Entomotoxic Activity of Prasiola crispa (Antarctic Algae) in Nauphoeta cinerea Cockroaches: Identification of Main Steroidal Compounds

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
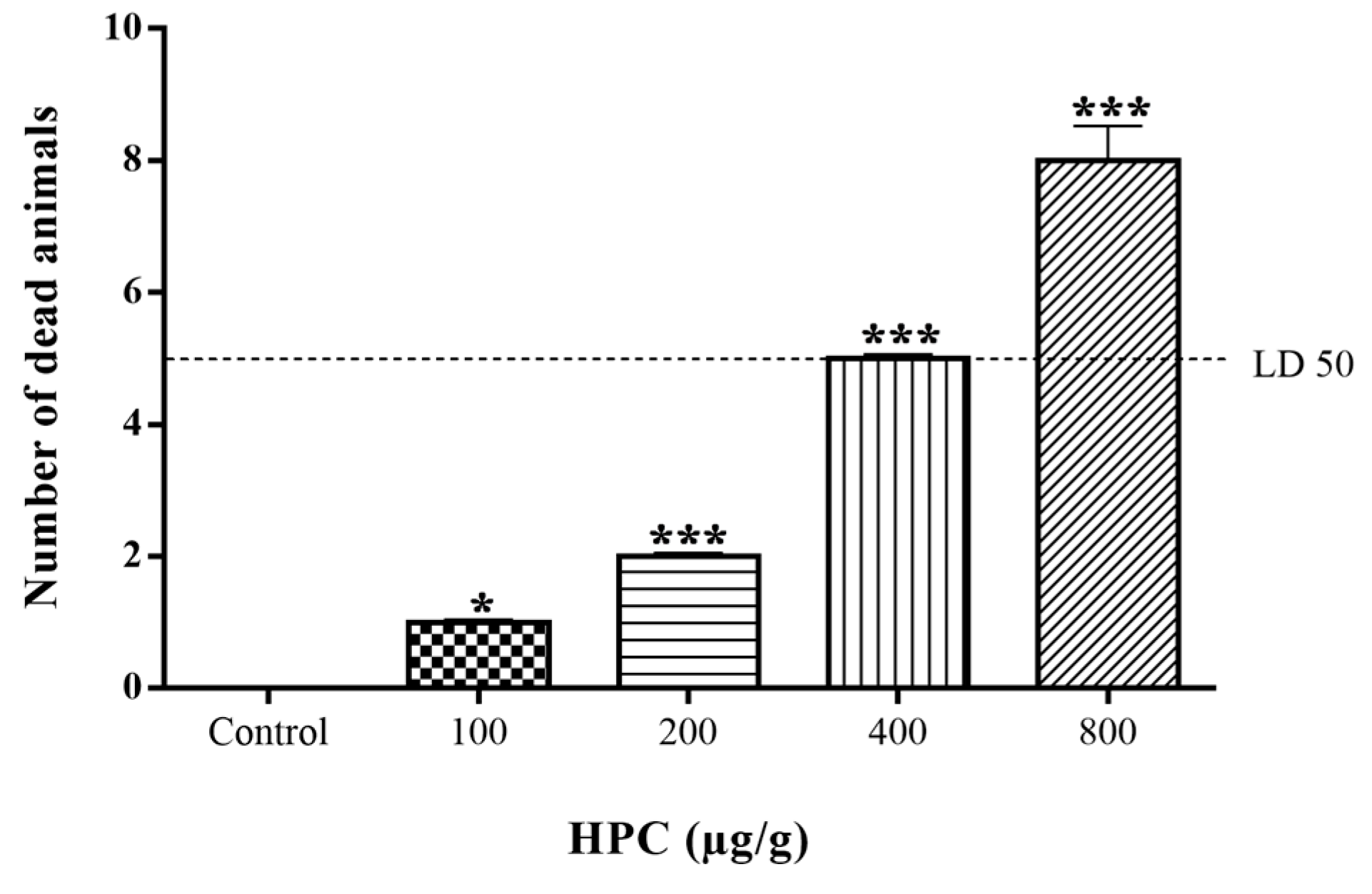
2.1. Insecticidal Activity of n-Hexane Extract of Prasiola crispa
2.2. The AChE Activity in Cockroach Brain Homogenates Was not Affected by HPC
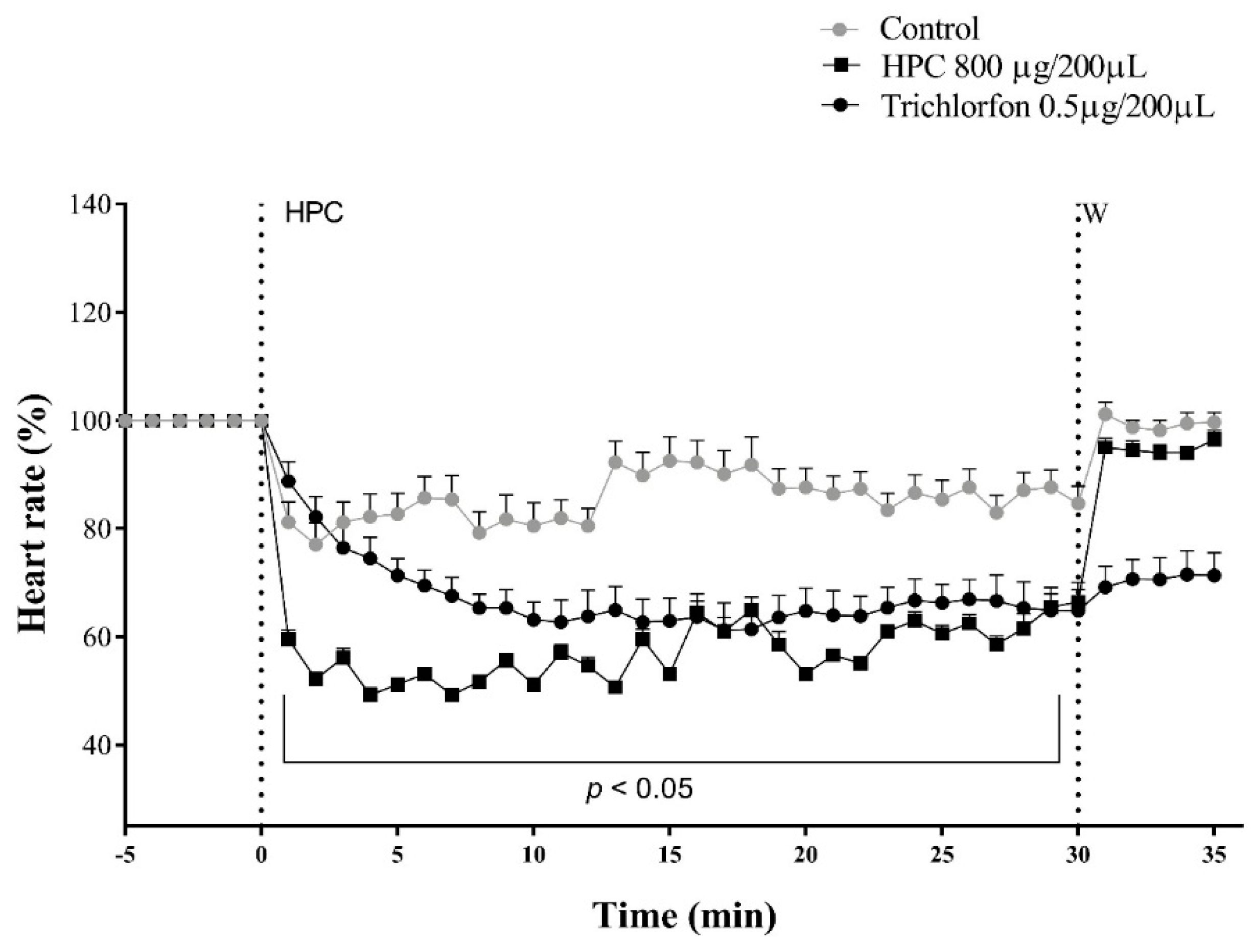
2.3. Effects of HPC on the Cockroach’s Heart Rate
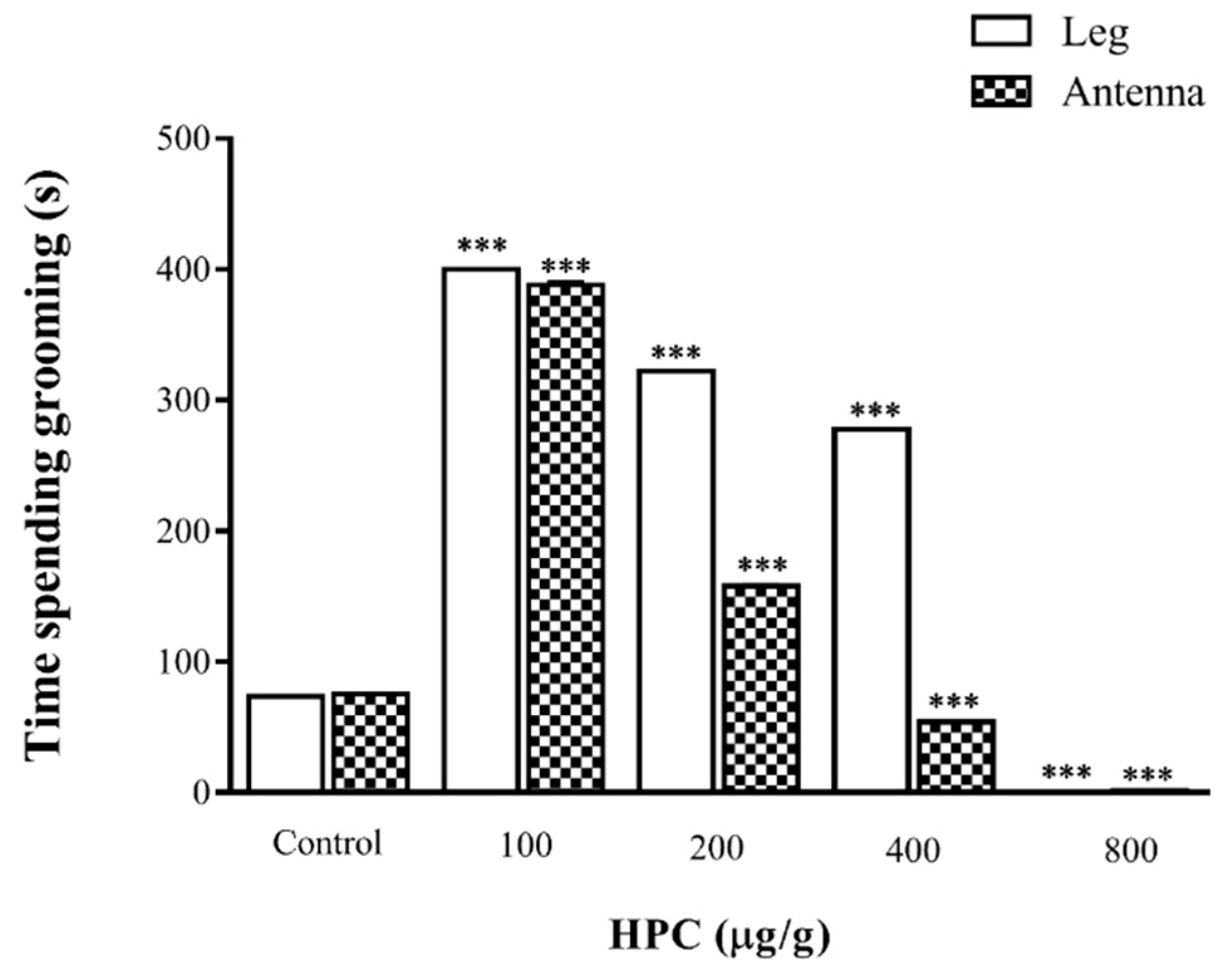
2.4. Effects of HPC on Cockroach Grooming Activity
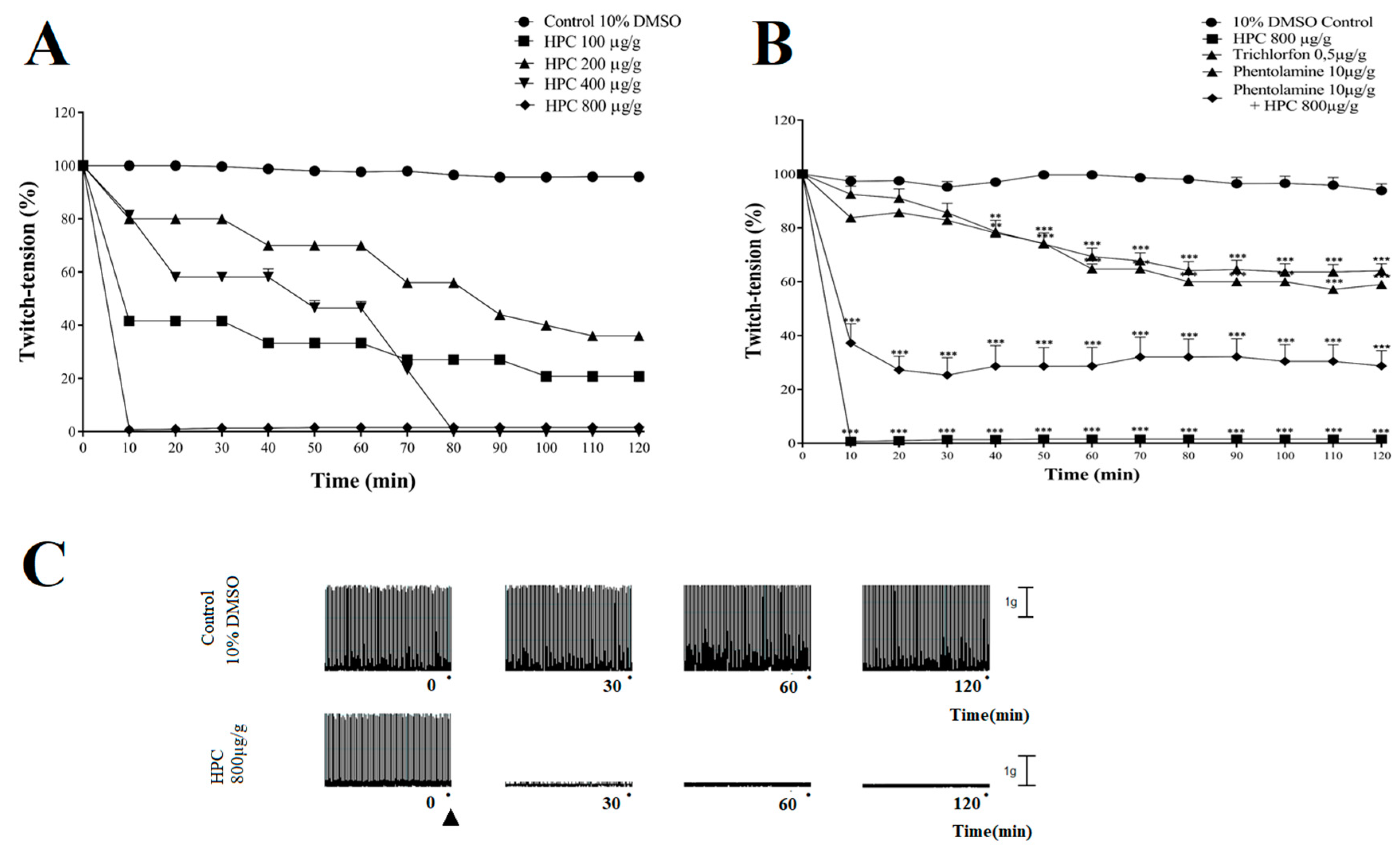
2.5. Neuromuscular Blockade Induced by HPC in N. cinerea Cockroaches
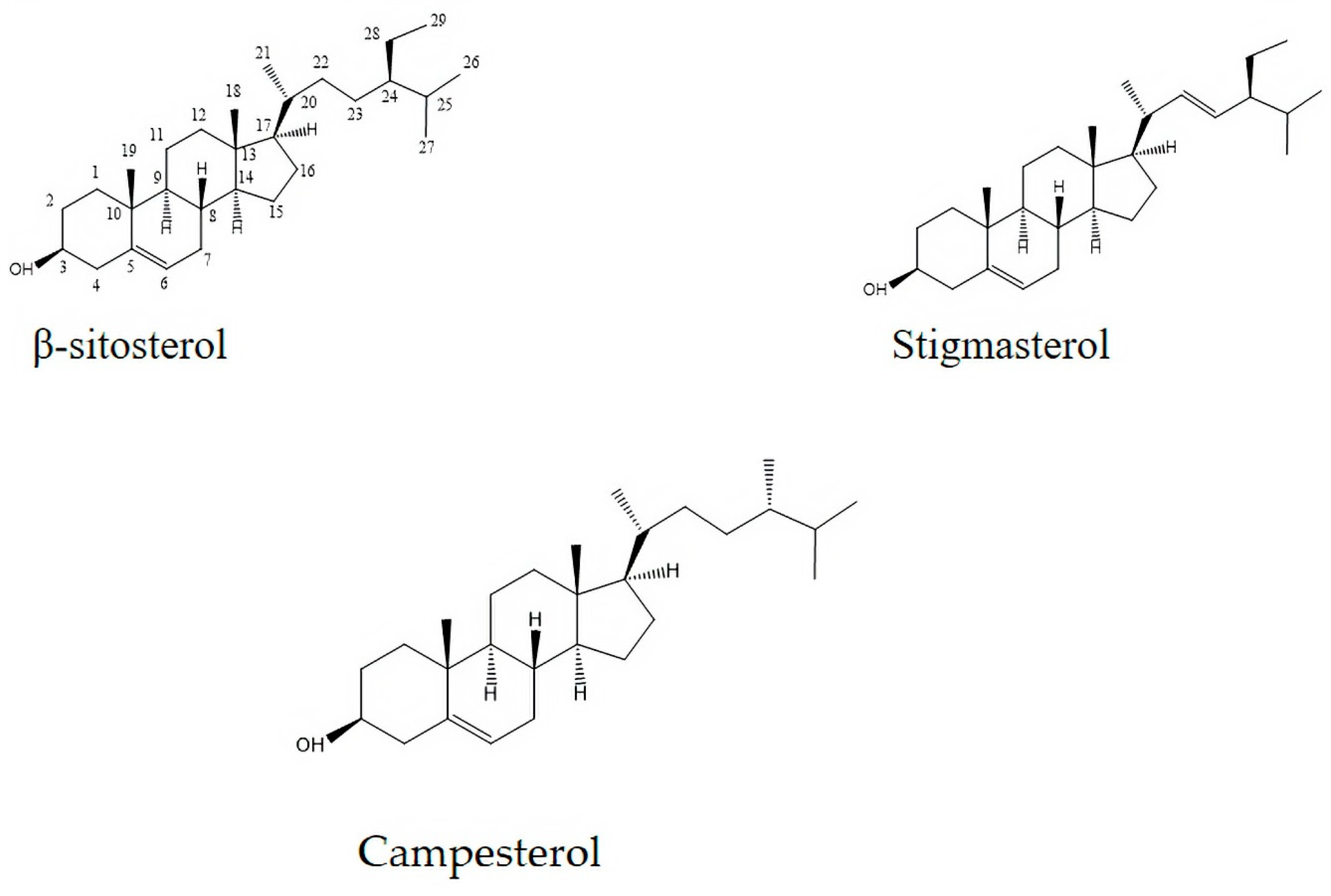
2.6. Phytosterols Identified in the Prasiola crispa n-Hexane Extract
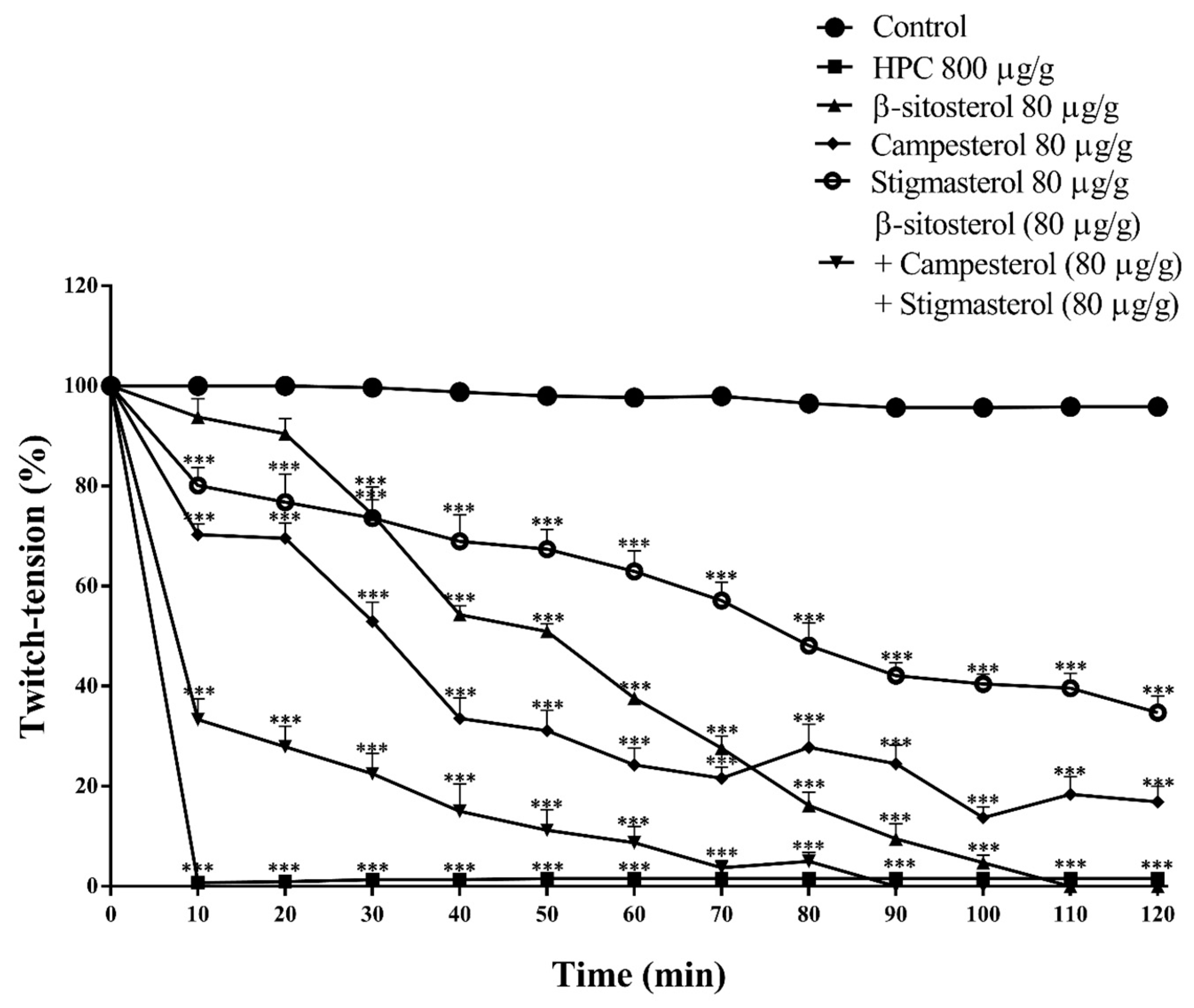
2.7. Effects of Phytosterols on Nerve-Muscle Preparation of Cockroaches
3. Discussion
4. Material and Methods
4.1. Experimental Animals
4.2. Reagents and Solutions
4.3. Algae Material
4.4. Extract Preparation
4.5. Assay for Insecticidal Activity
4.6. Assay for Insect Acetylcholinesterase Activity
4.7. Semi-Isolated Cockroach Heart Preparation
4.8. Grooming Activity
4.9. In Vivo Cockroach Metathoracic Coxal-Adductor Nerve-Muscle Preparation
4.10. Chemical Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The Antarctic—Extreme Environments—Climate Policy Watcher. Available online: https://www.climate-policy-watcher.org/extreme-environments/the-antarctic.html (accessed on 11 June 2019).
- Kejna, M. Trends of air temperature of the Antarctic during the period 1958–2000. Pol. Polar Res. 2003, 24, 99–126. [Google Scholar]
- Antarctic Environment—Australian Antarctic Division. Available online: http://www.antarctica.gov.au/about-antarctica/environment (accessed on 11 June 2019).
- Courtesy, P.; de Pesquisas Ambientais, A.; Headquarters, I.-A. Annual Activity Report; INCT-APA: Rio de Janeiro, Brazil, 2013. [Google Scholar]
- Amsler, C.D. Algal Chemical Ecology; Springer: Berlin/Heidelberg, Germany, 2008; Volume 468. [Google Scholar]
- Leal, M.C.; Munro, M.H.G.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- De Broyer, C.; Koubbi, P.; Griffiths, H.J.; Raymond, B.; Acoz, C.; Griffiths, H.; Raymond, B.; Udekem, C.; Van de Putte, A.; Danis, B.; et al. SCAR-Marine Biodiversity Information Network. In Biogeographic Atlas of the Southern Ocean; Scientific Committee on Antarctic Research: Cambridge, UK, 2014. [Google Scholar]
- Rindi, F.; McIvor, L.; Sherwood, A.R.; Friedl, T.; Guiry, M.D.; Sheath, R.G. Molecular Phylogeny of the Green Algal Order Prasiolales (Trebouxiophyceae, Chlorophyta) 1. J. Phycol. 2007, 43, 811–822. [Google Scholar] [CrossRef]
- Carvalho, E.L.; Wallau, G.L.; Rangel, D.L.; Machado, L.C.; Pereira, A.B.; Victoria, F.D.C.; Boldo, J.T.; Pinto, P.M. Phylogenetic positioning of the Antarctic alga Prasiola crispa (Trebouxiophyceae) using organellar genomes and their structural analysis. J. Phycol. 2017, 53, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, M.; Katashima, Y.; Aikawa, S.; Tanabe, Y.; Kudoh, S.; Kashino, Y.; Koike, H.; Satoh, K. Comparative Study on the Photosynthetic Properties of Prasiola (Chlorophyceae) and Nostoc (Cyanophyceae) from Antarctic and Non-Antarctic Sites 1. J. Phycol. 2010, 46, 466–476. [Google Scholar] [CrossRef]
- Kovácik, L.; Pereira, A.B. Green alga Prasiola crispa and its lichenized form Mastodia tesselata in Antarctic environment: General aspects. Beiheft. Nova Hedwigia 2001, 123, 465–478. [Google Scholar]
- Marinho RD, S.S.; Ramos CJ, B.; Leite JP, G.; Teixeira, V.L.; de Palmer Paixão IC, N.; Dal Belo, C.A.; Pereira, A.B.; Pinto, A.M.V. Antiviral activity of 7-keto-stigmasterol obtained from green Antarctic algae Prasiola crispa against equine herpesvirus 1. J. Appl. Phycol. 2017, 29, 555–562. [Google Scholar] [CrossRef]
- Zemolin, A.P.P.; Cruz, L.C.; Paula, M.T.; Pereira, B.K.; Albuquerque, M.P.; Victoria, F.C.; Pereira, A.B.; Posser, T.; Franco, J.L. Toxicity induced by Prasiola crispa to fruit fly Drosophila melanogaster and cockroach Nauphoeta cinerea: Evidence for Bioinsecticide action. J. Toxicol. Environ. Health Part A 2014, 77, 115–124. [Google Scholar] [CrossRef]
- Da Silva, A.C.R.; Pires, A.M.G.; Ramos, C.J.B.; Sanchez, E.F.; Cavalcanti, D.N.; Teixeira, V.L.; Fuly, A.L. The seaweed Prasiola crispa (Chlorophyta) neutralizes toxic effects of Bothrops jararacussu snake venom. J. Appl. Phycol. 2017, 29, 781–788. [Google Scholar] [CrossRef]
- Carrazoni, T.; de Brum Vieira, P.; da Silva, P.A. Mechanism of the Entomotoxic Activity Induced by Araucaria Angustifolia Methanolic Extract in Nauphoeta Cinerea Lobster Cockroaches. J. Bot. Res. 2017, 1, 38–49. [Google Scholar]
- Leal, A.P.; Oliveira, R.S.; Perin, A.P.A.; Borges, B.T.; de Brum Vieira, P.; dos Santos, T.G.; Vinadé, L.; Valsecchi, C.; Dal Belo, C.A. Entomotoxic activity of Rhinella icterica (Spix, 1824) toad skin secretion in Nauphoeta cinerea cockroaches: An octopamine-like modulation. Pestic. Biochem. Physiol. 2018, 148, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, D.S.; Rosa, M.E.; Zanatta, A.P.; Oliveira, R.S.; de Almeida, C.G.M.; Leal, A.P.; Sanz, M.; Fernandes, K.A.; de Souza, V.Q.; de Assis, D.R. Neurotoxic effects of sublethal concentrations of cyanobacterial extract containing anatoxin-a (s) on Nauphoeta cinerea cockroaches. Ecotoxicol. Environ. Saf. 2019, 171, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Chaubey, M.K. Role of phytoecdysteroids in insect pest management: A review. J. Agron. 2018, 17, 1–10. [Google Scholar] [CrossRef]
- Kortbeek, R.W.J.; van der Gragt, M.; Bleeker, P.M. Endogenous plant metabolites against insects. Eur. J. Plant Pathol. 2019, 154, 67–90. [Google Scholar] [CrossRef]
- Baker, B.J.; Amsler, C.D.; McClintock, J.B. Macroalgal chemical defenses in polar marine communities. In Algal Chemical Ecology; Springer: Berlin/Heidelberg, Germany, 2008; pp. 91–103. [Google Scholar]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Gade, S.; Rajamanikyam, M.; Vadlapudi, V.; Nukala, K.M.; Aluvala, R.; Giddigari, C.; Karanam, N.J.; Barua, N.C.; Pandey, R.; Upadhyayula, V.S. V Acetylcholinesterase inhibitory activity of stigmasterol & hexacosanol is responsible for larvicidal and repellent properties of Chromolaena odorata. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2017, 1861, 541–550. [Google Scholar]
- Ayaz, M.; Junaid, M.; Ullah, F.; Subhan, F.; Sadiq, A.; Ali, G.; Ovais, M.; Shahid, M.; Ahmad, A.; Wadood, A. Anti-Alzheimer’s studies on β-sitosterol isolated from Polygonum hydropiper L. Front. Pharmacol. 2017, 8, 697. [Google Scholar] [CrossRef] [PubMed]
- Elufioye, T.O.; Obuotor, E.M.; Agbedahunsi, J.M.; Adesanya, S.A. Anticholinesterase constituents from the leaves of Spondias mombin L. (Anacardiaceae). Biol. Targets Ther. 2017, 11, 107. [Google Scholar] [CrossRef]
- Wright, C.W.; Anderson, M.M.; Allen, D.; Phillipson, J.D.; Kirby, G.C.; Warhurst, D.C.; Chang, H.R. Quassinoids exhibit greater selectivity against Plasmodium falciparum than against Entamoeba histolytica, Giardia intestinalis or Toxoplasma gondii in vitro. J. Eukaryot. Microbiol. 1993, 40, 244–246. [Google Scholar] [CrossRef]
- Noriega, F.G. Juvenile hormone biosynthesis in insects: What is new, what do we know, and what questions remain? Int. Sch. Res. Not. 2014, 2014. [Google Scholar] [CrossRef]
- Rohit, M.; Ashok, T.; Vijaykumar, R.; Kashniyal, K. Molecular docking study of cassia tora, brassica campestris and calotropis procera as acetylcholinesterase inhibitor. Indian J. Pharm. Educ. 2016, 50, 116–122. [Google Scholar] [CrossRef]
- Miller, T. Response of cockroach cardiac neurons to cholinergic compounds. J. Insect Physiol. 1968, 14, 1713–1717. [Google Scholar] [CrossRef]
- Papaefthimiou, C.; Theophilidis, G. Octopamine—A single modulator with double action on the heart of two insect species (Apis mellifera macedonica and Bactrocera oleae): Acceleration vs. inhibition. J. Insect Physiol. 2011, 57, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Papaefthimiou, C.; Theophilidis, G. An in vitro method for recording the electrical activity of the isolated heart of the adult Drosophila melanogaster. Vitr. Cell. Dev. Biol. 2001, 37, 445–449. [Google Scholar] [CrossRef]
- Farooqui, T. Review of octopamine in insect nervous systems. Open Access Insect Physiol. 2012, 4, 1. [Google Scholar] [CrossRef]
- Evans, P.D.; Robb, S. Octopamine receptor subtypes and their modes of action. Neurochem. Res. 1993, 18, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.D. Multiple receptor types for octopamine in the locust. J. Physiol. 1981, 318, 99–122. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.D.; Maqueira, B. Insect octopamine receptors: A new classification scheme based on studies of cloned Drosophila G-protein coupled receptors. Invertebr. Neurosci. 2005, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Pitman, R.M. Transmitter substances in insects: A review. Comp. Gen. Pharmacol. 1971, 2, 347–371. [Google Scholar] [CrossRef]
- Rodríguez, V.; Mori, B.; Dörr, F.A.; Dal Belo, C.A.; Colepicolo, P.; Pinto, E. Effects of a cyanobacterial extract containing-anatoxin-a (s) on the cardiac rhythm of Leurolestes circunvagans. Rev. Bras. Farmacogn. 2012, 22, 775–781. [Google Scholar] [CrossRef]
- Schaefer, C.W.; Miles, J. Inhibition by acetylcholine of cockroach heart (Gromphadorhina portentosa). Comp. Gen. Pharmacol. 1970, 1, 426–430. [Google Scholar] [CrossRef]
- Carrazoni, T.; de Avila Heberle, M.; Perin, A.P.A.; Zanatta, A.P.; Rodrigues, P.V.; dos Santos, F.D.M.; de Almeida, C.G.M.; Vaz Breda, R.; dos Santos, D.S.; Pinto, P.M.; et al. Central and peripheral neurotoxicity induced by the Jack Bean Urease (JBU) in Nauphoeta cinerea cockroaches. Toxicology 2016, 368–369, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Libersat, F.; Pflueger, H.-J. Monoamines and the Orchestration of Behavior. Bioscience 2004, 54, 17–25. [Google Scholar] [CrossRef]
- Gal, R.; Libersat, F. A wasp manipulates neuronal activity in the sub-esophageal ganglion to decrease the drive for walking in its cockroach prey. PLoS ONE 2010, 5, e10019. [Google Scholar] [CrossRef] [PubMed]
- Weisel-Eichler, A.; Haspel, G.; Libersat, F. Venom of a parasitoid wasp induces prolonged grooming in the cockroach. J. Exp. Biol. 1999, 202, 957–964. [Google Scholar] [PubMed]
- Osborne, R.H. Insect neurotransmission: Neurotransmitters and their receptors. Pharmacol. Ther. 1996, 69, 117–142. [Google Scholar] [CrossRef]
- CARR, C.E.; FOURTNER, C.R. Pharmacological analysis of a monosynaptic reflex in the cockroach, Periplaneta americana. J. Exp. Biol. 1980, 86, 259–273. [Google Scholar]
- Evans, P.D. Biogenic amines in the insect nervous system. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 1980; Volume 15, pp. 317–473. ISBN 0065-2806. [Google Scholar]
- Orchard, I. Octopamine in insects: Neurotransmitter, neurohormone, and neuromodulator. Can. J. Zool. 1982, 60, 659–669. [Google Scholar] [CrossRef]
- Orchard, I.; Lange, A.B. Evidence for octopaminergic modulation of an insect visceral muscle. J. Neurobiol. 1985, 16, 171–181. [Google Scholar] [CrossRef]
- O’Shea, M.; Evans, P.D. Potentiation of neuromuscular transmission by an octopaminergic neurone in the locust. J. Exp. Biol. 1979, 79, 169–190. [Google Scholar]
- Theophilidis, G.; Burns, M.D. The innervation of the mesothoracic flexor tibiae muscle of the locust. J. Exp. Biol. 1983, 105, 373–388. [Google Scholar]
- Guixà-González, R.; Albasanz, J.L.; Rodriguez-Espigares, I.; Pastor, M.; Sanz, F.; Martí-Solano, M.; Manna, M.; Martinez-Seara, H.; Hildebrand, P.W.; Martín, M. Membrane cholesterol access into a G-protein-coupled receptor. Nat. Commun. 2017, 8, 14505. [Google Scholar] [CrossRef] [PubMed]
- Stu, G.D.; de Freitas, T.C.; de Avila Heberle, M.; de Assis, D.R.; Vinadé, L.; Pereira, A.B.; Franco, J.L.; Dal Belo, C.A. Modulation of dopaminergic neurotransmission induced by sublethal doses of the organophosphate trichlorfon in cockroaches. Ecotoxicol. Environ. Saf. 2014, 109, 56–62. [Google Scholar]
- Kagabu, S.; Murase, Y.; Imai, R.; Ito, N.; Nishimura, K. Effect of substituents at the 5-position of the pyridine ring of imidacloprid on insecticidal activity against Periplaneta americana. Pest Manag. Sci. Former. Pestic. Sci. 2007, 63, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Full, R.J.; Stokes, D.R.; Ahn, A.N.; Josephson, R.K. Energy absorption during running by leg muscles in a cockroach. J. Exp. Biol. 1998, 201, 997–1012. [Google Scholar]
- Giner, J.-L.; Zhao, H.; Beach, D.H.; Parish, E.J.; Jayasimhulu, K.; Kaneshiro, E.S. Comprehensive and definitive structural identities of Pneumocystis carinii sterols. J. Lipid Res. 2002, 43, 1114–1124. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C/H | Campesterol | β-sitosterol | Stigmasterol | |||
|---|---|---|---|---|---|---|
| C3 | 71.82 | 3.66 (1H, m) | 71.71 | 3.66 (1H, m) | 71.29 | 3.66 (1H, m) |
| C6 | 122.45 | 5.33 (1H;d;1.2) | 119.80 | 5.35 (1H, s) | 121.60 | 5.37 (1H;d; 5.33) |
| C18 | 11.96 | 0.68 (3H, s) | 11.94 | 0.68 (3H, s) | 11.21 | 0.70 (1H, s) |
| C19 | 19.21 | 1.00 (3H, s) | 18.28 | 1.00 (3H, s) | 20.80 | 1.01 (3H, s) |
| C21 | 18.61 | 0.90 (3H;d;6.5) | 18.83 | 0.92 (3H;d; 1.5) | 21.06 | 1.02 (3H, s) |
| C22 | 138.13 | 5.16 (1H;dd;15.0; 8.09) | ||||
| C23 | 129.21 | 5.16 (1H;dd;14.4; 8.09) | ||||
| C26 | 19.61 | 0.85 (3H;d;2.2) | 19.48 | 0.80 (3H;d; 3.7) | 20.40 | 0.80 (3H;d; 3.7) |
| C27 | 18.56 | 0.79 (3H;d;3.6) | 20.40 | 0.84 (3H;d;2.4) | 19.48 | 0.84 (3H;d; 2.4) |
| C28 | 14.00 | 0.77 (3H;d;3.7) | ||||
| C29 | 12.30 | 0.85 (3H;d; 2.2) | 12.20 | 0.80 (3H;d;3.7) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holken Lorensi, G.; Soares Oliveira, R.; Leal, A.P.; Zanatta, A.P.; Moreira de Almeida, C.G.; Barreto, Y.C.; Eduarda Rosa, M.; de Brum Vieira, P.; Brito Ramos, C.J.; de Carvalho Victoria, F.; et al. Entomotoxic Activity of Prasiola crispa (Antarctic Algae) in Nauphoeta cinerea Cockroaches: Identification of Main Steroidal Compounds. Mar. Drugs 2019, 17, 573. https://doi.org/10.3390/md17100573
Holken Lorensi G, Soares Oliveira R, Leal AP, Zanatta AP, Moreira de Almeida CG, Barreto YC, Eduarda Rosa M, de Brum Vieira P, Brito Ramos CJ, de Carvalho Victoria F, et al. Entomotoxic Activity of Prasiola crispa (Antarctic Algae) in Nauphoeta cinerea Cockroaches: Identification of Main Steroidal Compounds. Marine Drugs. 2019; 17(10):573. https://doi.org/10.3390/md17100573
Chicago/Turabian StyleHolken Lorensi, Graziela, Raquel Soares Oliveira, Allan P. Leal, Ana Paula Zanatta, Carlos Gabriel Moreira de Almeida, Yuri Correia Barreto, Maria Eduarda Rosa, Patrícia de Brum Vieira, Carlos José Brito Ramos, Filipe de Carvalho Victoria, and et al. 2019. "Entomotoxic Activity of Prasiola crispa (Antarctic Algae) in Nauphoeta cinerea Cockroaches: Identification of Main Steroidal Compounds" Marine Drugs 17, no. 10: 573. https://doi.org/10.3390/md17100573