.; Wu, S.-J.; Bao, Z.-H.; Wang, S.-F.; Ge, P.-H. Isolation and Structure Elucidation of Antifungal Metabolites from marine Paenibacillus polymyxa Strain L1–9. Acta Pharmacol. Sin. 2014, 44, 486–496. [Google Scholar] [CrossRef]
- Chakraborty, M.; Mahmud, N.U.; Gupta, D.R.; Tareq, F.S.; Shin, H.J.; Islam, T. Inhibitory Effects of Linear Lipopeptides from a Marine Bacillus subtilis on the Wheat Blast Fungus Magnaporthe oryzae Triticum. Front. Microbiol. 2020, 11, 665. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
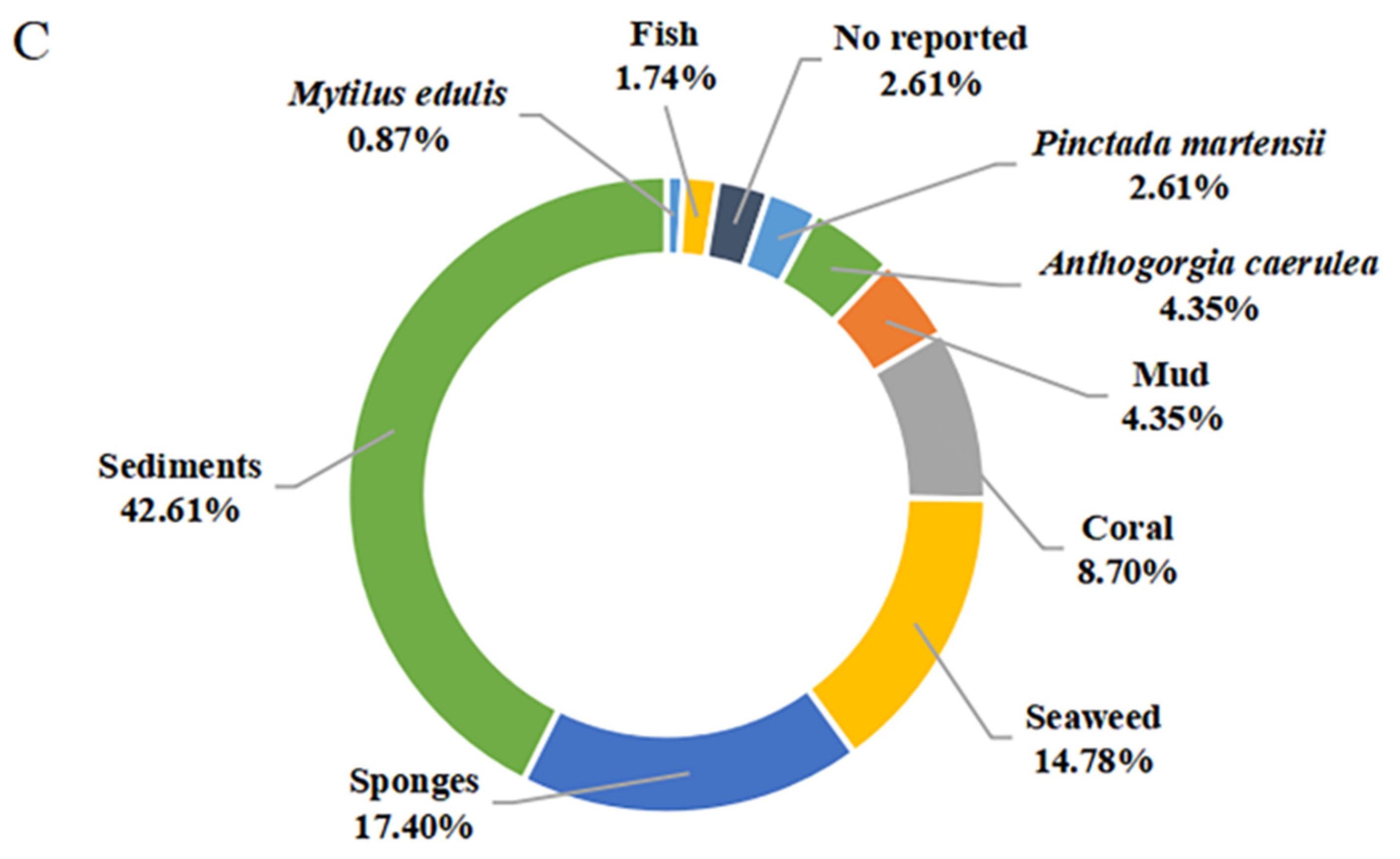{kind=link}
{kind=link}
| NO. | Classification | Producing Strain | Environmental Source | Antimicrobial Activities a | Ref. |
|---|---|---|---|---|---|
| 2 | Cyclic lipopeptide (surfactin) | B. licheniformis MB01 | sediments (the Bohai Sea, China) | V. parahaemolyticus (>3 cm), E. coli (>3 cm), V. cholerae (>3 cm), V. harveyi (>3 cm), P. aeruginosa (2–3 cm), S. aureus (2–3 cm), Proteus species (1–2 cm) (inhibition zone diameter) | [47] |
| 3 | Cyclic lipopeptide (surfactin) | B. licheniformis MB01 | sediments (the Bohai Sea, China) | V. parahaemolyticus (>3 cm), E. coli (>3 cm), V. cholerae (>3 cm), V. harveyi (>3 cm), P. aeruginosa (2–3 cm), S. aureus (2–3 cm), Proteus species (1–2 cm) (inhibition zone diameter) | [47] |
| 4 | Cyclic lipopeptide (surfactin) | B. licheniformis MB01 | sediments (the Bohai Sea, China) | V. parahaemolyticus (>3 cm), E. coli (>3 cm), V. cholerae (>3 cm), V. harveyi (>3 cm), P. aeruginosa (2–3 cm), S. aureus (2–3 cm), Proteus species (1–2 cm) (inhibition zone diameter) | [47] |
| 5 | Cyclic lipopeptide (surfactin) | B. licheniformis MB01 | sediments (the Bohai Sea, China) | V. parahaemolyticus (>3 cm), E. coli (>3 cm), V. cholerae (>3 cm), V. harveyi (>3 cm), P. aeruginosa (2–3 cm), S. aureus (2–3 cm), Proteus species (1–2 cm) (inhibition zone diameter) | [47] |
| 15 | Cyclic lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | MRSA ATCC25923 (8 μg/mL), MRSA XU2120 (16 μg/mL), MRSA SA1199B (8 μg/mL), MRSA RN4220 (32 μg/mL) (MIC) | [12] |
| 16 | Cyclic lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | MRSA ATCC25923 (16 μg/mL), MRSA XU2120 (16 μg/mL), MRSA SA1199B (32 μg/mL) (MIC) | [12] |
| 17 | Cyclic lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (16 μg/mL), B. subtilis (16 μg/mL), S. typhi (16 μg/mL), P. aeruginosa (16 μg/mL) (MIC) | [50] |
| 18 | Cyclic lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (16 μg/mL), B. subtilis (32 μg/mL), S. typhi (32 μg/mL), P. aeruginosa (16 μg/mL) (MIC) | [50] |
| 19 | Cyclic lipopeptide | Bacillus sp. BC028 | the blue mussel M. edulis (the western shore of Baltic Sea, Germany) | S. aureus NCTC 8325 (12 μM), E. faecalis JH212 (8 μM) (MIC) | [51] |
| 20 | Cyclic lipopeptide | Bacillus sp. SPB7 | the sponge S. officinali (the Palk Bay of Bengal, Mandapam coast, Tamil Nadu, India) | E. coli (16 μg/mL), S. aureus (22 μg/mL) (MIC) | [53] |
| 25 | Pyrrol | Bacillus sp. | sponges (Agatti island, | fish pathogens (GI50, 31.25 µg/mL) | [55] |
| Lakshadweep archipelago, India) | |||||
| 28 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. typhi (16 μg/mL), S. aureus (16 μg/mL), P. aeruginosa (16 μg/mL), B. subtilis (16 μg/mL) (MIC) | [35] |
| 29 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. typhi (32 μg/mL), S. aureus (16 μg/mL), P. aeruginosa (16 μg/mL), B. subtilis (32 μg/mL) (MIC) | [35] |
| 30 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. typhi (32 μg/mL), S. aureus (64 μg/mL), P. aeruginosa (64 μg/mL), B. subtilis (32 μg/mL) (MIC) | [35] |
| 31 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.03 µM), B. subtilis (0.03 µM), S. typhi (0.06 µM), P. aeruginosa (0.06 µM) (MIC) | [57] |
| 32 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.04 µM), B. subtilis (0.02 µM), S. typhi (0.02 µM), P. aeruginosa (0.04 µM) (MIC) | [57] |
| 33 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.04 µM), B. subtilis (0.04 µM), S. typhi (0.02 µM), P. aeruginosa (0.02 µM) (MIC) | [57] |
| 34 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.05 µM), B. subtilis (0.05 µM), S. typhi (0.05 µM), P. aeruginosa (0.09 µM) (MIC) | [58] |
| 35 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.05 µM), B. subtilis (0.05 µM), S. typhi (0.08 µM), P. aeruginosa (0.08 µM) (MIC) | [58] |
| 36 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.08 µM), B. subtilis (0.09 µM), S. typhi (0.09 µM), P. aeruginosa (0.09 µM) (MIC) | [58] |
| 37 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | S. aureus (0.08 µM), B. subtilis (0.05 µM), S. typhi (0.09 µM), P. aeruginosa (0.08 µM) (MIC) | [58] |
| 52 | Polyketide | B. subtilis | the brown seaweed S. myriocystum (Mannar Bay on the southeast coast of India) | V. parahemolyticus (24.33 ± 0.58 mm, 10 μg on disk), V. vulnificus (22.66 ± 0.58 mm), A. hydrophila (26.00 ± 1.00 mm) (inhibition zone diameter) | [64] |
| MTCC 10407 | |||||
| 53 | Polyketide | B. subtilis | the brown seaweed S. myriocystum (Mannar Bay on the southeast coast of India) | V. parahemolyticus (11.00 ± 1.00 mm), V. vulnificus (22.66 ± 0.58 mm), A. hydrophila (17.66 ± 0.58 mm) (inhibition zone diameter) | [64] |
| MTCC 10407 | |||||
| 54 | Polyketide | B. amyloliquefaciens | the brown seaweed P. gymnospora (Mannar Bay, Peninsular India) | V. vulnificus MTCC 1145 (16.33 ± 0.58 mm, 10 mcg on disk), V. parahaemolyticus ATCC® 17802™ (15.30 ± 1.15 mm), A. hydrophila MTCC 646 (12.67 ± 1.15 mm), V. harveyi MTCC 3438 (15.33 ± 0.58 mm), V. anguillarum (13.33 ± 1.15 mm), V. parahaemolyticus MTCC 451 (14.00 ± 1.00 mm) (inhibition zone diameter) | [63] |
| 55 | Polyketide | B. amyloliquefaciens | the brown seaweed P. gymnospora (Mannar Bay, Peninsular India) | V. vulnificus MTCC 1145 (14.67 ± 1.15 mm), V. parahaemolyticus ATCC® 17802™ (14.00 ± 1.00 mm), A. hydrophila MTCC 646 (14.67 ± 1.15 mm), V. harveyi MTCC 3438 (13.00 ± 1.00 mm), V. anguillarum (12.67 ± 1.54 mm), V. parahaemolyticus MTCC 451 (13.33 ± 1.15 mm) (inhibition zone diameter) | [63] |
| 56 | Polyketide | B. amyloliquefaciens | the brown seaweed P. gymnospora (Mannar Bay, Peninsular India) | V. vulnificus MTCC 1145 (17.33 ± 1.15 mm), V. parahaemolyticus ATCC® 17802™ (16.00 ± 1.00 mm), A. hydrophila MTCC 646 (13.33 ± 1.15 mm), V. harveyi MTCC 3438 (12.67 ± 1.15 mm), V. anguillarum (10.66 ± 1.15 mm), V. parahaemolyticus MTCC 451 (15.33 ± 0.58 mm) (inhibition zone diameter) | [63] |
| 57 | Polyketide | B. amyloliquefaciens | the brown seaweed P. gymnospora (Mannar Bay, Peninsular India) | V. vulnificus MTCC 1145 (13.67 ± 1.52 mm), V. parahaemolyticus ATCC® 17802™ (12.66 ± 1.15 mm), A. hydrophila MTCC 646 (11.33 ± 1.15 mm), V. harveyi MTCC 3438 (15.33 ± 0.58 mm), V. parahaemolyticus MTCC 451 (12.67 ± 1.54 mm) (inhibition zone diameter) | [63] |
| 68 | Macrolactin | B. subtilis MTCC10403 | the seaweed Anthophycus longifolius (Mannar Bay, Peninsular India) | A. hydrophilla (18 mm, 100 μg on disk), V. parahemolyticus ATCC 17802 (16 mm), V. vulnificus (14 mm) (inhibition zone diameter) | [68] |
| 69 | Macrolactin (difficidin) | B. amyloliquefaciens MTCC12713 | an intertidal macroalga Kappaphycus alverezii (Mannar Bay, Peninsular India) | MRSA ATCC33592 (0.005 µM), VREfs ATCC51299 (0.009 µM), P. aeruginosa ATCC27853 (0.006 µM), K. pneumonia ATCC13883 (0.009 µM), E. tarda MTCC2400 (0.005 µM), E. coli MTCC443 (0.009 µM), S. pyogenes MTCC1924 (0.009 µM), V. parahaemolyticus MTCC451 (0.006 µM) (MIC) | [13] |
| 70 | Macrolactin (difficidin) | B. amyloliquefaciens MTCC12713 | an intertidal macroalga Kappaphycus alverezii (Mannar Bay, Peninsular India) | MRSA ATCC33592 (0.002 µM), VREfs ATCC51299 (0.002 µM), P. aeruginosa ATCC27853 (0.004 µM), K. pneumonia ATCC13883 (0.002 µM), E. tarda MTCC2400 (0.004 µM), E. coli MTCC443 (0.004 µM), S. pyogenes MTCC1924 (0.007 µM), V. parahaemolyticus MTCC451 (0.004 µM) (MIC) | [13] |
| 71 | Macrolactin (difficidin) | B. amyloliquefaciens MTCC12713 | an intertidal macroalga Kappaphycus alverezii (Mannar Bay, Peninsular India) | MRSA ATCC33592 (0.002 µM), VREfs ATCC51299 (0.004 µM), P. aeruginosa ATCC27853 (0.002 µM), K. pneumonia ATCC13883 (0.002 µM), E. tarda MTCC2400 (0.004 µM), E. coli MTCC443 (0.004 µM), S. pyogenes MTCC1924 (0.005 µM), V. parahaemolyticus MTCC451 (0.002 µM) (MIC) | [13] |
| 72 | Macrolactin (difficidin) | B. amyloliquefaciens MTCC12713 | an intertidal macroalga Kappaphycus alverezii (Mannar Bay, Peninsular India) | MRSA ATCC33592 (0.002 µM), VREfs ATCC51299 (0.004 µM), P. aeruginosa ATCC27853 (0.002 µM), K. pneumonia ATCC13883 (0.002 µM), E. tarda MTCC2400 (0.004 µM), E. coli MTCC443 (0.004 µM), S. pyogenes MTCC1924 (0.004 µM), V. parahaemolyticus MTCC451 (0.002 µM) (MIC) | [13] |
| 73 | Thiopeptide | B. stratosphericus | no description | S. aureus KCTC 1927 (0.8 µg/mL), K. rhizophila KCTC 1915 (0.2 µg/mL), B. subtilis KCTC 1021 (0.8 µg/mL), E. coli KCTC 2441 (26 µg/mL), K. pneumoniae KCTC 2690 (26 µg/mL), S. typhimurium KCTC 2515 (26 µg/mL) (MIC) | [69] |
| 74 | Thiopeptide | B. stratosphericus | no description | S. aureus KCTC 1927 (0.1 µg/mL), K. rhizophila KCTC 1915 (0.05 µg/mL), B. subtilis KCTC 1021 (0.5 µg/mL), E. coli KCTC 2441 (26 µg/mL), K. pneumoniae KCTC 2690 (26 µg/mL), S. typhimurium KCTC 2515 (26 µg/mL) (MIC) | [69] |
| 75 | Long-chain amide | B. pumilus RJA1515 | sediments (at a depth of 84 m, Bamfield, British Columbia) | P. aeruginosa PA-01 (64 µg/mL), A. baumannii ATCC19606 (58 µg/mL) (MIC) | [70] |
| 76 | Long-chain amide | B. pumilus RJA1515 | sediments (at a depth of 84 m, Bamfield, British Columbia) | P. aeruginosa PA-01 (64 µg/mL), A. baumannii ATCC19606 (64 µg/mL) (MIC) | [70] |
| 77 | Long-chain amide | B. pumilus RJA1515 | sediments (at a depth of 84 m, Bamfield, British Columbia) | P. aeruginosa PA-01 (64 µg/mL), A. baumannii ATCC19606 (64 µg/mL) (MIC) | [70] |
| 78 | Long-chain amide | B. pumilus RJA1515 | sediments (at a depth of 84 m, Bamfield, British Columbia) | P. aeruginosa PA-01 (64 µg/mL), A. baumannii ATCC19606 (58 µg/mL) (MIC) | [70] |
| 82 | Glycolipid | B. licheniformis 09IDYM23 | sediments (at a depth 20 m, Ieodo, Korea) | P. aeruginosa (0.01 µM), E. coli (0.01 µM), B. cereus (0.01 µM), B. subtilis (0.03 µM), S. typhi (0.01 µM), S. aureus (0.03 µM) (MIC) | [71] |
| 83 | Glycolipid | B. licheniformis 09IDYM23 | sediments (at a depth 20 m, Ieodo, Korea) | P. aeruginosa (0.03 µM), E. coli (0.03 µM), B. cereus (0.03 µM), B.subtilis (0.05 µM), S. typhi (0.05 µM), S. aureus (0.03 µM) (MIC) | [71] |
| 86 | Other compounds | B. subtilis AD35 | marine water and sediment (Alexandria sea shore, Egypt) | S. typhimurium (16 µg/mL), MRSA (32 µg/mL), L. monocytogenes (0.25 µg/mL), A. hydrophila (0.5 µg/mL), S. aureus (8 µg/mL), S. epidermidis (4 µg/mL), E. coli (4 µg/mL), P. aeruginosa (8 µg/mL) (GI50) | [73] |
| B. polymyxa L1-9 | the mud of the intertidal mudflat (Lianyungang Port, China) | E. coli (5.2 ± 0.20 mm), B. subtilis (4.3 ± 0.11 mm), S. aureus (1.3 ± 0.02 mm) (inhibition zone diameter) | [74] | ||
| 87 | Other compounds | B. polymyxa L1-9 | the mud of the intertidal mudflat (Lianyungang Port, China) | E. coli (9.7 ± 0.20 mm), B. subtilis (6.6 ± 0.16 mm), S. aureus (2.3 ± 0.12 mm) (inhibition zone diameter) | [74] |
| NO. | Classification | Producing Strain | Environmental Source | Antifungal Activities a | Ref. |
|---|---|---|---|---|---|
| 1 | Cyclic lipopeptide (surfactin) | B. velezensis SH-B74 | sediments (CCTCC) | M. oryzae (concentration, 10–50 µM) | [46] |
| 6 | Cyclic lipopeptide (iturin) | B. amyloliquefaciens SH-B74 | sediments (the South China Sea, China) | V. mali, F. oxysporum f.sp. cucumerinum, R. solani (concentration, >2 mg/mL) | [22] |
| 8 | Cyclic lipopeptide (iturin) | B. mojavensis B0621A | the pearl oyster P. martensii (the South China Sea, China) | F. oxysporum f.sp. cucumerinum (1.0 mM, 5.63 ± 0.03 mm; 2.0 mM, 6.41 ± 0.56 mm), F. oxysporum f.sp. vasinfectum (2.0 mM, 5.94 ± 0.25 mm), F. oxysporum f.sp. vasinfectum SF2 (2.0 mM, 6.21 ± 0.36 mm) (inhibition zone diameter) | [48] |
| 9 | Cyclic lipopeptide (iturin) | B. mojavensis B0621A | the pearl oyster P. martensii (the South China Sea, China) | F. oxysporum f.sp. cucumerinum (0.5 Mm, 6.45 ± 0.13 mm; 1.0 mM, 6.52 ± 0.30 mm; 2.0 mM, 7.33 ± 0.24 mm), F. oxysporum f.sp. vasinfectum (0.5 mM, 6.46 ± 0.30 mm; 1.0 mM, 7.07 ± 0.10 mm; 2.0 mM, 7.33 ± 0.07 mm), F. oxysporum f.sp. vasinfectum SF2 (0.5 mM, 5.62 ± 0.40 mm; 1.0 mM, 6.63 ± 0.43 mm; 2.0 mM, 7.19 ± 0.34 mm), F. oxysporum f.sp. Cucumis melo L. (2.0 mM, 6.32 ± 0.37 mm) (inhibition zone diameter) | [48] |
| 10 | Cyclic lipopeptide (iturin) | B. mojavensis B0621A | the pearl oyster P. martensii (the South China Sea, China) | F. oxysporum f.sp. cucumerinum (0.5 mM, 5.43 ± 0.23 mm; 1.0 mM, 6.61 ± 0.31 mm; 2.0 mM, 8.02 ± 0.04 mm), F. oxysporum f.sp. vasinfectum (0.5 mM, 5.31 ± 0.17 mm; 1.0 mM, 6.12 ± 0.43 mm; 2.0 mM, 7.26 ± 0.45 mm), F. oxysporum f.sp. vasinfectum SF2 (0.5 mM, 5.45 ± 0.24 mm; 1.0 mM, 6.31 ± 0.20 mm; 2.0 mM, 6.89 ± 0.26 mm) (inhibition zone diameter) | [48] |
| 11 | Cyclic lipopeptide (plipastatin) | B. amyloliquefaciens SH-B74 | sediments (CCTCC) | B. cinerea (MIC, 50 µM) | [49] |
| 17 | Cyclic lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | P. capsici (IC50, 1 μg/mL), R. solani (MIC, 4 µg/mL), C. acutatum (MIC, 8 µg/mL), B. cinerea (MIC, 4 µg/mL) | [50] |
| 18 | Cyclic lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | P. capsici (IC50, 400 μg/mL), R. solani (MIC, 8 µg/mL), C. acutatum (MIC, 8 µg/mL), B. cinerea (MIC, 8 µg/mL) | [50] |
| 28 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | C. acutatum (8 μg/mL), B. cinerea (4 μg/mL), R. solani (4 μg/mL) (MIC) | [35] |
| 29 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | C. acutatum (8 μg/mL), B. cinerea (8 μg/mL), R. solani (8 μg/mL) (MIC) | [35] |
| 30 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | C. acutatum (16 μg/mL), B. cinerea (32 μg/mL), R. solani (32 μg/mL) (MIC) | [35] |
| 31 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | B. cinerea (0.03 µM), C. acutatum (0.03 µM), R. solani (0.06 µM) (MIC) | [57] |
| 32 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | B. cinerea (0.01 µM), C. acutatum (0.01 µM), R. solani (0.02 µM) (MIC) | [57] |
| MoT (MIC, 1.5 µg/disk) | [75] | ||||
| 33 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | B. cinerea (0.01 µM), C. acutatum (0.02 µM), R. solani (0.02 µM) (MIC) | [57] |
| 34 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | R. solani (0.02 µM), B. cinerea (0.06 µM), C. acutatum (0.04 µM) (MIC) | [58] |
| MoT (MIC, 10.0 µg/disk) | [75] | ||||
| 35 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | R. solani (0.04 µM), B. cinerea (0.06 µM), C. acutatum (0.04 µM) (MIC) | [58] |
| MoT (MIC, 10.0 µg/disk) | [75] | ||||
| 36 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | R. solani (0.08 µM), B. cinerea (0.08 µM), C. acutatum (0.06 µM) (MIC) | [58] |
| MoT (MIC, 2.5 µg/disk) | [75] | ||||
| 37 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | R. solani (0.08 µM), B. cinerea (0.04 µM), C. acutatum (0.06 µM) (MIC) | [58] |
| MoT (MIC, 2.5 µg/disk) | [75] | ||||
| 38 | Linear lipopeptide | B. amyloliquefaciens MTCC 10456 | seaweed (MTCC, Chandigarh, India) | M. furfur ATCC 44344 (50–100 µg/mL), M. furfur ATCC 12078 (50–110 µg/mL), M. globosa ATCC MYA 4612 (30–100 µg/mL) (MIC) | [44] |
| 47 | Nonribosomal peptide | B. subtilis B38 | not reported | C. albicans ATCC 10231 (MIC, 12.5 µg/mL) | [61] |
| 60 | Macrolactin | B. amyloliquefaciens MTCC 10456 | seaweed (MTCC, Chandigarh, India) | M. furfur ATCC 44344 (156–313 µg/mL), M. furfur ATCC 12078 (165–330 µg/mL), M. globosa ATCC MYA 4612 (138–275 µg/mL) (MIC) | [44] |
| 62 | Macrolactin | B. amyloliquefaciens MTCC 10456 | seaweed (MTCC, Chandigarh, India) | M. furfur ATCC 44344 (156–313 µg/mL), M. furfur ATCC 12078 (165–330 µg/mL), M. globosa ATCC MYA 4612 (138–275 µg/mL) (MIC) | [44] |
| 63 | Macrolactin | B. siamensis | the Anthogorgia caerulea (Beihai city, Guangxi, China) | S. scitamineum (EC50, 67.99 μg/mL) | [68] |
| 64 | Macrolactin | B. siamensis | the Anthogorgia caerulea (Beihai city, Guangxi, China) | S. scitamineum (EC50, 12.51 μg/mL) | [68] |
| 65 | Macrolactin | B. siamensis | the Anthogorgia caerulea (Beihai city, Guangxi, China) | S. scitamineum (EC50, 3.25 μg/mL) | [68] |
| 66 | Macrolactin | B. siamensis | the Anthogorgia caerulea (Beihai city, Guangxi, China) | S. scitamineum (EC50, 15.05 μg/mL) | [68] |
| 67 | Macrolactin | B. siamensis | the Anthogorgia caerulea (Beihai city, Guangxi, China) | S. scitamineum (EC50, 34.28 μg/mL) | [68] |
| 82 | Glycolipid | B. licheniformis 09IDYM23 | sediments (at a depth 20 m, Ieodo, Korea) | B. cinerea (0.03 µM), R. solani (0.03 µM), C. acutatum (0.03 µM), A. niger (0.05 µM) (IC50), C. albicans (0.05µM) (MIC) | [71] |
| 83 | Glycolipid | B. licheniformis 09IDYM23 | sediments (at a depth 20 m, Ieodo, Korea) | B. cinerea (0.05 µM), R. solani (0.03 µM), C. acutatum (0.03 µM), A.niger (0.05 µM) (IC50), C. albicans (0.03µM) (MIC) | [71] |
| 86 | Other compounds | B. polymyxa L1-9 | the mud of the intertidal mudflat (Lianyungang Port, China) | S. sclerotiorum (9.0 ± 0.3 mm), B. sorokiniana (8.7 ± 0.10 mm), R. solani (7.2 ± 0.11 mm), A. solani (6.0 ± 0.13 mm), B. maydis (5.3 ± 0.06 mm) (inhibition zone diameter) | [74] |
| 87 | Other compounds | B. polymyxa L1-9 | the mud of the intertidal mudflat (Lianyungang Port, China) | S. sclerotiorum (11.0 ± 0.11 mm), B. sorokiniana (10.4 ± 0.21 mm), R. solani (9.8 ± 0.12 mm), A. solani (8.4 ± 0.14 mm), B. maydis (7.2 ± 0.06 mm) (inhibition zone diameter) | [74] |
| NO. | Classification | Producing Strain | Environmental Source | Cytotoxic Activities a | Ref. |
|---|---|---|---|---|---|
| 12 | Cyclic lipopeptide | Streptomyces sp. and Bacillus sp. | the mud (the intertidal mudflat in Wando, Korea) | SK-HEP-1 (27 μM), HCT116 (28 μM), MDA-MB-231 (28 μM), A549 (38 μM), SNU638 (39 μM) (IC50) | [25] |
| 28 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | MDA-MB-231 (14.9 µg/mL), HCT-15 (11.4 µg/mL), PC-3 (10.8 µg/mL), NCI-H23 (11.2 µg/mL), NUGC-3 (11.8 µg/mL), ACHN (11.5 µg/mL) (GI50) | [35] |
| 29 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | MDA-MB-231 (16.1 µg/mL), HCT-15 (18.3 µg/mL), PC-3 (19.4 µg/mL), NCI-H23 (11.7 µg/mL), NUGC-3(13.9 µg/mL), ACHN (18.4 µg/mL) (GI50) | [35] |
| 30 | Linear lipopeptide | B. subtilis 109GGC020 | sediments (the Gageocho reef, Korea) | MDA-MB-231 (11.2 µg/mL), HCT-15 (23.2 µg/mL), PC-3 (11.7 µg/mL), NCI-H23 (10.9 µg/mL), NUGC-3 (10.5 µg/mL), ACHN (12.3 µg/mL) (GI50) | [35] |
| 45 | Nonribosomal peptide | Bacillus sp. PKU-MA00092 | sponges, corals and sediments (the South China Sea and the southern Coast of China) | MCF7 (4.2 ± 0.1 μM), HepG2 (8.2 ± 0.2 μM) (IC50) | [60] |
| 46 | Nonribosomal peptide | Bacillus sp. PKU-MA00093 | sponges, corals and sediments (the South China Sea and the southern Coast of China) | MCF7 (2.9 ± 0.1 μM), HepG2 (5.1 ± 0.2 μM) (IC50) | [60] |
| 47 | Nonribosomal peptide | Bacillus sp. PKU-MA00092 | sponges, corals and sediments (the South China Sea and the southern Coast of China) | MCF7 (3.3 ± 0.1 μM), HepG2 (4.9 ± 0.2 μM) (IC50) | [60] |
| 48 | Nonribosomal peptide | Bacillus sp. PKU-MA00092 | sponges, corals and sediments (the South China Sea and the southern Coast of China) | MCF7 (7.2 ± 0.2 μM) (IC50) | [60] |
| 84 | Other compounds | Bacillus sp. (SCO-147) | sediments (Suncheon Bay of Korea) | inhibitory activity (human melanoma cell line MNT-1 and melanin synthesis in mouse melanoma cell line B16F10) | [72] |
| 85 | Other compounds | Bacillus sp. (SCO-147) | sediments (Suncheon Bay of Korea) | inhibitory activity (human melanoma cell line MNT-1 and melanin synthesis in mouse melanoma cell line B16F10), enzyme inhibitory activity, and anti-pigmentary activity | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, S.; Chen, N.; Chai, Z.; Zhou, M.; Xiao, C.; Zhao, S.; Yang, X. Secondary Metabolites from Marine-Derived Bacillus: A Comprehensive Review of Origins, Structures, and Bioactivities. Mar. Drugs 2022, 20, 567. https://doi.org/10.3390/md20090567
Xiao S, Chen N, Chai Z, Zhou M, Xiao C, Zhao S, Yang X. Secondary Metabolites from Marine-Derived Bacillus: A Comprehensive Review of Origins, Structures, and Bioactivities. Marine Drugs. 2022; 20(9):567. https://doi.org/10.3390/md20090567
Chicago/Turabian StyleXiao, Shaoyujia, Nan Chen, Zixue Chai, Mengdie Zhou, Chenghaotian Xiao, Shiqin Zhao, and Xiliang Yang. 2022. "Secondary Metabolites from Marine-Derived Bacillus: A Comprehensive Review of Origins, Structures, and Bioactivities" Marine Drugs 20, no. 9: 567. https://doi.org/10.3390/md20090567