
Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction


 ,
,  ,
,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
1.1. Neuro-Immune Interactions in Host Defense
1.2. Interactions between the Immune Cells and the Nervous System in Different Tissues
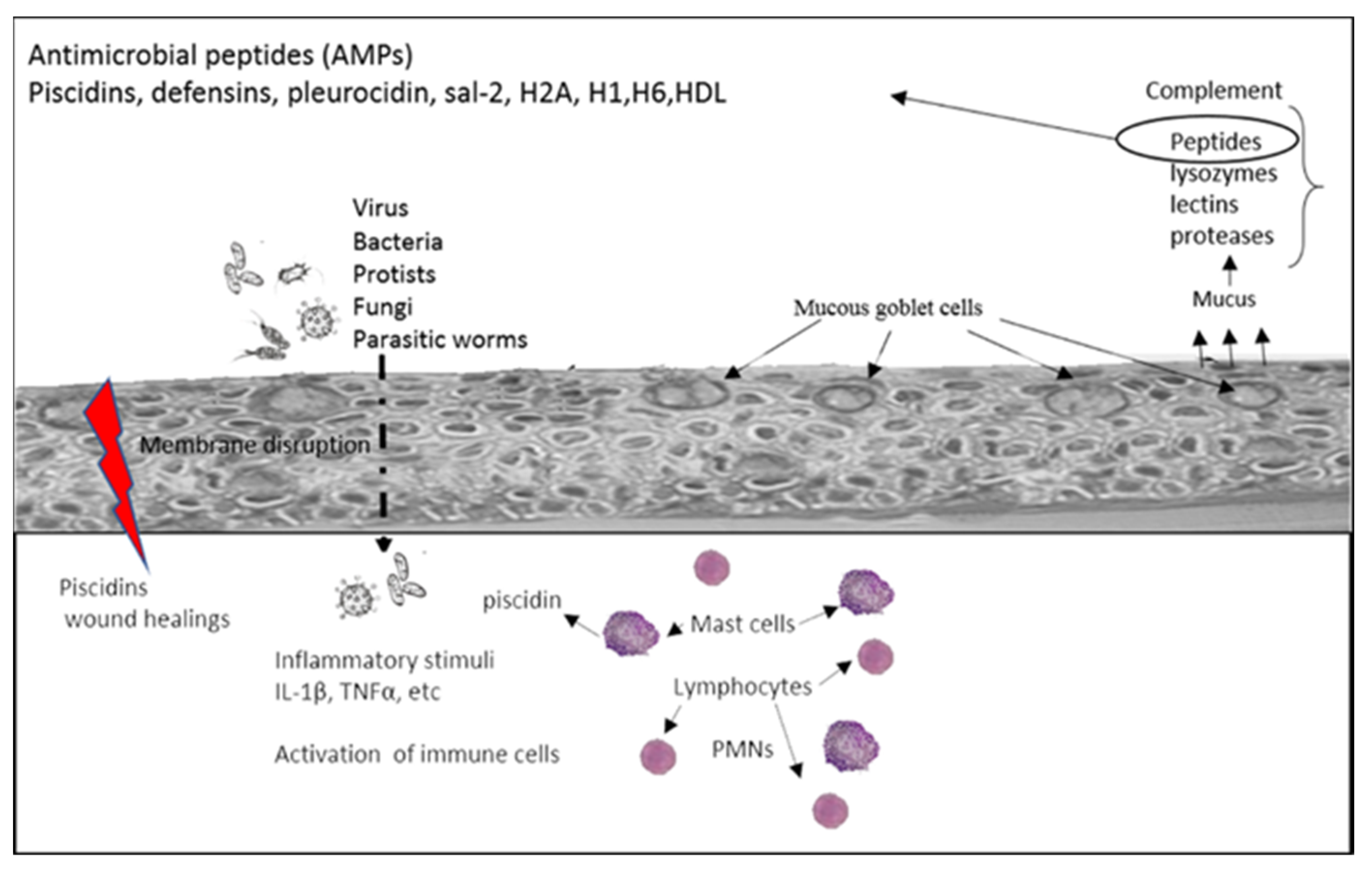
2. The Neuro-Immune Interaction at Barrier Surfaces
2.1. Neuro-Immune Circuits in Peripheral Tissues
2.2. Fish Mucosal Surfaces
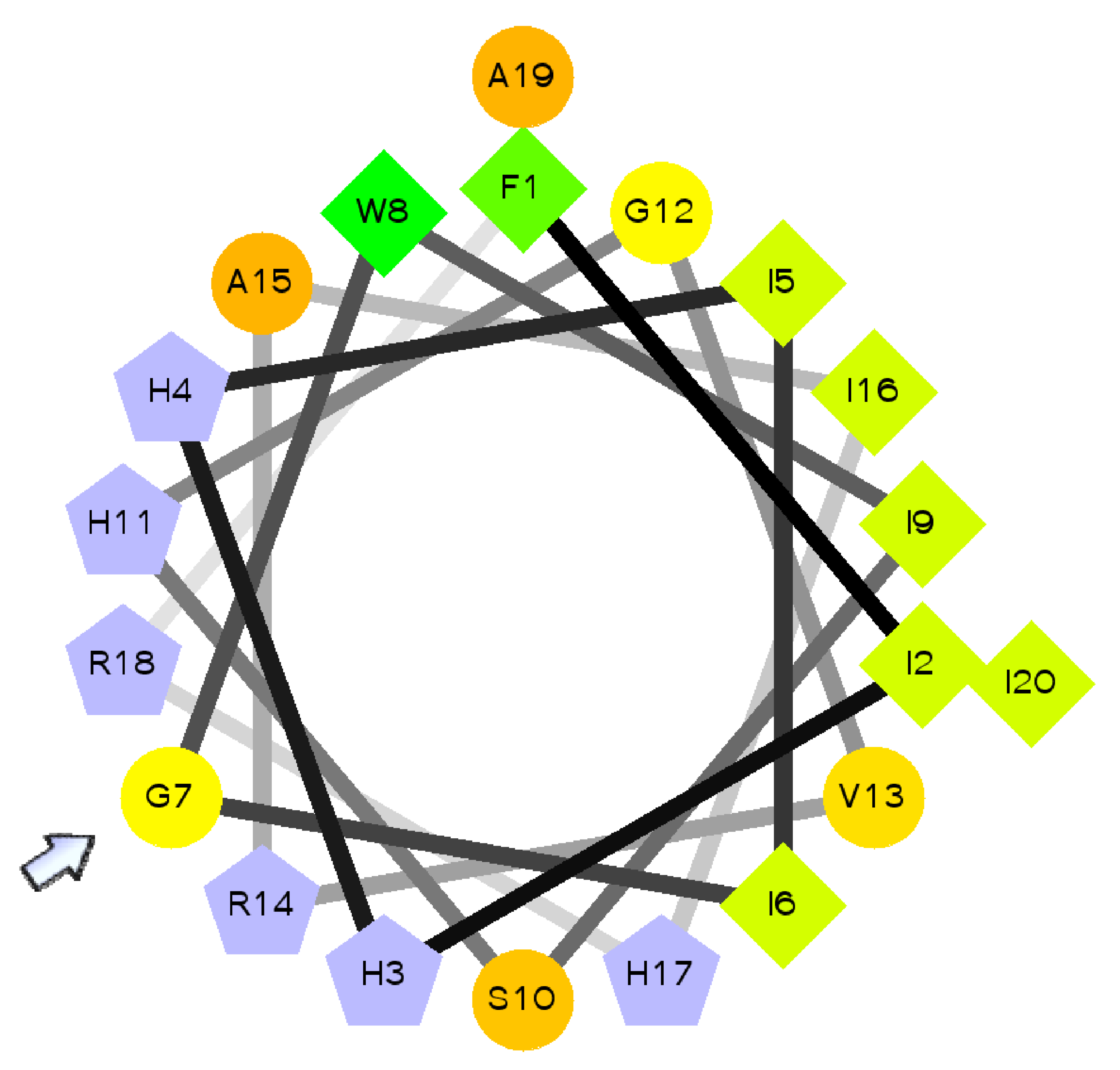
3. Piscidins and Neuropeptides as a Source of AMPs: Phylogeny, Cell Distribution, Molecular Structure and Immunomodulatory Function
4. Neuropeptides and AMPs and Immune-Associated Function
5. Neuroimmune Communication in the Gill-CNS Axis
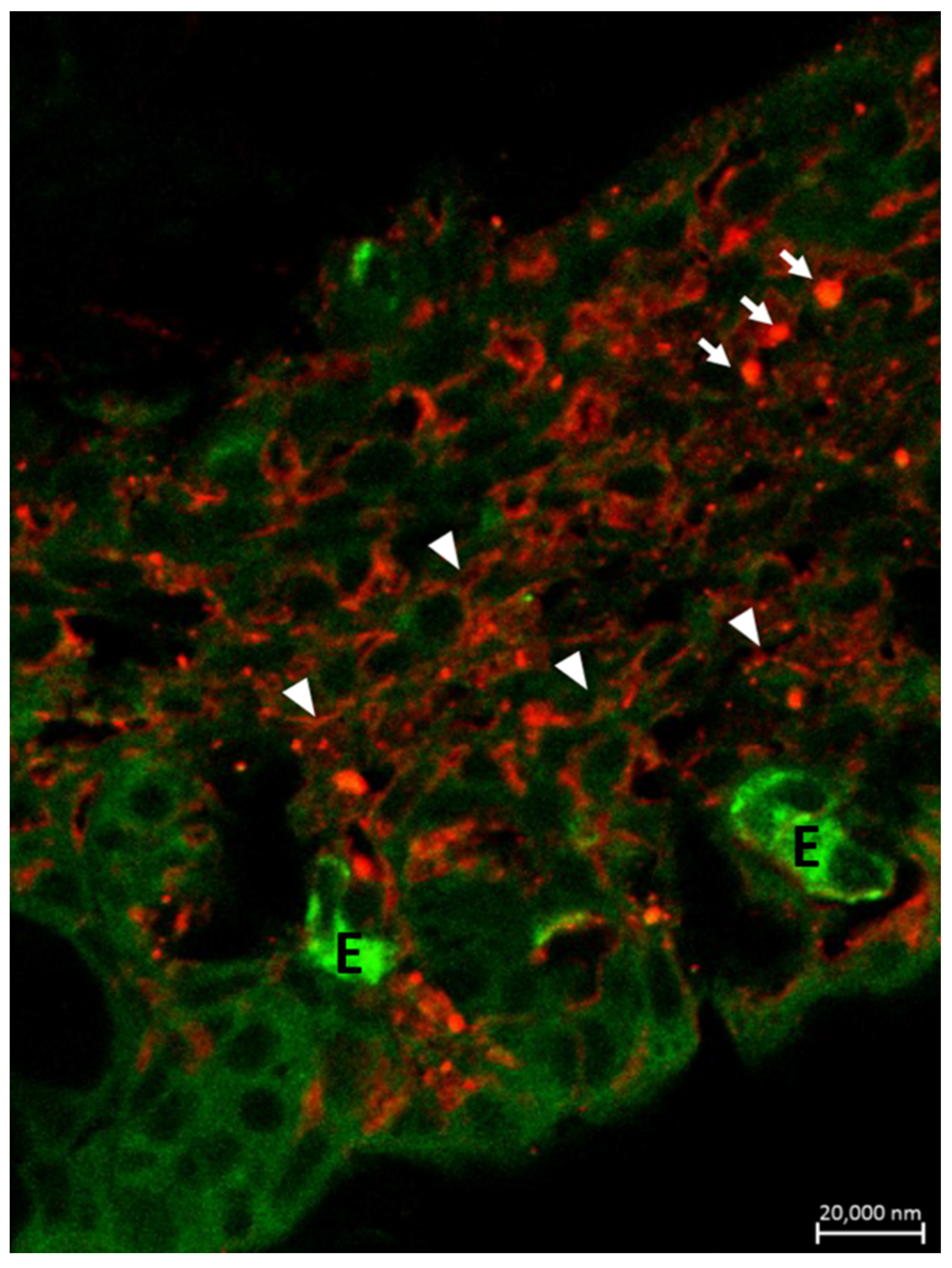
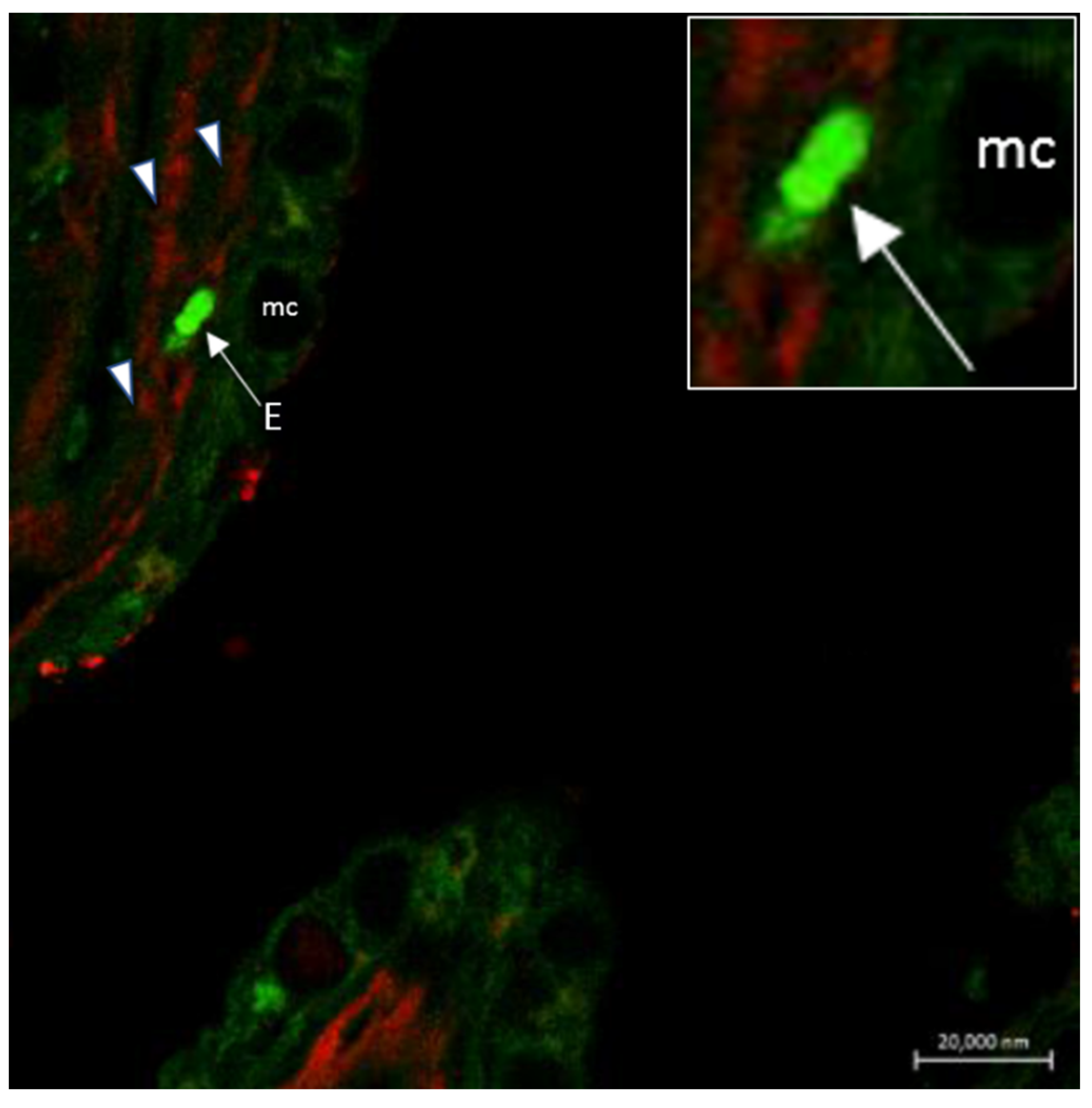
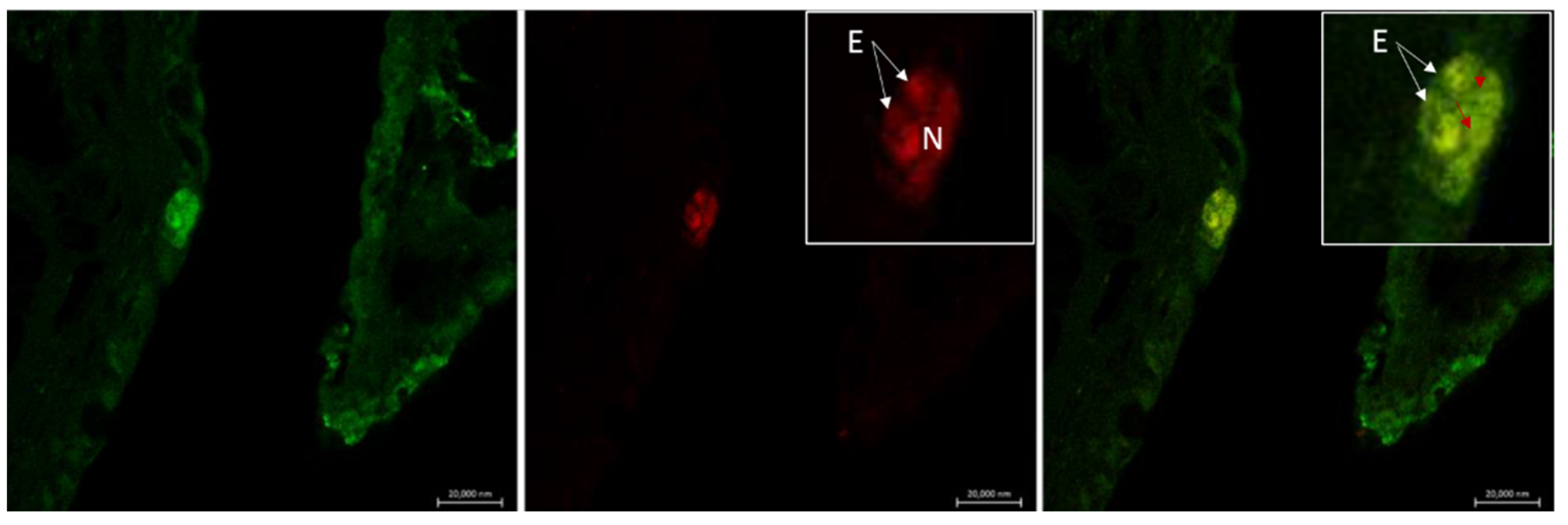
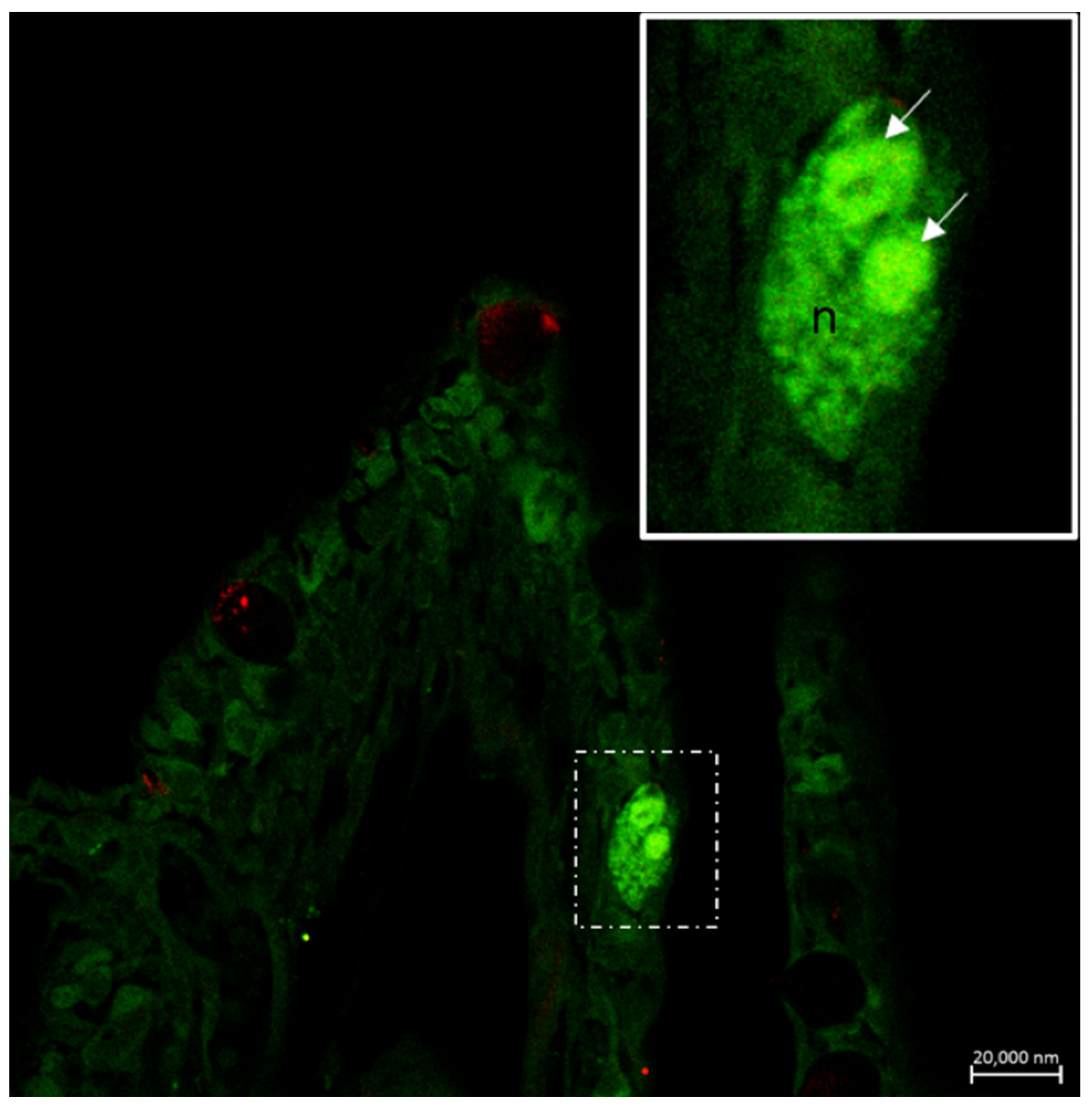
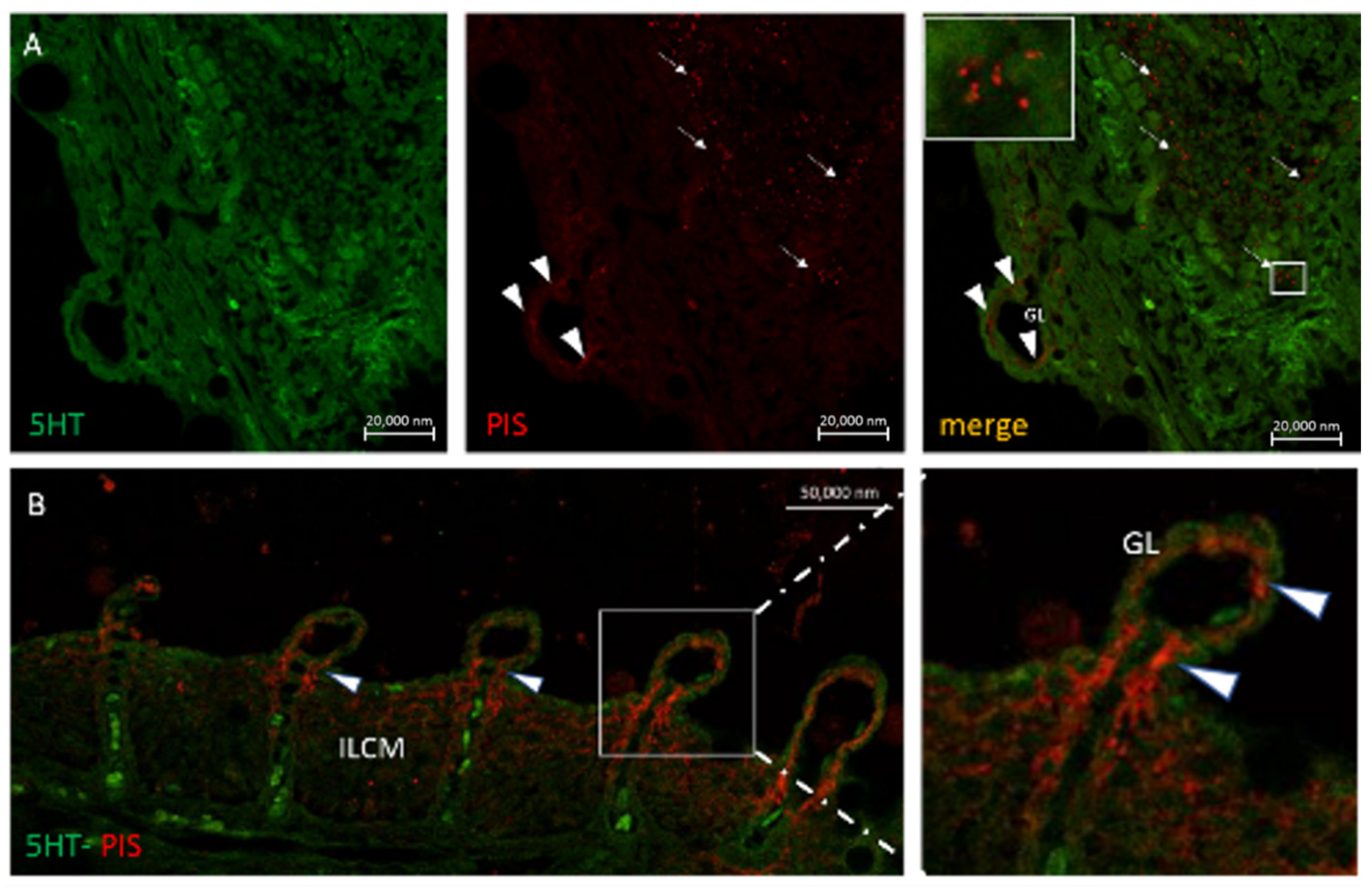
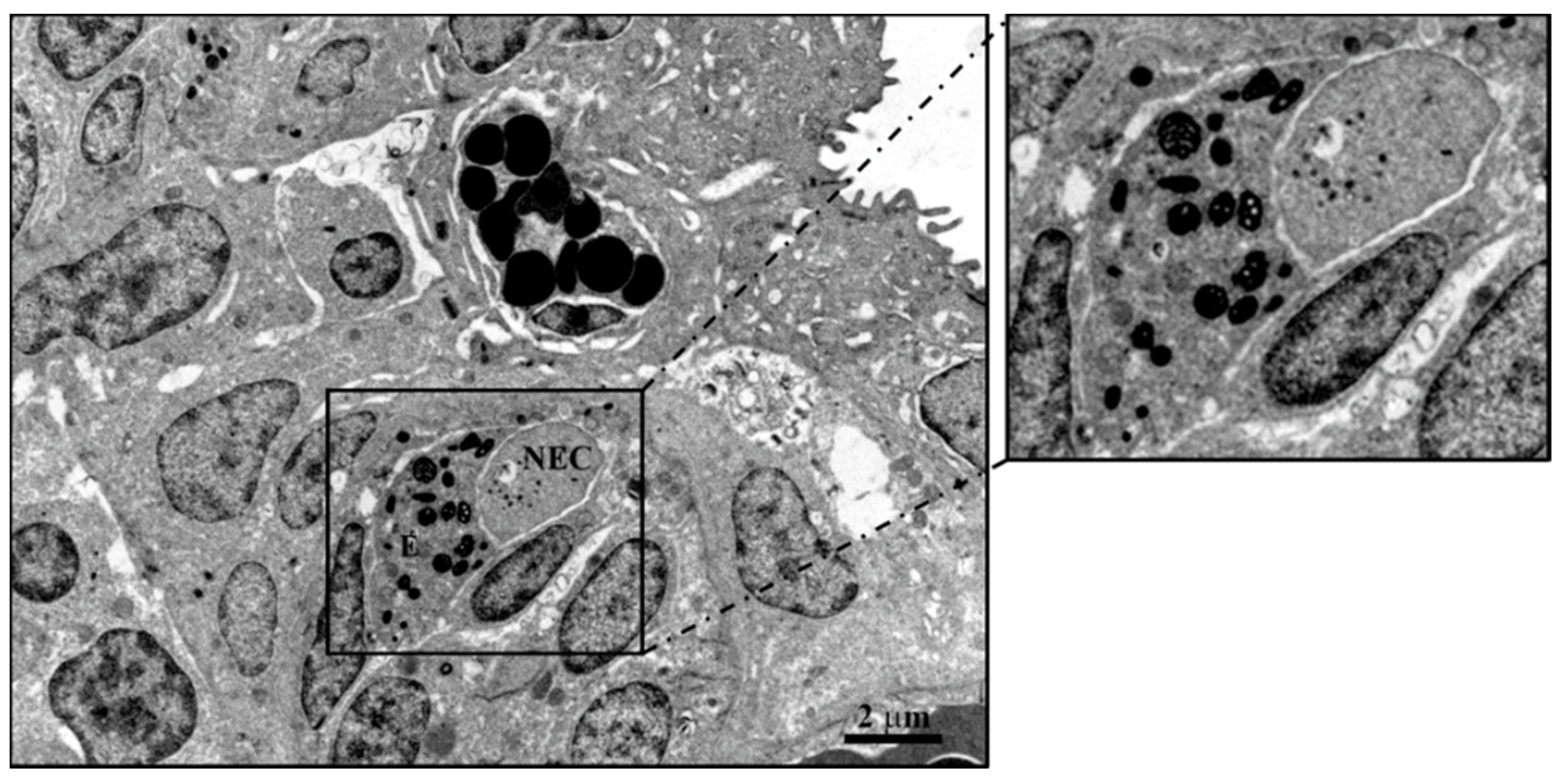
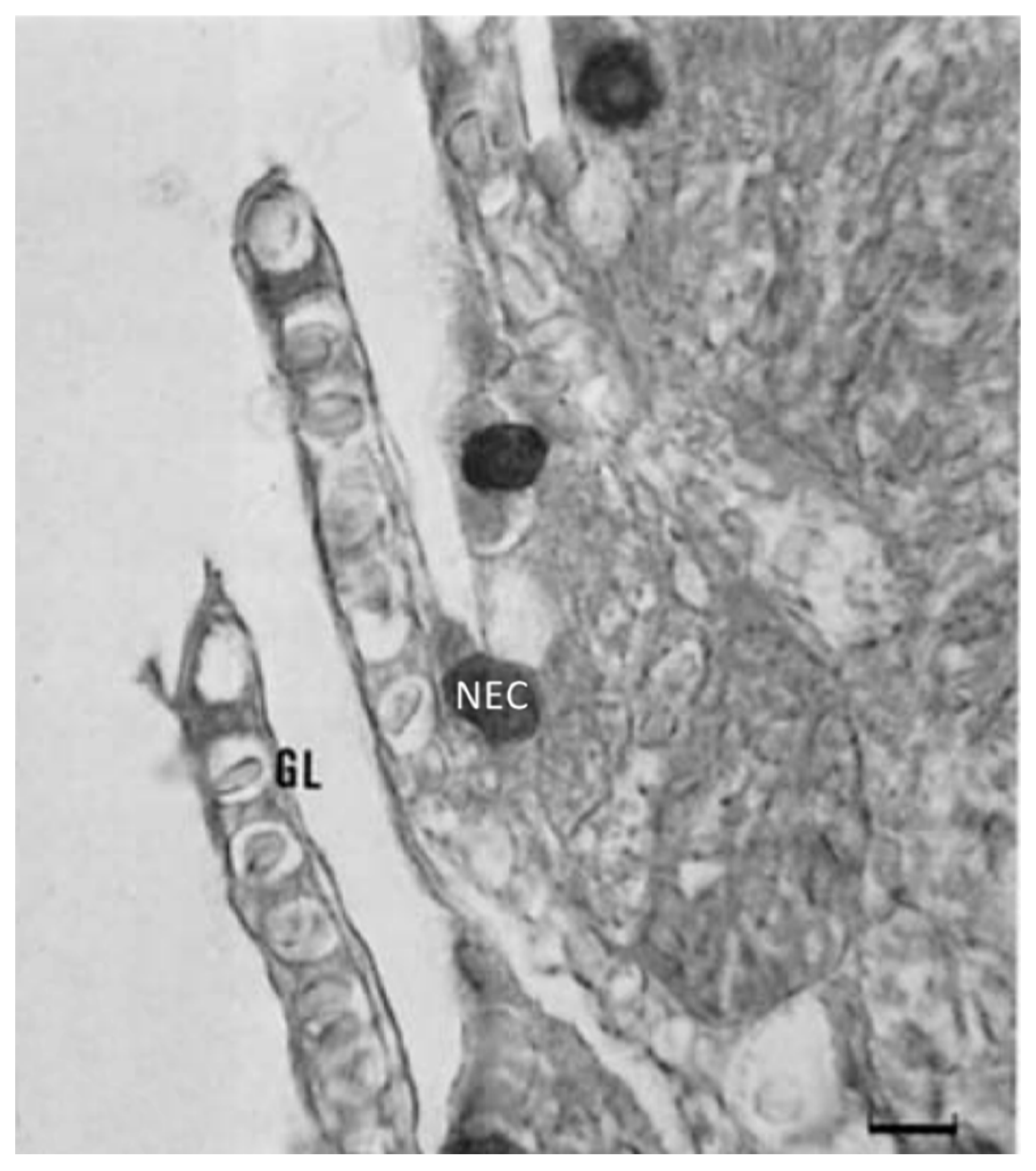
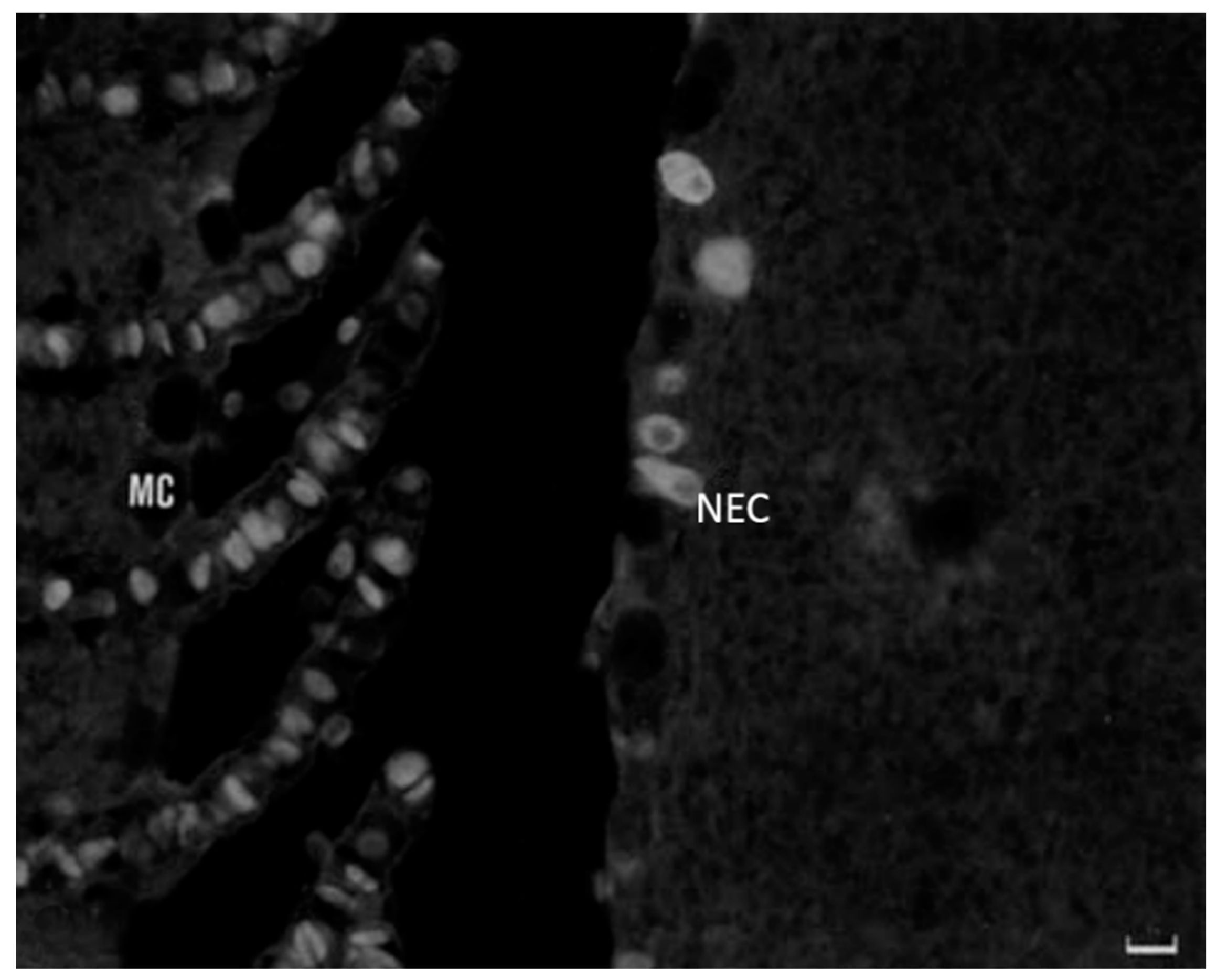
6. Neuroimmune Cell Crosstalk in the Gill
6.1. Neuropeptides
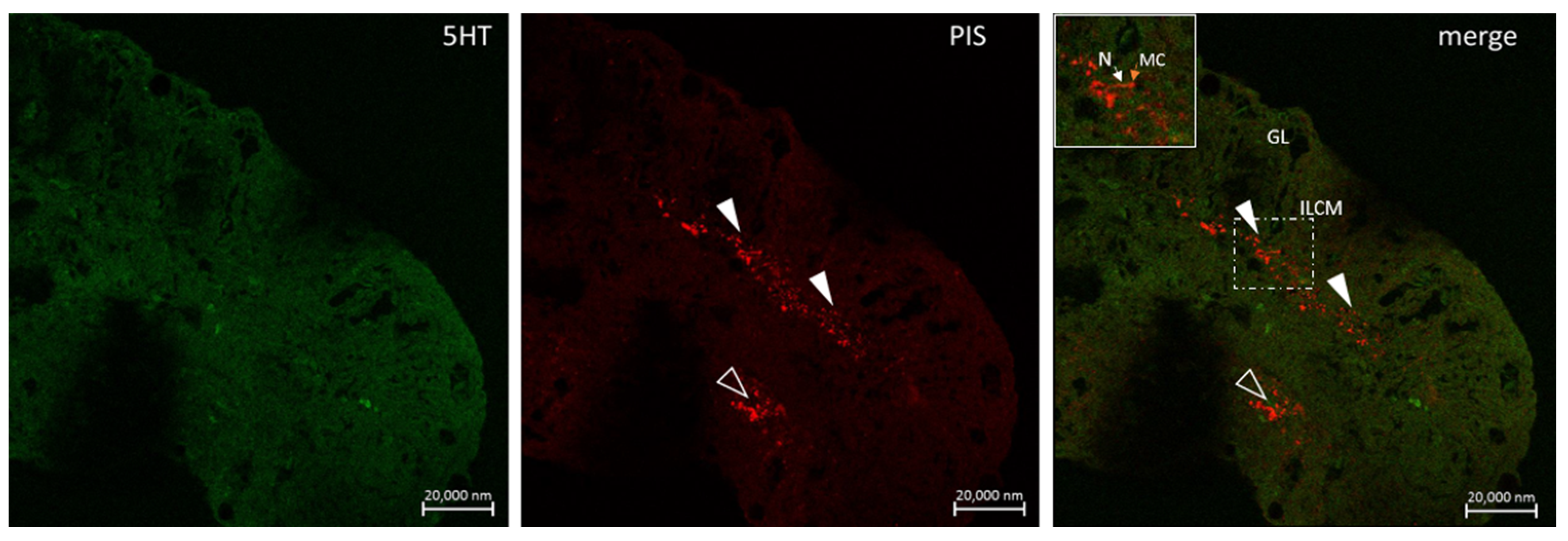
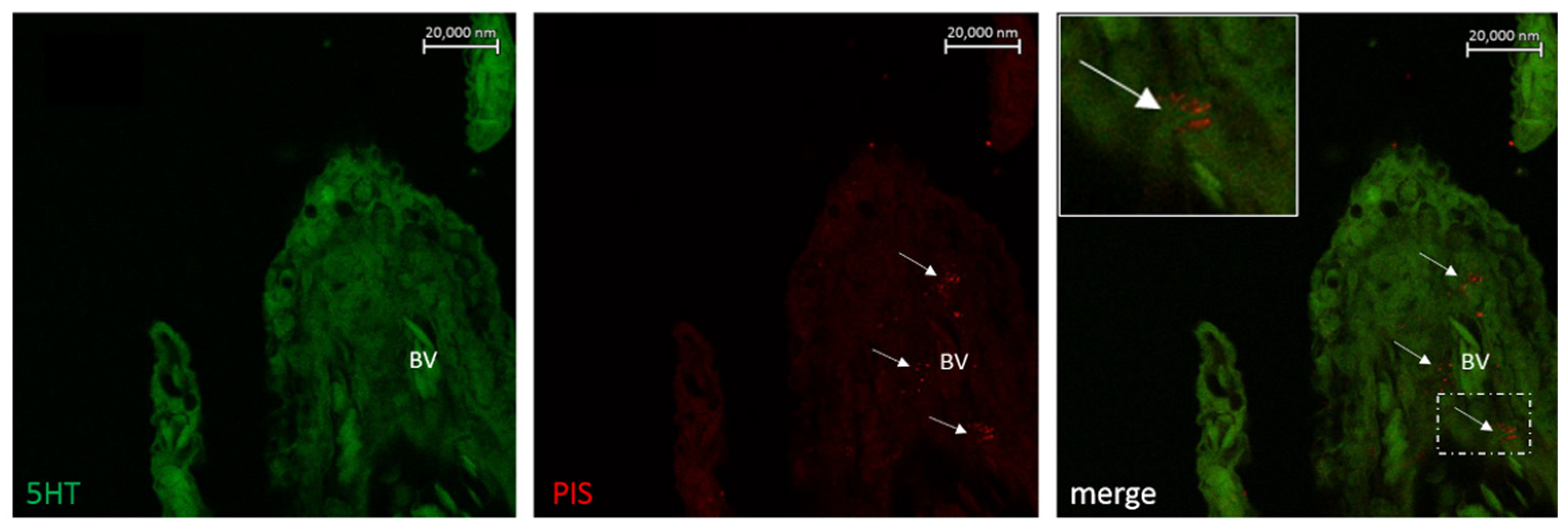
6.2. Neuro Mast Cell Crosstalk in the Skin
7. Conclusions
Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schluesener, H.J.; Su, Y.; Ebrahimi, A.; Pouladsaz, D. Antimicrobial peptides in the brain: Neuropeptides and amyloid. Front. Biosci. 2012, 4, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Kushwaha, S.; Rana, A.K.; Misra-Bhattacharya, S. Cofactor Independent Phosphoglycerate Mutase of Brugia malayi Induces a Mixed Th1/Th2 Type Immune Response and Inhibits Larval Development in the Host. BioMed Res. Int. 2014, 2014, 590281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Aidy, S.; Stilling, R.; Dinan, T.G.; Cryan, J.F. Microbiome to brain: Unravelling the multidirectional axes of communication. Microb. Endocrinol. Interkingdom Signal. Infect. Dis. 2016, 301–336. [Google Scholar] [CrossRef]
- Aresti Sanz, J.; El Aidy, S. Microbiota and gut neuropeptides: A dual action of antimicrobial activity and neuroimmune response. Psychopharmacology 2019, 236, 1597–1609. [Google Scholar] [CrossRef] [Green Version]
- Yoo, B.B.; Mazmanian, S.K. The Enteric Network: Interactions between the Immune and Nervous Systems of the Gut. Immunity 2017, 46, 910–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, T.; Goto, R.; Fujimoto, M.; Okada, N.; Hardt, W.-D. The Bactericidal Lectin RegIIIβ Prolongs Gut Colonization and Enteropathy in the Streptomycin Mouse Model for Salmonella Diarrhea. Cell Host Microbe 2017, 21, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Zaccone, G.; Lauweryns, J.M.; Fasulo, S.; Tagliafierro, G.; Ainis, L.; Licata, A. Immunocytochemical Localization of Serotonin and Neuropeptides in the Neuroendocrine Paraneurons of Teleost and Lungfish Gills. Acta Zool. 1992, 73, 177–183. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Capillo, G.; Icardo, J.M.; Fernandes, J.M.O.; Kiron, V.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Germana’, A.; et al. Neuroepithelial cells (NECs) and mucous cells express a variety of neurotransmitters and neurotransmitter receptors in the gill and respiratory air-sac of the catfish Heteropneustes fossilis (Siluriformes, Heteropneustidae): A possible role in local immune defence. Zoology 2021, 148, 125958. [Google Scholar] [CrossRef]
- Wei, Y.; Li, Y.; Yan, L.; Sun, C.; Miao, Q.; Wang, Q.; Xiao, X.; Lian, M.; Li, B.; Chen, Y.; et al. Alterations of gut microbiome in autoimmune hepatitis. Gut 2020, 69, 569. [Google Scholar] [CrossRef]
- Rességuier, J.; Dalum, A.S.; Du Pasquier, L.; Zhang, Y.; Koppang, E.O.; Boudinot, P.; Wiegertjes, G.F. Lymphoid Tissue in Teleost Gills: Variations on a Theme. Biology 2020, 9, 127. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, Q.; Huang, Z.; Ding, L.; Xu, Z. Immunoglobulins, Mucosal Immunity and Vaccination in Teleost Fish. Front. Immunol. 2020, 11, 2597. [Google Scholar] [CrossRef]
- Maki Joanne, L.; Dickerson Harry, W. Systemic and Cutaneous Mucus Antibody Responses of Channel Catfish Immunized against the Protozoan Parasite Ichthyophthirius multifiliis. Clin. Vaccine Immunol. 2003, 10, 876–881. [Google Scholar] [CrossRef] [Green Version]
- Ruangsri, J.; Fernandes, J.M.O.; Rombout, J.H.W.M.; Brinchmann, M.F.; Kiron, V. Ubiquitous presence of piscidin-1 in Atlantic cod as evidenced by immunolocalisation. BMC Vet. Res. 2012, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Ruangsri, J.; Lokesh, J.; Fernandes, J.M.O.; Kiron, V. Transcriptional regulation of antimicrobial peptides in mucosal tissues of Atlantic cod Gadus morhua L. in response to different stimuli. Aquac. Res. 2014, 45, 1893–1905. [Google Scholar] [CrossRef]
- Capillo, G.; Zaccone, G.; Cupello, C.; Fernandes, J.M.O.; Viswanath, K.; Kuciel, M.; Zuwala, K.; Guerrera, M.C.; Aragona, M.; Icardo, J.M.; et al. Expression of acetylcholine, its contribution to regulation of immune function and O2 sensing and phylogenetic interpretations of the African butterfly fish Pantodon buchholzi (Osteoglossiformes, Pantodontidae). Fish Shellfish. Immunol. 2021, 111, 189–200. [Google Scholar] [CrossRef]
- Katzenback, B.A. Antimicrobial Peptides as Mediators of Innate Immunity in Teleosts. Biology 2015, 4, 607. [Google Scholar] [CrossRef] [Green Version]
- Burleson, M.L. Sensory innervation of the Gills: O2-sensitive chemoreceptors and mechanoreceptors. Acta Histochem. 2009, 111, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Bailly, Y. Serotonergic neuroepithelial cells in fish gills: Cytology and innervation. In Airway Chemoreceptors in the Vertebrates; CRC Press: Boca Raton, FL, USA, 2019; pp. 61–97. ISBN 0429526709/9780429526701. [Google Scholar]
- Blake, K.J.; Jiang, X.R.; Chiu, I.M. Neuronal Regulation of Immunity in the Skin and Lungs. Trends Neurosci. 2019, 42, 537–551. [Google Scholar] [CrossRef]
- Branchfield, K.; Nantie, L.; Verheyden Jamie, M.; Sui, P.; Wienhold Mark, D.; Sun, X. Pulmonary neuroendocrine cells function as airway sensors to control lung immune response. Science 2016, 351, 707–710. [Google Scholar] [CrossRef] [Green Version]
- Sui, P.; Wiesner Darin, L.; Xu, J.; Zhang, Y.; Lee, J.; Van Dyken, S.; Lashua, A.; Yu, C.; Klein Bruce, S.; Locksley Richard, M.; et al. Pulmonary neuroendocrine cells amplify allergic asthma responses. Science 2018, 360, eaan8546. [Google Scholar] [CrossRef] [Green Version]
- Veiga-Fernandes, H.; Artis, D. Neuronal–immune system cross-talk in homeostasis. Science 2018, 359, 1465–1466. [Google Scholar] [CrossRef] [Green Version]
- Day, R.; Salzet, M. The neuroendocrine phenotype, cellular plasticity, and the search for genetic switches: Redefining the diffuse neuroendocrine system. Neuroendocrinol. Lett. 2002, 23, 447–451. [Google Scholar]
- Stefano, G.B.; Salzet, B.; Fricchione, G.L. Enkelytin and opioid peptide association in invertebrates and vertebrates: Immune activation and pain. Immunol. Today 1998, 19, 265–268. [Google Scholar] [CrossRef]
- Zaccone, G.; Mauceri, A.; Fasulo, S. Neuropeptides and nitric oxide synthase in the gill and the air-breathing organs of fishes. J. Exp. Zool. Part A Comp. Exp. Biol. 2006, 305, 428–439. [Google Scholar] [CrossRef]
- Maina, J.W.; Pringle, J.M.; Razal, J.M.; Nunes, S.; Vega, L.; Gallucci, F.; Dumée, L.F. Strategies for Integrated Capture and Conversion of CO2 from Dilute Flue Gases and the Atmosphere. ChemSusChem 2021, 14, 1805–1820. [Google Scholar] [CrossRef]
- Luger, T.A.; Lotti, T. Neuropeptides: Role in inflammatory skin diseases. J. Eur. Acad. Dermatol. Venereol. 1998, 10, 207–211. [Google Scholar] [CrossRef]
- Lee, E.Y.; Chan, L.C.; Wang, H.; Lieng, J.; Hung, M.; Srinivasan, Y.; Wang, J.; Waschek, J.A.; Ferguson, A.L.; Lee, K.-F.; et al. PACAP is a pathogen-inducible resident antimicrobial neuropeptide affording rapid and contextual molecular host defense of the brain. Proc. Natl. Acad. Sci. USA 2021, 118, e1917623117. [Google Scholar] [CrossRef]
- Gera, S.; Kankuri, E.; Kogermann, K. Antimicrobial peptides—Unleashing their therapeutic potential using nanotechnology. Pharmacol. Ther. 2021, 107990. [Google Scholar] [CrossRef]
- De Becker, G.; Moulin, V.; Pajak, B.; Bruck, C.; Francotte, M.; Thiriart, C.; Urbain, J.; Moser, M. The adjuvant monophosphoryl lipid A increases the function of antigen-presenting cells. Int. Immunol. 2000, 12, 807–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasiemski, A.; Salzet, M. Neuro-immune lessons from an annelid: The medicinal leech. Dev. Comp. Immunol. 2017, 66, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Veiga-Fernandes, H.; Mucida, D. Neuro-Immune Interactions at Barrier Surfaces. Cell 2016, 165, 801–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shouman, K.; Benarroch, E.E. Peripheral neuroimmune interactions: Selected review and some clinical implications. Clin. Auton. Res. 2021, 31, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Siriyappagouder, P.; Kiron, V.; Lokesh, J.; Rajeish, M.; Kopp, M.; Fernandes, J. The Intestinal Mycobiota in Wild Zebrafish Comprises Mainly Dothideomycetes While Saccharomycetes Predominate in Their Laboratory-Reared Counterparts. Front. Microbiol. 2018, 9, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, S.; Ozório, R.O.A.; Kollias, S.; Dhanasiri, A.K.; Lokesh, J.; Kiron, V.; Valente, L.M.P.; Fernandes, J.M.O. Changes in intestinal microbiota, immune- and stress-related transcript levels in Senegalese sole (Solea senegalensis) fed plant ingredient diets intercropped with probiotics or immunostimulants. Aquaculture 2016, 458, 149–157. [Google Scholar] [CrossRef]
- Lokesh, J.; Kiron, V.; Sipkema, D.; Fernandes, J.M.O.; Moum, T. Succession of embryonic and intestinal bacterial communities of Atlantic salmon. bioRxiv 2017. [Google Scholar] [CrossRef]
- Holzer, P.; Farzi, A. Neuropeptides and the microbiota-gut-brain axis. Microb. Endocrinol. Microbiota-Gut-Brain Axis Health 2014, 195–219. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, T.; Jacobsen, F.; Steinau, H.-U.; Steinstraesser, L. Host defense peptides and the new line of defence against multiresistant infections. Protein Pept. Lett. 2008, 15, 238–243. [Google Scholar] [CrossRef]
- Augustyniak, D.; Nowak, J.T.; Lundy, F. Direct and indirect antimicrobial activities of neuropeptides and their therapeutic potential. Curr. Protein Pept. Sci. 2012, 13, 723–738. [Google Scholar] [CrossRef] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Corrales, L.; Matson, V.; Flood, B.; Spranger, S.; Gajewski, T.F. Innate immune signaling and regulation in cancer immunotherapy. Cell Res. 2017, 27, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Current Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [Green Version]
- Salger, S.A.; Cassady, K.R.; Reading, B.J.; Noga, E.J. A diverse family of host-defense peptides (piscidins) exhibit specialized anti-bacterial and anti-protozoal activities in fishes. PLoS ONE 2016, 11, e0159423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauth, X.; Shike, H.; Burns, J.C.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; Van Olst, J.C.; Nizet, V.; Taylor, S.W.; Shimizu, C. Discovery and characterization of two isoforms of moronecidin, a novel antimicrobial peptide from hybrid striped bass. J. Biol. Chem. 2002, 277, 5030–5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salerno, G.; Parrinello, N.; Roch, P.; Cammarata, M. cDNA sequence and tissue expression of an antimicrobial peptide, dicentracin; a new component of the moronecidin family isolated from head kidney leukocytes of sea bass, Dicentrarchus labrax. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 146, 521–529. [Google Scholar] [CrossRef]
- Acosta, J.; Montero, V.; Carpio, Y.; Velázquez, J.; Garay, H.E.; Reyes, O.; Cabrales, A.; Masforrol, Y.; Morales, A.; Estrada, M.P. Cloning and functional characterization of three novel antimicrobial peptides from tilapia (Oreochromis niloticus). Aquaculture 2013, 372, 9–18. [Google Scholar] [CrossRef]
- Ruangsri, J.; Salger, S.A.; Caipang, C.M.A.; Kiron, V.; Fernandes, J.M.O. Differential expression and biological activity of two piscidin paralogues and a novel splice variant in Atlantic cod (Gadus morhua L.). Fish Shellfish. Immunol. 2012, 32, 396–406. [Google Scholar] [CrossRef]
- Noga, E.J.; Silphaduang, U.; Park, N.G.; Seo, J.K.; Stephenson, J.; Kozlowicz, S. Piscidin 4, a novel member of the piscidin family of antimicrobial peptides. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 152, 299–305. [Google Scholar] [CrossRef]
- Silphaduang, U.; Noga, E.J. Peptide antibiotics in mast cells of fish. Nature 2001, 414, 268–269. [Google Scholar] [CrossRef]
- Sung, W.S.; Lee, D.G. Mechanism of decreased susceptibility for Gram-negative bacteria and synergistic effect with ampicillin of indole-3-carbinol. Biol. Pharm. Bull. 2008, 31, 1798–1801. [Google Scholar] [CrossRef] [Green Version]
- Colorni, A.; Ullal, A.; Heinisch, G.; Noga, E.J. Activity of the antimicrobial polypeptide piscidin 2 against fish ectoparasites. J. Fish Dis. 2008, 31, 423–432. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Koti, P.N.; Sugam, R.; Neog, K.; Hejazi, M. Cooperation or rivalry? Impact of alternative development pathways on India’s long-term electricity generation and associated water demands. Energy 2020, 192, 116708. [Google Scholar] [CrossRef]
- Fernandes, D.; van Heerden, W.; Ribeiro, A.; Brandão, T.; Mello, E.; Rivera, C.; van Heerden, M.; Gondak, R.; Santos-Silva, A.; Vargas, P. Different methods of cell quantification can lead to different results: A comparison of digital methods using a pilot study of dendritic cells in HIV-positive patients. Med. Oral Patol. Oral Y Cir. Bucal 2020, 23, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Sousa, R.; Jain, E.; Cohen, O.; Kim, D.; Buendia-Buendia, J.; Winer, E.; Lin, N.; Tolaney, S.M.; Wagle, N. Prevalence and mutational determinants of high tumor mutation burden in breast cancer. Ann. Oncol. 2020, 31, 387–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raju, S.V.; Sarkar, P.; Kumar, P.; Arockiaraj, J. Piscidin, fish antimicrobial peptide: Structure, classification, properties, mechanism, gene regulation and therapeutical importance. Int. J. Pept. Res. Ther. 2021, 27, 91–107. [Google Scholar] [CrossRef]
- Avci, F.G.; Sariyar Akbulut, B.; Ozkirimli, E. Membrane Active Peptides and Their Biophysical Characterization. Biomolecules 2018, 8, 77. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Mao, Y.; Niu, S.-F.; Pan, Y.; Zheng, W.-H.; Wang, J. Molecular characterisation and biological activity of an antiparasitic peptide from Sciaenops ocellatus and its immune response to Cryptocaryon irritans. Mol. Immunol. 2022, 141, 1–12. [Google Scholar] [CrossRef]
- Noga, E.J.; Ullal, A.J.; Corrales, J.; Fernandes, J.M.O. Application of antimicrobial polypeptide host defenses to aquaculture: Exploitation of downregulation and upregulation responses. Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 44–54. [Google Scholar] [CrossRef]
- Chen, W.; Cotten, M.L. Expression, purification, and micelle reconstitution of antimicrobial piscidin 1 and piscidin 3 for NMR studies. Protein Expr. Purif. 2014, 102, 63–68. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Lin, W.-J.; Lin, T.-L. A fish antimicrobial peptide, tilapia hepcidin TH2-3, shows potent antitumor activity against human fibrosarcoma cells. Peptides 2009, 30, 1636–1642. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Lin, W.-J.; Wu, J.-L.; Her, G.M.; Hui, C.-F. Epinecidin-1 peptide induces apoptosis which enhances antitumor effects in human leukemia U937 cells. Peptides 2009, 30, 2365–2373. [Google Scholar] [CrossRef]
- Salger, S.A.; Reading, B.J.; Baltzegar, D.A.; Sullivan, C.V.; Noga, E.J. Molecular characterization of two isoforms of piscidin 4 from the hybrid striped bass (Morone chrysops × Morone saxatilis). Fish Shellfish. Immunol. 2011, 30, 420–424. [Google Scholar] [CrossRef]
- Peng, K.C.; Lee, S.H.; Hour, A.L.; Pan, C.Y.; Lee, L.H.; Chen, J.Y. Five different piscidins from Nile tilapia, Oreochromis niloticus: Analysis of their expressions and biological functions. PLoS ONE 2012, 7, e50263. [Google Scholar] [CrossRef]
- Yoshida, K.; Mukai, Y.; Niidome, T.; Takashi, C.; Tokunaga, Y.; Hatakeyama, T.; Aoyagi, H. Interaction of pleurocidin and its analogs with phospholipid membrane and their antibacterial activity. J. Pept. Res. 2001, 57, 119–126. [Google Scholar] [CrossRef]
- Sun, Y.; Forrest, S.R. High-efficiency white organic light emitting devices with three separate phosphorescent emission layers. Appl. Phys. Lett. 2007, 91, 263503. [Google Scholar] [CrossRef]
- Rathinakumar, R.; Walkenhorst, W.F.; Wimley, W.C. Broad-Spectrum Antimicrobial Peptides by Rational Combinatorial Design and High-Throughput Screening: The Importance of Interfacial Activity. J. Am. Chem. Soc. 2009, 131, 7609–7617. [Google Scholar] [CrossRef] [Green Version]
- Browne, M.A.; Dissanayake, A.; Galloway, T.S.; Lowe, D.M.; Thompson, R.C. Ingested microscopic plastic translocates to the circulatory system of the mussel, Mytilus edulis (L.). Environ. Sci. Technol. 2008, 42, 5026–5031. [Google Scholar] [CrossRef]
- Narayana, J.L.; Huang, H.-N.; Wu, C.-J.; Chen, J.-Y. Epinecidin-1 antimicrobial activity: In vitro membrane lysis and In vivo efficacy against Helicobacter pylori infection in a mouse model. Biomaterials 2015, 61, 41–51. [Google Scholar] [CrossRef]
- Li, M.V.; Chen, W.; Harmancey, R.N.; Nuotio-Antar, A.M.; Imamura, M.; Saha, P.; Taegtmeyer, H.; Chan, L. Glucose-6-phosphate mediates activation of the carbohydrate responsive binding protein (ChREBP). Biochem. Biophys. Res. Commun. 2010, 395, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.-Y.; Tsai, T.-Y.; Su, B.-C.; Hui, C.-F.; Chen, J.-Y. Study of the antimicrobial activity of tilapia piscidin 3 (TP3) and TP4 and their effects on immune functions in hybrid tilapia (Oreochromis spp.). PLoS ONE 2017, 12, e0169678. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lu, X.-J.; Chai, F.-C.; Chen, J. Molecular characterization and functional analysis of a piscidin gene in large yellow croaker (Larimichthys crocea). Zool Res. 2016, 37, 347–355. [Google Scholar] [CrossRef]
- Iijima, N.; Tanimoto, N.; Emoto, Y.; Morita, Y.; Uematsu, K.; Murakami, T.; Nakai, T. Purification and characterization of three isoforms of chrysophsin, a novel antimicrobial peptide in the gills of the red sea bream, Chrysophrys major. Eur. J. Biochem. 2003, 270, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Han, H.M.; Gopal, R.; Park, Y. Design and membrane-disruption mechanism of charge-enriched AMPs exhibiting cell selectivity, high-salt resistance, and anti-biofilm properties. Amino Acids 2016, 48, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wu, S.; Jing, C.; Zhang, N.; Liang, D.; Xu, A. Identification, synthesis and characterization of a novel antimicrobial peptide HKPLP derived from Hippocampus kuda Bleeker. J. Antibiot. 2012, 65, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, S.-F.; Jin, Y.; Xu, X.; Qiao, Y.; Wu, Y.; Mao, Y.; Su, Y.-Q.; Wang, J. Characterization of a novel piscidin-like antimicrobial peptide from Pseudosciaena crocea and its immune response to Cryptocaryon irritans. Fish Shellfish. Immunol. 2013, 35, 513–524. [Google Scholar] [CrossRef]
- McDonald, M.; Mannion, M.; Pike, D.; Lewis, K.; Flynn, A.; Brannan, A.M.; Browne, M.J.; Jackman, D.; Madera, L.; Power Coombs, M.R.; et al. Structure–function relationships in histidine-rich antimicrobial peptides from Atlantic cod. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 1451–1461. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.M.; Ruangsri, J.; Kiron, V. Atlantic cod piscidin and its diversification through positive selection. PLoS ONE 2010, 5, e9501. [Google Scholar] [CrossRef] [Green Version]
- Salzet, M.; Verger-Bocquet, M. Elements of angiotensin system are involved in leeches and mollusks immune response modulation. Mol. Brain Res. 2001, 94, 137–147. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Wang, H.-X.; Wang, Y.-P. Gut microbiota-brain axis. Chin. Med. J. 2016, 129, 2373–2380. [Google Scholar] [CrossRef]
- Grenham, S.; Clarke, G.; Cryan, J.; Dinan, T. Brain–Gut–Microbe Communication in Health and Disease. Front. Physiol. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Montiel-Castro, A.; González-Cervantes, R.; Bravo-Ruiseco, G.; Pacheco-Lopez, G. The microbiota-gut-brain axis: Neurobehavioral correlates, health and sociality. Front. Integr. Neurosci. 2013, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Downing, J.E.G.; Miyan, J.A. Neural immunoregulation: Emerging roles for nerves in immune homeostasis and disease. Immunol. Today 2000, 21, 281–289. [Google Scholar] [CrossRef]
- Das, P.K.; Salinas, I. Fish nasal immunity: From mucosal vaccines to neuroimmunology. Fish Shellfish. Immunol. 2020, 104, 165–171. [Google Scholar] [CrossRef]
- Sundin, L.; Nilsson, S. Branchial innervation. J. Exp. Zool. 2002, 293, 232–248. [Google Scholar] [CrossRef]
- Tian, L.; Ma, L.; Kaarela, T.; Li, Z. Neuroimmune crosstalk in the central nervous system and its significance for neurological diseases. J. Neuroinflamm. 2012, 9, 155. [Google Scholar] [CrossRef] [Green Version]
- Ransohoff, R.M.; Brown, M.A. Innate immunity in the central nervous system. J. Clin. Investig. 2012, 122, 1164–1171. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Verri, W.A.; Chiu, I.M. Nociceptor Sensory Neuron–Immune Interactions in Pain and Inflammation. Trends Immunol. 2017, 38, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, E.Z.M.; Jamur, M.C.; Oliver, C. Mast Cell Function: A New Vision of an Old Cell. J. Histochem. Cytochem. 2014, 62, 698–738. [Google Scholar] [CrossRef]
- Kim, N.; Yun, M.; Oh, Y.J.; Choi, H.-J. Mind-altering with the gut: Modulation of the gut-brain axis with probiotics. J. Microbiol. 2018, 56, 172–182. [Google Scholar] [CrossRef]
- Mazzoli, R.; Pessione, E. The Neuro-endocrinological Role of Microbial Glutamate and GABA Signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef] [Green Version]
- Bhandage, A.K.; Barragan, A. GABAergic signaling by cells of the immune system: More the rule than the exception. Cell. Mol. Life Sci. 2021, 78, 5667–5679. [Google Scholar] [CrossRef]
- Xu, J.; Zanvit, P.; Hu, L.; Tseng, P.-Y.; Liu, N.; Wang, F.; Liu, O.; Zhang, D.; Jin, W.; Guo, N.; et al. The Cytokine TGF-β Induces Interleukin-31 Expression from Dermal Dendritic Cells to Activate Sensory Neurons and Stimulate Wound Itching. Immunity 2020, 53, 371–383.e375. [Google Scholar] [CrossRef]
- Liu, C.-W.; Su, B.-C.; Chen, J.-Y. Tilapia piscidin 4 (TP4) reprograms M1 macrophages to M2 phenotypes in cell models of Gardnerella Vaginalis-induced vaginosis. J. Front. Immunol. 2021, 5213. [Google Scholar] [CrossRef]
- Rakers, S.; Niklasson, L.; Steinhagen, D.; Kruse, C.; Schauber, J.; Sundell, K.; Paus, R. Antimicrobial Peptides (AMPs) from Fish Epidermis: Perspectives for Investigative Dermatology. J. Investig. Dermatol. 2013, 133, 1140–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunjani, N.; Ahearn-Ford, S.; Dube, F.S.; Hlela, C.; O’Mahony, L. Mechanisms of microbe-immune system dialogue within the skin. Genes Immun. 2021, 22, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R. Neuroimmune Interactions: From the Brain to the Immune System and Vice Versa. Physiol Rev. 2018, 98, 477–504. [Google Scholar] [CrossRef] [PubMed]
- Zaccone, G.; Lauriano, E.R.; Capillo, G.; Kuciel, M. Air-breathing in fish: Air-breathing organs and control of respiration: Nerves and neurotransmitters in the air-breathing organs and the skin. Acta Histochem. 2018, 120, 630–641. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
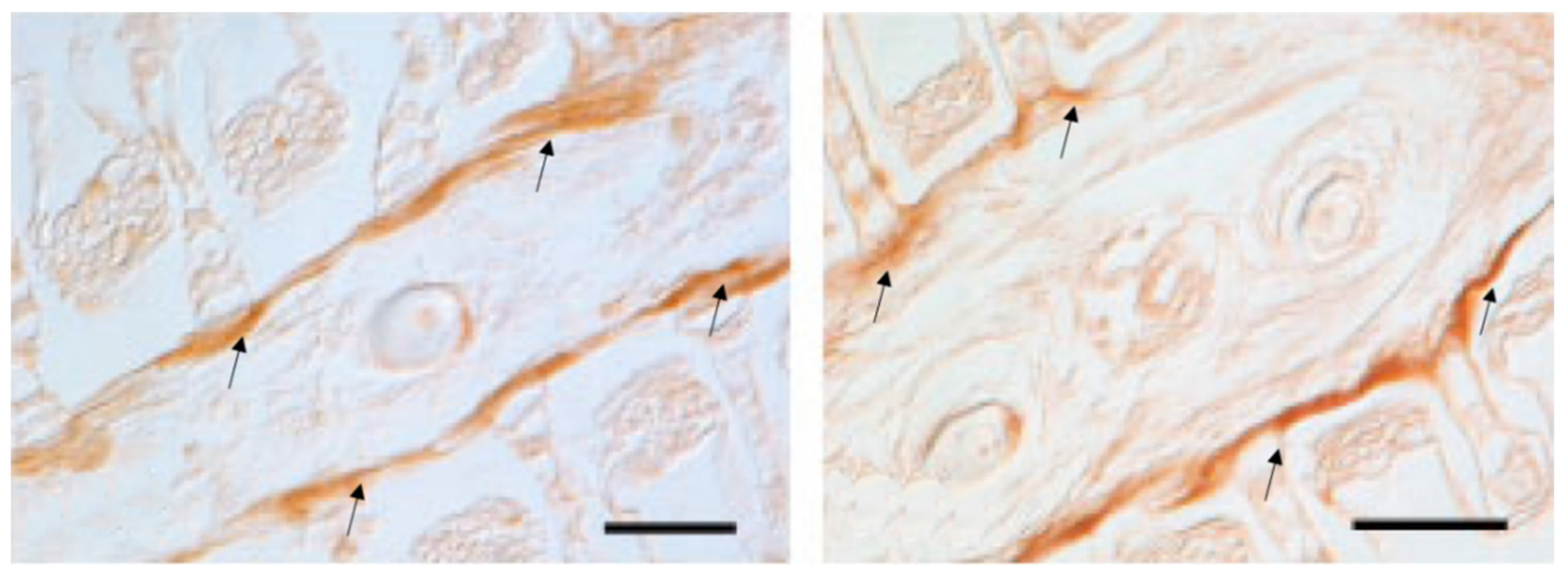{kind=link}
{kind=link}
| Peptide | Sequence | Properties | References |
|---|---|---|---|
| Piscidin 1 | FFHHIFRGIVHVGKTIHRLVTG | Anti-viral, anti-bacterial, anti-fungal, anti-parasitic and anticancer | [44,57] |
| Piscidin 2 | FFHHIFRGIVHVGKTIHKLVTG-NH2 | Anti-viral, anti-bacterial, anti-fungal, anti-mold and anti-parasitic | [48,58,59,60,61] |
| Piscidin 3 | FIHHIFRGIVHAGRSIGRFLTG | Anti-viral, anti-bacterial, anti- fungal and antiparasitic | [42,58,59,60] |
| Piscidin 4 | FFRHLFRGAKAIFRGARQGXRAHKVVSRYRNRDVPETDNNQEEP | Lowest hemolysis | [61,62] |
| Piscidin 5 | LIGSLFRGAKAIFRGARQGWRSHKA | Anti-bacterial, anti-fungal and anti-parasitic | [63,64] |
| Piscidin 6 | N/A | Antibacterial | [42] |
| Piscidin 7 | N/A | Antibacterial | [42] |
| Pathogens | Upregulation/ Downregulation | Organism | References |
|---|---|---|---|
| Gram-negative bacteria | |||
| Aeromonas salmonicida | Upregulation | Gadus morhua | [65] |
| Edwardsiella tarda | Up and downregulation | Oplegnathus fasciatus | [66] |
| LPS | Upregulation | Siniperca chuatsi, Chionodraco hamatus, Epinephelus coioides | [67,68,69] |
| Vibrio anguillarum | Up and downregulation | Dicentrarchus labrax | [42] |
| Gram-positive bacteria | |||
| Streptococcus iniae | Upregulation | Oplegnathus fasciatus | [66] |
| Virus and viral analogue | |||
| Lymphocystis iridovirus | Upregulation | Sparus aurata | [70] |
| Poly I:C | Upregulation | Chionodraco hamatus, Epinephelus coioides | [67,68] |
| Red seabream iridovirus (RSIV) | Upregulation | Oplegnathus fasciatus | [66] |
| Parasites | |||
| Acanthocephalus lucii | Upregulation | Perca fuviatilis | [71] |
| Chondracanthus, goldsmid | Upregulation | Latris lineata | [72] |
| Cryptocaryon irritans | Upregulation | Pseudosciaena crocea | [73] |
| Ergasilus sp. | Upregulation | Sparus aurata | [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaccone, G.; Capillo, G.; Fernandes, J.M.O.; Kiron, V.; Lauriano, E.R.; Alesci, A.; Lo Cascio, P.; Guerrera, M.C.; Kuciel, M.; Zuwala, K.; et al. Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction. Mar. Drugs 2022, 20, 145. https://doi.org/10.3390/md20020145
Zaccone G, Capillo G, Fernandes JMO, Kiron V, Lauriano ER, Alesci A, Lo Cascio P, Guerrera MC, Kuciel M, Zuwala K, et al. Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction. Marine Drugs. 2022; 20(2):145. https://doi.org/10.3390/md20020145
Chicago/Turabian StyleZaccone, Giacomo, Gioele Capillo, Jorge Manuel Oliveira Fernandes, Viswanath Kiron, Eugenia Rita Lauriano, Alessio Alesci, Patrizia Lo Cascio, Maria Cristina Guerrera, Michal Kuciel, Krystyna Zuwala, and et al. 2022. "Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction" Marine Drugs 20, no. 2: 145. https://doi.org/10.3390/md20020145