
Astaxanthin Confers a Significant Attenuation of Hippocampal Neuronal Loss Induced by Severe Ischemia-Reperfusion Injury in Gerbils by Reducing Oxidative Stress

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Neuroprotection by ATX
2.1.1. Findings by Nissl Staining
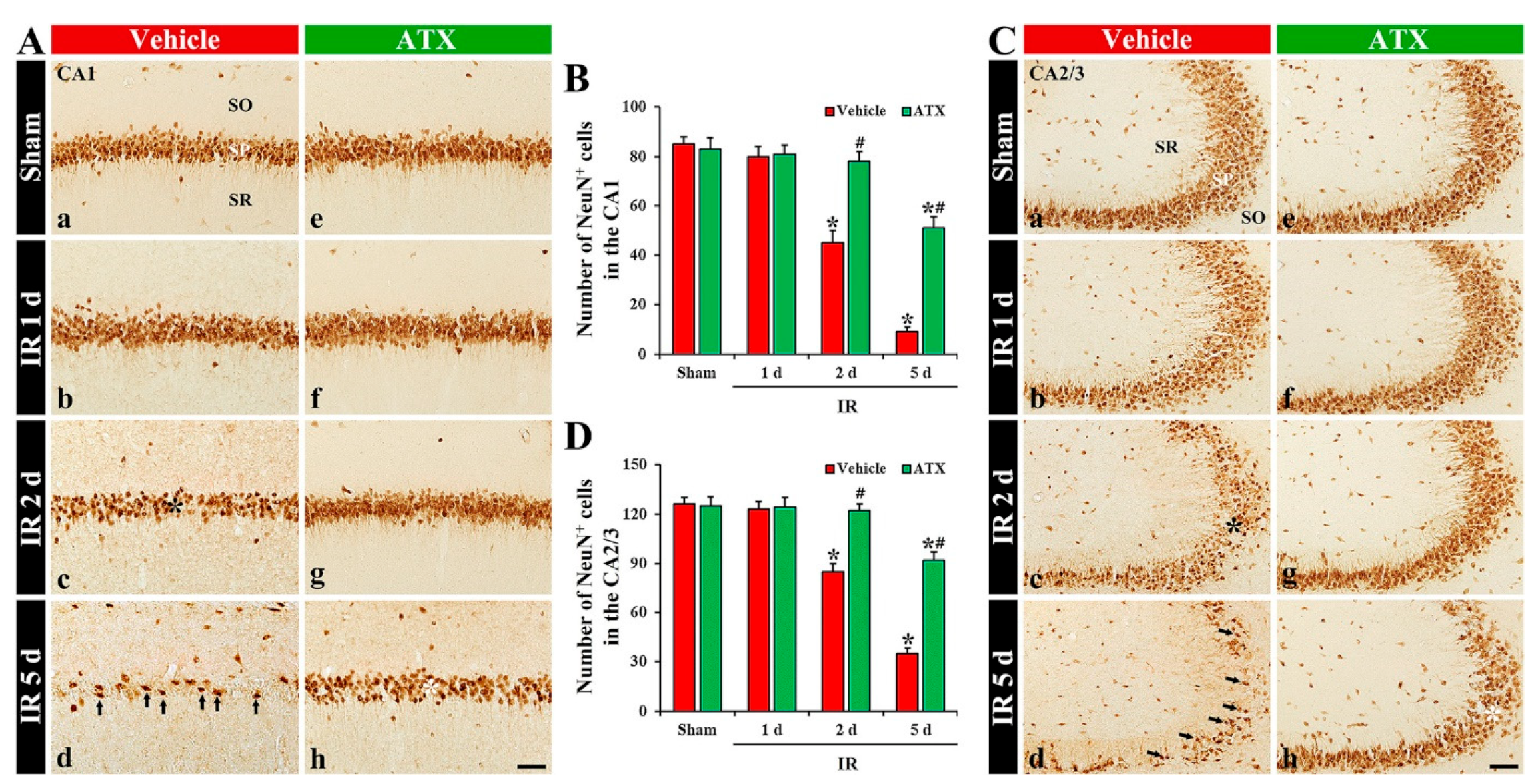
2.1.2. Findings by Neuronal Nuclei (NeuN, a Marker for Mature Neurons) Immunohistochemistry
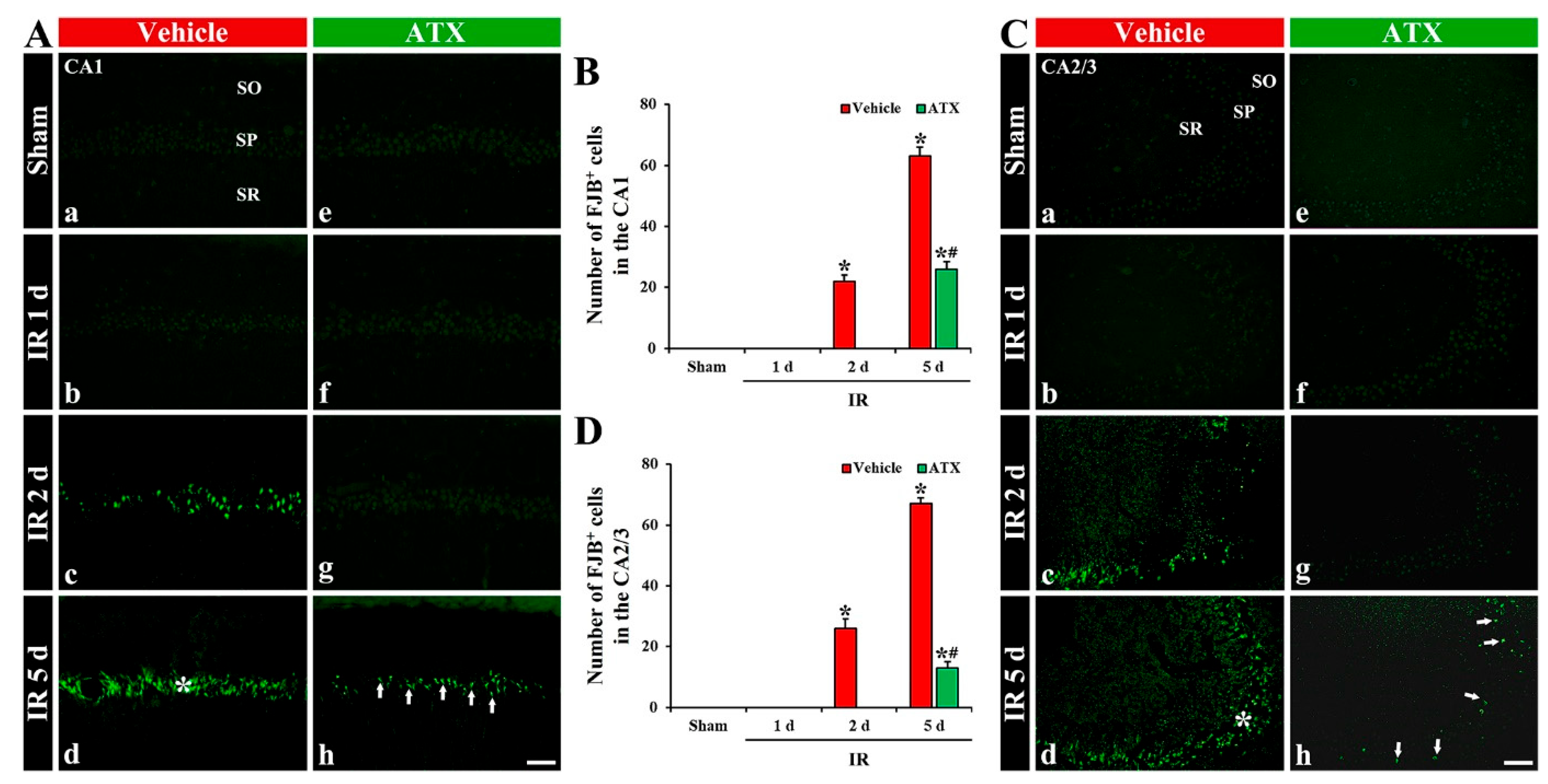
2.1.3. Findings by Fluoro-Jade B (FJB) Histofluorescence
2.2. Attenuation of Oxidative DNA Damage and Lipid Peroxidation by ATX
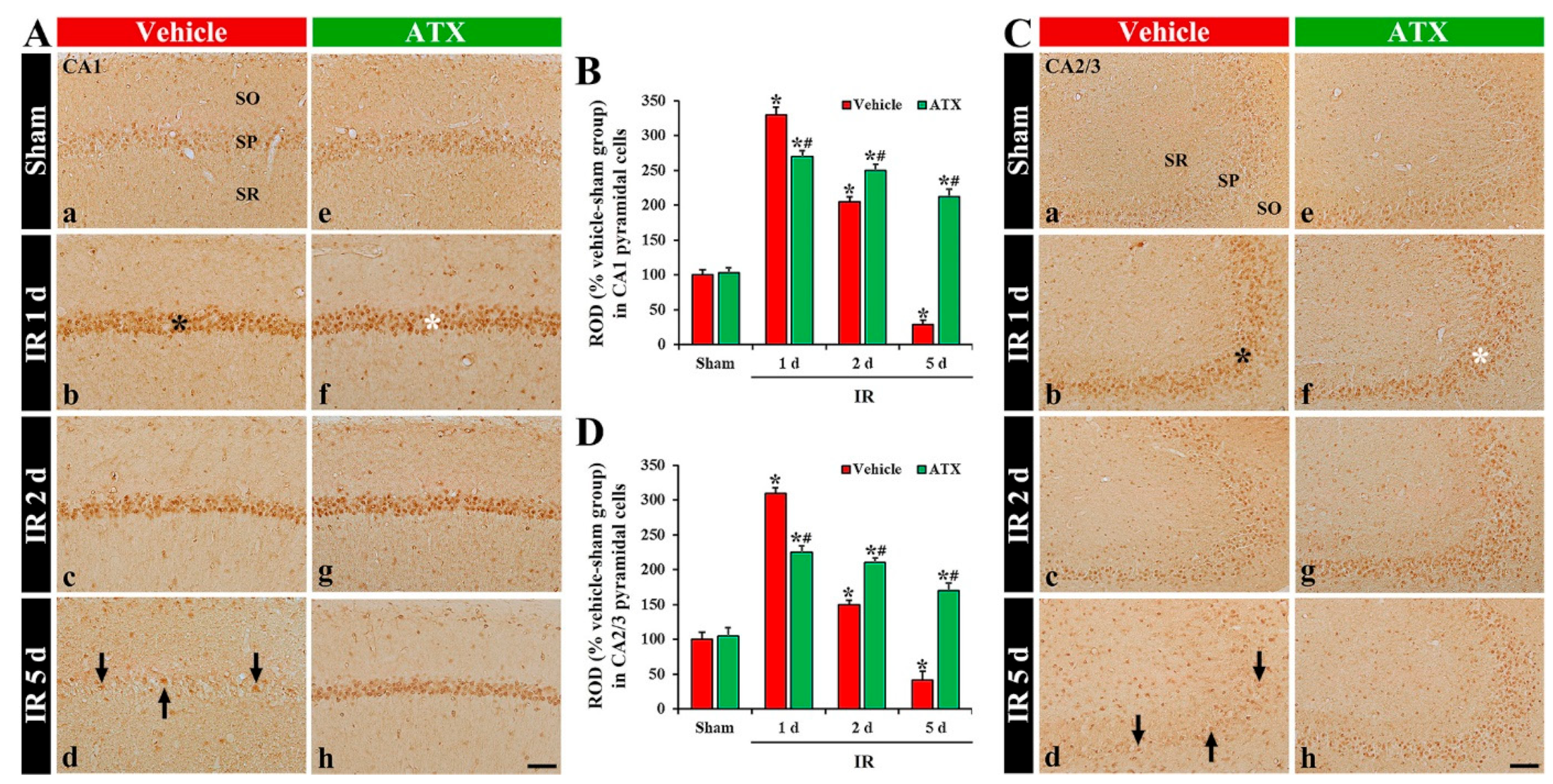
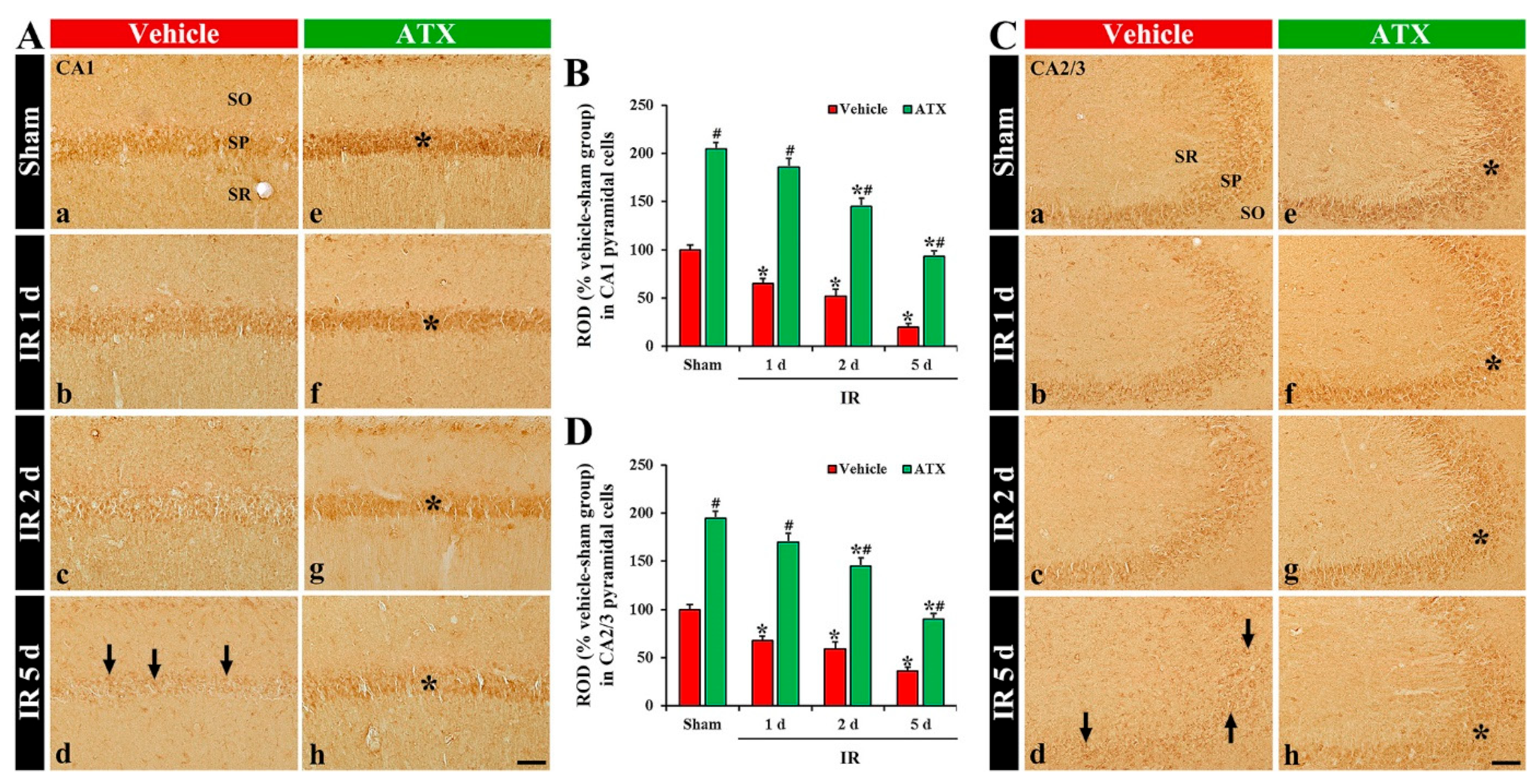
2.2.1. 8-Hydroxydeoxyguanosine (8OHdG a Marker of Oxidative DNA Damage) Immunoreactivity
2.2.2. 4-Hydroxy-2-Nonenal (4HNE, a Marker for Lipid Peroxidation) Immunoreactivity
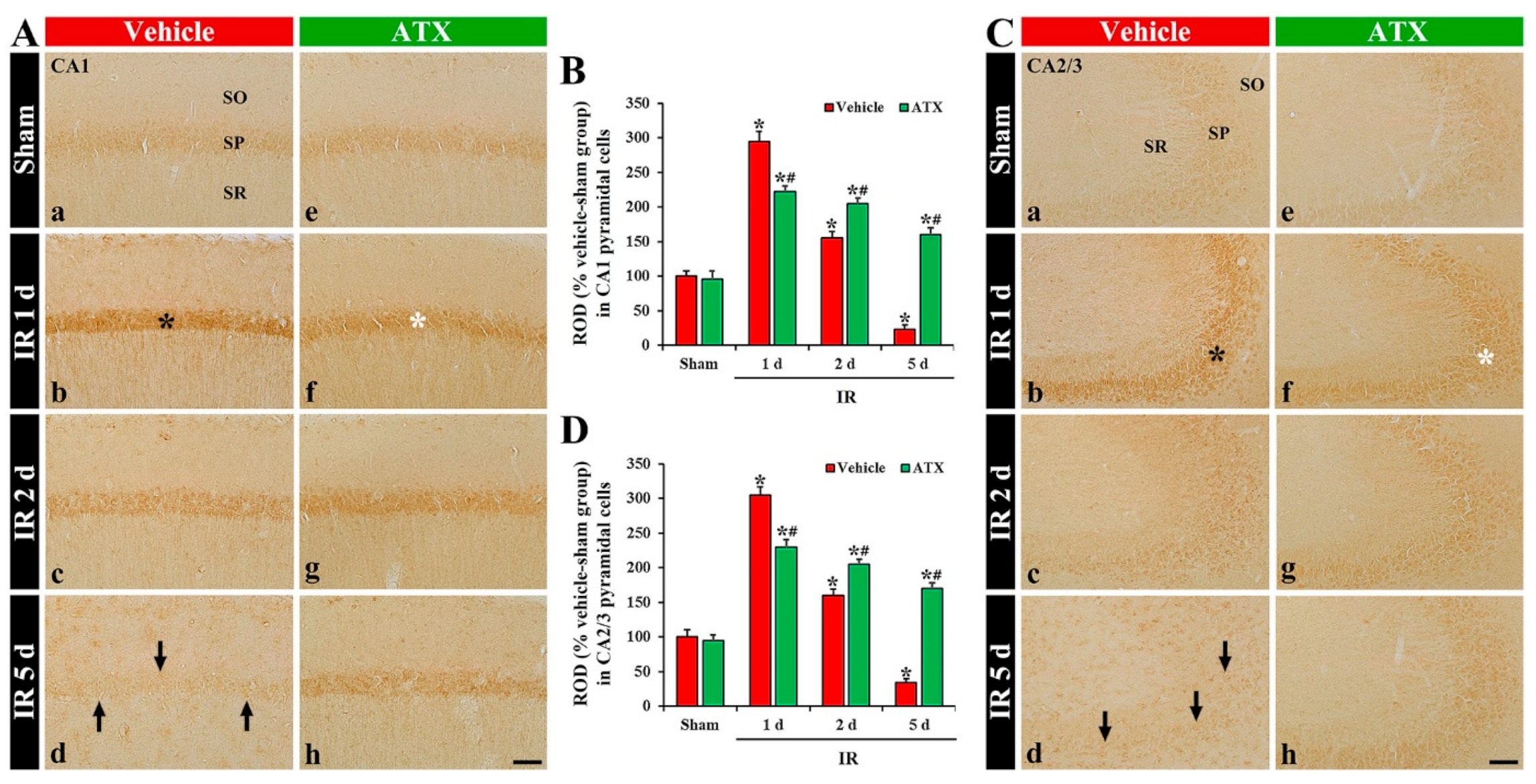
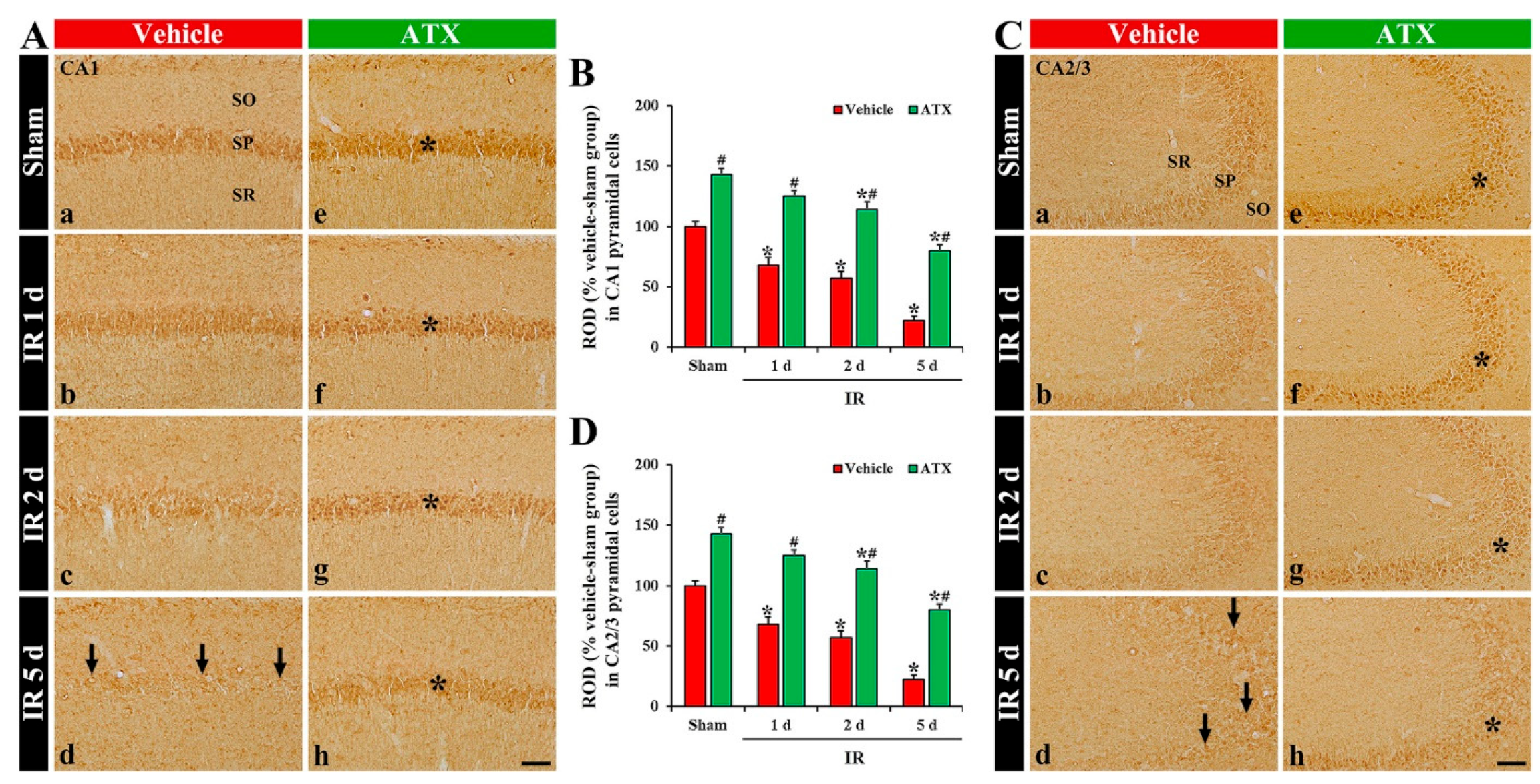
2.3. Increase of SOD1 and SOD2 Expressions by ATX
2.3.1. SOD1 Immunoreactivity
2.3.2. SOD2 Immunoreactivity
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Groups and Treatment of ATX
4.3. Induction of Severe IR Injury
4.4. Preparation of Histological Sections
4.5. Nissl Staining
4.6. FJB Staining
4.7. Immunohistochemistry
4.8. Analyses of Data
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4HNE | 4-hydroxy-2-nonenal |
| 8OHdG | 8-hydroxydeoxyguanosine |
| ATX | astaxanthin |
| CA | cornu Ammonis |
| CV | cresyl violet |
| DG | dentate gyrus |
| FJB | Fluoro-Jade B |
| IR | ischemia and reperfusion |
| NeuN | neuronal nuclei |
| ROD | relative optical density |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| SO | stratum oriens |
| SP | stratum pyramidale |
| SR | stratum radiatum |
References
- Kawai, K.; Nitecka, L.; Ruetzler, C.A.; Nagashima, G.; Joo, F.; Mies, G.; Nowak, T.S., Jr.; Saito, N.; Lohr, J.M.; Klatzo, I. Global cerebral ischemia associated with cardiac arrest in the rat: I. Dynamics of early neuronal changes. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 1992, 12, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahul, A.B.; Joshi, P.C.; Kumar, A.; Chakravarty, S. Transient global cerebral ischemia differentially affects cortex, striatum and hippocampus in bilateral common carotid arterial occlusion (bccao) mouse model. J. Chem. Neuroanat. 2018, 92, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Globus, M.Y.; Busto, R.; Martinez, E.; Valdes, I.; Dietrich, W.D.; Ginsberg, M.D. Comparative effect of transient global ischemia on extracellular levels of glutamate, glycine, and gamma-aminobutyric acid in vulnerable and nonvulnerable brain regions in the rat. J. Neurochem. 1991, 57, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Park, J.H.; Ahn, J.H.; Kim, I.H.; Cho, J.H.; Choi, J.H.; Yoo, K.Y.; Lee, C.H.; Hwang, I.K.; Cho, J.H.; et al. New gabaergic neurogenesis in the hippocampal ca1 region of a gerbil model of long-term survival after transient cerebral ischemic injury. Brain Pathol. 2016, 26, 581–592. [Google Scholar] [CrossRef]
- Ahn, J.H.; Ohk, T.G.; Kim, D.W.; Kim, H.; Song, M.; Lee, T.K.; Lee, J.C.; Yang, G.E.; Shin, M.C.; Cho, J.H.; et al. Fluoro-jade b histofluorescence staining detects dentate granule cell death after repeated five-minute transient global cerebral ischemia. Metab. Brain Dis. 2019, 34, 951–956. [Google Scholar] [CrossRef]
- Lee, C.H.; Ahn, J.H.; Lee, T.K.; Sim, H.; Lee, J.C.; Park, J.H.; Shin, M.C.; Cho, J.H.; Kim, D.W.; Won, M.H.; et al. Comparison of neuronal death, blood-brain barrier leakage and inflammatory cytokine expression in the hippocampal ca1 region following mild and severe transient forebrain ischemia in gerbils. Neurochem. Res. 2021, 46, 2852–2866. [Google Scholar] [CrossRef]
- Lee, T.K.; Kim, H.; Song, M.; Lee, J.C.; Park, J.H.; Ahn, J.H.; Yang, G.E.; Kim, H.; Ohk, T.G.; Shin, M.C.; et al. Time-course pattern of neuronal loss and gliosis in gerbil hippocampi following mild, severe, or lethal transient global cerebral ischemia. Neural Regen. Res. 2019, 14, 1394–1403. [Google Scholar]
- Chen, H.; Yoshioka, H.; Kim, G.S.; Jung, J.E.; Okami, N.; Sakata, H.; Maier, C.M.; Narasimhan, P.; Goeders, C.E.; Chan, P.H. Oxidative stress in ischemic brain damage: Mechanisms of cell death and potential molecular targets for neuroprotection. Antioxid. Redox Signal. 2011, 14, 1505–1517. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yang, S. Targeting oxidative stress for the treatment of ischemic stroke: Upstream and downstream therapeutic strategies. Brain Circ. 2016, 2, 153–163. [Google Scholar]
- Kim, B.; Lee, T.K.; Park, C.W.; Kim, D.W.; Ahn, J.H.; Sim, H.; Lee, J.C.; Yang, G.E.; Kim, J.D.; Shin, M.C.; et al. Pycnogenol((r)) supplementation attenuates memory deficits and protects hippocampal ca1 pyramidal neurons via antioxidative role in a gerbil model of transient forebrain ischemia. Nutrients 2020, 12, 2477. [Google Scholar] [CrossRef]
- Margaill, I.; Plotkine, M.; Lerouet, D. Antioxidant strategies in the treatment of stroke. Free. Radic. Biol. Med. 2005, 39, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.Y.; Shan, W.G.; Wang, S.F.; Hu, M.M.; Chen, Y. Effects of astaxanthin on blood coagulation, fibrinolysis and platelet aggregation in hyperlipidemic rats. Pharm. Biol. 2017, 55, 663–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farruggia, C.; Kim, M.B.; Bae, M.; Lee, Y.; Pham, T.X.; Yang, Y.; Han, M.J.; Park, Y.K.; Lee, J.Y. Astaxanthin exerts anti-inflammatory and antioxidant effects in macrophages in nrf2-dependent and independent manners. J. Nutr. Biochem. 2018, 62, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.S.; Lignell, A.; Pettersson, A.; Elfving, E.; Soni, M.G. Safety assessment of astaxanthin-rich microalgae biomass: Acute and subchronic toxicity studies in rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2008, 46, 3030–3036. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef]
- Manabe, Y.; Komatsu, T.; Seki, S.; Sugawara, T. Dietary astaxanthin can accumulate in the brain of rats. Biosci. Biotechnol. Biochem. 2018, 82, 1433–1436. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Niu, H.; Shao, A.; Wu, C.; Dixon, B.J.; Zhang, J.; Yang, S.; Wang, Y. Astaxanthin as a potential neuroprotective agent for neurological diseases. Mar. Drugs 2015, 13, 5750–5766. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ding, C.; Zhang, S.; Xu, Y. Neuroprotective effects of astaxanthin against oxygen and glucose deprivation damage via the pi3k/akt/gsk3beta/nrf2 signalling pathway in vitro. J. Cell. Mol. Med. 2020, 24, 8977–8985. [Google Scholar] [CrossRef]
- Pan, L.; Zhou, Y.; Li, X.F.; Wan, Q.J.; Yu, L.H. Preventive treatment of astaxanthin provides neuroprotection through suppression of reactive oxygen species and activation of antioxidant defense pathway after stroke in rats. Brain Res. Bull. 2017, 130, 211–220. [Google Scholar] [CrossRef]
- Xue, Y.; Qu, Z.; Fu, J.; Zhen, J.; Wang, W.; Cai, Y.; Wang, W. The protective effect of astaxanthin on learning and memory deficits and oxidative stress in a mouse model of repeated cerebral ischemia/reperfusion. Brain Res. Bull. 2017, 131, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Taheri, F.; Sattari, E.; Hormozi, M.; Ahmadvand, H.; Bigdeli, M.R.; Kordestani-Moghadam, P.; Anbari, K.; Milanizadeh, S.; Moghaddasi, M. Dose-dependent effects of astaxanthin on ischemia/reperfusion induced brain injury in mcao model rat. Neurochem. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Cakir, E.; Cakir, U.; Tayman, C.; Turkmenoglu, T.T.; Gonel, A.; Turan, I.O. Favorable effects of astaxanthin on brain damage due to ischemia- reperfusion injury. Comb. Chem. High Throughput Screen. 2020, 23, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Sun, H.; Wei, H.; Dong, M.; Zhang, Y.; Xu, W.; Fang, Y.; Zhao, J. Astaxanthin alleviates spinal cord ischemia-reperfusion injury via activation of pi3k/akt/gsk-3beta pathway in rats. J. Orthop. Surg. Res. 2020, 15, 275. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Shin, B.N.; Cho, J.H.; Lee, T.K.; Kim, I.H.; Noh, Y.; Kim, S.S.; Lee, H.A.; Kim, Y.M.; Kim, H.; et al. Brain ischemic preconditioning protects against moderate, not severe, transient global cerebral ischemic injury. Metab. Brain Dis. 2018, 33, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T. Delayed neuronal death in the gerbil hippocampus following ischemia. Brain Res. 1982, 239, 57–69. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, Y.J.; Kwon, K.H. Neuroprotective effects of astaxanthin in oxygen-glucose deprivation in sh-sy5y cells and global cerebral ischemia in rat. J. Clin. Biochem. Nutr. 2010, 47, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Herguido, M.J.; Carceller, F.; Roda, J.M.; Avendano, C. Hippocampal cell loss in transient global cerebral ischemia in rats: A critical assessment. Neuroscience 1999, 93, 71–80. [Google Scholar] [CrossRef]
- Atochin, D.N.; Chernysheva, G.A.; Aliev, O.I.; Smolyakova, V.I.; Osipenko, A.N.; Logvinov, S.V.; Zhdankina, A.A.; Plotnikova, T.M.; Plotnikov, M.B. An improved three-vessel occlusion model of global cerebral ischemia in rats. Brain Res. Bull. 2017, 132, 213–221. [Google Scholar] [CrossRef]
- Gyo, K. Experimental study of transient cochlear ischemia as a cause of sudden deafness. World J. Otorhinolaryngol. 2013, 3, 1–15. [Google Scholar] [CrossRef]
- Jarrott, D.M.; Domer, F.R. A gerbil model of cerebral ischemia suitable for drug evaluation. Stroke 1980, 11, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Q.; Kosten, T.R.; Zhang, X.Y. Free radicals, antioxidant defense systems, and schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 46, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Huang, D.; Tang, X.; Liu, Y.; Luo, Q.; Liu, C.; Huang, H.; Chen, W.; Qi, Z. Beclin1 exerts protective effects against cerebral ischemiareperfusion injury by promoting DNA damage repair through a nonautophagydependent regulatory mechanism. Int. J. Mol. Med. 2022, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Park, C.W.; Ahn, J.H.; Lee, T.K.; Park, Y.E.; Kim, B.; Lee, J.C.; Kim, D.W.; Shin, M.C.; Park, Y.; Cho, J.H.; et al. Post-treatment with oxcarbazepine confers potent neuroprotection against transient global cerebral ischemic injury by activating nrf2 defense pathway. Biomed. Pharmacother. 2020, 124, 109850. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, T.K.; Kim, D.W.; Sim, H.; Lee, J.C.; Kim, J.D.; Ahn, J.H.; Lee, C.H.; Kim, Y.M.; Won, M.H.; et al. Neuroprotective effects of salicin in a gerbil model of transient forebrain ischemia by attenuating oxidative stress and activating pi3k/akt/gsk3beta pathway. Antioxidants 2021, 10, 629. [Google Scholar] [CrossRef]
- Yousefi-Manesh, H.; Dehpour, A.R.; Nabavi, S.M.; Khayatkashani, M.; Asgardoon, M.H.; Derakhshan, M.H.; Moradi, S.A.; Sheibani, M.; Tavangar, S.M.; Shirooie, S.; et al. Therapeutic effects of hydroalcoholic extracts from the ancient apple mela rosa dei monti sibillini in transient global ischemia in rats. Pharmaceuticals 2021, 14, 1106. [Google Scholar] [CrossRef]
- Jia, J.Y.; Tan, Z.G.; Liu, M.; Jiang, Y.G. Apurinic/apyrimidinic endonuclease 1 (ape1) contributes to resveratrolinduced neuroprotection against oxygenglucose deprivation and reoxygenation injury in ht22 cells: Involvement in reducing oxidative DNA damage. Mol. Med. Rep. 2017, 16, 9786–9794. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Won, M.H. Neuroprotection of antioxidant enzymes against transient global cerebral ischemia in gerbils. Anat. Cell Biol. 2014, 47, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Noshita, N.; Lewen, A.; Gasche, Y.; Ferrand-Drake, M.; Fujimura, M.; Morita-Fujimura, Y.; Chan, P.H. Overexpression of copper/zinc superoxide dismutase in transgenic rats protects vulnerable neurons against ischemic damage by blocking the mitochondrial pathway of caspase activation. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.N.; Kindy, M.S.; Holtsberg, F.W.; St Clair, D.K.; Yen, H.C.; Germeyer, A.; Steiner, S.M.; Bruce-Keller, A.J.; Hutchins, J.B.; Mattson, M.P. Mitochondrial manganese superoxide dismutase prevents neural apoptosis and reduces ischemic brain injury: Suppression of peroxynitrite production, lipid peroxidation, and mitochondrial dysfunction. J. Neurosci. Off. J. Soc. Neurosci. 1998, 18, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.H.; Park, J.H.; Ahn, J.H.; Cho, J.H.; Kim, I.H.; Lee, J.C.; Won, M.H.; Lee, C.H.; Hwang, I.K.; Kim, J.D.; et al. Pretreated quercetin protects gerbil hippocampal ca1 pyramidal neurons from transient cerebral ischemic injury by increasing the expression of antioxidant enzymes. Neural Regen. Res. 2017, 12, 220–227. [Google Scholar] [PubMed]
- Lee, T.K.; Kang, I.J.; Kim, B.; Sim, H.J.; Kim, D.W.; Ahn, J.H.; Lee, J.C.; Ryoo, S.; Shin, M.C.; Cho, J.H.; et al. Experimental pretreatment with chlorogenic acid prevents transient ischemia-induced cognitive decline and neuronal damage in the hippocampus through anti-oxidative and anti-inflammatory effects. Molecules 2020, 25, 3578. [Google Scholar] [CrossRef]
- Yang, B.B.; Zou, M.; Zhao, L.; Zhang, Y.K. Astaxanthin attenuates acute cerebral infarction via nrf-2/ho-1 pathway in rats. Curr. Res. Transl. Med. 2021, 69, 103271. [Google Scholar] [CrossRef]
- Radtke-Schuller, S.; Schuller, G.; Angenstein, F.; Grosser, O.S.; Goldschmidt, J.; Budinger, E. Brain atlas of the mongolian gerbil (Meriones unguiculatus) in ct/mri-aided stereotaxic coordinates. Brain Struct. Funct. 2016, 221 (Suppl. 1), 1–272. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Ahn, J.H.; Lee, T.K.; Park, C.W.; Kim, B.; Lee, J.C.; Kim, D.W.; Shin, M.C.; Cho, J.H.; Lee, C.H.; et al. Laminarin pretreatment provides neuroprotection against forebrain ischemia/reperfusion injury by reducing oxidative stress and neuroinflammation in aged gerbils. Mar. Drugs 2020, 18, 213. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Shin, M.C.; Kim, D.W.; Kim, H.; Song, M.; Lee, T.K.; Lee, J.C.; Kim, H.; Cho, J.H.; Kim, Y.M.; et al. Antioxidant properties of fucoidan alleviate acceleration and exacerbation of hippocampal neuronal death following transient global cerebral ischemia in high-fat diet-induced obese gerbils. Int. J. Mol. Sci. 2019, 20, 554. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Dilutions | Suppliers |
|---|---|---|
| Mouse anti-neuronal nuclei (NeuN) | 1:1000 | Chemicon, Temecula, CA, USA |
| Goat anti-8-hydroxydeoxyguanosine (8OHdG) | 1:500 | Millipore, Billerica, MA, USA |
| Mouse 4-hydroxy-2-nonenal (4HNE) | 1:800 | Alexis Biochemicals, San Diego, CA, USA |
| Sheep anti-Cu, Zn-superoxide dismutase (SOD1) | 1:800 | Calbiochem, La Jolla, CA, USA |
| Sheep anti-Mn-superoxide dismutase (SOD2) | 1:800 | Calbiochem, La Jolla, CA, USA |
| Secondary Antibodies | Dilutions | Suppliers |
| Biotinylated horse anti-mouse IgG | 1:250 | Vector Laboratories Inc., Burlingame, CA, USA |
| Biotinylated horse anti-goat IgG | 1:250 | Vector Laboratories Inc., Burlingame, CA, USA |
| Biotinylated rabbit anti-sheep IgG | 1:250 | Vector Laboratories Inc., Burlingame, CA, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.H.; Lee, T.-K.; Kim, D.W.; Ahn, J.H.; Lee, C.-H.; Kim, J.-D.; Shin, M.C.; Cho, J.H.; Lee, J.-C.; Won, M.-H.; et al. Astaxanthin Confers a Significant Attenuation of Hippocampal Neuronal Loss Induced by Severe Ischemia-Reperfusion Injury in Gerbils by Reducing Oxidative Stress. Mar. Drugs 2022, 20, 267. https://doi.org/10.3390/md20040267
Park JH, Lee T-K, Kim DW, Ahn JH, Lee C-H, Kim J-D, Shin MC, Cho JH, Lee J-C, Won M-H, et al. Astaxanthin Confers a Significant Attenuation of Hippocampal Neuronal Loss Induced by Severe Ischemia-Reperfusion Injury in Gerbils by Reducing Oxidative Stress. Marine Drugs. 2022; 20(4):267. https://doi.org/10.3390/md20040267
Chicago/Turabian StylePark, Joon Ha, Tae-Kyeong Lee, Dae Won Kim, Ji Hyeon Ahn, Choong-Hyun Lee, Jong-Dai Kim, Myoung Cheol Shin, Jun Hwi Cho, Jae-Chul Lee, Moo-Ho Won, and et al. 2022. "Astaxanthin Confers a Significant Attenuation of Hippocampal Neuronal Loss Induced by Severe Ischemia-Reperfusion Injury in Gerbils by Reducing Oxidative Stress" Marine Drugs 20, no. 4: 267. https://doi.org/10.3390/md20040267