Modulation of Glial Responses by Furanocembranolides: Leptolide Diminishes Microglial Inflammation in Vitro and Ameliorates Gliosis In Vivo in a Mouse Model of Obesity and Insulin Resistance

 , ,
, ,
Abstract
:
1. Introduction
2. Results
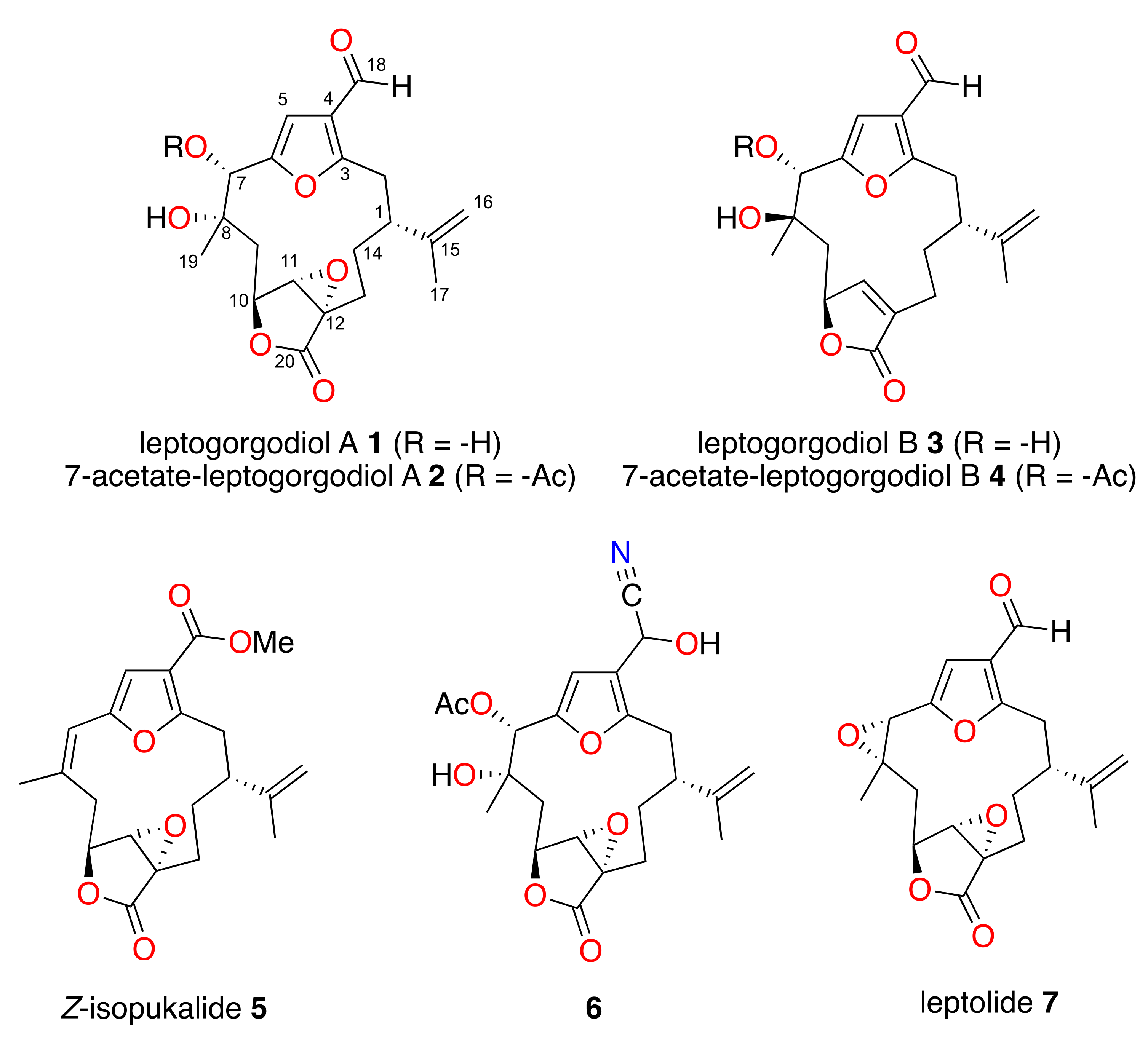
2.1. Characterization of Novel Furanocembranolides
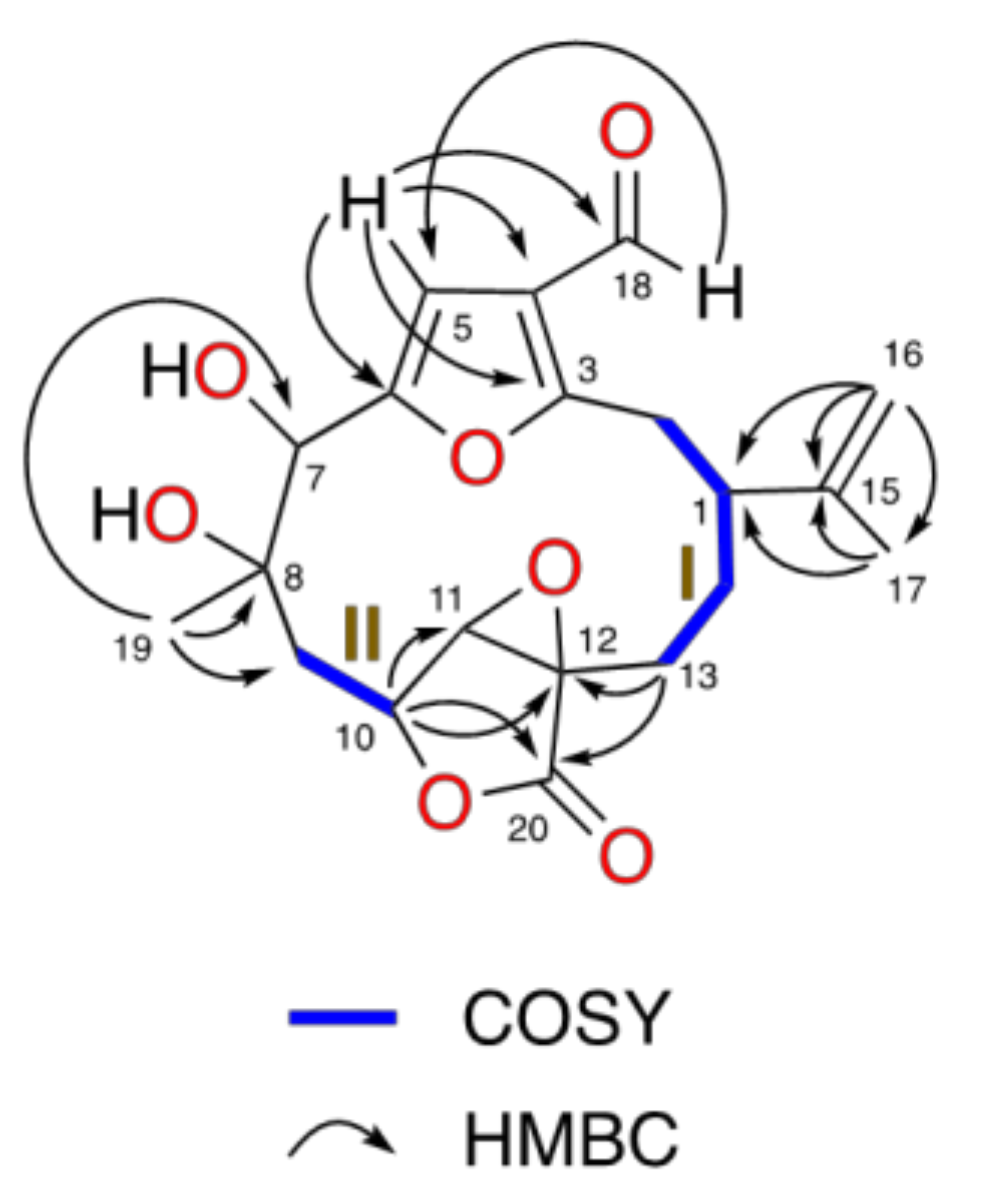
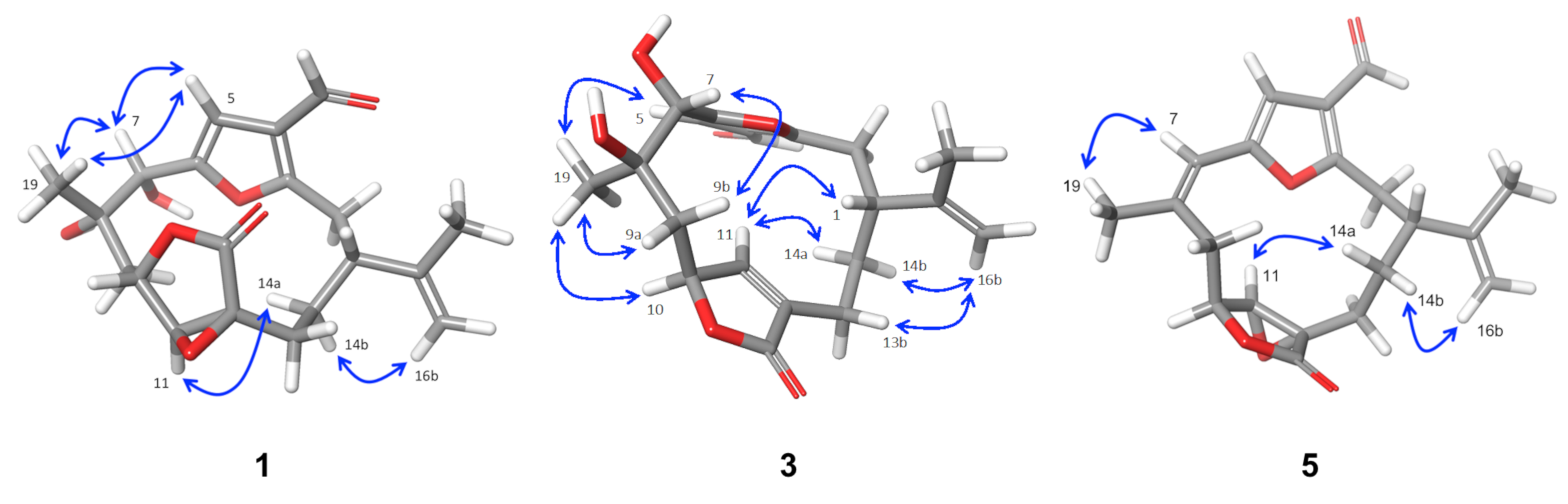
2.2. Relative Configuration of Compounds 1–6
2.3. Absolute Configuration
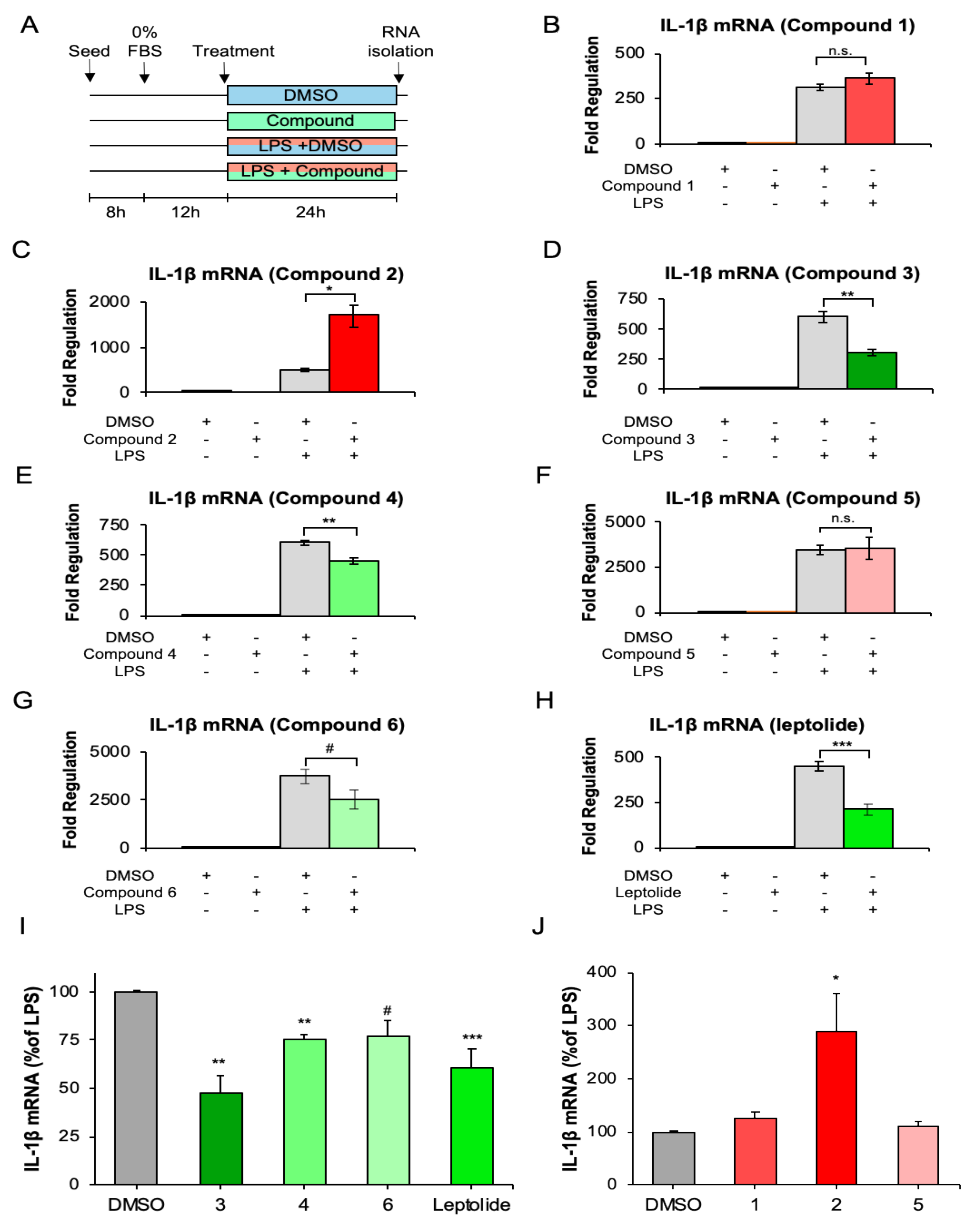
2.4. Furanocembranolides Modulate LPS-Induced Inflammatory Responses on Microglial Cells
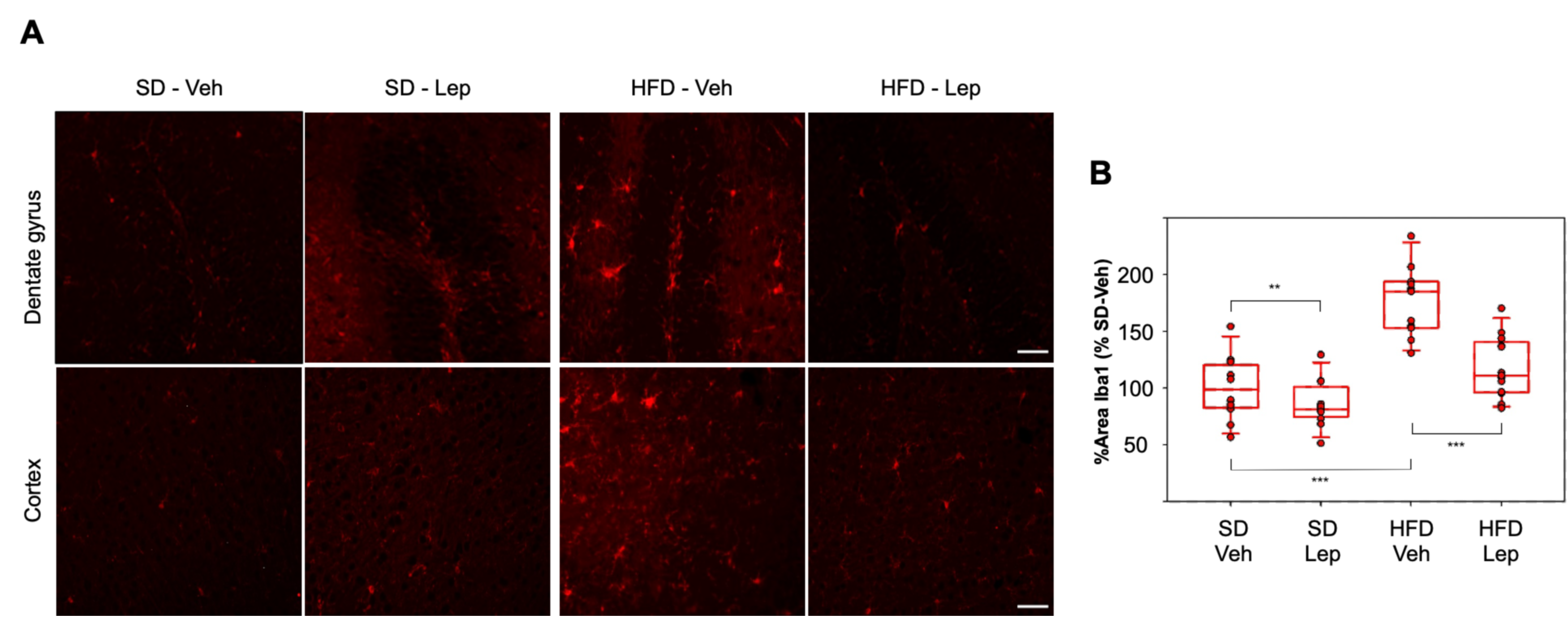
2.5. Leptolide Treatment in Vivo Ameliorates Microgliosis in a Mouse Model of Obesity and Insulin Resistance Triggered by HFD (High Fat Diet)
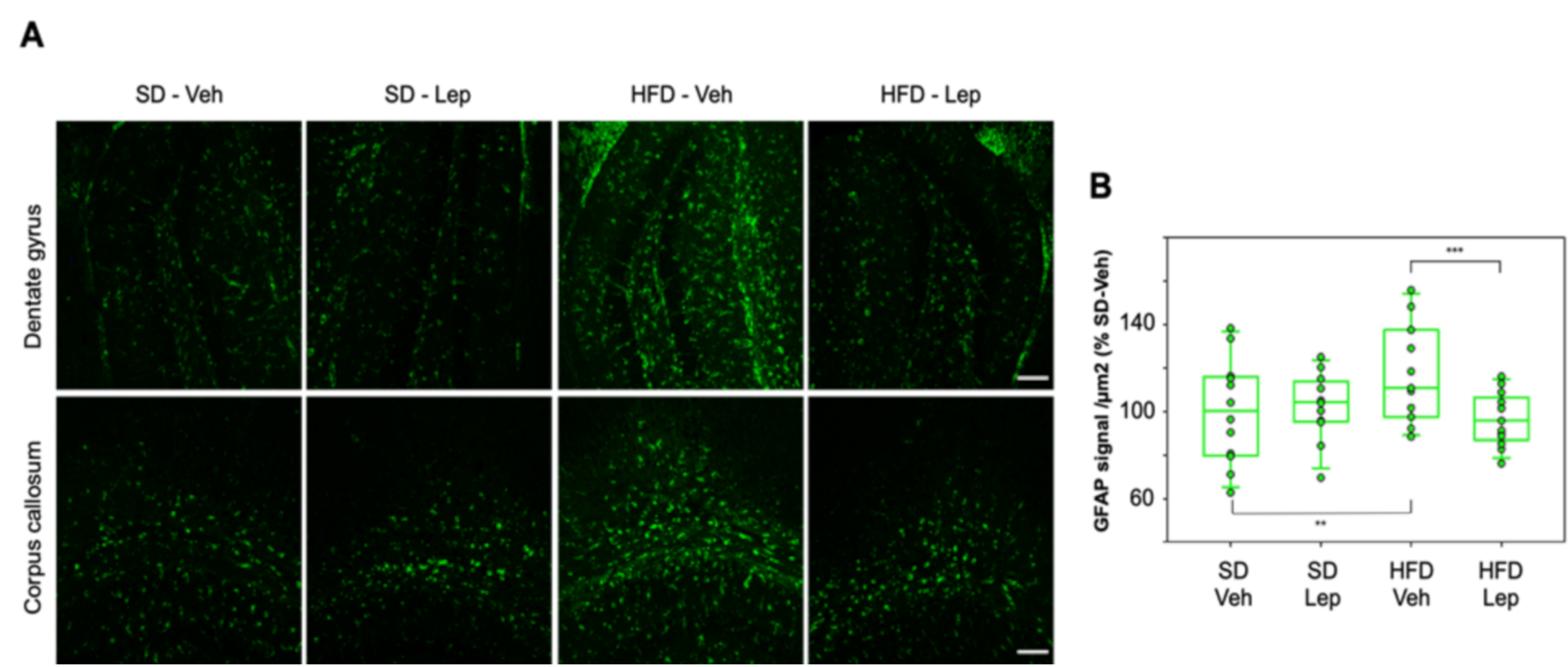
2.6. Leptolide Treatment in Vivo Also Reduces Astrogliosis
3. Discussion
4. Experimental Section
4.1. General Experimental Procedures
4.2. Collection, Extraction and Isolation
4.3. Reagents Preparation
4.4. Cell Culture and Treatments
4.5. MTT Viability Assay
4.6. RNA Isolation from Cell Cultures
4.7. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.8. Animal Procedures
4.9. Immunohistochemistry
4.10. Image Acquisition
4.11. Image Quantification
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division World Population Ageing 2019: Highlights; 2019. Available online: https://www.un.org/en/development/desa/population/publications/pdf/ageing/WorldPopulationAgeing2019-Highlights.pdf (accessed on 27 May 2020).
- McManus, R.M.; Heneka, M.T. Role of neuroinflammation in neurodegeneration: New insights. Alzheimers Res. Ther. 2017, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yacoubian, T.A. Chapter 1-Neurodegenerative Disorders: Why Do We Need New Therapies? In Drug Discovery Approaches for the Treatment of Neurodegenerative Disorders; Adejare, A., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 1–16. ISBN 978-0-12-802810-0. [Google Scholar]
- Chen, W.-W.; Zhang, X.; Huang, W.-J. Role of neuroinflammation in neurodegenerative diseases (Review). Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, V.H.; Teeling, J. Microglia and macrophages of the central nervous system: The contribution of microglia priming and systemic inflammation to chronic neurodegeneration. Semin. Immunopathol. 2013, 35, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [Google Scholar] [CrossRef]
- Boche, D.; Perry, V.H.; Nicoll, J.A.R. Review: Activation patterns of microglia and their identification in the human brain: Microglia in human brain. Neuropathol. Appl. Neurobiol. 2013, 39, 3–18. [Google Scholar] [CrossRef]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef]
- Prinz, M.; Priller, J. Microglia and brain macrophages in the molecular age: From origin to neuropsychiatric disease. Nat. Rev. Neurosci. 2014, 15, 300–312. [Google Scholar] [CrossRef]
- Du, L.; Zhang, Y.; Chen, Y.; Zhu, J.; Yang, Y.; Zhang, H.-L. Role of Microglia in Neurological Disorders and Their Potentials as a Therapeutic Target. Mol. Neurobiol. 2017, 54, 7567–7584. [Google Scholar] [CrossRef]
- Cevenini, E.; Monti, D.; Franceschi, C. Inflamm-ageing. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 14–20. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Kim, J.; Min, K.-J.; Seol, W.; Jou, I.; Joe, E. Astrocytes in injury states rapidly produce anti-inflammatory factors and attenuate microglial inflammatory responses. J. Neurochem. 2018, 115, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Dorta, E.; Díaz-Marrero, A.R.; Brito, I.; Cueto, M.; D’Croz, L.; Darias, J. The oxidation profile at C-18 of furanocembranolides may provide a taxonomical marker for several genera of octocorals. Tetrahedron 2007, 63, 9057–9062. [Google Scholar] [CrossRef]
- Díaz-Marrero, A.R.; Porras, G.; Cueto, M.; D’Croz, L.; Lorenzo, M.; San-Martín, A.; Darias, J. Leptogorgolide, a biogenetically interesting 1,4-diketo-cembranoid that reinforces the oxidation profile of C-18 as taxonomical marker for octocorals. Tetrahedron 2009, 65, 6029–6033. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.J.; Wu, T.Y.; Su, T.R.; Wen, Z.H.; Chen, J.J.; Fang, L.S.; Wu, Y.C.; Sung, P.J. Terpenoids from the Octocoral Sinularia gaweli. Int. J. Mol. Sci. 2015, 16, 19508–19517. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.-H.; Chou, K.-J.; Huang, C.-Y.; Wen, Z.-H.; Hsu, C.-H.; Wu, Y.-C.; Dai, C.-F.; Sheu, J.-H. Bioactive Cembranoids from the Soft Coral Sinularia crassa. Mar. Drugs 2011, 9, 1955–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Procaccini, C.; Santopaolo, M.; Faicchia, D.; Colamatteo, A.; Formisano, L.; de Candia, P.; Galgani, M.; De Rosa, V.; Matarese, G. Role of metabolism in neurodegenerative disorders. Metabolism 2016, 65, 1376–1390. [Google Scholar] [CrossRef]
- Yin, F.; Yao, J.; Brinton, R.D.; Cadenas, E. The Metabolic-Inflammatory Axis in Brain Aging and Neurodegeneration; Frontiers Research Topics; Frontiers Media SA: Lausanne, Switzerland, 2017; ISBN 978-2-88945-253-8. [Google Scholar]
- Pugazhenthi, S.; Qin, L.; Reddy, P.H. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1037–1045. [Google Scholar] [CrossRef]
- Ramos-Rodriguez, J.J.; Jimenez-Palomares, M.; Murillo-Carretero, M.I.; Infante-Garcia, C.; Berrocoso, E.; Hernandez-Pacho, F.; Lechuga-Sancho, A.M.; Cozar-Castellano, I.; Garcia-Alloza, M. Central vascular disease and exacerbated pathology in a mixed model of type 2 diabetes and Alzheimer’s disease. Psychoneuroendocrinology 2015, 62, 69–79. [Google Scholar] [CrossRef]
- Ramos-Rodriguez, J.J.; Ortiz, O.; Jimenez-Palomares, M.; Kay, K.R.; Berrocoso, E.; Murillo-Carretero, M.I.; Perdomo, G.; Spires-Jones, T.; Cozar-Castellano, I.; Lechuga-Sancho, A.M.; et al. Differential central pathology and cognitive impairment in pre-diabetic and diabetic mice. Psychoneuroendocrinology 2013, 38, 2462–2475. [Google Scholar] [CrossRef]
- Villa-Pérez, P.; Cueto, M.; Díaz-Marrero, A.R.; Lobatón, C.D.; Moreno, A.; Perdomo, G.; Cózar-Castellano, I. Leptolide Improves Insulin Resistance in Diet-Induced Obese Mice. Mar. Drugs 2017, 15. [Google Scholar] [CrossRef] [Green Version]
- López-Acosta, J.F.; Moreno-Amador, J.L.; Jiménez-Palomares, M.; Díaz-Marrero, A.R.; Cueto, M.; Perdomo, G.; Cózar-Castellano, I. Epoxypukalide Induces Proliferation and Protects against Cytokine-Mediated Apoptosis in Primary Cultures of Pancreatic β-Cells. PloS ONE 2013, 8, e52862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Acosta, J.F.; Villa-Pérez, P.; Fernández-Díaz, C.M.; de Román, D.L.; Díaz-Marrero, A.R.; Cueto, M.; Perdomo, G.; Cózar-Castellano, I. Protective effects of epoxypukalide on pancreatic β-cells and glucose metabolism in STZ-induced diabetic mice. Islets 2015, 7, e1078053. [Google Scholar] [CrossRef] [Green Version]
- PCModel; (v.9.2) Serena Software. Maestro v. 11.5.010, Schrödinger; LLC: New York, NY, USA, 2018.
- Gallardo, A.B.; Díaz-Marrero, A.R.; de la Rosa, J.M.; D’Croz, L.; Perdomo, G.; Cózar-Castellano, I.; Darias, J.; Cueto, M. Chloro-Furanocembranolides from Leptogorgia sp. Improve Pancreatic Beta-Cell Proliferation. Mar. Drugs 2018, 16, 49. [Google Scholar] [CrossRef] [Green Version]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, K.K.W. Glial Fibrillary acidic protein: From intermediate filament assembly and gliosis to neurobiomarker. Trends Neurosci. 2015, 38, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.-Y.; Wen, Z.-H.; Wang, S.-K.; Chiou, S.-F.; Hsu, C.-H.; Dai, C.-F.; Duh, C.-Y. Anti-inflammatory cembranolides from the soft coral Lobophytum durum. Bioorg. Med. Chem. 2009, 17, 3763–3769. [Google Scholar] [CrossRef]
- Lai, K.-H.; You, W.-J.; Lin, C.-C.; El-Shazly, M.; Liao, Z.-J.; Su, J.-H. Anti-Inflammatory Cembranoids from the Soft Coral Lobophytum crassum. Mar. Drugs 2017, 15, 327. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Liu, X.; Jiang, M.; Luo, G.; Wu, Z.; Chen, B.; Li, J.; Liu, L.; Chen, S. Anti-Inflammatory Cembrane-Type Diterpenoids and Prostaglandins from Soft Coral Lobophytum sarcophytoides. Mar. Drugs 2019, 17, 481. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Bent, R.; Moll, L.; Grabbe, S.; Bros, M. Interleukin-1 Beta-A Friend or Foe in Malignancies? IJMS 2018, 19, 2155. [Google Scholar] [CrossRef] [Green Version]
- Marucha, P.T.; Zeff, R.A.; Kreutzer, D.L. Cytokine-induced IL-1 beta gene expression in the human polymorphonuclear leukocyte: Transcriptional and post-transcriptional regulation by tumor necrosis factor and IL-1. J. Immunol. 1991, 147, 2603. [Google Scholar]
- Donnelly, R.P.; Fenton, M.J.; Kaufman, J.D.; Gerrard, T.L. IL-1 expression in human monocytes is transcriptionally and posttranscriptionally regulated by IL-4. J. Immunol. 1991, 146, 3431. [Google Scholar] [PubMed]
- Qin, X.; Jiang, X.; Jiang, X.; Wang, Y.; Miao, Z.; He, W.; Yang, G.; Lv, Z.; Yu, Y.; Zheng, Y. Micheliolide inhibits LPS-induced inflammatory response and protects mice from LPS challenge. Sci. Rep. 2016, 6, 23240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Guo, W.; Li, L.; Xu, C.; Yang, D.; Wang, S.; Lu, Y.; Zhang, Q.; Zhai, J.; Fan, H.; et al. Micheliolide Derivative DMAMCL Inhibits Glioma Cell Growth In Vitro and In Vivo. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascua-Maestro, R.; Corraliza-Gomez, M.; Diez-Hermano, S.; Perez-Segurado, C.; Ganfornina, M.D.; Sanchez, D. The MTT-formazan assay: Complementary technical approaches and in vivo validation in Drosophila larvae. Acta Histochem. 2018, 120, 179–186. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.J.; Foster, T.D.; Thomas, W.E. Cellular forms and functions of brain microglia. Brain Res. Bull. 1994, 34, 73–78. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | ||
|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 40.1, CH | 3.24, m | 40.2, CH | 3.27, dd (11.2, 11.2) |
| 2 | 31.9, CH2 | 3.00, m 3.05, m | 31.6, CH2 | 2.96, dd (2.9, 16.8) 3.05, dd (11.8, 16.8) |
| 3 | 162.4, C | - | 162.6, C | - |
| 4 | 123.2, C | - | 124.3, C | - |
| 5 | 106.4, CH | 6.69, s | 107.0, CH | 6.63, s |
| 6 | 155.2, C | - | 151.8, C | - |
| 7 | 73.2, CH | 5.28, s | 74.6, CH | 6.21, s |
| 8 | 74.5, C | - | 73.9, C | - |
| 9 | 40.5, CH2 | 1.51, dd (11.0, 15.2) 1.86, dd (5.0, 15.2) | 40.8, CH2 | 1.45, dd (12.2, 14.9) 1.86, dd (5.4, 14.9) |
| 10 | 74.5, CH | 4.85, dd (5.0, 11.0) | 74.3, CH | 4.81, dd (5.8, 10.7) |
| 11 | 63.0, CH | 3.70, s | 62.9, CH | 3.72, s |
| 12 | 61.0, C | - | 61.0, C | - |
| 13 | 22.6, CH2 | a: 1.39, m b: 2.49, dd (11.2, 14.7) | 22.5 CH2 | a: 1.36, m b: 2.47, dd (11.5, 14.7) |
| 14 | 29.2, CH2 | a: 1.26, m b: 1.88, m | 28.8 CH2 | a: 1.19, m b: 1.83, m |
| 15 | 144.9, C | - | 144.7, C | - |
| 16 | 114.1, CH2 | a: 4.97, dd (1.6, 1.6) b: 5.10, s | 114.2, CH2 | a: 4.97, dd, (1.6, 1.6) b: 5.10, s |
| 17 | 18.8, CH3 | 1.77, s | 18.8 CH3 | 1.76, s |
| 18 | 184.7, CH | 9.89, s | 184.6, CH | 9.87, s |
| 19 | 22.5, CH3 | 1.34, s | 23.4, CH3 | 1.39, s |
| 20 | 172.5, C | - | 171.4, C | - |
| 7-CH3CO | 170.1, C | - | ||
| 7-CH3CO | 21.0, CH3 | 2.12, s | ||
| No. | 3 | 4 | ||
|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 43.8, CH | 2.22, m | 43.8, CH | 2.25, m |
| 2 | 31.8, CH2 | 2.81, dd (2.2, 14.1) 3.09, dd (11.6, 14.8) | 31.8, CH2 | 2.89, dd (2.9, 14.8) 3.06, dd (12.2, 14.8) |
| 3 | 162.9, C | - | 163.5, C | - |
| 4 | 125.6, C | - | 125.3, C | - |
| 5 | 106.4, CH | 6.78, s | 106.9, CH | 6.70, s |
| 6 | 154.6, C | - | 151.5, C | - |
| 7 | 75.6, CH | 4.57, s | 75.9, CH | 5.59, s |
| 8 | 73.7, C | - | 72.8, C | - |
| 9 | 43.0, CH | b: 1.87, dd (11.6, 14.8) a: 2.57, dd (4.4, 14.8) | 43.2, CH2 | b: 1.95, dd (11.9, 15.1) a: 2.60, dd (4.0, 15.1) |
| 10 | 78.8, CH | 4.96, m | 78.5, CH | 4.96, m |
| 11 | 148.7, CH | 5.81, s | 148.4, CH | 5.85, s |
| 12 | 133.6, C | - | 133.8, C | - |
| 13 | 21.7 CH2 | a: 2.10, m b: 2.31, m | 21.8, CH2 | a: 2.13, m b: 2.36, m |
| 14 | 28.5 CH2 | a: 1.49, m b: 1.82, m | 28.7, CH2 | a: 1.49, m b: 1.87, m |
| 15 | 145.7, C | - | 145.5, C | - |
| 16 | 113.2, CH2 | a: 4.82, s b: 4.87, dd (1.6, 1.6) | 113.3, CH2 | a: 4.84, s b: 4.89, dd (1.6, 1.6) |
| 17 | 19.2 CH3 | 1.77 s, br s | 19.3, CH3 | 1.78, s |
| 18 | 184.6, CH | 9.94, s | 184.3, C | 9.95, s |
| 19 | 19.7, CH3 | 1.40, s | 21.0, CH3 | 1.48, s |
| 20 | 173.5, C | - | 171.9, C | - |
| 7-CH3CO | 169.5, C | - | ||
| 7-CH3CO | 21.1, CH3 | 2.16, s | ||
| No. | 5 | 6 * | ||
|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 43.5, CH | 2.56 m | 41.0, CH | 3.13, m |
| 2 | 32.8, CH2 | 2.83, dd (2.6, 16.6) 3.39, dd (12.5, 16.6) | 30.7, CH2 | 2.73, m 2.65, m |
| 3 | 159.8, C | - | 152.0 / 152.2, C | - |
| 4 | 115.9, C | - | 118.4 / 118.3, C | - |
| 5 | 110.4, CH | 6.45, s | 108.5 / 108.8, CH | 6.45 / 6.49, s |
| 6 | 150.4, C | - | 150.2 / 150.4, CH | - |
| 7 | 117.4, CH | 6.18, s | 75.0 / 74.9, CH | 6.17, s |
| 8 | 129.4, C | - | 73.9 / 74.0, C | - |
| 9 | 36.6, CH2 | 3.56, m 2.58, m | 40.9 / 40.8, CH | 1.88 m 1.55, m |
| 10 | 76.9, CH | 4.60, dd (4.5, 12.8) | 74.6 / 74.5, CH | 4.79, m |
| 11 | 61.3, CH | 3.81, s | 62.9, CH | 3.76, s |
| 12 | 60.8, C | - | 60.9 / 61.0, C | - |
| 13 | 21.7, CH2 | 1.59, m 2.62, m | 22.5, CH2 | 1.38, m 2.46, m |
| 14 | 28.8, CH2 | a: 1.05, m b: 1.65, m | 28.5 / 28.6, CH2 | a: 1.23, m b: 1.81, m |
| 15 | 144.3, C | - | 145.1, C | - |
| 16 | 113.8, CH2 | a: 4.89, s b: 4.99, dd (1.6, 1.6) | 113.9, CH2 | a: 4.95, s b: 5.06, s |
| 17 | 19.4, CH3 | 1.76, s | 18.8, CH3 | 1.74, s |
| 18 | 164.3, C | - | 56.1, CH | 5.38 / 5.39, s |
| 19 | 25.2, CH3 | 2.01, s | 23.5 / 23.6, CH3 | 1.40, s |
| 20 | 172.2, C | - | 171.8, C | - |
| 21 | 51.7, CH3 | 3.82 s | - | |
| 7-CH3CO | 170.4, C | |||
| 7-CH3CO | 21.1, CH3 | 2.15, s | ||
| 18-CN | 117.9 / 117.6, C | |||
 |  | ||
|---|---|---|---|
| No. | 2 | 6 | 4 |
| δH-7 | 6.21, s | 6.17, s | 5.59, s |
| δH-9 | 1.45, dd 1.86, dd | 1.55, m 1.88 m | 1.95, dd 2.60, dd |
| δH-19 | 1.39, s | 1.35, s | 1.48 s |
| δC-7 | 74.6 | 74.0 | 75.9 |
| δC-9 | 40.8 | 40.5 | 43.2 |
| δC-19 | 23.4 | 22.4 | 21.0 |
| No. | δR | δS | ΔδR-S |
|---|---|---|---|
| δH-18 | 9.84 | 9.77 | +0.070 |
| δH-5 | 6.53 | 6.08 | +0.45 |
| δH-19 | 1.10 | 1.30 | −0.20 |
| δH-10 | 4.60 | 4.72 | −0.12 |
| Gene Name (ID) | Primer Name | Sequence (5′-3′) |
|---|---|---|
| IL-1β (NM_008361.4) | Mouse IL1β-Forward | TGTAATGAAAGACGGCACACCCAC |
| Mouse IL1β-Reverse | GGCTTGTGCTCTGCTTGTGAGG | |
| Rpl18 (NM_009077.2) | Mouse Rpl18-Forward | TTCCGTCTTTCCGGACCT |
| Mouse Rpl18-Reverse | TCGGCTCATGAACAACCTCT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corraliza-Gómez, M.; Gallardo, A.B.; Díaz-Marrero, A.R.; de la Rosa, J.M.; D’Croz, L.; Darias, J.; Arranz, E.; Cózar-Castellano, I.; Ganfornina, M.D.; Cueto, M. Modulation of Glial Responses by Furanocembranolides: Leptolide Diminishes Microglial Inflammation in Vitro and Ameliorates Gliosis In Vivo in a Mouse Model of Obesity and Insulin Resistance. Mar. Drugs 2020, 18, 378. https://doi.org/10.3390/md18080378
Corraliza-Gómez M, Gallardo AB, Díaz-Marrero AR, de la Rosa JM, D’Croz L, Darias J, Arranz E, Cózar-Castellano I, Ganfornina MD, Cueto M. Modulation of Glial Responses by Furanocembranolides: Leptolide Diminishes Microglial Inflammation in Vitro and Ameliorates Gliosis In Vivo in a Mouse Model of Obesity and Insulin Resistance. Marine Drugs. 2020; 18(8):378. https://doi.org/10.3390/md18080378
Chicago/Turabian StyleCorraliza-Gómez, Miriam, Amalia B. Gallardo, Ana R. Díaz-Marrero, José M. de la Rosa, Luis D’Croz, José Darias, Eduardo Arranz, Irene Cózar-Castellano, María D. Ganfornina, and Mercedes Cueto. 2020. "Modulation of Glial Responses by Furanocembranolides: Leptolide Diminishes Microglial Inflammation in Vitro and Ameliorates Gliosis In Vivo in a Mouse Model of Obesity and Insulin Resistance" Marine Drugs 18, no. 8: 378. https://doi.org/10.3390/md18080378