Massive Gene Expansion and Sequence Diversification Is Associated with Diverse Tissue Distribution, Regulation and Antimicrobial Properties of Anti-Lipopolysaccharide Factors in Shrimp

 , and
, and
Abstract
:1. Introduction
2. Results
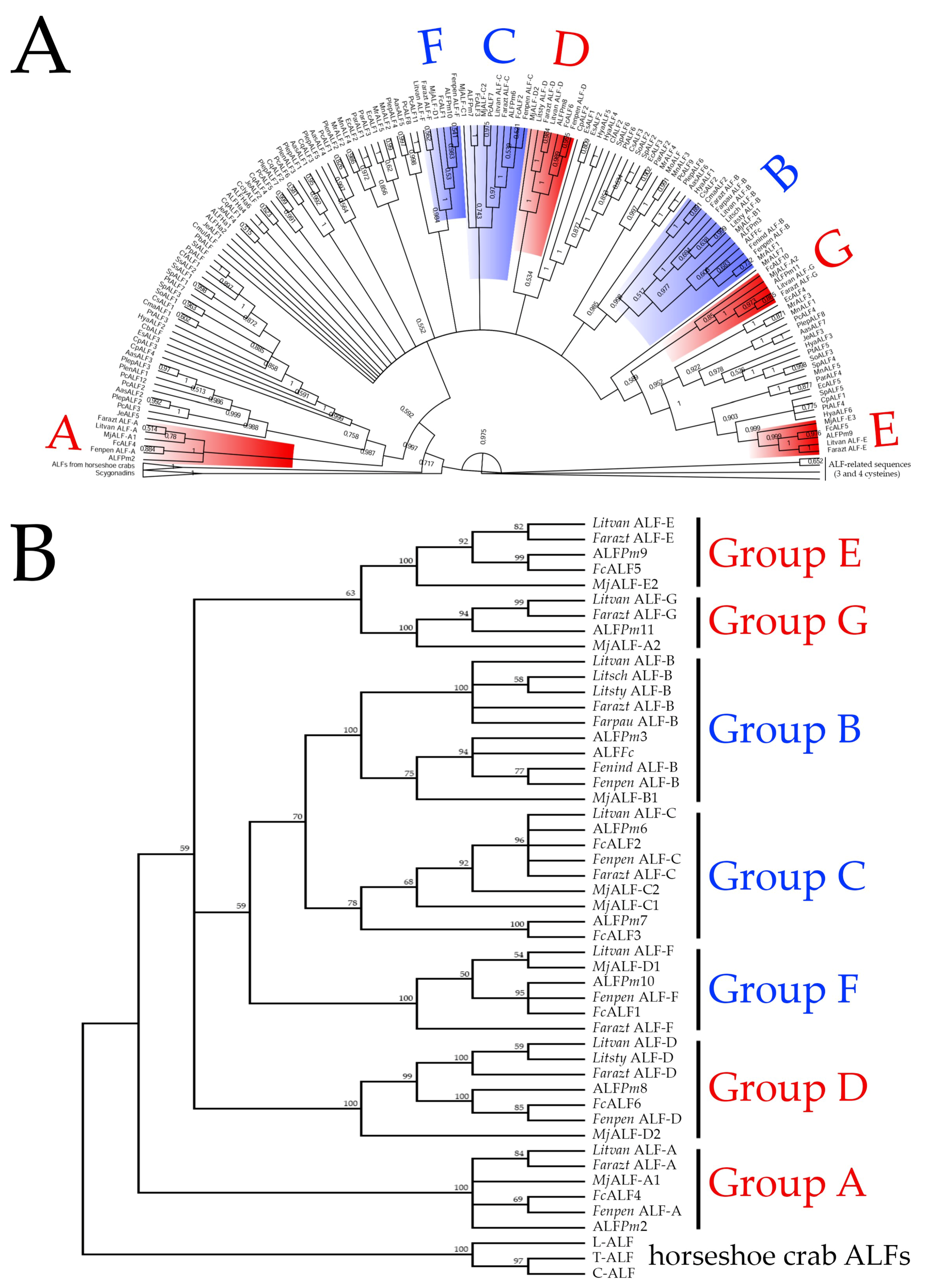
2.1. ALFs from Penaeid Shrimp Comprise a Diverse Family Composed of Seven Members
2.2. ALF Sequence Diversity Is Gene-Encoded
2.3. ALFs Evolved from Gene Duplication Events before Shrimp Speciation
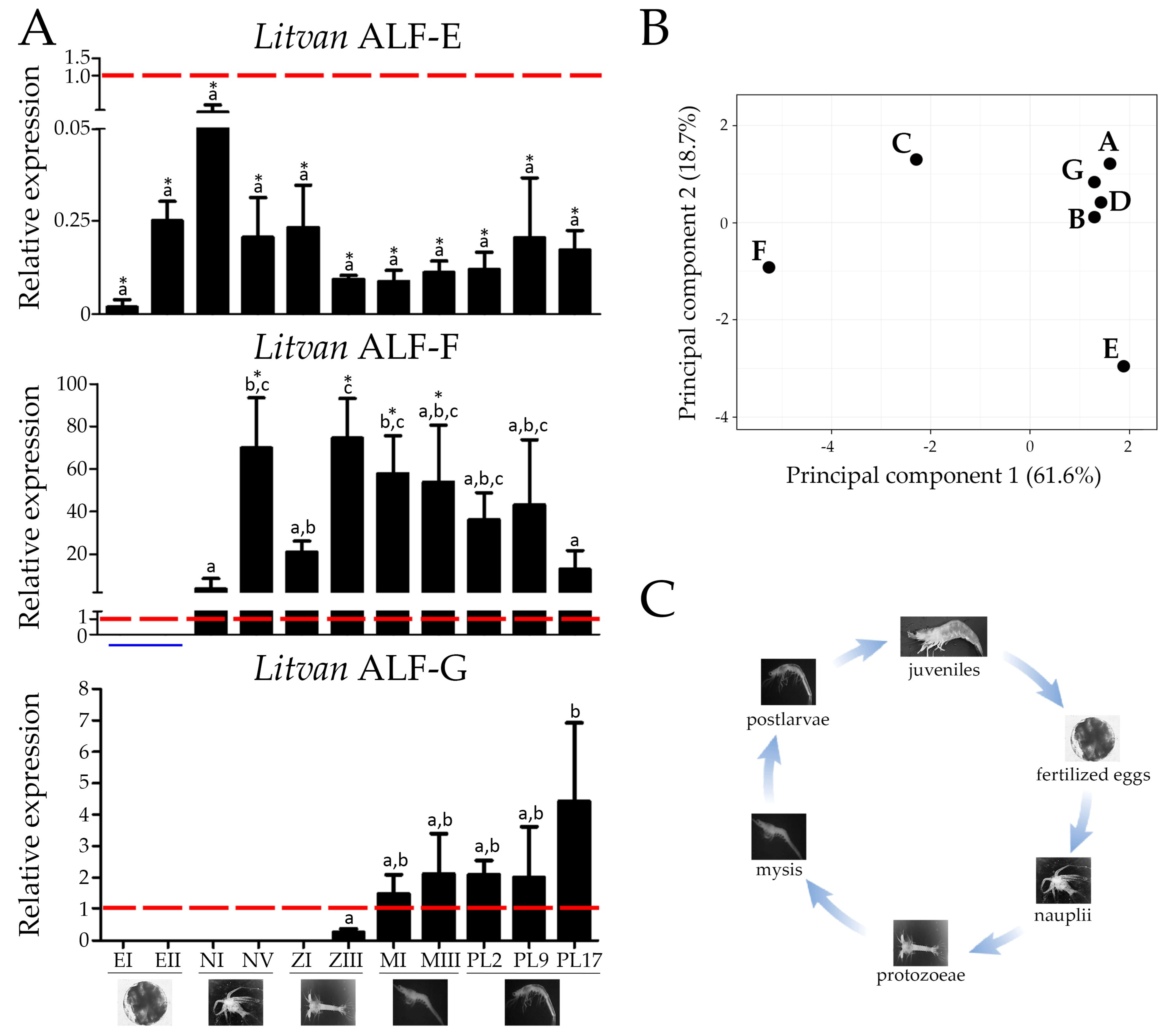
2.4. ALFs Are All Expressed in Individual Shrimps and Differentially Modulated in Response to Tissue Damage
2.5. Some ALF Genes Are Transcribed Early in Shrimp Development, while Others Are Mainly Expressed in Juveniles
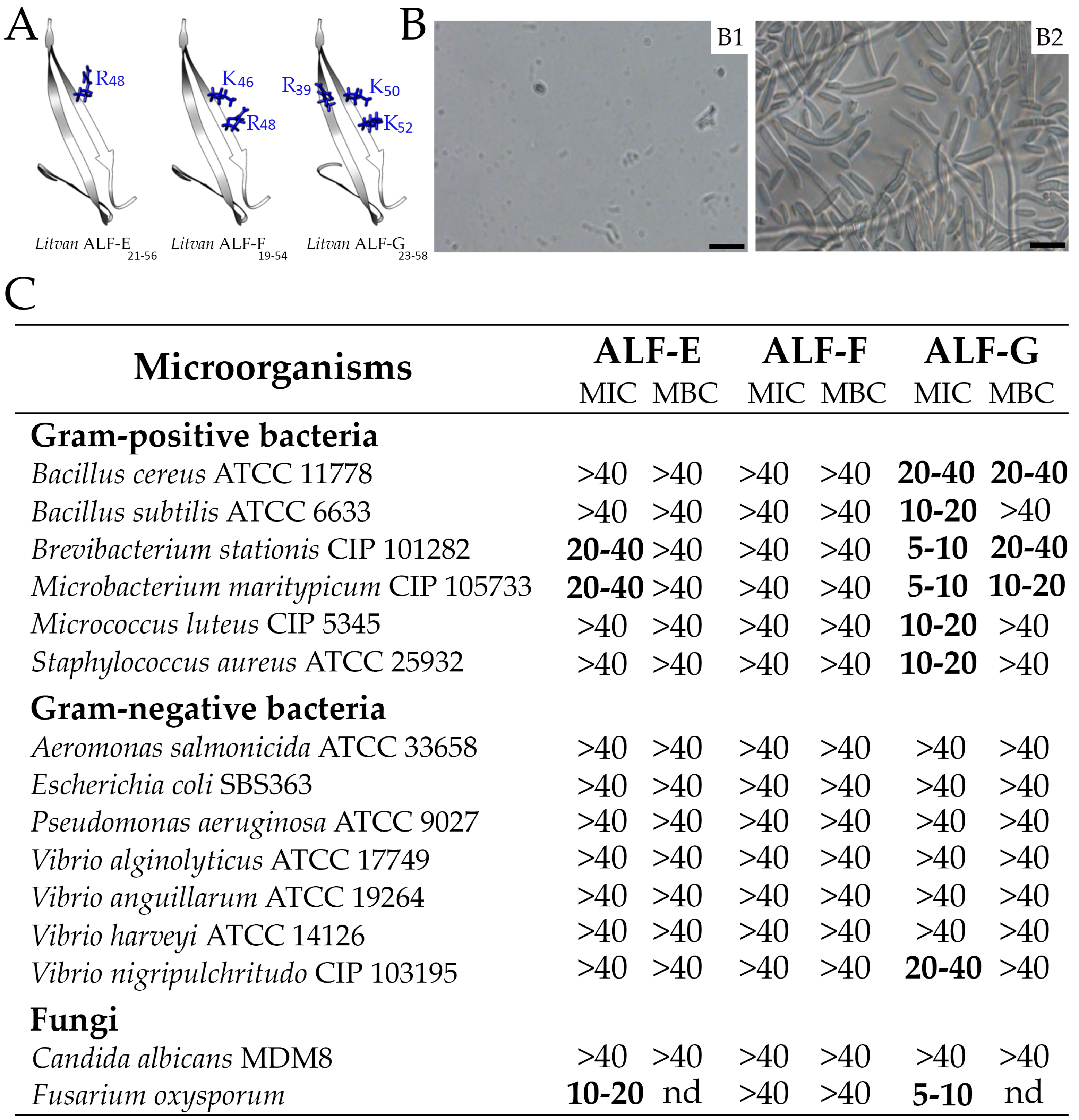
2.6. Sequence Diversity of Shrimp ALFs Results in Distinct Antimicrobial Properties
3. Discussion
4. Materials and Methods
4.1. Database Searches and Phylogenetic Reconstructions
4.2. Animals and Tissue Collection
4.3. Experimental Infections
4.4. Semiquantitative RT-PCR Analysis for Tissue Distribution of Gene Expression
4.5. Fluorescence-Based Reverse Transcription Real-Time Quantitative PCR (RT-qPCR)
4.6. Peptide Synthesis, Oxidation and Characterization
4.7. Antibacterial and Antifungal Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tanaka, S.; Nakamura, T.; Morita, T.; Iwanaga, S. Limulus anti-LPS factor: an anticoagulant which inhibits the endotoxin-mediated activation of Limulus coagulation system. Biochem. Biophys. Res. Commun. 1982, 105, 717–723. [Google Scholar] [CrossRef]
- Morita, T.; Ohtsubo, S.; Nakamura, T.; Tanaka, S.; Iwanaga, S.; Ohashi, K.; Niwa, M. Isolation and biological activities of limulus anticoagulant (anti-LPS factor) which interacts with lipopolysaccharide (LPS). J. Biochem. 1985, 97, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef]
- Supungul, P.; Klinbunga, S.; Pichyangkura, R.; Jitrapakdee, S.; Hirono, I.; Aoki, T.; Tassanakajon, A. Identification of immune-related genes in hemocytes of black tiger shrimp (Penaeus monodon). Mar. Biotechnol. 2002, 4, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: antimicrobial peptides and melanization. Dev. Comp. Immunol. 2018, 80, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Boze, H.; Chemardin, P.; Padilla, A.; Moulin, G.; Tassanakajon, A.; Pugniere, M.; Roquet, F.; Destoumieux-Garzón, D.; Gueguen, Y.; et al. NMR structure of rALF-Pm3, an anti-lipopolysaccharide factor from shrimp: Model of the possible lipid A-binding site. Biopolymers 2008, 91, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, P.; Rosa, R.D.; Destoumieux-Garzón, D. An intimate link between antimicrobial peptide sequence diversity and binding to essential components of bacterial membranes. Biochim. Biophys. Acta Biomembr. 2016, 1858, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial peptides in marine invertebrate health and disease. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150300. [Google Scholar] [CrossRef] [PubMed]
- Somboonwiwat, K.; Marcos, M.; Tassanakajon, A.; Romestand, B.; Gueguen, Y. Recombinant expression and anti-microbial activity of anti-lipopolysaccharide factor (ALF) from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2005, 29, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Vergnes, A.A.; de Lorgeril, J.; Goncalves, P.; Perazzolo, L.M.; Sauné, L.; Romestand, B.; Fievet, J.; Gueguen, Y.; Bachère, E.; et al. Functional divergence in shrimp anti-lipopolysaccharide factors (ALFs): from recognition of cell wall components to antimicrobial activity. PLoS ONE 2013, 8, e67937. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-S.; Zhang, Q.; Zhao, Y.-R.; Jia, W.-M.; Zhao, X.-F.; Wang, J.-X. A new group of anti-lipopolysaccharide factors from Marsupenaeus japonicus functions in antibacterial response. Dev. Comp. Immunol. 2015, 48, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Barracco, M.A. Antimicrobial peptides in crustaceans. Invertebr. Surviv. J. 2010, 7, 262–284. [Google Scholar]
- Pilotto, M.; Goncalves, A.; Vieira, F.; Seifert, W.; Bachère, E.; Rosa, R.; Perazzolo, L. Exploring the Impact of the Biofloc Rearing System and an Oral WSSV Challenge on the Intestinal Bacteriome of Litopenaeus vannamei. Microorganisms 2018, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, P.; Guertler, C.; Bachère, E.; de Souza, C.R.B.; Rosa, R.D.; Perazzolo, L.M. Molecular signatures at imminent death: hemocyte gene expression profiling of shrimp succumbing to viral and fungal infections. Dev. Comp. Immunol. 2014, 42, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Silveira, A.S.; Matos, G.M.; Falchetti, M.; Ribeiro, F.S.; Bressan, A.; Bachère, E.; Perazzolo, L.M.; Rosa, R.D. An immune-related gene expression atlas of the shrimp digestive system in response to two major pathogens brings insights into the involvement of hemocytes in gut immunity. Dev. Comp. Immunol. 2018, 79, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Quispe, R.L.; Justino, E.B.; Vieira, F.N.; Jaramillo, M.L.; Rosa, R.D.; Perazzolo, L.M. Transcriptional profiling of immune-related genes in Pacific white shrimp (Litopenaeus vannamei) during ontogenesis. Fish Shellfish Immunol. 2016, 58, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Guo, S.; Li, F.; Xiang, J. Functional Diversity of Anti-Lipopolysaccharide Factor Isoforms in Shrimp and Their Characters Related to Antiviral Activity. Mar. Drugs 2015, 13, 2602–2616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponprateep, S.; Tharntada, S.; Somboonwiwat, K.; Tassanakajon, A. Gene silencing reveals a crucial role for anti-lipopolysaccharide factors from Penaeus monodon in the protection against microbial infections. Fish Shellfish Immunol. 2012, 32, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchai, W.; Chaiyapechara, S.; Klinbunga, S.; Thongda, W.; Tangphatsornruang, S.; Yoocha, T.; Jarayabhand, P.; Jiravanichpaisal, P. Differentially expressed transcripts in stomach of Penaeus monodon in response to AHPND infection. Dev. Comp. Immunol. 2016, 65, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux, D.; Muñoz, M.; Cosseau, C.; Rodriguez, J.; Bulet, P.; Comps, M.; Bachère, E. Penaeidins, antimicrobial peptides with chitin-binding activity, are produced and stored in shrimp granulocytes and released after microbial challenge. J. Cell Sci. 2000, 113, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Vandenbulcke, F.; Gueguen, Y.; Bachère, E. Expression of penaeidin antimicrobial peptides in early larval stages of the shrimp Penaeus vannamei. Dev. Comp. Immunol. 2003, 27, 283–289. [Google Scholar] [CrossRef]
- Barreto, C.; Coelho, J.R.; Yuan, J.; Xiang, J.; Perazzolo, L.M.; Rosa, R.D. Specific Molecular Signatures for Type II Crustins in Penaeid Shrimp Uncovered by the Identification of Crustin-Like Antimicrobial Peptides in Litopenaeus vannamei. Mar. Drugs 2018, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Somboonwiwat, K.; Bachère, E.; Rimphanitchayakit, V.; Tassanakajon, A. Localization of anti-lipopolysaccharide factor (ALFPm3) in tissues of the black tiger shrimp, Penaeus monodon, and characterization of its binding properties. Dev. Comp. Immunol. 2008, 32, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- De la Vega, E.; O’Leary, N.; Shockey, J.E.; Robalino, J.; Payne, C.; Browdy, C.L.; Warr, G.W.; Gross, P.S.; O’Leary, N.A.; Shockey, J.E.; et al. Anti-lipopolysaccharide factor in Litopenaeus vannamei (LvALF): A broad spectrum antimicrobial peptide essential for shrimp immunity against bacterial and fungal infection. Mol. Immunol. 2008, 45, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, J.D.; Zhao, X.F.; Vasta, G.R.; Wang, J.X. A shrimp C-type lectin inhibits proliferation of the hemolymph microbiota by maintaining the expression of antimicrobial peptides. J. Biol. Chem. 2014, 289, 11779–11790. [Google Scholar] [CrossRef] [PubMed]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, C.A.; Guzmán, F.; Cárdenas, C.; Marshall, S.H.; Mercado, L. Antimicrobial activity of trout hepcidin. Fish Shellfish Immunol. 2014, 41, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Hetru, C.; Bulet, P. Strategies for the Isolation and Characterization of Antimicrobial Peptides of Invertebrates. Methods Mol. Biol. 1997, 78, 35–49. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon |
|---|---|---|---|
| LvActin 1 | TAATCCACATCTGCTGGAAGGTGG | TCACCAACTGGGATGACATGG | 846 bp |
| LvActin 2 | CCACGAGACCACCTACAAC | AGCGAGGGCAGTGATTTC | 142 bp |
| LvEF1α 2 | TGGCTGTGAACAAGATGGACA | TTGTAGCCCACCTTCTTGACG | 103 bp |
| LvL40 2 | GAGAATGTGAAGGCCAAGATC | TCAGAGAGAGTGCGACCATC | 104 bp |
| LvRpS6 2 | AGCAGATACCCTTGGTGAAG | GATGCAACCACGGACTGAC | 193 bp |
| Litvan ALF-A 1,2 | CTGATTGCTCTTGTGCCACG | TGACCCATGAACTCCACCTC | 113 bp |
| Litvan ALF-B 1,2 | GTGTCTCCGTGTTGACAAGC | ACAGCCCAACGATCTTGCTG | 123 bp |
| Litvan ALF-C 1,2 | ATGCGAGTGTCTGTCCTCAG | TGAGTTTGTTCGCGATGGCC | 115 bp |
| Litvan ALF-D 1,2 | TGTGTTGGTTGTGGCACTGG | CAACGAGGTCAATGTCACCG | 131 bp |
| Litvan ALF-E 1,2 | TGCTACGTGAATCGCAGTCC | CGCTTCCTCTTCCGACAATG | 100 bp |
| Litvan ALF-F 1,2 | AAGCTCTCATTCCTGGTCGG | GGGTGTAACGAAGTACGTGC | 180 bp |
| Litvan ALF-G 1,2 | CCGCTGCATGTCAAGTATCC | TCAGCAGTAGCAGTGTCAGC | 140 bp |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matos, G.M.; Schmitt, P.; Barreto, C.; Farias, N.D.; Toledo-Silva, G.; Guzmán, F.; Destoumieux-Garzón, D.; Perazzolo, L.M.; Rosa, R.D. Massive Gene Expansion and Sequence Diversification Is Associated with Diverse Tissue Distribution, Regulation and Antimicrobial Properties of Anti-Lipopolysaccharide Factors in Shrimp. Mar. Drugs 2018, 16, 381. https://doi.org/10.3390/md16100381
Matos GM, Schmitt P, Barreto C, Farias ND, Toledo-Silva G, Guzmán F, Destoumieux-Garzón D, Perazzolo LM, Rosa RD. Massive Gene Expansion and Sequence Diversification Is Associated with Diverse Tissue Distribution, Regulation and Antimicrobial Properties of Anti-Lipopolysaccharide Factors in Shrimp. Marine Drugs. 2018; 16(10):381. https://doi.org/10.3390/md16100381
Chicago/Turabian StyleMatos, Gabriel Machado, Paulina Schmitt, Cairé Barreto, Natanael Dantas Farias, Guilherme Toledo-Silva, Fanny Guzmán, Delphine Destoumieux-Garzón, Luciane Maria Perazzolo, and Rafael Diego Rosa. 2018. "Massive Gene Expansion and Sequence Diversification Is Associated with Diverse Tissue Distribution, Regulation and Antimicrobial Properties of Anti-Lipopolysaccharide Factors in Shrimp" Marine Drugs 16, no. 10: 381. https://doi.org/10.3390/md16100381