
Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents
by
Kai-Ling Wang
1,2,†, 
Zheng-Rong Dou
1,†,
Gao-Fen Gong
1,
Hai-Feng Li
1,
Bei Jiang
1 and
Ying Xu
2,* 1
Yunnan Key Laboratory of Screening and Research on Anti-Pathogenic Plant Resources from West Yunnan (Cultivation), Institute of Materia Medica, College of Pharmacy, Dali University, Dali 671000, China
2
Shenzhen Key Laboratory of Marine Bioresource & Eco-Environmental Science, Shenzhen Engineering Laboratory for Marine Algal Biotechnology, College of Life Sciences and Oceanography, Shenzhen University, Shenzhen 518060, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Mar. Drugs 2022, 20(2), 90; https://doi.org/10.3390/md20020090
Submission received: 25 November 2021
/
Revised: 13 January 2022
/
Accepted: 14 January 2022
/
Published: 21 January 2022
(This article belongs to the Special Issue Marine Natural Products with Antifouling Activity, 2nd Edition)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Bacteria growing inside biofilms are more resistant to hostile environments, conventional antibiotics, and mechanical stresses than their planktonic counterparts. It is estimated that more than 80% of microbial infections in human patients are biofilm-based, and biofouling induced by the biofilms of some bacteria causes serious ecological and economic problems throughout the world. Therefore, exploring highly effective anti-biofilm compounds has become an urgent demand for the medical and marine industries. Marine microorganisms, a well-documented and prolific source of natural products, provide an array of structurally distinct secondary metabolites with diverse biological activities. However, up to date, only a handful of anti-biofilm natural products derived from marine microorganisms have been reported. Meanwhile, it is worth noting that some promising antifouling (AF) compounds from marine microbes, particularly those that inhibit settlement of fouling invertebrate larvae and algal spores, can be considered as potential anti-biofilm agents owing to the well-known knowledge of the correlations between biofilm formation and the biofouling process of fouling organisms. In this review, a total of 112 anti-biofilm, anti-larval, and anti-algal natural products from marine microbes and 26 of their synthetic analogues are highlighted from 2000 to 2021. These compounds are introduced based on their microbial origins, and then categorized into the following different structural groups: fatty acids, butenolides, terpenoids, steroids, phenols, phenyl ethers, polyketides, alkaloids, flavonoids, amines, nucleosides, and peptides. The preliminary structure-activity relationships (SAR) of some important compounds are also briefly discussed. Finally, current challenges and future research perspectives are proposed based on opinions from many previous reviews.
1. Introduction
Bacterial biofilms are surface, or interphase-attached microorganism communities that are encapsulated in a self-secreted extracellular matrix comprising largely of proteins, polysaccharides, nucleic acids, and lipids [1,2]. Biofouling, defined as the undesirable growth of micro- and macro-organisms on submerged surfaces, causes serious ecological and economic problems throughout the world [3]. In natural seawater, all substrata are quickly fouled by marine fouling organisms, which is a complex process [4,5]. During the biofouling process, the formation of a biofilm acts as the glue that binds macro-foulers to the surface [2,5,6]. Biofilm formation consisting of microbes and microalgae is often the first step in the progression of biofouling development and is also increasingly recognized to play an essential role in the colonization of marine macro-foulers [5,6,7,8]. Within biofilms, microorganisms interact with a settlement of larvae of fouling invertebrates and algal spores [7,9].
Microbial biofilms formed by certain kinds of pathogenic and fouling bacteria play an adverse role in the medical and marine industries. Over 80% of microbial infections in the body are induced by biofilms [10]. The CDC declares that biofilms have become one of the most important medical hurdles of the century [11]. At the same time, it should be noted that biofouling based on the biofilm formation of fouling microbes causes enormous economic losses annually to the maritime and medical industries [2,5,6,7,8,9,10,11]. There is an urgent demand for exploring effective anti-biofilm agents to combat biofilms. Marine microorganisms, owing to their unique living environment, are extremely rich in natural products with diverse biological activities [12]. However, to our best knowledge, only a handful of anti-biofilm secondary metabolites of marine microbes and their mechanisms of action have been reported in the past few decades [13,14,15]. A massive amount of genomic and proteomic data generated allowing for further illustrations of the unknown molecular mechanisms in the association of biofilm formation and drug resistance [14,15]. The present review primarily focuses on modes of action of some anti-biofilm compounds via interfering with the quorum-sensing pathways, the disruption of extracellular polymeric substances, and adhesion mechanisms reported in the literature from 2000 to 2021.
As mentioned in many previous reviews [5,6,7,9], the development of a biofilm on a substratum could attract the biofouling of marine invertebrates (such as the barnacle Balanus amphitrite, the bryozoan Bugula neritina, the polychaete Hydroides elegans, the mussel Mytilus coruscus, and so on) and algae through physically modifying surfaces and releasing chemical compounds. That means numerous antifouling (AF) compounds, especially some environmentally friendly anti-larval/anti-algal agents with an EC50 value of <5 μg/mL and a LC50/EC50 ratio of >50, could serve as a potential source for the exploitation of highly effective and low- or non-toxic anti-biofilm candidates. More encouragingly, in the past two decades, marine microorganisms have provided a large number of potential AF natural products, some of which can be considered a promising source for the further exploration of potent anti-biofilm agents according to some previous reviews [5,7,16,17,18,19,20].
This review focuses on the anti-biofilm, anti-larval, and anti-algal secondary metabolites of marine microorganisms from 2000 up to the end of 2021. A total of 112 important active natural products, including 12 anti-biofilm compounds and 122 AF compounds, together with their 26 synthetic analogs, are described and highlighted with examples. Meanwhile, the preliminary analysis of the structure-relationship (SAR) of some compounds is discussed briefly. All selected compounds are sequentially introduced based on their chemical structure types in the relevant sections of this review. The further direction in the search for anti-biofilm/AF natural products from marine microbes is prosed in the concluding remarks.
2. Anti-Biofilm and Antifouling Natural Products from Marine Bacteria
Marine bacteria are well-recognized as the largest microbial group in the marine environment. Several anti-larval and anti-algal natural products have been isolated from diverse marine bacteria, including the species of the bacterial genera Shewanella, Pseudovibrio, Pseudoalteromonas, and Pseudomonas, and of the actinobacterial genus Streptomyces [21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42]. All secondary metabolites of these marine bacteria are described based on their chemical structures, including the following: fatty acids, butenolides, terpenoids, steroids, alkaloids, glycolipids, benzenoids, flavonoids, polyketides, and enzymes (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8).
2.1. Fatty Acids
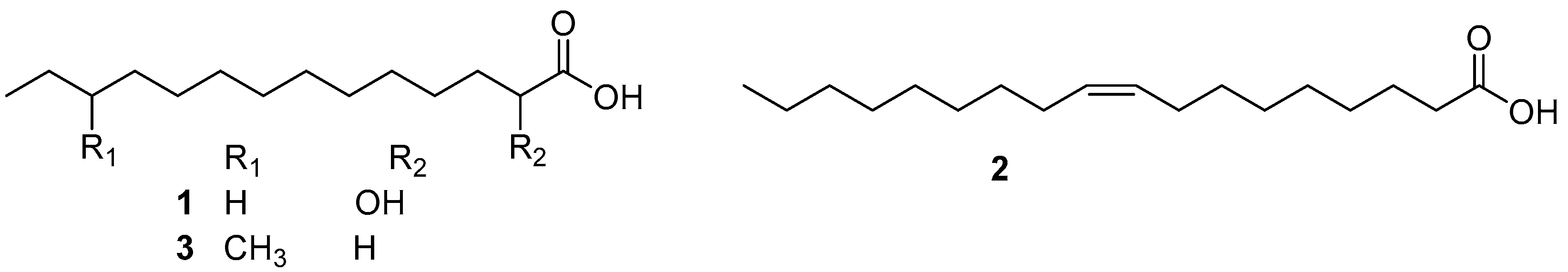
Isolation of AF compounds from the chloroform extract of a marine bacterium, Shewanella oneidensis SCH0402, yielded two fatty acids (S)-2-hydroxymyristic acid (HMA) (1) and cis-9-oleic acid (COA) (2) (Figure 1). During the one-and-a-half-year period of the field trial, both displayed inhibition for the germination of the representative soft fouling macro-algae Ulva pertusa spores at respective concentrations of 10 and 100 μg/mL, and strong AF activity as potent as TBTO against a wide range of micro- and macro-foulers. More importantly, these two fatty acids can be completely biodegraded, suggesting that this class of compounds can be used as environmentally friendly substitutes for toxic antifoulants [21]. Similarly, as an HMA-like homolog, 12-methyltetradecanoid acid (12-MTA) (3) (Figure 1), obtained from a deep-sea Streptomyces sp. through bioassay-guided isolation procedure, was found to have effective inhibition on larval settlement of H. elegans with EC50 value of 0.6 μg/mL, while its toxicity was very low with a high LC50/EC50 ratio of >133.5. Further study on AF mechanism revealed that the inhibitory effects of 12-MTA on the settlement of H. elegans larvae might be related to the down-regulation of GTPase-activating gene and up-regulation of ATP synthase gene [22].
2.2. Butenolides
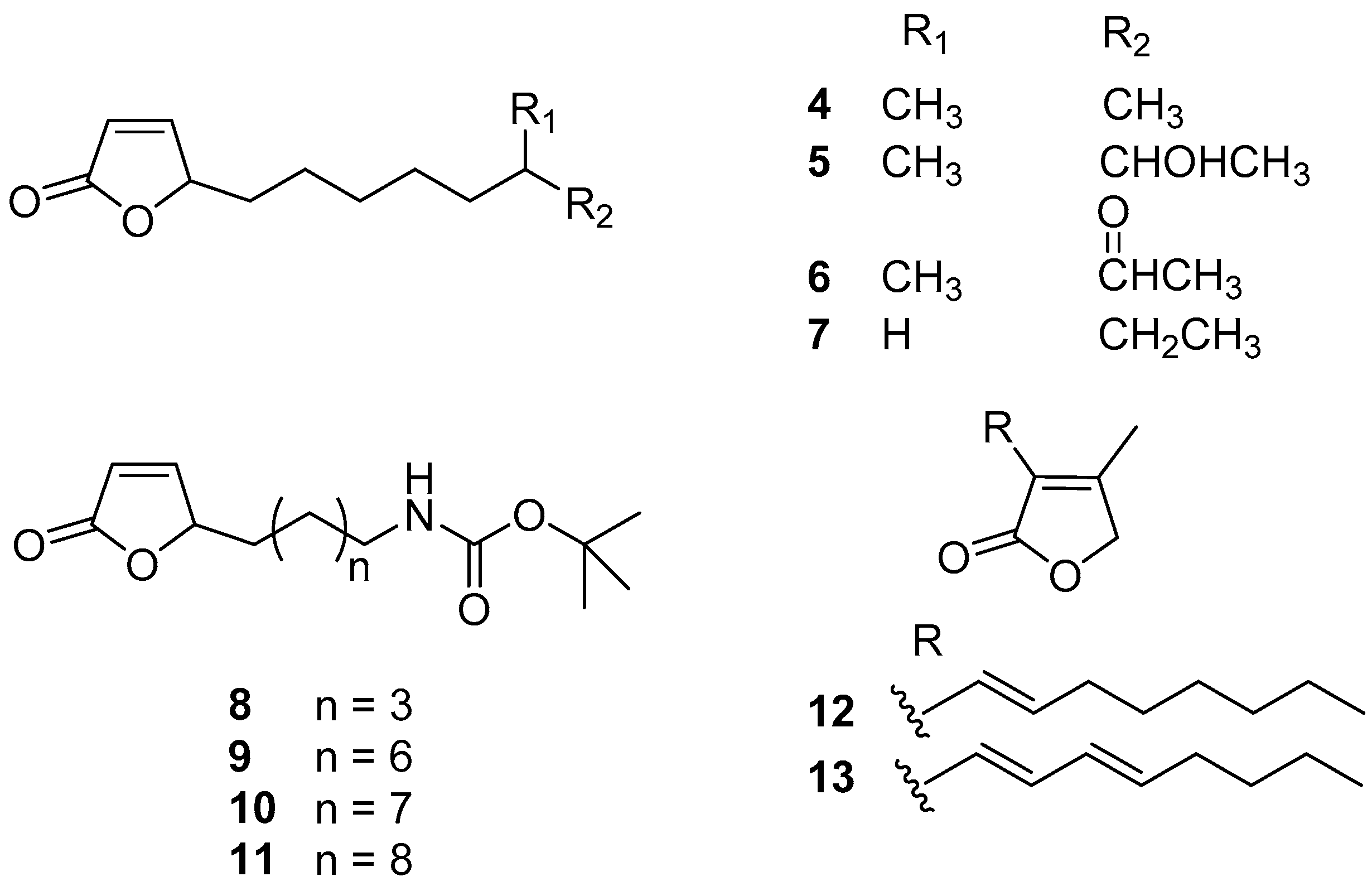
Actinobacteria is the largest phylum in the bacterial domain and has great potential for producing novel and bioactive natural products including antifoulants. A series of butenolides (Figure 2) were isolated from two actinobacterial strains, a deep-sea derived Streptomyces albidoflavus UST040711-291 and a North Sea derived Streptomyces sp. GWS-BW-H5. Among these butenolides, compounds 4–6 prevented larval settlement of B. amphitrite at concentrations less than 10.00 μg/mL, and their toxicity was weak [23,24]. Analysis of SAR revealed that 2-furanone ring, the common moiety, may be essential for AF activity, while the lipophilicity of the alkyl side-chain linked to the mother nucleus may explain their varying potency of AF activity. Based on these findings, compound 7 (5-octylfuran-2(5H)-one), named butenolide, structured with a straight alkyl chain exhibited significant and non-toxic anti-larval activity against settlement of B. amphitrite, B. neritina, and H. elegans larvae with EC50 values of 0.518, 0.199, and 0.0168 μg/mL and LC50/EC50 ratios higher than 97, 250, and 119, respectively. More importantly, this compound also displayed outstanding AF activity even at a concentration of 5% (w/w) in field tests, indicating that it might be a promising candidate for AF purposes [24]. Because of the excellent AF activity of butenolide, a dozen analogues of butenolide were synthesized for AF bioassays, among which the Boc-butenolide (8) structurally modified with a Boc-protecting-group at the terminal of the alkyl side-chain was demonstrated to have similar AF capabilities to butenolide in larval settlement bioassays but with significantly lower toxicity at high concentration of 25 μg/mL. Meanwhile, Boc-butenolide has good environmental stability with a higher melting point at 132 °C higher than that of butenolide at 23 °C. The coverage of biofouler on the Boc-butenolide coatings was low after 2 months in a marine field test, indicating the AF potential of Boc-butenolide [25]. The other three synthesized butenolide derivatives 9–11 showed significant inhibition for larval settlement of B. amphitrite with EC50 values of 0.663, 0.722, and 0.827 μg/mL, respectively, whereas they were less toxic with the LC50/EC50 ratios >61, 73, and 63, respectively [26]. In addition, another two known natural butenolides, compounds 12 and 13, were isolated from a seaweed epibiotic bacterium S. violaceoruber SCH-09. Both of them displayed strong anti-larval activity against the mussel M. edulis with EC50 values of 0.02 and 0.1 μg/mL, while their toxicity is very low with high therapeutic ratios >92 [27]. Moreover, based on the investigations of mode of AF action from proteome and phosphoproteome profiles, Zhang et al. [28] and Qian et al. [29] suggested that butenolide prevented larval settlement of macro-fouling organisms B. amphitrite and B. neritina by modulating proteins involved in stress regulation and energy metabolism in larvae.
Recently, Ding et al. [30] reported anti-biofilm activity of this compound against mixed marine bacterial species in a field experiment. After one month, the panels coated with paints containing 10% (w/w) of butenolide were free of biofilm formation, and only a small amount of biofilm occurred on the panels with 2.5 and 5% of butenolide. According to the results of metatranscriptomic analysis, the microbial community structure of biofilm was visualized at the phylum level, including Bacteroidetes/Chlorobi, Chloroflexi, Gammaproteobacteria, Cyanobacteria, Actinobacteria, Chlamydiae/Verrucomicrobia, and Fibrobacteres/Acidobacteria. Soon afterwards, Yin et al. [31] reported that butenolide could have a broad spectrum of anti-biofilm activity against both Gram-positive and Gram-negative pathogenic model species, including Escherichia coli, Pseudomonas aeruginosa, and methicillin-resistant Staphylococcus aureus (MRSA) with MBIC values ranging from 50 to 800 mg/L. This compound could effectively inhibit biofilm formation and eradicate mature biofilms of all the tested strains. The results of confocal laser scanning microscope (CLSM) and scanning electron microscope (SEM) showed that butenolide greatly removed biofilms by decreasing biofilm coverage and thickness and destroying biofilm matrix of microbial strains. Perhaps more significantly, it was demonstrated that butenolide had the potential to work with commonly used antibiotics for fighting biofilm infections due to its synergistic/enhancing anti-biofilm manner as a tetracycline enhancer. Based on these findings, butenolides should be considered as an attractive anti-biofilm candidate.
2.3. Terpenoids and Steroids
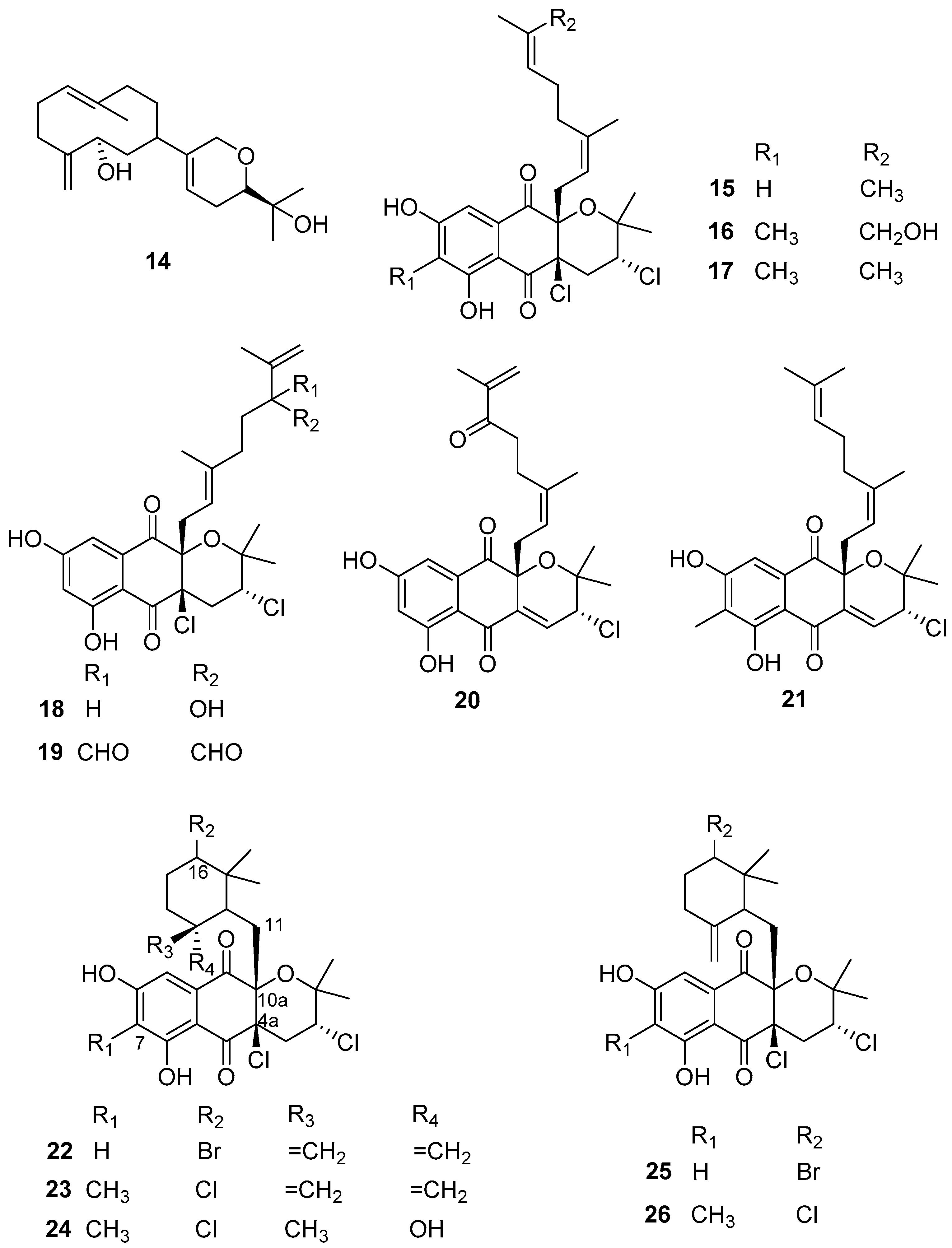
Some active anti-larval, anti-biofilm, and anti-bacterial terpenes (Figure 3) have been obtained from Streptomyces actinomycetes in recent years. The active AF diterpene compound 14, namely lobocompactol, was obtained from a marine-derived actinobacterial strain Streptomyces cinnabarinus PK209 and found to have excellent anti-algal activity against the alga U. pertusa and the diatom Navicula annexa with EC50 values of 0.18 and 0.43 μg/mL and LC50/EC50 ratios of 46.2 and 71.6, respectively. Lobocompactol (14) also inhibited the growth of the fouling bacteria Pseudomonas aeruginosa KNP-5 and Pseudomonas sp. KNP-8 with MIC values of 66 μg/mL and 112 μg/mL, respectively [32]. Napyradiomycins belongs to a class of hybrid isoprenoids and/or meroterpenoids whose structures contain one or more terpene moieties bound to a non-terpenoid scaffold [33]. They are known for their antimicrobial and anticancer activities [34]. Until now, to our best knowledge, 61 napyradiomycin derivatives have been discovered and elucidated [33,34]. Recently, 12 napyradiomycins 15–26 isolated from the ethyl acetate extracts of S. aculeolatus strains PTM-029 and PTM-420 were reported as potential AF and anti-biofilm components by Pereira et al. [34]. Specifically, compounds 15, 16, 18, and 24, and mixtures of 17 and 21, 19 and 20, 22 and 25, and 23 and 26 displayed AF activity against Mytilus galloprovincialis plantigrade larval settlements with EC50 values ranging from 0.10 to 6.34 µg/mL. Among these active napyradiomycins, the most effective ones were 15, 24, and 26, showing EC50 value < 1 µg/mL (EC50 values of 0.66, 0.10, and 0.95 µg/mL, respectively), and the mixtures of compounds 22 and 25, and compounds 23 and 26 with EC50 values of 0.73 µg/mL and 0.45 µg/mL, respectively. Importantly, all these compounds displayed low- or non-toxicity to the larvae of M. galloprovincialis with LC50/EC50 ratios >15 with the exception of compound 26. Meanwhile, some of these AF napyradiomycins showed anti-biofilm activities against the five species of marine fouling bacteria, including Marinobacter hydrocarbonoclasticus DSM 8798, Cobetia marina DSM 4741, Micrococcus luteus DSM 20030 and ATCC 4698, Pseudooceanicola batsensis DSM 15984, and Phaeobacter inhibens DSM 17395. Among these compounds, napyradiomycin (18), the mixtures of compounds 19 and 20, and compounds 22 and 25 were considered the most promising anti-biofilm components without inhibiting the growth of fouling bacteria assayed at the same concentration. Specifically, compound 18 showed significant biofilm inhibition (>90%) of M. hydrocarbonoclasticus at the lowest tested concentration of 0.98 µg/mL. The mixture of compounds 19 and 20 had no anti-bacterial activity at a concentration of 3.91 µg/mL, while it could prevent biofilm formation of M. luteus and M. hydrocarbonoclasticus DSM 8798 by 82.5 ± 5.1% and 43.8 ± 9.4%, respectively. The mixture of compounds 22 and 25 displayed anti-biofilm activity against M. hydrocarbonoclasticus was about 60% at all tested concentrations ranging from 0.98–15.60 µg/mL with no detected anti-bacterial activity, and completely abolished biofilm formation of M. luteus (100%) at the lowest concentration of 0.98 µg/mL but inhibited its growth by 70.5 ± 1.9%. Also, for the same mixture, significant inhibition of biofilm formation of P. inhibens (38.7 ± 0.7%) was observed at a concentration of 15.60 µg/mL, while not showing anti-bacterial effectiveness. The mixture of napyradiomycins 17 and 21 showed no anti-bacterial activity on these fouling microbes, but weak biofilm inhibition (>20%) for M. luteus and C. marina.
As shown in Figure 3, the chemical structure of napyradiomycins is characterized by the presence of a semi-naphthoquinone core, and herein all the reported napyradiomycins 15–26 consist of a prenyl unit attached at C-4a that is cyclized to form a tetrahydropyran ring and a monoterpenoid substituent at C-10a. The SAR analysis revealed that compounds 23, 24, and 26 containing the methyl group in the core structure at C-7 showed higher anti-larval activity. The 6-membered ring cyclized by the linear monoterpenoid subunit at C-10a in the most active compounds 22–26 may serve as an important functional group for enhancing the anti-larval activity of these napyradiomycin derivatives. Interestingly, three napyradiomycins 18, 22, and 25 with the highest anti-biofilm activity were obtained from the strain PTM-420, and contained a hydrogen atom at C-7. Conversely, napyradiomycins from PTM-029 generally had higher anti-biofilm activity with the methyl group at C-7. Additionally, napyradiomycins 22 and 25 stand out due to their broad-spectrum AF activity against macro- and micro-fouling organisms, which may be related with their chemical feature of a bromine substitute at C-16. Altogether, these above-mentioned natural anti-biofilm napyradiomycins should be further developed as promising anti-biofilm candidates that will not contribute to antibiotic/biocide resistance.
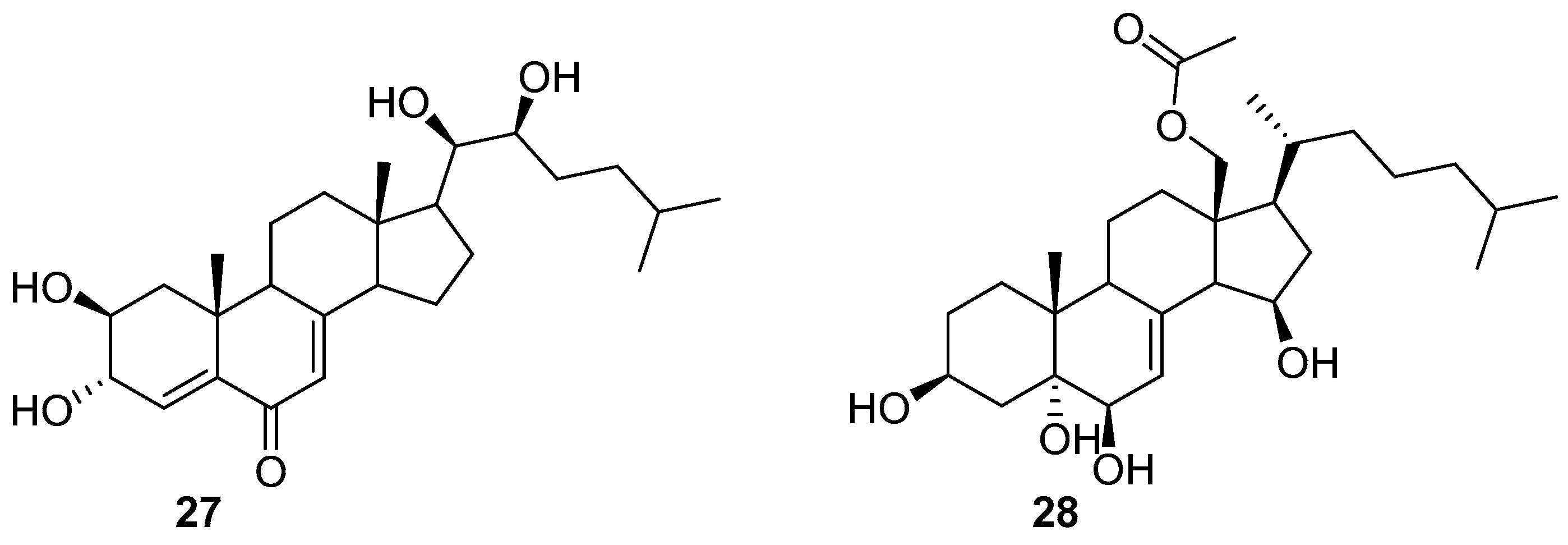
Only two steroids 27 and 28 (Figure 4) were isolated from a filamentous bacterium Leucothrix mucor, and found to prevent the spore settlement of the algae U. pertusa zoospores with EC50 values of 1.2 and 2.1 μg/mL as well as the growth of a biofouling diatom Navicula annexa with EC50 values of 5.2 and 7.5 μg/mL, respectively. The two compounds also exhibited anti-bacterial activity against the fouling bacterial strains Pseudomonas aeruginosa KNP-3 with MIC values of 32 and 56 μg/mL and Alteromonas sp. KNS-8 with MIC values of 66 and 90 μg/mL, respectively [35].
2.4. Anthraquinones
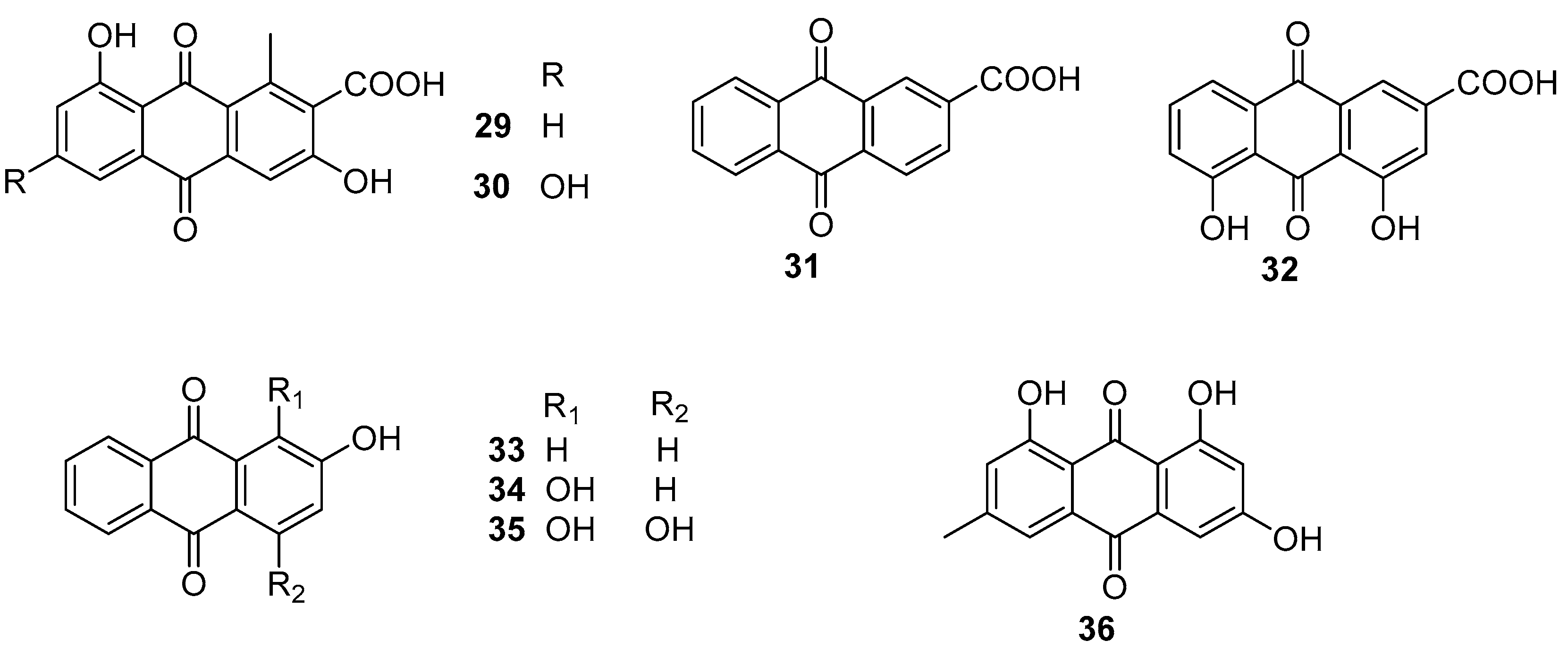
Two natural anthraquinones 29 and 30 (Figure 5) isolated from a rare actinobacterial strain Kitasatospora albolonga R62, together with their 10 commercial analogs were evaluated for antibiofilm activities against MRSA [36]. Among them, compounds 31–36 (Figure 5) could all effectively inhibit the formation of MRSA biofilms by >50%. It is noted that the inhibition rates of compounds 32 and 33 were up to 75.2% (±3.3%) and 96.4% (±0.3%). Also, both of these two compounds showed excellent eradication activity for preformed MRSA biofilms at concentrations of 200 and 50 mg/mL. The structure-activity relationship analysis of these anthraquinones suggested that the hydroxyl group attached at C-2 position of the anthraquinone skeleton should be considered as a key group for inhibiting biofilm formation at high concentrations, and the carboxyl group at the same C-2 position should play an important role in improving the eradication activity of these anthraquinones. The results of RNA sequencing indicated that the disruption of phosphate homeostasis of MRSA might be involved in the mode of action of compounds 32 and 33.
2.5. Alkaloids
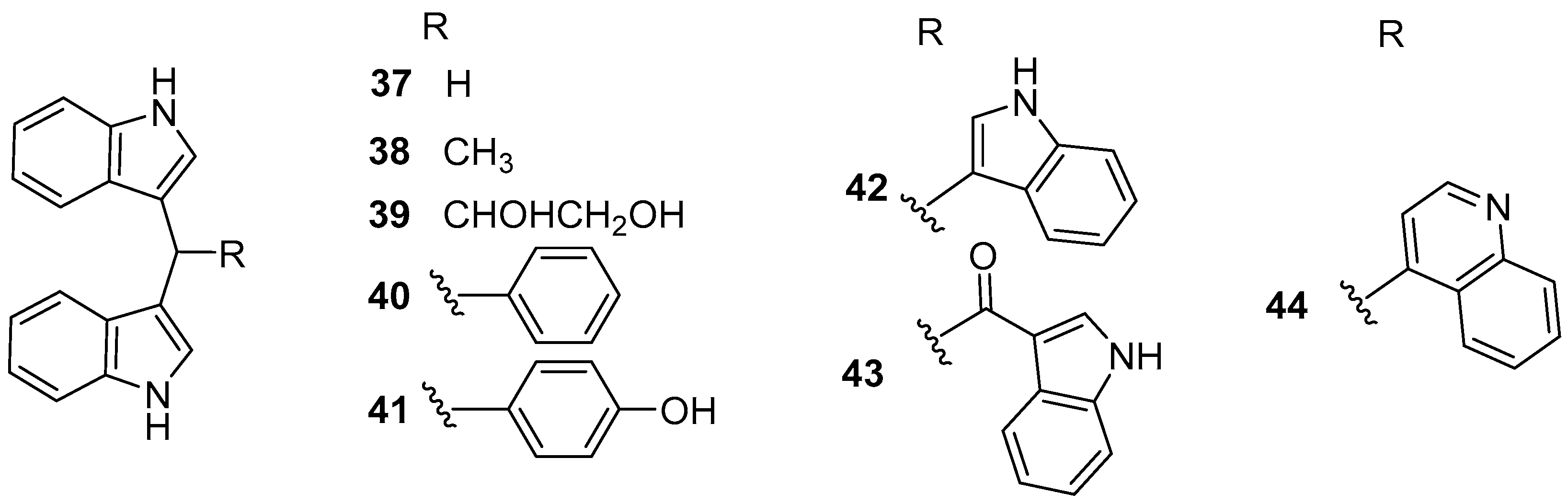
Eight bisindole alkaloids 37–44 (Figure 6), belonging to di (1H-indol-3-yl) methane (DIM) family, were isolated from a Red Sea ascidian-derived bacterium Pseudovibrio denitrificans UST4-50 [37]. All of them exhibited moderate to strong inhibitory activity against larval settlement of B. amphitrite with EC50 values ranging from 0.63 to 5.68 μg/mL, among which DIM (37) and 4-(di(1H-indol-3-yl) methyl)phenol (DIM-Ph-4-OH) (40) displayed excellent AF activity against larval attachment of B. neritina with EC50 values of 0.62 and 0.42 μg/mL, respectively, while their toxicity was very low with both LC50/EC50 ratios >60. Interestingly, the inhibitory ability of these two compounds for settlement of B. amphitrite cyprids was reversible, indicating special AF effects on macro-fouling invertebrates’ larvae. More encouragingly, the mixed paints containing 10–15% (w/w) DIM and basal ingredients showed comparable AF activity to the positive antifoulant Sea-Nine 211™ for 5 months or even longer in the field testing, which suggested that some potential AF DIMs might be considered as promising antifoulant candidates. The preliminary SAR of these DIMs was also discussed. It was found that the mother nucleus, di(1H-indol-3-yl) methylene, might be the functional group responsible for anti-larval activity, and the phenolic hydroxyl group linked to the Ph-C1′ would play a crucial role in increasing the anti-larval activity of these DIMs. Moreover, the supply issue of DIM has been easily solved by chemical synthesis or market purchases because of its simple structure. Further studies on AF characteristics of DIM and DIM-Ph-4-OH, such as the anti-algal activity against fouling algae, anti-biofilm activity against fouling bacteria, the evaluation of toxicity toward non-target organisms, AF mechanism using proteomic methods, and so on are in progress in our laboratory.
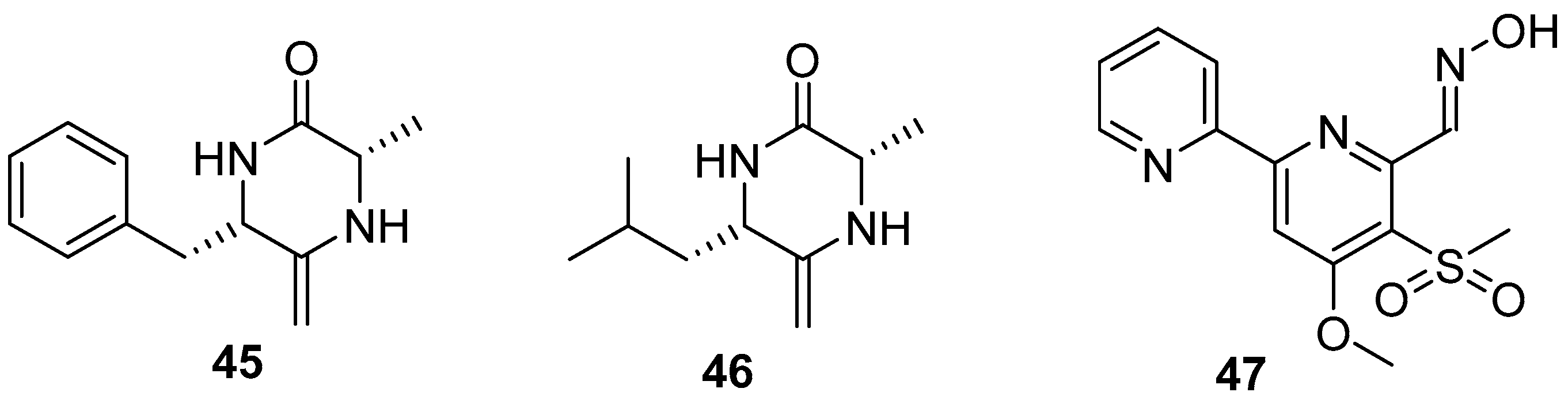
Two diketopiperazines (DKPs) (Figure 7), (6S,3S)-6-benzyl-3-methyl-2,5-diketopiperazine (bmDKP) (45) and (6S, 3S)-6-isobutyl-3-methyl-2,5-diketopiperazine (imDKP) (46), were obtained from a seaweed Undaria pinnatifida–derived Streptomyces praecox 291-11. Both bmDKP and imDKP could inhibit zoospore settlement of the seaweed U. pertusa with EC50 values of 2.2 and 3.1 μg/mL and the growth of the diatom N. annexa with EC50 values of 0.8 and 1.1 μg/mL, respectively. Compared to the low LC50/EC50 ratios of positive controls (being 2 and 6, respectively) toward the target algae, both compounds showed higher therapeutic ratios with LC50/EC50 ratios of 17.7 and 21 to U. pertusa and 263 and 120.2 to N. annexa, which indicated that these two compounds might be considered as non-toxic anti-algal compounds [38]. As a novel compound, Maipomycin A (MaiA) (47) (Figure 7), bearing a unique 2,20-bipyridine structure and an unusual oxime functionality, was isolated from a rare actinomycete strain Kibdelosporangium phytohabitans XY-R10. This compound showed strong anti-biofilm activities against Gram-negative bacteria including Acinetobacter baumannii ATCC 19606 and Pseudomonas aeruginosa ATCC 27853 with MBIC values of 8 and 32 mg/mL, respectively. More interestingly, it has demonstrated that this natural product might exert its antibiofilm activity through chelating irons [39].
2.6. Glycolipids, Benzenoids, Flavonoids, Polyketides, and Enzymes
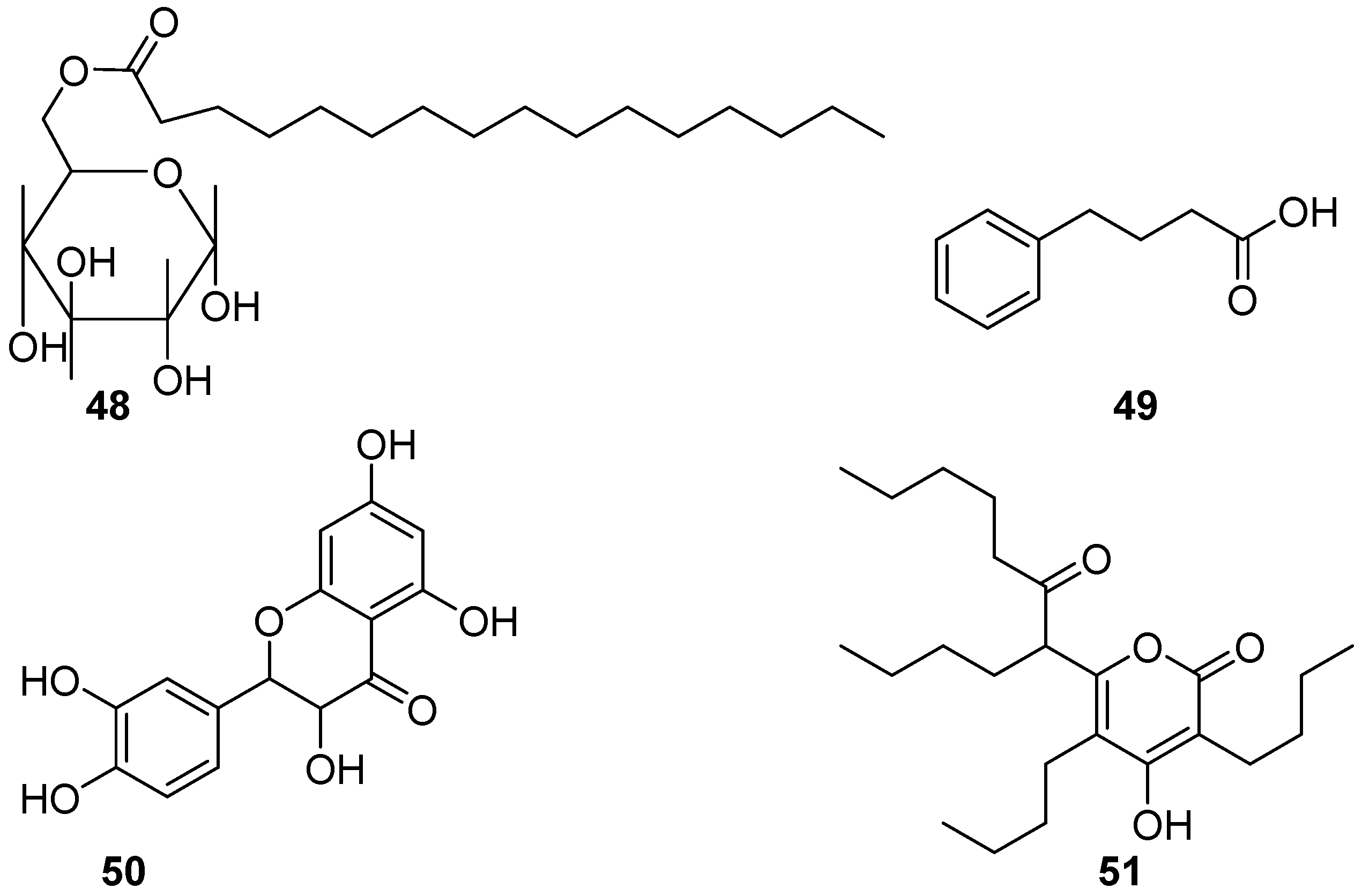
Besides some bioactive compounds mentioned above, few of the glycolipids, benzenoids, flavonoids, and polyketides (Figure 8) derived from marine bacteria have also been evaluated for anti-biofilm and AF activities, and revealed a good level of effectiveness. A glycolipid surfactant 48 composed of glucose and palmitic acid was produced by a tropical marine strain of Serratia marcescens [40]. It showed anti-biofilm potential for preventing the adhesion of the marine biofouling bacterium Bacillus pumilus TiO1 with EC50 value of 6.25 μg/mL, and disrupting preformed biofilms of this culture in microtiter plates at concentrations ranging from 50 to 100 μg/mL. Interestingly, a marine-derived bacterial strain S6-15 that actually affiliates with the above-mentioned fouling species B. pumilus, could produce a novel 4-phenylbutanoic acid (49), which combated biofilm formation in all tested Gram-positive and Gram-negative species at concentrations ranging from 10–15 μg/mL [41]. Although dozens of flavonoids are widely distributed in the secondary metabolites of marine microorganisms and reported as bioactive compounds, herein only one flavonoid compound taxifolin (50) from a mangrove derived Streptomyces sampsonii PM33 was found to have AF activity [42]. In field experiment, marine surfaces of the test PVC panels coated with taxifolin (0.5 and 1.0 mg) did not have adherence of micro- or macro-fouling organisms even after 4 weeks. Further toxicity assays based on zebrafish models showed the lower toxicity of taxifolin at effective concentrations. These results indicate that taxifolin should be a promising candidate for the development of eco-friendly antifoulants. During the AF natural products project screening, the polyketide compound elasnin (51) highly produced by Streptomyces mobaraensis DSM 40847 exhibited strong biofilm inhibition activity against four marine bacterial Gram-positive strains Staphylococcus aureus B04, S. hominis N32, S. arlettae OM, and Microbacterium esteraromaticum N22 with MBIC90 and MBIC50 ranged from 2.5 to 5 µg/mL and 1.25 to 5 µg/mL, whereas three gram-positive strains Idiomarina sediminum N28, Pseudoalteromonas sp. L001, and Escherichia coli N57 with MBIC90 of 5 to 10 µg/mL and MBIC50 of 1.25 to 10 µg/mL, respectively [43]. The larval settlement of Balanus amphitrite was also effectively inhibited at concentrations above 12.5 µg/mL after 24 h exposure. Encouragingly, elasnin-based coatings showed significant performance in reducing the species richness and diversity of biofilms and the attachment of large biofouling organisms in the marine environment with a concentration of 2.5 wt% in the first 2 weeks. In addition, a low toxicity of elasnin towards B. amphitrite larvae was observed with a mortality rate of larvae around 10% under the concentration of 25 µg/mL compared with the control groups. These findings indicate that elasnin has a huge development potential for anti-biofilm and AF agents. So far, only a novel protease produced by a deep-sea bacterial strain Pseudoalteromonas issachenkonii UST041101-043, showed drastically strong AF effects on the settlement of B. neritina larvae with EC50 value of 0.5 ng/mL, and significant inhibition for larval settlement of the barnacle B. amphitrite and the bryozoan Schizoporella sp. at a very low concentration of 100 ng/mL [44].
3. Antifouling Natural Products from Marine Cyanobacteria
Marine cyanobacteria are a bountiful source of novel bioactive metabolites, but the dearth of AF compounds isolated from cyanobacteria is still surprising nowadays. During the last two decades, only seven anti-larval secondary metabolites were obtained from marine-derived cyanobacteria [45,46,47,48,49], and there was no relevant report about anti-biofilm natural products. According to the chemical structures, these anti-larval are categorized into butenolides and polyketide–polypeptide structural families (Figure 9 and Figure 10).
3.1. Butenolides
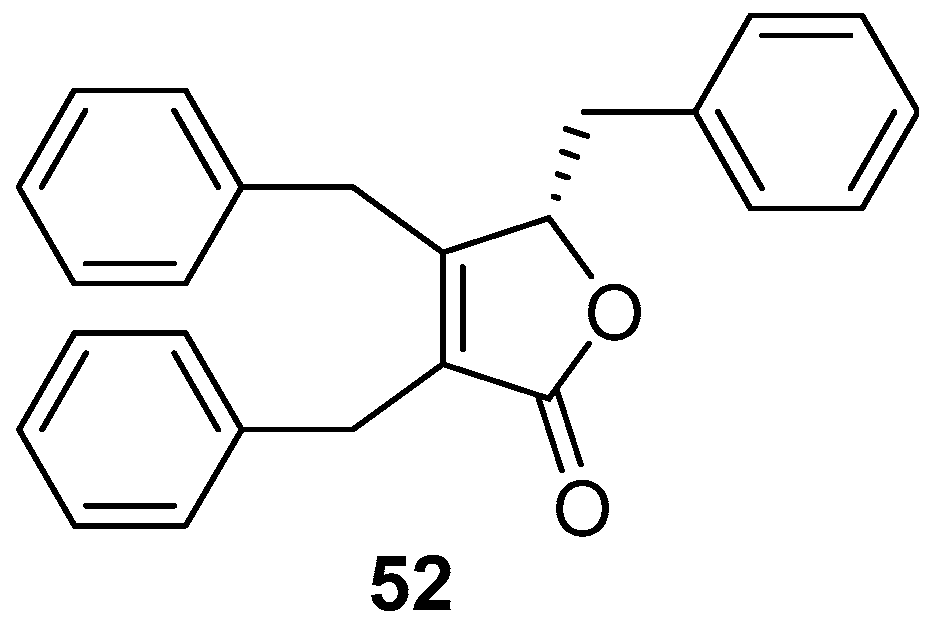
A well-known butenolide compound maculalactone A (52) (Figure 9), as a “natural” marine AF agent from the marine cyanobacterium Kyrtuthrix maculans, exhibited excellent inhibitory activity against specific bivalve settlers at concentrations of 0.1% and 1% (w/w) in the preliminary field investigation, but was toxic to the naupliar larvae of the barnacles B. amphitrite, Tetraclita japonica, and Ibla cumingii with LC50 values ranging from 1.1–5.2 μg/mL [45]. And the stereochemistry of this natural product has been recently assigned by total synthesis [46].
3.2. Polyketide–Polypeptide Structural Compounds
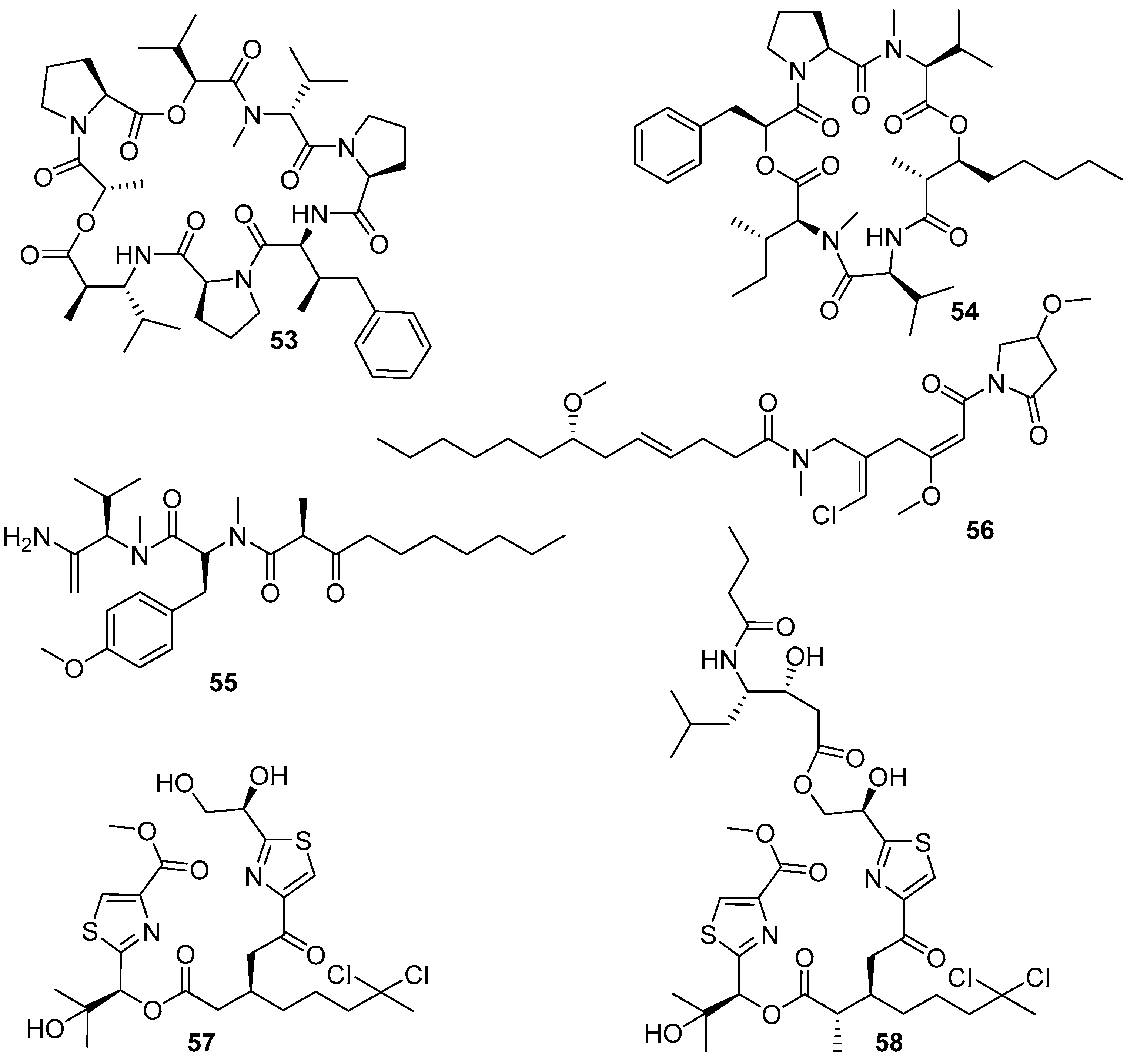
The polyketide–polypeptide structural compound dolastatin 16 (53) (Figure 10), together with its three derivatives 54–56 (Figure 10), was isolated from the benthic filamentous marine cyanobacterium Lyngbya majuscule. All these four compounds showed anti-larval activity against the barnacle B. amphitrite larvae with EC50 values of 0.003, 10.6, 0.54, and 2.6 μg/mL, respectively. The most attractive dolastatin 16 has a good therapeutic ratio with LC50/EC50 ratio > 6000, and barnacle settlement on the dolastatin 16-treated plates was significantly reduced compared to negative controls at all the tested concentrations of 10.0, 1.0, 0.1, and 0.01 μg/mL, thus indicating its potential development prospects as a non-toxic antifoulant. Meanwhile, compounds 55 and 56 also displayed low toxicity with LC50/EC50 values of 167.7 and 77.6 except compound 54 with LC50/EC50 value of 6.7 [47]. Recently, natural dolastatin 16 was also found in the secondary metabolites of another marine cyanobacterium Okeania sp. [48], and the total synthesis of dolastatin 16 has been successfully achieved [50]. Both the synthetic and Okeania sp.-derived compounds showed similarly remarkable AF activity against B. amphitrite cyprid larvae to that of the previous L. majuscula-derived dolastatin 16, which further indicated that dolastatin 16 might serve as a potent AF compound for antifoulant candidates without concern about supply issues. Additionally, the strain of Okeania sp. could produce a series of AF lyngbyabellins, also belonging to the polyketide–polypeptide family, of which lyngbyabellins O (57) showed the most inhibitory effects on larval settlement of B. amphitrite with an EC50 value of 0.24 μg/mL, while its analogue lyngbyabellins P (58) was less active with an EC50 value of 0.62 μg/mL [48]. The investigation of SAR proposed that the acyclic form without a side-chain in compound 57 might be a functional structural framework for improving anti-larval activity of lyngbyabellins.
4. Anti-Biofilm and Antifouling Natural Products from Marine Fungi
Marine fungi are a main source of secondary metabolites with various biological activities and chemical structures, and offer more diverse anti-biofilm and AF natural products than marine bacteria and cyanobacteria. In this part, all natural products from marine fungi and their analogues are categorized into terpenoids, phenols, phenyl ethers, polyketides, alkaloids, amines, nucleosides, and peptides (Figure 11, Figure 12, Figure 13, Figure 14, Figure 15, Figure 16, Figure 17, Figure 18, Figure 19 and Figure 20).
4.1. Terpenoids
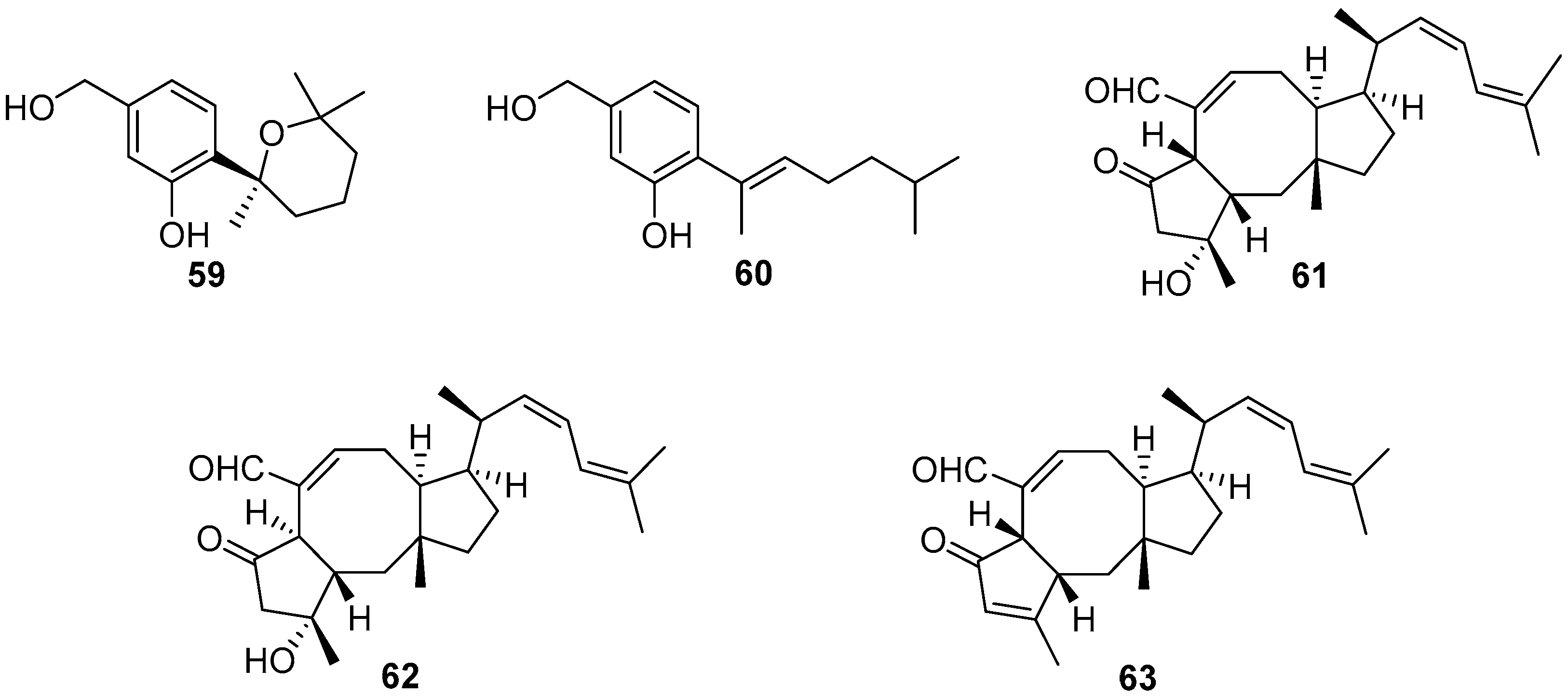
Four bisabolane-type sesquiterpenoids were isolated from a sponge-derived fungus Aspergillus sp., among which compounds 59 and 60 (Figure 11) could completely prevent larval settlement of B. amphitrite at a concentration of 25.0 μg/mL [51]. However, compound 59 was observed to have an obvious toxic effect on the larvae at its effective concentration. Three sesterterpenes named ophiobolin K (61), 6-epi-ophiobolin K (62), and 6-epi-ophiobolin G (63) (Figure 11) were obtained from the fermentation broth of a marine-derived fungus Emericella variecolor, and demonstrated to inhibit biofilm formation of Mycobacterium species to different degrees. Among them, ophiobolin K exhibited the best anti-biofilm activity against M. smegmatis and M. bovis BCG with MIC values of 1.58 and 3.15 μg/mL, respectively [52]. More importantly, ophiobolin K also has the ability to restore the antibacterial activity of isoniazid against M. smegmatis by preventing biofilm formation.
4.2. Phenols and Phenyl Ethers
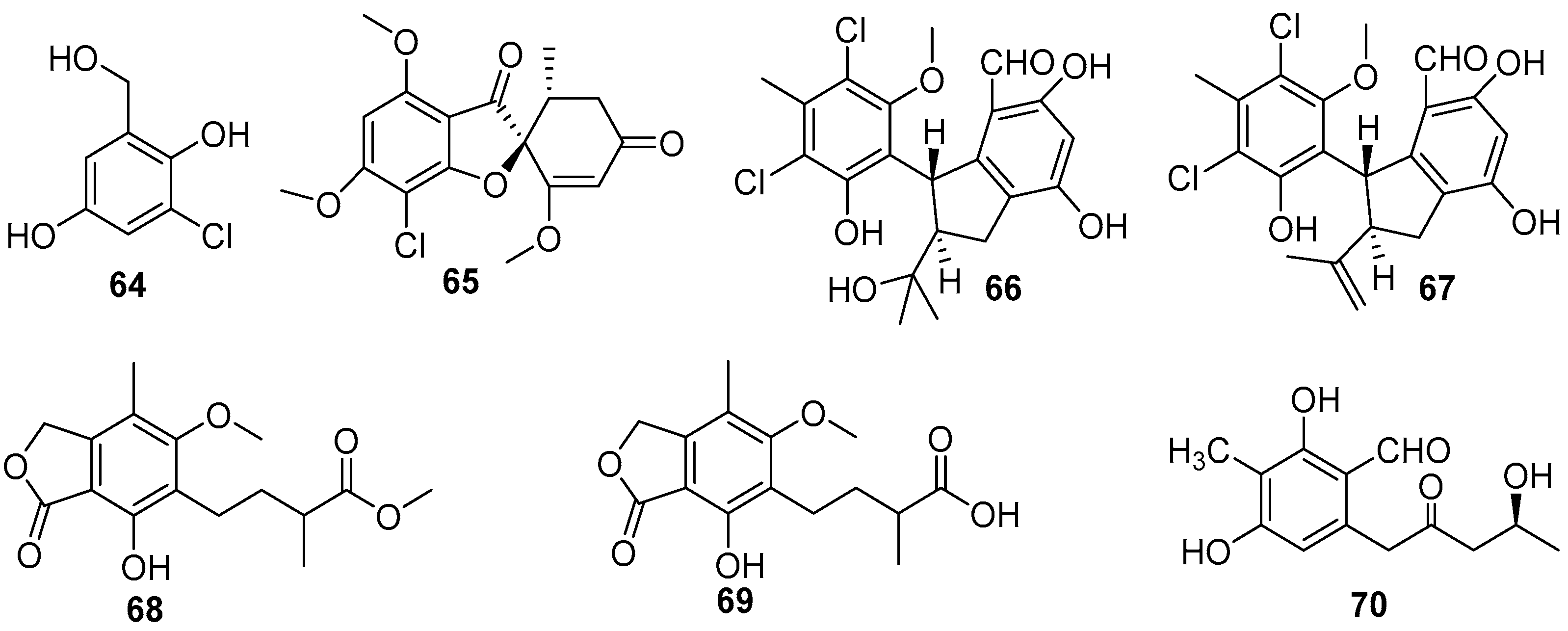
Over the past two decades, dozens of benzenoids, especially phenols and phenyl ethers, have been isolated from marine fungi, and evaluated for AF activity, which led to the discovery of some effective anti-larval compounds. A series of phenol compounds were obtained from several species of four fungal genera, consisting of Ampelomyces, Sarcophyton, Pestalotiopsis, Aspergillus, and Penicillium [53,54,55,56]. Bioassay-guided isolation of crude extracts of Ampelomyces and Pestalotiopsis strains yielded four AF chlorinated phenols 64–67 (Figure 12) [53,54]. The compound 3-chloro-2, 5-dihydroxybenzyl alcohol (64) from Ampelomyces sp. UST040128 could inhibit larval attachment of B. amphitrite and H. elegans with EC50 values ranging from 3.192 to 3.812 μg/mL and from 0.672 to 0.78 μg/mL respectively. Meanwhile, its toxicity toward the B. amphitrite cyprids is very low with LC50 value of 266.682 μg/mL, yet it is highly toxic to the larvae of H. elegans with LC50 value of 2.642 μg/mL [51]. The amibromdole (65) was obtained from a soft coral-derived fungus Sarcophyton sp., and exhibited weak anti-larval activity against settlement of B. amphitrite larvae with EC50 value of 16.70 μg/mL [45]. Another two chlorinated phenols (±)-pestalachlorides E (66) and F (67) were isolated from Pestalotiopsis sp. ZJ-2009-7-6, showing excellent inhibition on settlement of B. amphitrite larvae with EC50 values of 1.65 and 0.55 μg/mL, and were not toxic to the cyprids with respective LC50/EC50 ratios >30.3 and >18.2 [47]. Three phenolic acids 68–70 (Figure 12), were obtained from two deep-sea-derived fungal strains Penicillium brevicompactum DFFSCS025 and Aspergillus versicolor SCSIO 41502, and these compounds showed anti-larval activity against B. neritina with respective EC50 values of 0.004, 0.006, and 3.40 μg/mL, but their toxicity was very low with all LC50/EC50 ratios >100 [55,56].
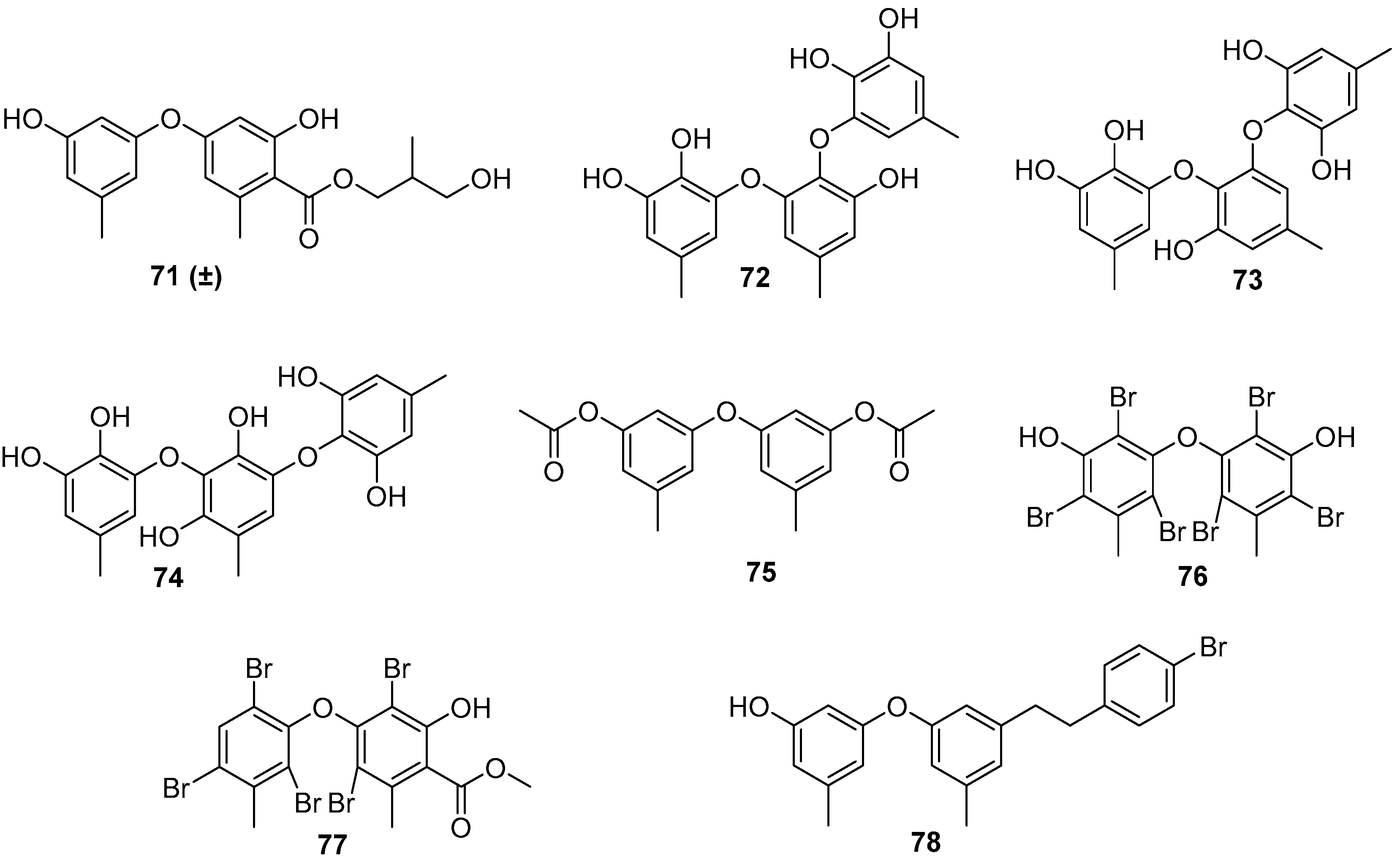
As for the marine fungi-sourced natural products, it should be noted that the fungal genus Aspergillus may be a potent source for the exploitation of AF phenyl ethers [56,57]. Five phenyl ether compounds 71–73 (Figure 13) from the deep-sea-derived fungus A. versicolor SCSIO 41502 were found to have anti-larval activity against settlement of B. neritina larvae with respective EC50 values of 1.28, 2.61, 5.48, 1.59, and 3.40 μg/mL, and LC50/EC50 ratios >100 [56]. Another six phenyl ethers, as characteristic secondary metabolites of Aspergillus sp. XS-20090066, together with their seven synthetic derivatives were demonstrated to have moderate to strong anti-larval activity against settlement of B. amphitrite larvae with EC50 values ≤ 14.11 μg/mL [57]. Among them, compounds 75–78 (Figure 13)were particularly promising, as they showed EC50 values ≤ 0.96 μg/mL and LC50/EC50 ratios > 20. Further studies of SAR indicated that the presence of the ester group and the introduction of bromine atoms at C-4 may be beneficial for enhancing anti-larval activity of these phenyl ethers.
4.3. Polyketides
In recent twenty years, polyketides, as one of the largest families of natural products from marine microorganisms, have provided many anti-biofilm and AF compounds with structural diversity, including benzylazaphilones, dihydroisocoumarins, anthraquinones, xanthones, and 14-membered resorcylic acid lactones (RALs) (Figure 14, Figure 15 and Figure 16). These compounds were mainly isolated from five fungal genera, including Aspergillus, Xylariaceae, Penicillium, Cochliobolus, and Eurotium.
4.3.1. Benzylazaphilones and Dihydroisocoumarins
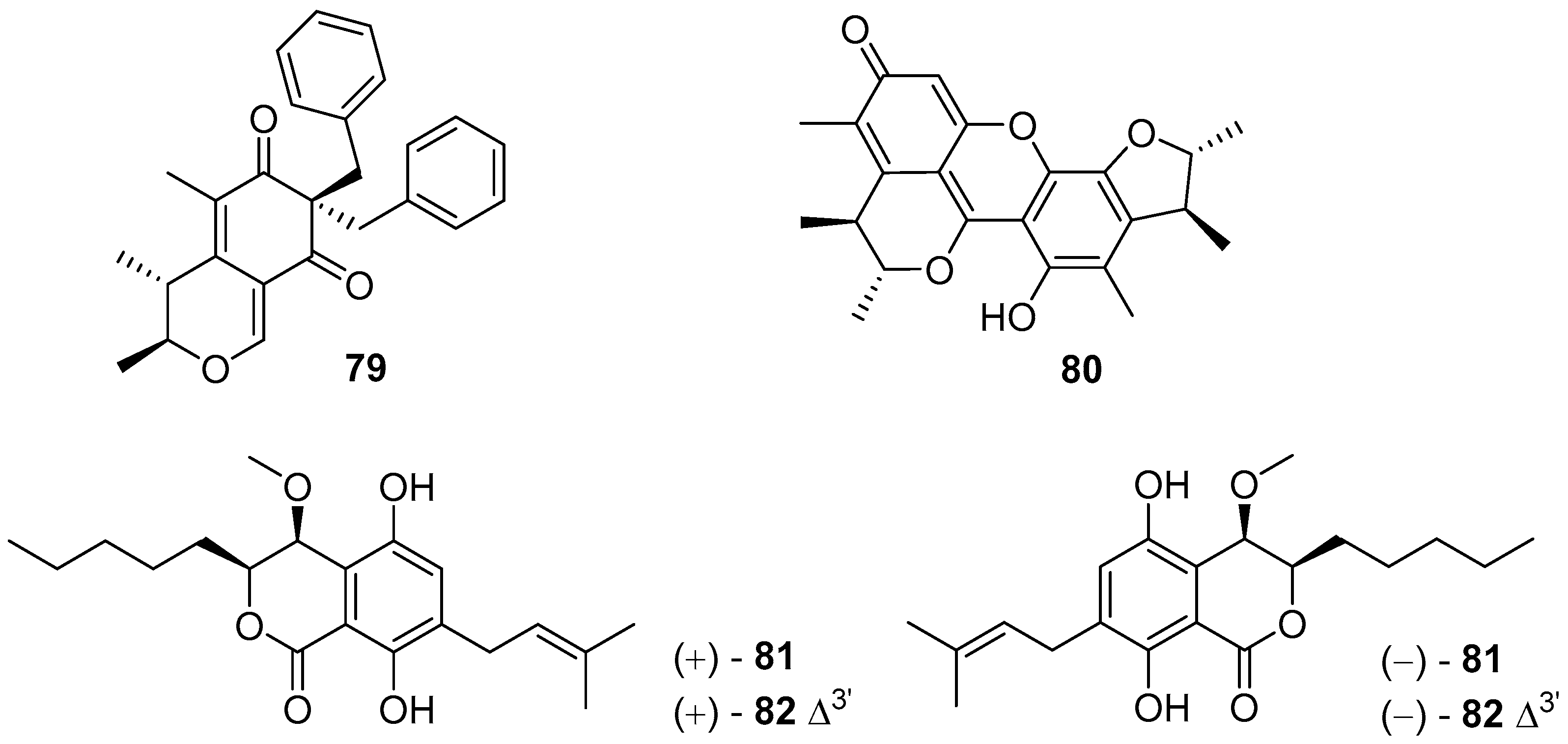
Aspergilone A (79) (Figure 14), a novel benzylazaphilone analogue from a gorgonian-associated fungus Aspergillus sp., could inhibit larval settlement of B. amphitrite with EC50 value of 7.68 μg/mL [58]. Dicitrinin A (80) (Figure 14), isolated from Xylariaceae sp., exhibited highly promising and non-toxic anti-larval activity against B. neritina larvae, with EC50 value of 1.76 μg/mL and a high LC50/EC50 ratio > 56 [59]. Four pairs of rare dihydroisocoumarin metabolites (±)-eurotiumides B (±81) and (±)-eurotiumides D (±82) (Figure 14) from a gorgonian-derived fungus Eurotium sp. XS-200900E6 were evaluated for AF activity against settlement of B. amphitrite larvae, and were all active in inhibiting larval settlement with EC50 values of 1.5, 0.7, 2.3, and 1.9 μg/mL, respectively. Importantly, their toxicities were low with LC50/EC50 ratios > 20 [60].
4.3.2. Anthraquinones and Xanthones
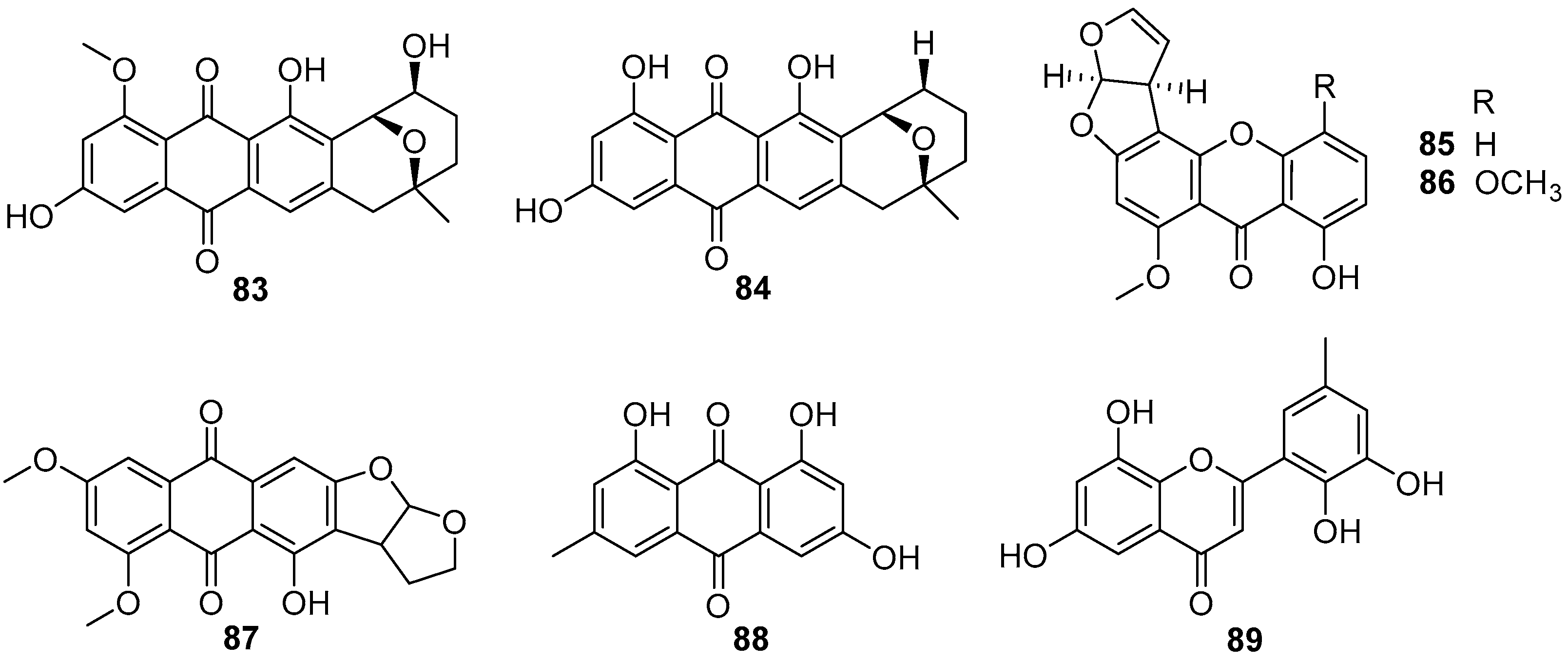
Although anthraquinones and xanthonesthis as prominent secondary metabolites are isolated from cultural broth of some fungal strains, herein only eight compounds from two fungal genera of Penicillium and Aspergillus were investigated for their inhibitory effects on larval settlement of B. amphitrite [57,61,62,63]. Two Aspergillus-sourced anthraquinones averufin (83) and 8-O-methylnidurufin (84) (Figure 15) showed anti-larval activity against B. amphitrite with EC50 values of 2.03 and 3.39 μg/mL, respectively, but their toxicity was high with LC50/EC50 ratios >8.75 [57]. Other screenings of AF natural products from marine fungi yielded several anthraquinones and xanthones with strong to moderate inhibition abilities for settlement of B. amphitrite larvae, among which sterigmatocystin (85) and methoxysterigmatocystin (86) (Figure 15), isolated from Aspergillus strains, were the most highly active with EC50 values <0.125 μg/mL, but paralyzed the cyprid larvae at these effective concentrations [61]. Also, the xanthone 6,8-di-O-methyl versiconol (87) (Figure 15) from an unidentified mangrove endophytic fungus ZSUH-36, and two anthraquinones 88 and 89 (Figure 15) from a gorgonian-derived fungus Penicillium sp. SCSGAF 0023 showed anti-larval activity against B. amphitrite with respective EC50 values of 5.13, 6.10, and 6.70 μg/mL [62,63].
4.3.3. 14-Membered Resorcylic Acid Lactones
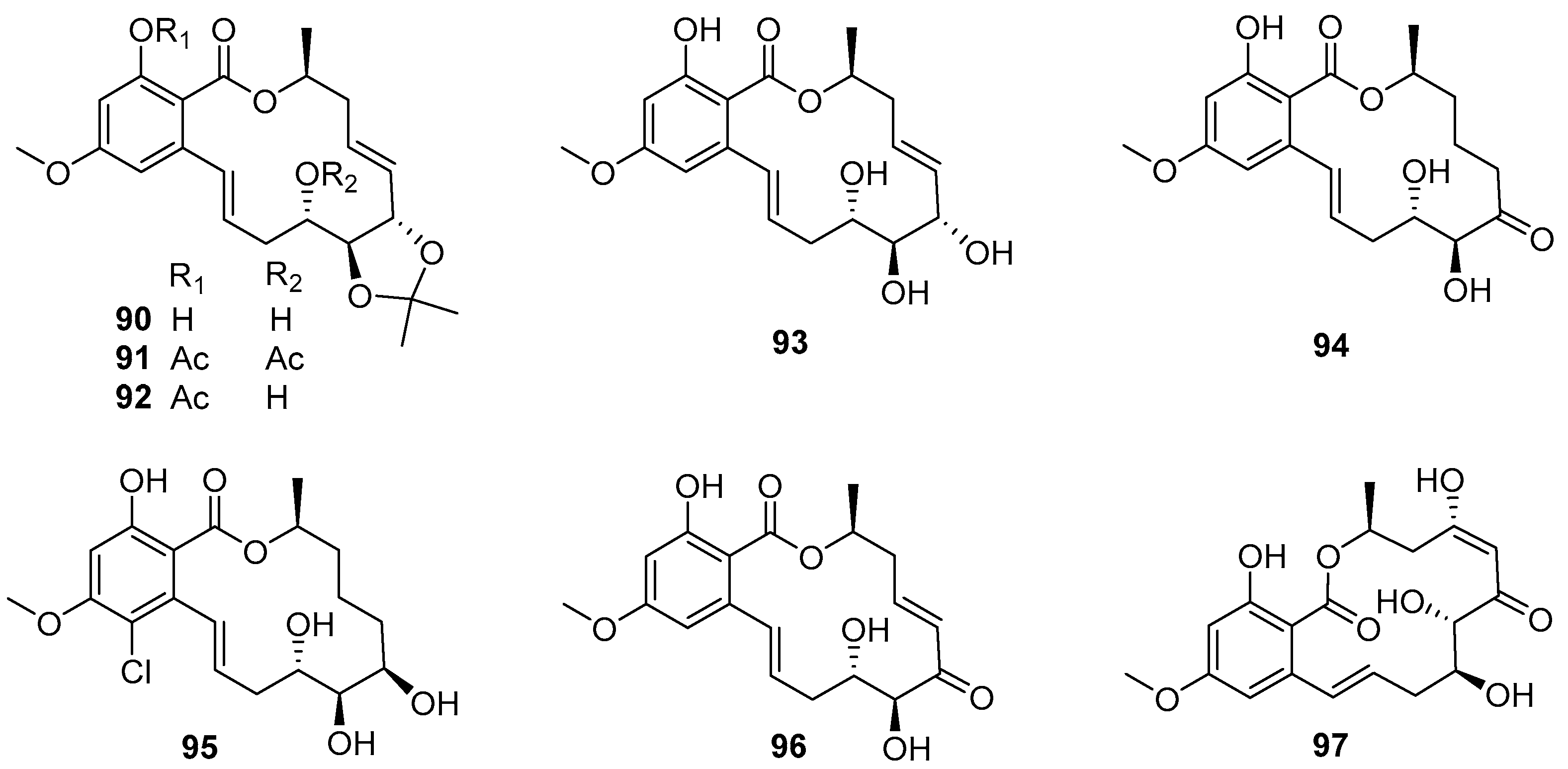
Cochliomycins 90–95 (Figure 16), belonging to 14-membered resorcylic acid lactones (RALs) isolated from a gorgonian-derived fungus Cochliobolus lunatus HQ215514, were first demonstrated to have AF activity against larval settlement of B. amphitrite with EC50 values ranging from 1.2 to 17.9 μg/mL by Shao et al. [64]. Among these compounds, the most active one was cochliomycin A (90), with an EC50 value of 1.2 μg/mL, while its toxicity was low with LC50/EC50 ratio > 16.7. Proteomic analysis of B. amphitrite cyprids treated with cochliomycin A revealed the up- and down-regulation of proteins involved in the NO/cGMP pathway [65]. Later, some other similarly structural cochliomycins were obtained from a sea anemone-derived fungus C. lunatus TA26-46, and showed AF activity against B. amphitrite larvae with EC50 values ranging from 1.82 to 22.5 μg/mL, and the most active LL-Z1640-2 (96) (Figure 16) could inhibit larval settlement of B. amphitrite with EC50 value of 1.82 μg/mL and LC50/EC50 ratio > 50 [66]. As part of the ongoing research toward the AF cochliomycins from fungal strains of C. lunatus, another new compound cochliomycin G (97) (Figure 16) was found to have potent anti-algal activity against Chlorella vulgaris, Chaetoceros socialis, and Navicula exigua with EC50 values of 1.09, 0.92, and 0.61 μg/mL, respectively [67]. In addition, the results of SAR analysis suggested that the acetonide moiety, cis-enone moiety, and hydroxy groups should be essential for improving AF activity of these cochliomycins.
4.4. Alkaloids
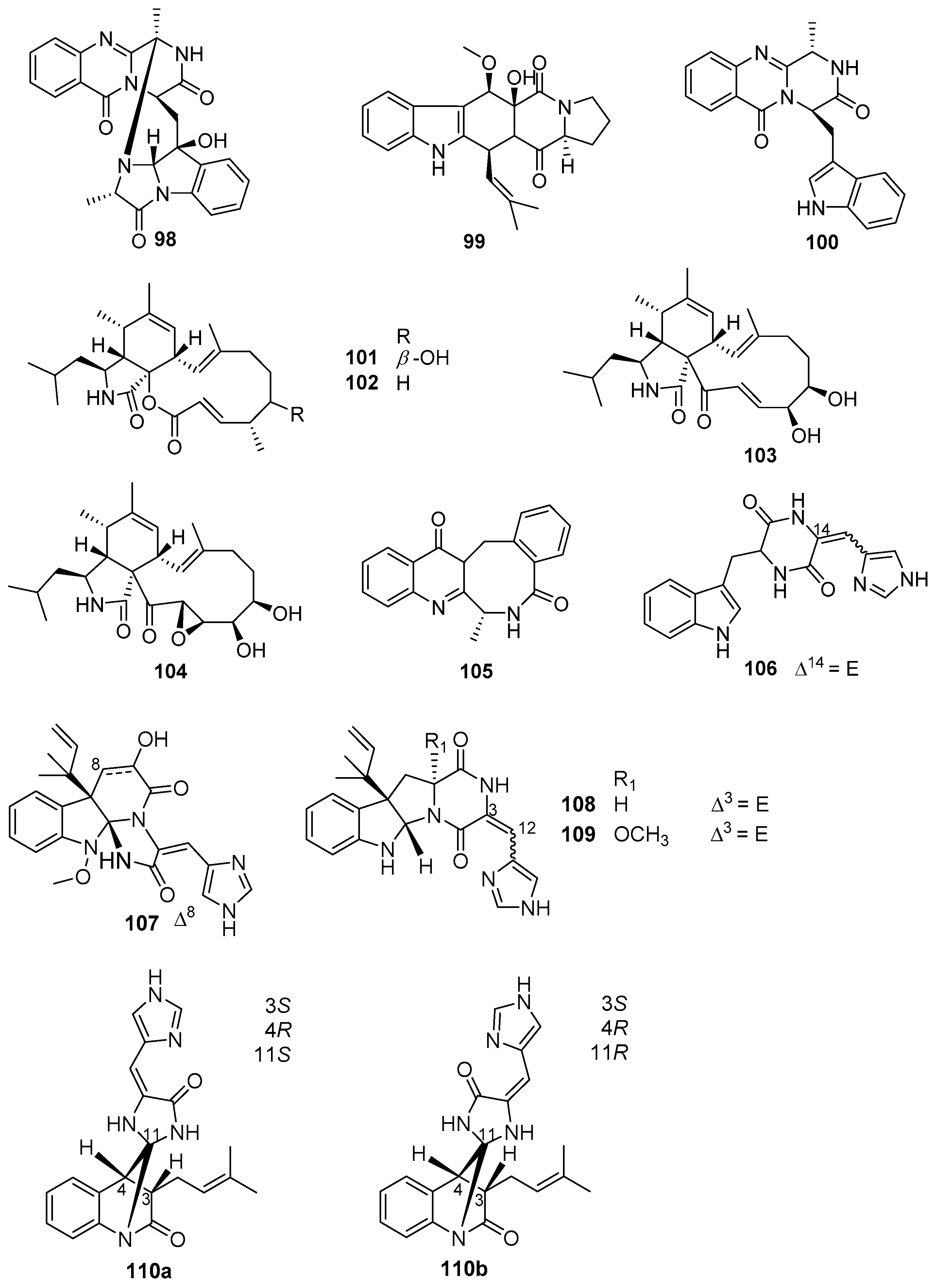
Alkaloids, as a predominant member of marine microbes-derived AF natural products, were isolated from various species of marine fungi that are affiliated with the genera Aspergillus, Penicillium, Scopulariopsis, and Eurotium. Among these fungal strains, Aspergillus and Penicillium have been mostly well-studied [68,69,70,71]. Eight alkaloids 98–105 (Figure 17) produced by some strains belonging to the genus Aspergillus exhibited AF activity against larval attachment of B. amphitrite and B. neritina [68], of which compounds 98–100 from A. sydowii SCSIO 00305 displayed moderate anti-larval activity against settlement of B. neritina larvae with EC50 values ranging from 8.2 to 15.3 μg/mL. A series of cytochalasin alkaloids were obtained from a soft coral Sarcophyton sp.-derived fungus A. elegans ZJ-2008010, and evaluated for AF activity toward B. amphitrite larvae [69]. Among them, cytochalasins 101–104 showed anti-larval activity with EC50 values of 14.13, 5.59, 2.49, and 15.45 μg/mL, respectively. Analysis of SAR revealed that the electrophilic α,β-unsaturated ketone moiety and the double-bond at C-19 and C-20 in the most active aspochalasin D (103) might play an important role in increasing anti-larval activity of these cytochalasins. The benzodiazepine alkaloid (+)-circumdatin F (105) from A. westerdijkiae SCSIO 05233 was reported as an inhibitor for settlement of B. amphitrite larvae with EC50 value of 8.81 μg/mL [69]. Meanwhile, two strains SCSIO 00258 and OUCMDZ-776 of Penicillium were also found to produce some anti-larval secondary metabolites of alkaloids [68,71]. Among them, four indole alkaloids 106–109 (Figure 17) isolated from SCSIO 00258 showed anti-larval activity against B. amphitrite larvae with EC50 values ranging from 1.1 to 17.5 μg/mL, and the meleagrin (107) was considered as a potent AF candidate of ecofriendly antifoulants with a low EC50 value of 1.1 μg/mL and a high LC50/EC50 ratio > 22.6 [68]. In addition, a novel AF alkaloid penispirolloid A (110a/b) (Figure 17) possessing a unique spiro imidazolidinyl skeleton was isolated from a halotolerant fungus Penicillium sp. OUCMDZ-776, and also could prevent larval settlement of B. neritina with EC50 value of 2.40 μg/mL [71].
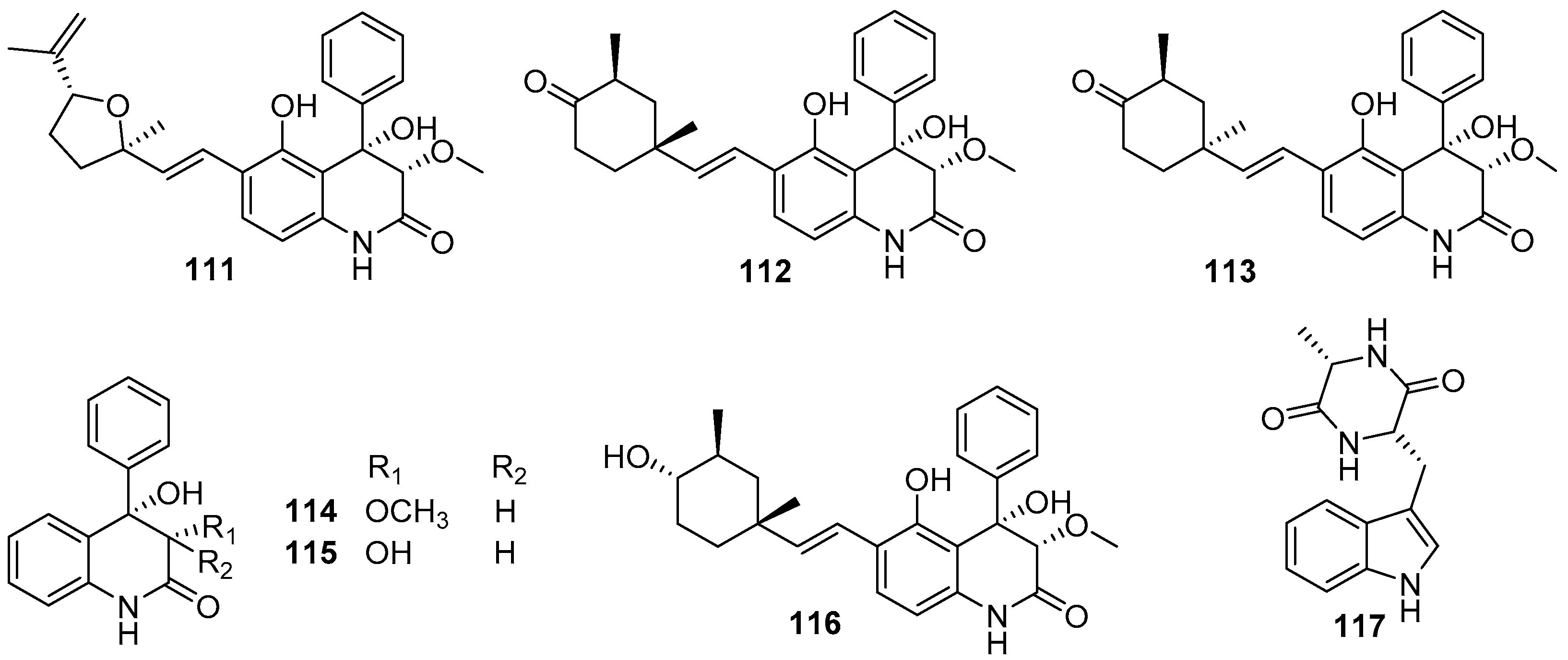
Besides those above-mentioned anti-larval alkaloids from Aspergillus and Penicillium strains, some other types of AF alkaloids were also obtained from various marine fungi. Several dihydroquinolin-2-one alkaloids were isolated from a gorgonian-derived fungus Scopulariopsis sp., all of which except aflaquinolone G displayed highly effective inhibition for larval settlement of B. amphitrite, with EC50 values of compounds 111–115 (Figure 18) being 0.007 pg/mL, 0.012 ng/mL, 0.001 ng/mL, 0.280 μg/mL, and 0.219 μg/mL, respectively [72]. Perhaps more notably, these active compounds were non-toxic with LC50/EC50 ratios ranging from 57 to 1200. Further chemical investigation of the fungus Scopulariopsis sp. led to the discovery of another new dihydroquinolin-2(1H)-one with a monoterpenoid side chain, scopuquinolone B (116) (Figure 18) [73]. This compound also showed outstanding anti-larval activity against the settlement of B. amphitrite larvae with EC50 value of 0.045 μg/mL and LC50/EC50 ratio of 222. Recently, the alkaloid cyclo-L-Trp-L-Ala (117) (Figure 18) from the sponge-associated fungus Eurotium chevalieri MUT 2316 was subjected to an enzymatic assay based on tyrosinase, which plays an important role in mussel byssus production. This compound displayed strong inhibition of tyrosinase with an LOEC value of 0.01 μg/mL, suggesting its potent anti-larval activity toward mussels [74].
4.5. Amines
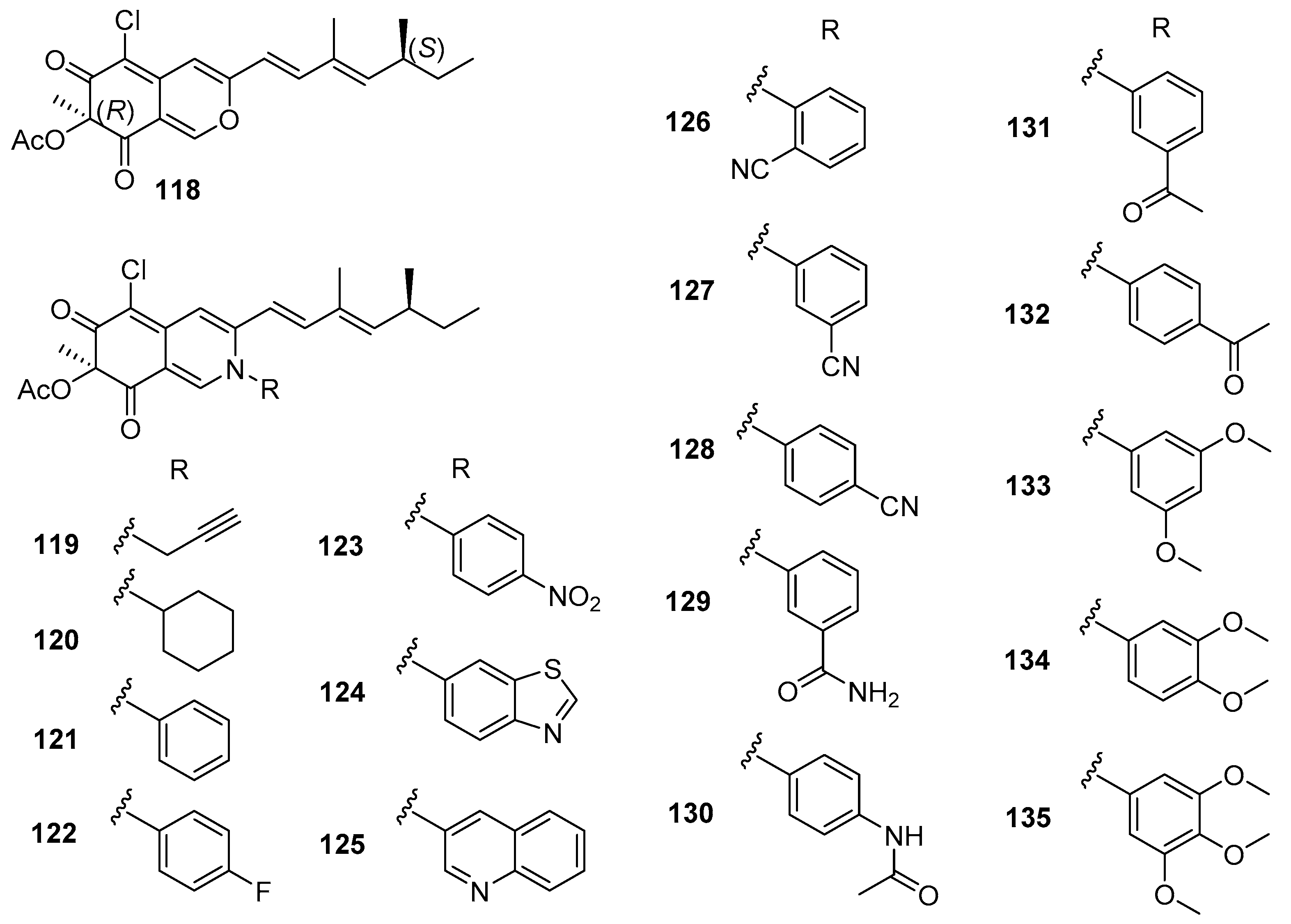
The natural compound (+)-sclerotiorin (118) (Figure 19), isolated from a gorgonian coral-derived fungus Penicillium sclerotiorum CHNSCLM-0013, was reported as an AF compound for larval settlement of B. amphitrite with EC50 value of 5.6 μg/mL and LC50/EC50 ratio > 8.9 by Wei et al. [75]. In order to explore environmentally friendly AF amine derivatives, a series of 30 sclerotioramine derivatives were first synthesized, and evaluated for anti-larval activity against B. amphitrite larvae. A total of 25 semisynthetic derivatives showed anti-larval activity with EC50 values ranging from 0.47–18.2 μg/mL and LC50/EC50 ratios >2.7. Among these synthetic AF sclerotioramines, compounds 119–135 (Figure 19) showed potent anti-larval activities, which were stronger than that of the natural product (+)-sclerotiorin. The most active ones were the aliphatic amino-derivative 119 and the aromatic amino-derivative 134 with EC50 values of 0.94 and 0.47 μg/mL and LC50/EC50 ratios >53.2, indicating that both of them could be further developed as AF candidates as antifoulants. More interestingly, it should be mentioned that most of the potential anti-larval sclerotioramines belong to the aromatic amines except for compounds 119 and 120, which suggest that the substitution of aromatic groups at N-2 should be crucial for increasing AF activities of these derivatives.
4.6. Peptides and Nucleosides
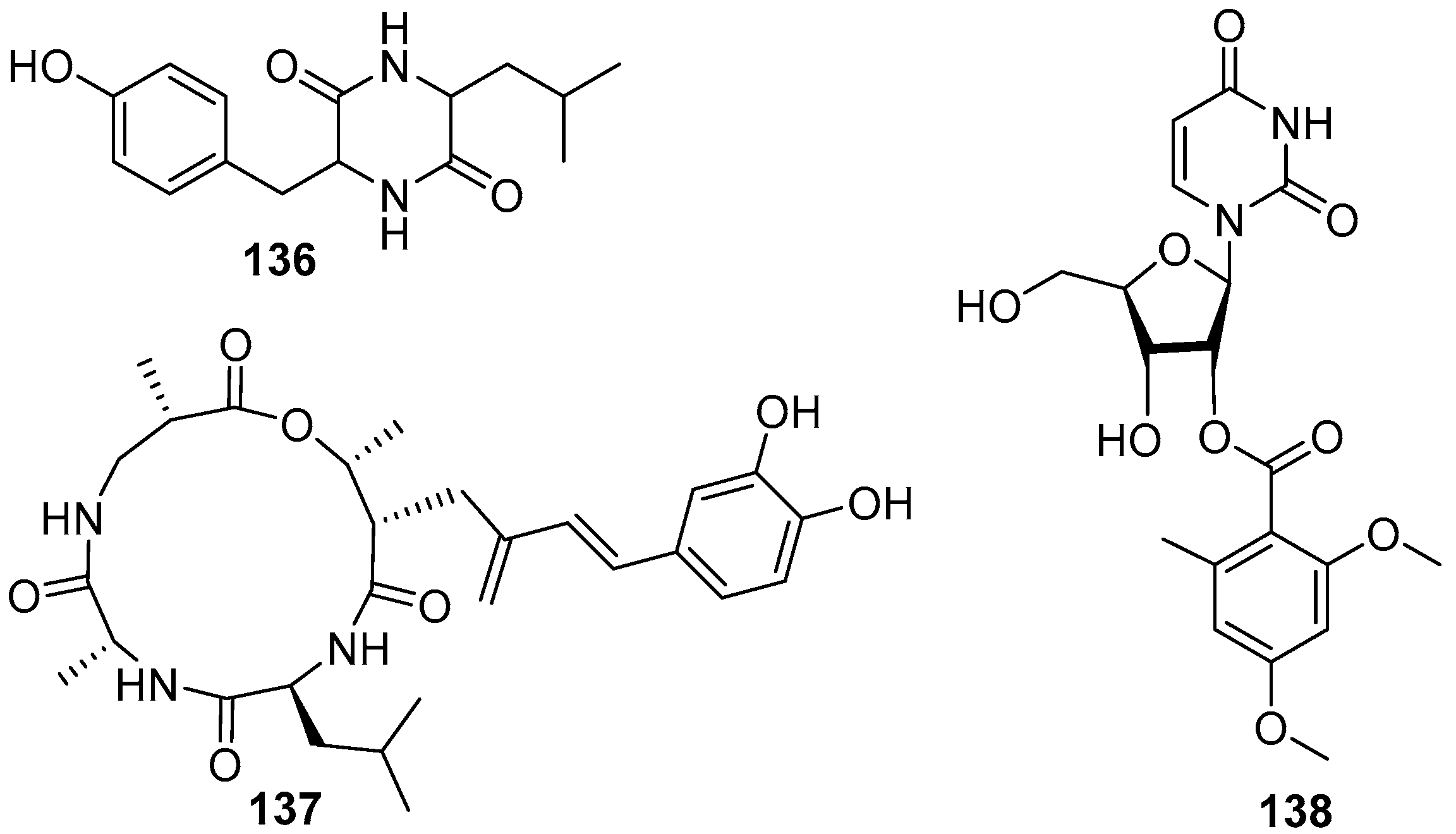
A dipeptide compound cis-cyclo (Leucyl-Tyrosyl) (136) (Figure 20) was isolated from a marine fungus Penicillium sp. F37, and found to be capable of inhibiting ~80% biofilm formation of a pathogenic bacterium Staphylococcus epidermidis at the concentration of 1.0 mg/mL, while the growth of the target strain was not interfered [76]. At the same time, a cyclic tetrapeptide aspergillipeptide C (137) (Figure 20), isolated from a fungus Aspergillus sp. SCSGAF 0076 associated with a gorgonian Melitodes squamata, displayed anti-larval activity against B. neritina with EC50 value of 11 μg/mL and LC50/EC50 ratio > 25 [77]. Only one AF nucleoside compound diacetylkipukasin E (138) (Figure 20) was obtained from marine microorganisms. This compound was isolated from a gorgonian Dichotella gemmacea-derived fungus Aspergillus versicolor, and displayed weak inhibitory effect on settlement of B. amphitrite larvae with EC50 value of 22.5 μg/mL [78].
5. Conclusions and Looking Ahead
In the last two decades, few studies on anti-biofilm natural products from marine microorganisms have been reported, except for the twelve compounds belonging to the five chemical types described in this review. Among these anti-biofilm natural products, the most effective ones are butenolide (7) and terpenoid compounds napyradiomycins 18–20 and 25, with the exception of ophiobolin K (61). They are all considered highly promising AF compounds for further development of antifoulant candidates. As mentioned at the beginning, biofilms play essential roles in biofouling of macro- and micro-foulers because of inducing the settlement of invertebrates’ larvae and algae spores, which indicates that some effective anti-larval/anti-algal compounds may possess a promising potential for anti-biofilm agents. Herein, a total of 112 important anti-larval and anti-algal natural products, together with their 26 synthetic analogues, are shown from Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13, Figure 14, Figure 15, Figure 16, Figure 17, Figure 18, Figure 19 and Figure 20. According to Qian et al. [17] and Fusetani et al. [18], a compound with an EC50 value of <5 μg/mL and a LC50/EC50 ratio of >50 can be considered a highly promising candidate for environmentally benign AF agents, but those with lower AF concentrations and higher therapeutic ratios are more desirable. As such, some significantly efficient and non-toxic anti-larval/anti-algal compounds from marine microbes, including the following: Streptomyces-derived butenolides and terpenoids, Pseudovibrio-derived bisindole alkaloids, Pseudoalteromonas-derived protease, cyanobacteria-derived polyketide–polypeptide structural compounds, fungi-derived phenols and phenyl ethers, Xylariaceae-derived benzylazaphilones and Cochliobolus lunatus-derived 14-Membered RALs belonging to the polyketides, Scopulariopsis-derived alkaloids, and semisynthetic sclerotioramines based on Eurotium chevalieri-derived amine, are worthy of exploitation for their potential as anti-biofilm and AF candidates. Undoubtedly, more and more AF natural products from marine microorganisms will be discovered in the future due to their great AF applications in the medical and marine industries. It is a feasible and time-saving way to get a certain amount of anti-biofilm agents by screening for anti-biofilm activity of marine microbes-derived AF natural products.
The main challenges for the development of anti-biofilm/AF compounds have been highlighted in previous reviews [2,7,8,12,13,14,15,16,17,18,19,20]. As we have been advancing our knowledge of how to bioprospect for potent anti-biofilm/AF from marine microorganisms, a few of the issues remaining in current research or application studies should be addressed in future research. First, although the so-called “continuous supply” is one of the major factors that warrants microbe-derived anti-biofilm/AF agents for biological assays and preclinical/clinical/field trials, a few promising and effective natural products have complex structures and low fermentation yields, which hinders their further development. To solve the bottleneck problem of supply, genetic engineering technology can be employed for the culture and fermentation of microbes to increase the yield of target substances, and synthetic/semi-synthetic strategies could also be developed to satisfy the demand for large quantities of compounds for application in commercial or medicinal areas. Second, for those potent anti-biofilm/AF natural products based on the evaluation through biofilm assays/antifouling activity evaluation platforms in laboratories, further in vivo outcomes/field experiments should be performed since the results could be very different. Third, the basic mechanistic aspects of how anti-biofilm/AF compounds exert their activity should be well understood. Over the past twenty years, a number of genes and proteins that may be involved in biofilm formation or fouling of invertebrate larvae and algae have been identified through genomic and proteomic approaches [22,27,29,65,79,80,81,82]. In the near future, more efforts need to be invested to decipher the specific signaling pathways that are either conserved, or unique to the biofilm formation/eradication, and settlement/anti-settlement processes, which will facilitate the introduction of anti-biofilm/AF to the market. Fourth, comprehensive toxicity tests need to be established. For anti-biofilm agents, before advancing to costly animal models, simple nematode models can be used for the preliminary toxicological analysis [82,83,84,85,86,87,88,89]. Caenorhabditis elegans, as a tiny model organism, is often used in toxicity studies of small molecules owing to its complete genome, short lifespan, and simple physiology [84,90,91]. For antifouling compounds, their acute toxicity to larvae of model macro-foulers, chronic toxicity to some model organisms (such as zebrafish), as well as their degradation kinetics of in the marine ecosystem need to be evaluated before commercialization. Finally, it has been demonstrated that bacterial quorum sensing (QS) plays an important role in biofilm formation and infectious processes [2,14,15]. Hence, development of efficient and safe anti-biofilm agents via interfering with QS to inhibit biofilm formation and reduce virulence without inhibiting basic growth of bacteria may be a promising solution to treat bacterial infections.
Last but not least, in addition to purified compounds, dozens of crude extracts from marine microbes were also found to have good anti-biofilm/AF activity in the past decades, which provided potential sources for further exploitation as novel anti-biofilm/AF compounds. For example, the crude extract of an actinobacterial strain Glycomyces sediminimaris UTMC 2460 caused 93.2% and 71.4% reduction in biofilm biomass of two fouling bacterial strains Kocuria sp. UTMC 2449 and Mesorhizobium sp. UTMC 2518 with MIC value of 100 µg/mL, respectively, and it was not toxic toward Artemia salina larvae [92]. According to Viju et al. [93], the coating developed with 15.8 and 27.5% (w/w) crude extract of the bacterium Pseudomonas putida showed a significant reduction in the recruitment of fouling organisms over a period of 50 days under natural marine conditions. For these promising and effective crude extracts of marine bacteria mentioned above, it is necessary to carry out further investigation of separation, purification, and chemical identification of the active antibiofilm/AF compounds present in these extracts.
Overall, it is well recognized that AF natural products from marine microorganisms could provide a promising source for anti-biofilm compounds. Nowadays, there are still many obstacles restricting the development of anti-biofilm/AF agents in medical and industrial applications. To meet these major issues as mentioned in this review, collaborative endeavors involving natural products chemistry with organic chemistry, microbiology, pharmacology, and biology will help to build up a fairly systematic research system, and facilitate an increase in marine microorganisms-derived natural products reaching the clinical trials/market as anti-biofilm therapeutics/antifoulants.
Author Contributions
Data curation, K.-L.W., Z.-R.D., G.-F.G. and H.-F.L.; writing—original draft preparation, K.-L.W., Z.-R.D. and G.-F.G.; writing—review and editing, K.-L.W., Z.-R.D., H.-F.L., B.J. and Y.X.; supervision, B.J. and Y.X.; funding acquisition, K.-L.W., Y.X. and B.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Nos. 32060032 and 41706155), Shenzhen Science and Technology Program (JCYJ20210324093409025), the Natural Science Foundation of Yunnan Province of China (No. 202001AT070022), the Scientific Research Foundation of Dali University (KYBS2021099), and the Development and Utilization of Characteristic Medicinal Plants in Western Yunnan & Bai Nationality Medicines (No. ZKLX2019106).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Stowe, S.D.; Richards, J.J.; Tucker, A.T.; Thompson, R.; Melander, C.; Cavanagh, J. Anti-biofilm compounds derived from marine sponges. Mar. Drugs 2011, 9, 2010–2035. [Google Scholar] [CrossRef] [Green Version]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology—past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coat. 2004, 50, 75–104. [Google Scholar] [CrossRef]
- Wahl, M. Marine epibiosis. I. Fouling and antifouling: Some basic aspects. Mar. Ecol. Prog. Ser. 1989, 58, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Dahms, H.U.; Qian, P.-Y. Inhibition of biofouling by marine microorganisms and their metabolites. Biofouling 2006, 22, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.Y.; Lau, S.; Dahms, H.U.; Dobretsov, S.; Harder, T. Marine biofilms as mediators of colonization by marine macroorganisms: Implications for antifouling and aquaculture. Mar. Biotechnol. 2007, 9, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Abed, R.M.M.; Teplitski, M. Mini-review: Inhibition of biofouling by marine microorganisms. Biofouling 2013, 29, 423–441. [Google Scholar] [CrossRef]
- Satheesh, S.; Ba-akdah, M.A.; Al-Sofyani, A.A. Natural antifouling compound production by microbes associated with marine macroorganisms—A review. Electron. J. Biotechnol. 2016, 21, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Teplitski, M.; Alagely, A.; Gunasekera, S.P.; Paul, V.J. Malyngolide from the cyanobacterium Lyngbya majuscula interferes with quorum sensing circuitry. Environ. Microbiol. Rep. 2010, 2, 739–744. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Issues in Healthcare Settings: CDC’s Seven Healthcare Safety Challenges; United States Department of Health and Human Services: Atlanta, GA, USA, 2001.
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural products as inspiration for the development of bacterial antibiofilm agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control biofilm-forming pathogens. Front. Microbiol. 2020, 11, 2640. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Biofouling and antifouling. Nat. Prod. Rep. 2004, 21, 94–104. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Xu, Y.; Fusetani, N. Natural products as antifouling compounds: Recent progress and future perspectives. Biofouling 2009, 26, 223–234. [Google Scholar] [CrossRef]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep. 2011, 28, 400–410. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Li, Z.; Xu, Y.; Li, Y.; Fusetani, N. Mini-review: Marine natural products and their synthetic analogs as antifouling compounds: 2009–2014. Biofouling 2015, 31, 101–122. [Google Scholar] [CrossRef]
- Wang, K.-L.; Wu, Z.-H.; Wang, Y.; Wang, C.-Y.; Xu, Y. Mini-Review: Antifouling natural products from marine microorganisms and their synthetic analogs. Mar. Drugs 2017, 15, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, H.D.; Ganti, V.S.; Paudel, B.; Lee, Y.K.; Lee, H.K.; Hong, Y.K.; Shin, H.W. Isolation of antifouling compounds from the marine bacterium, Shewanella oneidensis SCH0402. World J. Microbiol. Biotechnol. 2007, 23, 243–249. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Li, X.; Xiao, X.; Qian, P.-Y. Inhibitory effects of a branched-chain fatty acid on larval settlement of the polychaete Hydroides elegans. Mar. Biotechnol. 2009, 11, 495–504. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Martens, T.; Brinkhoff, T.; Simon, M.; Schulz, S. Volatiles released by a Streptomyces species isolated from the North Sea. Chem. Biodivers. 2005, 2, 837–865. [Google Scholar] [CrossRef]
- Xu, Y.; He, H.; Schulz, S.; Liu, X.; Fusetani, N.; Xiong, H.; Xiao, X.; Qian, P.-Y. Potent antifouling compounds produced by marine Streptomyces. Bioresour. Technol. 2010, 101, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.Y.; Cheng, J.; Liu, X.; Ma, C.; Qian, P.Y. Synthetic analogue of butenolide as an antifouling agent. Mar. Drugs 2021, 19, 481. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, F.; Xu, Y.; Matsumura, K.; Han, Z.; Liu, L.; Lin, W.; Jia, Y.; Qian, P.-Y. Structural optimization and evaluation of butenolides as potent antifouling agents: Modification of the side chain affects the biological activities of compounds. Biofouling 2012, 28, 857–864. [Google Scholar] [CrossRef]
- Hong, Y.K.; Cho, J.Y. Effect of seaweed epibiotic bacterium Streptomyces violaceoruber SCH-09 on marine fouling organisms. Fish. Sci. 2013, 79, 469–475. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Y.; Arellano, S.M.; Xiao, K.; Qian, P.-Y. Comparative proteome and phosphoproteome analyses during cyprid development of the barnacle Balanus (=Amphibalanus) amphitrite. J. Proteome Res. 2010, 9, 3146–3157. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Wong, Y.H.; Zhang, Y. Changes in the proteome and phosphoproteome expression in the bryozoan Bugula neritina larvae in response to the antifouling agent butenolide. Proteomics 2010, 10, 3435–3446. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Ma, C.; Zhang, W.; Chiang, H.; Qian, P.-Y. Anti-biofilm effect of a butenolide polymer coating and metatranscriptomic analyses. Biofouling 2018, 34, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.; Liang, J.; Zhang, W.; Zhang, L.; Hu, Z.L.; Zhang, Y.; Ying, X. Butenolide, a marine-derived broad-spectrum antibiofilm agent against both gram-positive and gram-negative pathogenic bacteria. Mar. Biotechnol. 2019, 21, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.Y.; Kim, M.S. Induction of antifouling diterpene production by Streptomyces cinnabarinus PK209 in co-culture with marine-derived Alteromonas sp. KNS-16. Biosci. Biotechnol. Biochem. 2012, 76, 1849–1854. [Google Scholar] [CrossRef] [Green Version]
- Bauermeister, A.; Pereira, F.; Grilo, I.R.; Godinho, C.C.; Paulino, M.; Almeida, V.; Gobbo-Neto, L.; Prieto-Davo, A.; Sobral, R.G.; Lopes, N.P.; et al. Intra-clade metabolomic profiling of MAR4 Streptomyces from the Macaronesia Atlantic region reveals a source of anti-biofilm metabolites. Environ. Microbiol. 2019, 21, 1099–1112. [Google Scholar] [CrossRef]
- Pereira, F.; Almeida, J.R.; Paulino, M.; Grilo, I.R.; Macedo, H.; Cunha, I.; Sobral, R.G.; Vasconcelos, V.; Gaudêncio, S.P. Antifouling napyradiomycins from marine-derived actinomycetes Streptomyces aculeolatus. Mar. Drugs 2020, 18, 63. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.Y. Antifouling steroids isolated from red alga epiphyte filamentous bacterium Leucothrix mucor. Fish. Sci. 2012, 78, 683–689. [Google Scholar] [CrossRef]
- Song, Z.-M.; Zhang, J.-L.; Zhou, K.; Yue, L.-M.; Zhang, Y.; Wang, C.-Y.; Wang, K.-L.; Xu, Y. Anthraquinones as potential antibiofilm agents against Methicillin-resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 709826. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-L.; Xu, Y.; Lu, L.; Li, Y.; Han, Z.; Zhang, J.; Shao, C.-L.; Wang, C.-Y.; Qian, P.-Y. Low-toxicity diindol-3-ylmethanes as potent antifouling compounds. Mar. Biotechnol. 2015, 17, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kang, J.Y.; Hong, Y.K.; Baek, H.H.; Shin, H.W.; Kim, M.S. Isolation and structural determination of the antifouling diketopiperazines from marine-derived Streptomyces praecox 291-11. Biosci. Biotechnol. Biochem. 2012, 76, 1116–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-L.; Liang, X.-Y.; Zhang, S.-L.; Song, Z.-M.; Wang, C.-Y.; Xu, Y. Maipomycin A, a novel natural compound with promising anti-biofilm activity against Gram-negative pathogenic bacteria. Front. Microbiol. 2021, 11, 3480. [Google Scholar] [CrossRef] [PubMed]
- Dusane, D.H.; Pawar, V.S.; Nancharaiah, Y.V.; Venugopalan, V.P.; Kumar, A.R.; Zinjarde, S.S. Anti-biofilm potential of a glycolipid surfactant produced by a tropical marine strain of Serratia marcescens. Biofouling 2011, 27, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Nithya, C.; Devi, M.G.; Pandian, S.K. A novel compound from the marine bacterium Bacillus pumilus S6-15 inhibits biofilm formation in gram-positive and gram-negative species. Biofouling 2011, 27, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Gopikrishnan, V.; Radhakrishnan, M.; Shanmugasundaram, T.; Gopikrishnan, V.; Radhakrishnan, M.; Shanmugasundaram, T.; Ramakodi, M.P.; Balagurunathan, R. Isolation, characterization and identification of antibiofouling metabolite from mangrove derived Streptomyces sampsonii PM33. Sci. Rep. 2019, 9, 12975. [Google Scholar] [CrossRef]
- Long, L.; Wang, R.; Chiang, H.Y.; Ding, W.; Li, Y.X.; Chen, F.; Qian, P.Y. Discovery of antibiofilm activity of elasnin against marine biofilms and its application in the marine antifouling coatings. Mar. Drugs 2021, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Xiong, H.; Xu, Y.; Levin, L.A.; Qian, P.-Y. Novel antifoulants: Inhibition of larval attachment by proteases. Mar. Biotechnol. 2007, 9, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Wong, H.F.; Hutchinson, N.; Lee, S.C.; Chan, B.K.K.; Williams, G.A. Chemistry and biology of maculalactone A from the marine cyanobacterium Kyrtuthrix maculans. Phytochem. Rev. 2004, 3, 381–400. [Google Scholar] [CrossRef]
- Bader, S.L.; Luescher, M.U.; Gademann, K. Synthesis of maculalactone A and derivatives for environmental fate tracking studies. Org. Biomol. Chem. 2015, 13, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.T.; Goh, B.P.L.; Tripathi, A.; Lim, M.G.; Dickinson, G.H.; Lee, S.S.C.; Teo, S.L.M. Natural antifoulants from the marine cyanobacterium Lyngbya majuscula. Biofouling 2010, 26, 685–695. [Google Scholar] [CrossRef]
- Petitbois, J.G.; Casalme, L.O.; Lopez, J.A.V.; Alarif, W.M.; Abdel-Lateff, A.; Al-Lihaibi, S.S.; Yoshimura, E.; Nogata, Y.; Umezawa, T.; Matsuda, F.; et al. Serinolamides and lyngbyabellins from an Okeania sp. cyanobacterium collected from the Red Sea. J. Nat. Prod. 2017, 80, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.; Pereira, S.; Ribeiro, T.; Plowman, J.E.; Thomas, A.; Clerens, S.; Campos, A.; Vasconcelos, V.; Almeida, J.R. A multi-bioassay integrated approach to assess the antifouling potential of the cyanobacterial metabolites portoamides. Mar. Drugs 2019, 17, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casalme, L.O.; Yamauchi, A.; Sato, A.; Petitbois, J.G.; Nogata, Y.; Yoshimura, E.; Okino, T.; Umezawa, T.; Matsuda, F. Total synthesis and biological activity of dolastatin 16. Org. Biomol. Chem. 2017, 15, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, Y.; Shao, C.-L.; Yang, R.-Y.; Zheng, C.-J.; Chen, Y.-Y.; Fu, X.-M.; Qian, P.-Y.; She, Z.-G.; Voogd, N.J.D.; et al. Antibacterial bisabolane-type sesquiterpenoids from the sponge-derived fungus Aspergillus sp. Mar. Drugs 2012, 10, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Arai, M.; Niikawa, H.; Kobayashi, M. Marine-derived fungal sesterterpenes, ophiobolins, inhibit biofilm formation of Mycobacterium species. J. Nat. Med. 2013, 67, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Kwong, T.F.N.; Miao, L.; Li, X.; Qian, P.-Y. Novel antifouling and antimicrobial compound from a marine-derived fungus Ampelomyces sp. Mar. Biotechnol. 2006, 8, 634–640. [Google Scholar] [CrossRef]
- Xing, Q.; Gan, L.-S.; Mou, X.-F.; Wang, W.; Wang, C.-Y.; Wei, M.-Y.; Shao, C.-L. Isolation, resolution and biological evaluation of pestalachlorides E and F containing both point and axial chirality. RSC Adv. 2016, 6, 22653–22658. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Nong, X.; Wang, J.; Qi, S.-H. Brevianamides and mycophenolic acid derivatives from the deep-sea-derived fungus penicillium brevicompactum DFFSCS025. Mar. Drugs 2017, 15, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Nong, X.; Ren, Z.; Zhe, R.; Wang, J.; Zhang, X.; Qi, S.-H. Anti-HSV-1, antioxidant and antifouling phenolic compounds from the deep-sea-derived fungus Aspergillus versicolor SCSIO 41502. Bioorg. Med. Chem. Lett. 2017, 27, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Y.; Wang, K.-L.; Qian, P.-Y.; Xu, Y.; Chen, M.; Zheng, J.-J.; Liu, M.; Shao, C.-L.; Wang, C.-Y. Antifouling phenyl ethers and other compounds from the invertebrates and their symbiotic fungi collected from the South China Sea. AMB Express 2016, 6, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.-L.; Wang, C.-Y.; Wei, M.-Y.; Gu, Y.-C.; She, Z.-G.; Qian, P.-Y.; Lin, Y.-C. Aspergilones A and B, two benzylazaphilones with an unprecedented carbon skeleton from the gorgonian-derived fungus Aspergillus sp. Bioorg. Med. Chem. Lett. 2011, 21, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Nong, X.-H.; Zheng, Z.-H.; Zhang, X.-Y.; Lu, X.-H.; Qi, S.-H. Polyketides from a marine-derived fungus Xylariaceae sp. Mar. Drugs 2013, 11, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Shao, C.-L.; Wang, K.-L.; Xu, Y.; She, Z.-G.; Wang, C.-Y. Dihydroisocoumarin derivatives with antifouling activities from a gorgonian-derived Eurotium sp. fungus. Tetrahedron 2014, 70, 9132–9138. [Google Scholar] [CrossRef]
- Li, Y.X.; Wu, H.X.; Xu, Y.; Shao, C.-L.; Wang, C.-Y.; Qian, P.-Y. Antifouling activity of secondary metabolites isolated from Chinese marine organisms. Mar. Biotechnol. 2013, 15, 552–558. [Google Scholar] [CrossRef]
- Shao, C.-L.; She, Z.-G.; Guo, Z.; Peng, H.; Cai, X.; Zhou, S.; Gu, Y.-C.; Lin, Y.-C. 1H and 13C NMR assignments for two anthraquinones and two xanthones from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2007, 45, 434–438. [Google Scholar] [CrossRef]
- Bao, J.; Sun, Y.-L.; Zhang, X.-Y.; Han, Z.; Gao, H.-C.; He, F.; Qian, P.-Y.; Qi, S.-H. Antifouling and antibacterial polyketides from marine gorgonian coral-associated fungus Penicillium sp. SCSGAF 0023. J. Antibiot. 2013, 66, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.-L.; Wu, H.-X.; Wang, C.-Y.; Liu, Q.-A.; Xu, Y.; Wei, M.-Y.; Qian, P.-Y.; Gu, Y.-C.; Zheng, C.-J.; She, Z.-G.; et al. Potent antifouling resorcylic acid lactones from the gorgonian-derived fungus Cochliobolus lunatus. J. Nat. Prod. 2011, 74, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-L.; Zhang, G.; Sun, J.; Xu, Y.; Han, Z.; Liu, L.-L.; Shao, C.-L.; Liu, Q.-A.; Wang, C.-Y.; Qian, P.-Y. Cochliomycin A inhibits the larval settlement of Amphibalanus amphitrite by activating the NO/cGMP pathway. Biofouling 2016, 32, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.A.; Shao, C.-L.; Gu, Y.-C.; Blum, M.; Gan, L.S.; Wang, K.-L.; Chen, M.; Wang, C.-Y. Antifouling and fungicidal resorcylic acid lactones from the Sea Anemone-derived fungus Cochliobolus lunatus. J. Agric. Food Chem. 2014, 62, 3183–3191. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-F.; Xue, X.-J.; Qi, Y.-X.; Wu, N.-N.; Shao, C.-L. Cochliomycin G, a 14-membered resorcylic acid lactone from a marine-derived fungus Cochliobolus lunatus. Nat. Prod. Res. 2019, 35, 490–493. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Han, Z.; Peng, J.; Qian, P.-Y.; Qi, S.-H. Antifouling indole alkaloids from two marine derived Fungi. Nat. Prod. Commun. 2013, 8, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.-J.; Shao, C.-L.; Wu, L.Y.; Chen, M.; Wang, K.-L.; Zhao, D.-L.; Sun, X.-P.; Chen, G.-Y.; Wang, C.-Y. Bioactive phenylalanine derivatives and cytochalasins from the soft coral-derived fungus, Aspergillus elegans. Mar. Drugs 2013, 11, 2054–2068. [Google Scholar] [CrossRef] [Green Version]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Xian, J.Y.; Liu, Y. Westerdijkin A, a new hydroxyphenylacetic acid derivative from deep sea fungus Aspergillus westerdijkiae SCSIO 05233. Nat. Prod. Res. 2015, 29, 158–162. [Google Scholar] [CrossRef]
- He, F.; Liu, Z.; Yang, J.; Fu, P.; Peng, J.; Zhu, W.-M.; Qi, S.-H. A novel antifouling alkaloid from halotolerant fungus Penicillium sp. OUCMDZ-776. Tetrahedron Lett. 2012, 53, 2280–2283. [Google Scholar] [CrossRef]
- Shao, C.-L.; Xu, R.-F.; Wang, C.-Y.; Qian, P.-Y.; Wang, K.-L.; Wei, M.-Y. Potent antifouling marine dihydroquinolin-2(1H)-one-containing alkaloids from the gorgonian coral-derived fungus Scopulariopsis sp. Mar. Biotechnol. 2015, 17, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Mou, X.-F.; Liu, X.; Xu, R.-F.; Wei, M.-Y.; Fang, Y.-W.; Shao, C.-L. Scopuquinolone B, a new monoterpenoid dihydroquinolin-2(1H)-one isolated from the coral-derived scopulariopsis sp. fungus. Nat. Prod. Res. 2018, 32, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Bovio, E.; Fauchon, M.; Toueix, Y.; Mehiri, M.; Varese, G.C.; Hellio, C. The sponge-associated fungus Eurotium chevalieri MUT 2316 and its bioactive molecules: Potential applications in the field of antifouling. Mar. Biotechnol. 2019, 21, 743–752. [Google Scholar] [CrossRef]
- Wei, M.-Y.; Wang, C.-F.; Wang, K.-L.; Qian, P.-Y.; Wang, C.-Y.; Shao, C.-L. Preparation, structure, and potent antifouling activity of sclerotioramine derivatives. Mar. Biotechnol. 2017, 19, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Scopel, M.; Abraham, W.R.; Henriques, A.T.; Macedo, A.J. Dipeptide cis-cyclo(Leucyl-Tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorg. Med. Chem. Lett. 2013, 23, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Zhang, X.-Y.; Xu, X.-Y.; He, F.; Nong, X.-H.; Qi, S.-H. New cyclic tetrapeptides and asteltoxins from gorgonian-derived fungus Aspergillus sp. SCSGAF 0076. Tetrahedron 2013, 69, 2113–2117. [Google Scholar] [CrossRef]
- Chen, M.; Fu, X.M.; Kong, C.-J.; Wang, C.-Y. Nucleoside derivatives from the marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 2014, 28, 895–900. [Google Scholar] [CrossRef]
- Zhang, H.; Wong, Y.H.; Wang, H.; Chen, Z.; Arellano, S.M.; Ravasi, T.; Qian, P.Y. Quantitative proteomics identify molecular targets that are crucial in larval settlement and metamorphosis of Bugula neritina. J. Proteome Res. 2011, 10, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, L.S.; Zhang, G.; Xu, Y.; Lee, O.O.; Matsumura, K.; Qian, P.Y. The regulatory role of the NO/cGMP signal transduction cascade during larval attachment and metamorphosis of the barnacle Balanus (=Amphibalanus) amphitrite. J. Exp. Biol. 2012, 215, 3813–3822. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.F.; Zhang, H.; He, L.; Liu, C.; Xu, Y.; Qian, P.Y. Butenolide inhibits marine fouling by altering the primary metabolism of three target organisms. ACS Chem. Biol. 2012, 7, 1049–1058. [Google Scholar] [CrossRef]
- Han, Z.; Sun, J.; Zhang, Y.; He, F.; Xu, Y.; Matsumura, K.; He, L.S.; Qiu, J.W.; Qi, S.H.; Qian, P.Y. iTRAQ-based proteomic profiling of the barnacle Balanus amphitrite in response to the antifouling compound meleagrin. J. Proteome. Res. 2013, 12, 2090–2100. [Google Scholar] [CrossRef]
- Artal-Sanz, M.; de Jong, L.; Tavernarakis, N. Caenorhabditis elegans: A versatile platform for drug discovery. Biotechnol. J. 2006, 1, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Boyd, W.A.; McBride, S.J.; Rice, J.R.; Snyder, D.W.; Freedman, J.H. A high-throughput method for assessing chemical toxicity using a Caenorhabditis elegans reproduction assay. Toxicol. Appl. Pharmacol. 2010, 245, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.R.; Kwok, T.C.Y.; Howard, A.; Houston, E.; Johanson, K.; Chan, A.; Cutler, S.R.; McCourt, P.; Roy, P.J. High-throughput screening of small molecules for bioactivity and target identification in Caenorhabditis elegans. Nat. Protoc. 2006, 1, 1906–1914. [Google Scholar] [CrossRef]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Scientific Frontiers in Developmental Toxicology and Risk Assessment; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- National Toxicology Program. A National Toxicology Program for the 21st Century: Roadmap to Achieve the NTP Vision; National Institute of Environmental Health Sciences: Research Triangle, NC, USA, 2004.
- Bischof, L.J.; Huffman, D.L.; Aroian, R.V. Assays for Toxicity Studies in C. elegans with Bt Crystal Proteins. In C. elegans: Methods and Applications; Strange, K., Ed.; Humana Press: Totowa, NJ, USA, 2006; Volume 351, pp. 139–154. [Google Scholar]
- Breger, J.; Fuchs, B.B.; Aperis, G.; Moy, T.I.; Ausubel, F.M.; Mylonakis, E. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 2007, 3, e18. [Google Scholar] [CrossRef]
- Heidarian, S.; Mohammadipanah, F.; Maghsoudlou, A.; Dashti, Y.; Challis, G. Anti-microfouling activity of Glycomyces sediminimaris UTMC 2460 on dominant fouling bacteria of Iran marine habitats. Front. Microbiol. 2019, 9, 3148. [Google Scholar] [CrossRef] [PubMed]
- Viju, N.; Satheesh, S.; Punitha, S.M.J. Antifouling activities of antagonistic marine bacterium Pseudomonas putida associated with an Octopus. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 87, 1113–1124. [Google Scholar] [CrossRef]
Figure 1.
Chemical structures of fatty acids 1 and 2 marine bacteria Shewanella oneidensis SCH0402, and 12-methyltetradecanoid acid (3) from Streptomyces sp.
Figure 1.
Chemical structures of fatty acids 1 and 2 marine bacteria Shewanella oneidensis SCH0402, and 12-methyltetradecanoid acid (3) from Streptomyces sp.

Figure 2.
Chemical structures of butenolides 4–13. Among them, compounds 4–6 were isolated from marine bacteria Streptomyces albidoflavus UST040711-291 and Streptomyces sp. GWS-BW-H5, compounds 7–11 are synthetic ones, and compounds 12 and 13 were isolated from a marine bacterium S. violaceoruber SCH-09.
Figure 2.
Chemical structures of butenolides 4–13. Among them, compounds 4–6 were isolated from marine bacteria Streptomyces albidoflavus UST040711-291 and Streptomyces sp. GWS-BW-H5, compounds 7–11 are synthetic ones, and compounds 12 and 13 were isolated from a marine bacterium S. violaceoruber SCH-09.

Figure 3.
Chemical structures of terpenoids 14–26 from actinobacterial species of Streptomyces. Among them, the diterpene compound 14 was isolated from S. cinnabarinus PK209, the napyradiomycins 15, 16, 18–20, 22 and 25 having a hydrogen atom at C-7 in the semi-naphthoquinone core structure were obtained from S. aculeolatus strain PTM-420, and another napyradiomycin compounds 17, 21, 23, 24 and 26 containing methyl group at C-7 were obtained from S. aculeolatus strain PTM-029.
Figure 3.
Chemical structures of terpenoids 14–26 from actinobacterial species of Streptomyces. Among them, the diterpene compound 14 was isolated from S. cinnabarinus PK209, the napyradiomycins 15, 16, 18–20, 22 and 25 having a hydrogen atom at C-7 in the semi-naphthoquinone core structure were obtained from S. aculeolatus strain PTM-420, and another napyradiomycin compounds 17, 21, 23, 24 and 26 containing methyl group at C-7 were obtained from S. aculeolatus strain PTM-029.

Figure 4.
Chemical structures of steroid compound 27 and 28 from a filamentous bacterium Leucothrix mucor.
Figure 4.
Chemical structures of steroid compound 27 and 28 from a filamentous bacterium Leucothrix mucor.

Figure 5.
Chemical structures of anthraquinones 29–36 from a rare actinobacterium Kitasatospora albolonga R62.
Figure 5.
Chemical structures of anthraquinones 29–36 from a rare actinobacterium Kitasatospora albolonga R62.

Figure 6.
Chemical structures of bisindole alkaloids 37–44 from a marine bacterium Pseudovibrio denitrificans UST4-50.
Figure 6.
Chemical structures of bisindole alkaloids 37–44 from a marine bacterium Pseudovibrio denitrificans UST4-50.

Figure 7.
Chemical structures of diketopiperazine alkaloids 45 and 46 from Streptomyces praecox 291-11, and maipomycin A 47 from a rare actinomycete Kibdelosporangium phytohabitans XY-R10.
Figure 7.
Chemical structures of diketopiperazine alkaloids 45 and 46 from Streptomyces praecox 291-11, and maipomycin A 47 from a rare actinomycete Kibdelosporangium phytohabitans XY-R10.

Figure 8.
Chemical structures of the glycolipid surfactant 48 from a marine strain of Serratia marcescens, the benzenoid compound 4-phenylbutanoic acid (49) from a bacterium Bacillus pumilus strain S6-15, the flavonoid compound taxifolin (50) from actinobacterial Streptomyces sampsonii PM33, and the polyketide compound elasnin (51) from S. mobaraensis DSM 40847.
Figure 8.
Chemical structures of the glycolipid surfactant 48 from a marine strain of Serratia marcescens, the benzenoid compound 4-phenylbutanoic acid (49) from a bacterium Bacillus pumilus strain S6-15, the flavonoid compound taxifolin (50) from actinobacterial Streptomyces sampsonii PM33, and the polyketide compound elasnin (51) from S. mobaraensis DSM 40847.

Figure 9.
Chemical structures of the butenolide compound maculalactone A (52) from a marine cyanobacterium Kyrtuthrix maculans.
Figure 9.
Chemical structures of the butenolide compound maculalactone A (52) from a marine cyanobacterium Kyrtuthrix maculans.

Figure 10.
Chemical structures of 53–58 belonging to the polyketide–polypeptide structural family from marine cyanobacteria. Among them, compounds 53–56 were isolated from Lyngbya majuscula, and the dolastatin 16 (53) and lyngbyabellins 57 and 58 were isolated from Okeania sp.
Figure 10.
Chemical structures of 53–58 belonging to the polyketide–polypeptide structural family from marine cyanobacteria. Among them, compounds 53–56 were isolated from Lyngbya majuscula, and the dolastatin 16 (53) and lyngbyabellins 57 and 58 were isolated from Okeania sp.

Figure 11.
Chemical structures of bisabolane-type sesquiterpenoids 59 and 60 from Aspergillus sp., and sesterterpenes 61–63 belonging to the polyketide–polypeptide structural family from Emericella variecolor.
Figure 11.
Chemical structures of bisabolane-type sesquiterpenoids 59 and 60 from Aspergillus sp., and sesterterpenes 61–63 belonging to the polyketide–polypeptide structural family from Emericella variecolor.

Figure 12.
Chemical structures of chlorinated phenols 64–70 from marine fungi. Among them, compound 61 was isolated from Ampelomyces sp. UST040128, compounds 65 was isolated from Sarcophyton sp., and compounds 66 and 67 were isolated from Pestalotiopsis sp. ZJ-2009-7-6, and compounds 68–70 were isolated from strains Penicillium brevicompactum DFFSCS025 and Aspergillus versicolor SCSIO 41502.
Figure 12.
Chemical structures of chlorinated phenols 64–70 from marine fungi. Among them, compound 61 was isolated from Ampelomyces sp. UST040128, compounds 65 was isolated from Sarcophyton sp., and compounds 66 and 67 were isolated from Pestalotiopsis sp. ZJ-2009-7-6, and compounds 68–70 were isolated from strains Penicillium brevicompactum DFFSCS025 and Aspergillus versicolor SCSIO 41502.

Figure 13.
Chemical structures of phenyl ethers 71–78 from fungal strains of Aspergillus. Among them, compounds 71–74 were isolated from A. versicolor SCSIO 41502, and compounds 75–78 were synthetic derivatives of natural products from Aspergillus sp. XS-20090066.
Figure 13.
Chemical structures of phenyl ethers 71–78 from fungal strains of Aspergillus. Among them, compounds 71–74 were isolated from A. versicolor SCSIO 41502, and compounds 75–78 were synthetic derivatives of natural products from Aspergillus sp. XS-20090066.

Figure 14.
Chemical structures of benzylazaphilone analogue 79 from Aspergillus sp., compound 80 from Xylariaceae sp., and (±)-eurotiumides B (±81) and D (±82) from Eurotium sp. XS-200900E6.
Figure 14.
Chemical structures of benzylazaphilone analogue 79 from Aspergillus sp., compound 80 from Xylariaceae sp., and (±)-eurotiumides B (±81) and D (±82) from Eurotium sp. XS-200900E6.

Figure 15.
Chemical structures of anthraquinones and xanthones 83–86 from strains of Aspergillus, compound 87 from an unidentified fungal strain ZSUH-36, and anthraquinones 88 and 89 from Penicillium sp. SCSGAF 0023.
Figure 15.
Chemical structures of anthraquinones and xanthones 83–86 from strains of Aspergillus, compound 87 from an unidentified fungal strain ZSUH-36, and anthraquinones 88 and 89 from Penicillium sp. SCSGAF 0023.

Figure 16.
Chemical structures of 14-membered resorcylic acid lactones 90–97 from marine fungal strains of Cochliobolus lunatus.
Figure 16.
Chemical structures of 14-membered resorcylic acid lactones 90–97 from marine fungal strains of Cochliobolus lunatus.

Figure 17.
Chemical structures of alkaloids 98–110a/b from marine fungal strains of Aspergillus and Penicillium. Among them, compounds 98–100 were isolated from A. sydowii SCSIO 00305, cytochalasins 101–104 were isolated from A. elegans ZJ-2008010, (+)-circumdatin F (105) was isolated from A. alliaceus, A. westerdijkiae DFFSCS013 and A. westerdijkiae SCSIO 05233, and indole alkaloids 106–109 and penispirolloid A (110a/b) were obtained from Penicillium sp. SCSIO 00258 and OUCMDZ-776, respectively.
Figure 17.
Chemical structures of alkaloids 98–110a/b from marine fungal strains of Aspergillus and Penicillium. Among them, compounds 98–100 were isolated from A. sydowii SCSIO 00305, cytochalasins 101–104 were isolated from A. elegans ZJ-2008010, (+)-circumdatin F (105) was isolated from A. alliaceus, A. westerdijkiae DFFSCS013 and A. westerdijkiae SCSIO 05233, and indole alkaloids 106–109 and penispirolloid A (110a/b) were obtained from Penicillium sp. SCSIO 00258 and OUCMDZ-776, respectively.

Figure 18.
Chemical structures of alkaloids 111–116 from marine fungi Scopulariopsis sp., and the Cyclo-L-Trp-L-Ala (117) from Eurotium chevalieri MUT 2316.
Figure 18.
Chemical structures of alkaloids 111–116 from marine fungi Scopulariopsis sp., and the Cyclo-L-Trp-L-Ala (117) from Eurotium chevalieri MUT 2316.

Figure 19.
Chemical structures of the natural (+)-sclerotiorin (118) from a fungus Penicillium sclerotiorum CHNSCLM-0013 and its semisynthetic sclerotioramine derivatives 119–135.
Figure 19.
Chemical structures of the natural (+)-sclerotiorin (118) from a fungus Penicillium sclerotiorum CHNSCLM-0013 and its semisynthetic sclerotioramine derivatives 119–135.

Figure 20.
Chemical structures of the dipeptide cis-cyclo (Leucyl-Tyrosyl) (136) from Penicillium sp. F37, the cyclic tetrapeptide aspergillipeptide C (137) from Aspergillus sp. SCSGAF 0076, and the nucleoside diacetylkipukasin E (138) from Aspergillus versicolor.
Figure 20.
Chemical structures of the dipeptide cis-cyclo (Leucyl-Tyrosyl) (136) from Penicillium sp. F37, the cyclic tetrapeptide aspergillipeptide C (137) from Aspergillus sp. SCSGAF 0076, and the nucleoside diacetylkipukasin E (138) from Aspergillus versicolor.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, K.-L.; Dou, Z.-R.; Gong, G.-F.; Li, H.-F.; Jiang, B.; Xu, Y. Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents. Mar. Drugs 2022, 20, 90. https://doi.org/10.3390/md20020090
AMA Style
Wang K-L, Dou Z-R, Gong G-F, Li H-F, Jiang B, Xu Y. Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents. Marine Drugs. 2022; 20(2):90. https://doi.org/10.3390/md20020090
Chicago/Turabian StyleWang, Kai-Ling, Zheng-Rong Dou, Gao-Fen Gong, Hai-Feng Li, Bei Jiang, and Ying Xu. 2022. "Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents" Marine Drugs 20, no. 2: 90. https://doi.org/10.3390/md20020090
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.