1. Introduction
Obesity is associated with a higher risk of developing diabetes and cardiovascular disease. At the cellular level, enlargement of the adipose tissue mass has been characterized by an increase in the size (hypertrophy) or number (hyperplasia) of adipocytes. The triglyceride (TG) content in adipocytes reflects the balance between lipogenesis and lipolysis, which is largely related to cell volume. When adipocytes reach a critical size threshold, preadipocytes in close proximity to the adipocytes will respond to positive energy balance by proliferating and then differentiating into adipocytes to store the excess energy [
1]. Early in life, adipose tissue expansion occurs primarily through hyperplasia. However, humans and rodents have the capacity to form new fat cells from preadipocytes throughout life. Several mechanisms reduce the risk of obesity, including reduced food intake, decreased intestine adsorption, suppressed lipogenesis, enhanced lipolysis and fatty acid oxidation, increased energy expenditure and inhibited preadipocyte proliferation, differentiation, and pharmacological treatment [
2,
3,
4].
Monascus species has been used as the traditional food fungus in Eastern Asia for several centuries.
Monascus-fermented products are gradually developed as the popular and important functional food for the prevention of cardiovascular disease. Red mold dioscorea (RMD) known as the
Monascus purpureus-fermented
Disocorea batatas was proven as the strong hypolipidemic functional food in the previous study [
5]. However, we found that RMD had only a weak effect on anti-obesity, which limited the development of RMD for the prevention of metabolic syndrome. Monascin and ankaflavin isolated form
Monascus-fermented product were proven to prevent obesity development via the suppressions of differentiation and lipogenesis in our
in vitro and
in vivo studies [
5,
6]. Therefore, enhancing monascin and ankaflavin levels in RMD may straighten the anti-obesity of RMD.
Deep ocean water (DOW) generally means ocean water from a depth of more than 200 m in depth. The character of DOW includes high purity, cold temperature, abundant nutrients, and minerals [
7,
8]. Currently, DOW has been applied to food, agriculture, cosmetic and medical field in many countries such as Taiwan, Japan, Korea and America. Due to its high contents of minerals such as magnesium (Mg), calcium (Ca), potassium (K), zinc (Zn),
etc. [
9,
10,
11,
12,
13]. A previous study used DOW as the culture water of
Monascus in order to straighten the hypolipidemic function [
14]. DOW-cultured RMD (DOW-RMD) using DOW as the culture water has greater effect on lowering serum total cholesterol (TC), triglyceride (TG), low density lipoprotein cholesterol (LDL-C) levels and raising high density lipoprotein cholesterol (HDL-C) levels than reverse osmosis water-cultured RMD (ROW-RMD). Furthermore, greater anti-atherosclerosis effect, and anti-fatty liver effect are performed by DOW-RMD treatment than ROW-RMD treatment group [
14].
According to above-mentioned study, RMD had strong hypolipidemic effect but not anti-obesity effects. RMD cultured in DOW contains substantial amounts of monascin and ankaflavin, and relatively low levels of citrinin. In addition, DOW enhances the production of monascin and ankaflavin. Previous research has shown that monascin and ankaflavin were the effective compounds that perform hypolipidemic and anti-obesity effects. Furthermore, DOW was previously shown to possess anti-obesity capability. Therefore, DOW and the DOW-enhanced functional metabolites in RMD may improve the anti-obesity effect. In this study, animal test was adopted to examine whether DOW-RMD has better anti-obesity effect than ultra-pure water (UPW)-cultured RMD (UPW-RMD) does. Moreover, the effect of ions composition of DOW and DOW-influenced functional metabolites change of RMD on the differentiation and lipogenesis regulation were investigated using 3T3-L1 pre-adipocytes.
3. Discussion
DOW, enriched with minerals and trace elements, has been applied in the manufacturing of various fermented food products, such as sake, soya sauce, and miso. Moreover, DOW is composed many major elements, Mg
2+, Ca
2+, K
+, Zn
2+, Fe
2+,
etc., that enhances the secondary metabolite production of
Monascus purpureus [
14]. In this study, DOW and UPW were used to culture RMD. The analytical results indicated that DOW-cultured RMD contains a relatively high level of monascin and comparatively low amount of citrinin. Monascin yield increased by 22%, and citrinin yield was reduced by 33%, suggesting that certain minerals or trace elements in DOW may enhance
Monascus to produce higher amounts of monascin and inhibit the production of citrinin [
14]. However, these salts such as NaCl, MgCl
2, KCl, and
etc. in DOW may also induce stress due to decreased water availability (water activity). The previous studies indicated that the growth and secondary metabolites formation of fungus were stimulated by the stress [
16,
17]. Furthermore, the previous studies indicated that salts have a major impact on the nature and extent of the biosphere, because solutes radically influence water activity and exert other activities that also affect biological systems (e.g., ionic, kosmotropic, chaotropic and those that affect cell turgor), and as a consequence can be major stressors of cellular systems. MgCl
2 known as a chaotropic agent disrupts the structure of macromolecules such as proteins and nucleic acids. The extreme chaotropicity of MgCl
2 at high concentrations not only denatures macromolecules, but also preserves the more stable ones: such indicator molecules, hitherto regarded as evidence of life, may thus be misleading signatures in chaotropic environments [
18,
19,
20]. Compared to DOW, UPW has a high water activity that is supra-optimal and stressful for growth and metabolism of fungus [
16,
21]. Therefore, DOW including salts may act a stressor with lower water activity that may cause up-regulation of growth, secondary metabolites production, and pharmaceutical activities [
21,
22,
23].
Regarding the reduction of body-fats, compared to UPW-RMD, DOW-RMD possessed better ability of lowering weight gain, food intake, feed efficiency, body-fat pad, cross-sectional area of adipocytes, serum TG level, and liver TC level. Previous research has confirmed that
Monascus was able to reduce body-fats; however, the ability of RMD to reduce body-fat was not significant [
24]. In this study, DOW-RMD, but not UPW-RMD, demonstrated the ability to significantly reduce weight gain, body-fat pad, and the cross-sectional area of adipocytes. This result suggested that DOW use in RMD fermentation can enhance the anti-obesity effect of RMD.
Numerous factors explain the reason that DOW contributed higher anti-obesity to DOW-RMD, one of which is that the water absorbed during the fermentation is retained in RMD, which therefore improved the anti-obesity function in DOW-RMD. According to a previous study, drinking DOW reduces the PPAR-γ expression in adipocytes [
12]. Furthermore, in an
in vitro cell test, Hwang
et al. (2009) [
25] investigated the mechanism of how DOW influences 3T3-L1 pre-adipocytes. The results verified that DOW effectively inhibited the proliferation of pre-adipocytes and significantly reduced the accumulation of lipid droplets. Moreover, DOW was able to reduce the regulation of adipocyte transcription factors; consequently, differentiation of pre-adipocytes was inhibited.
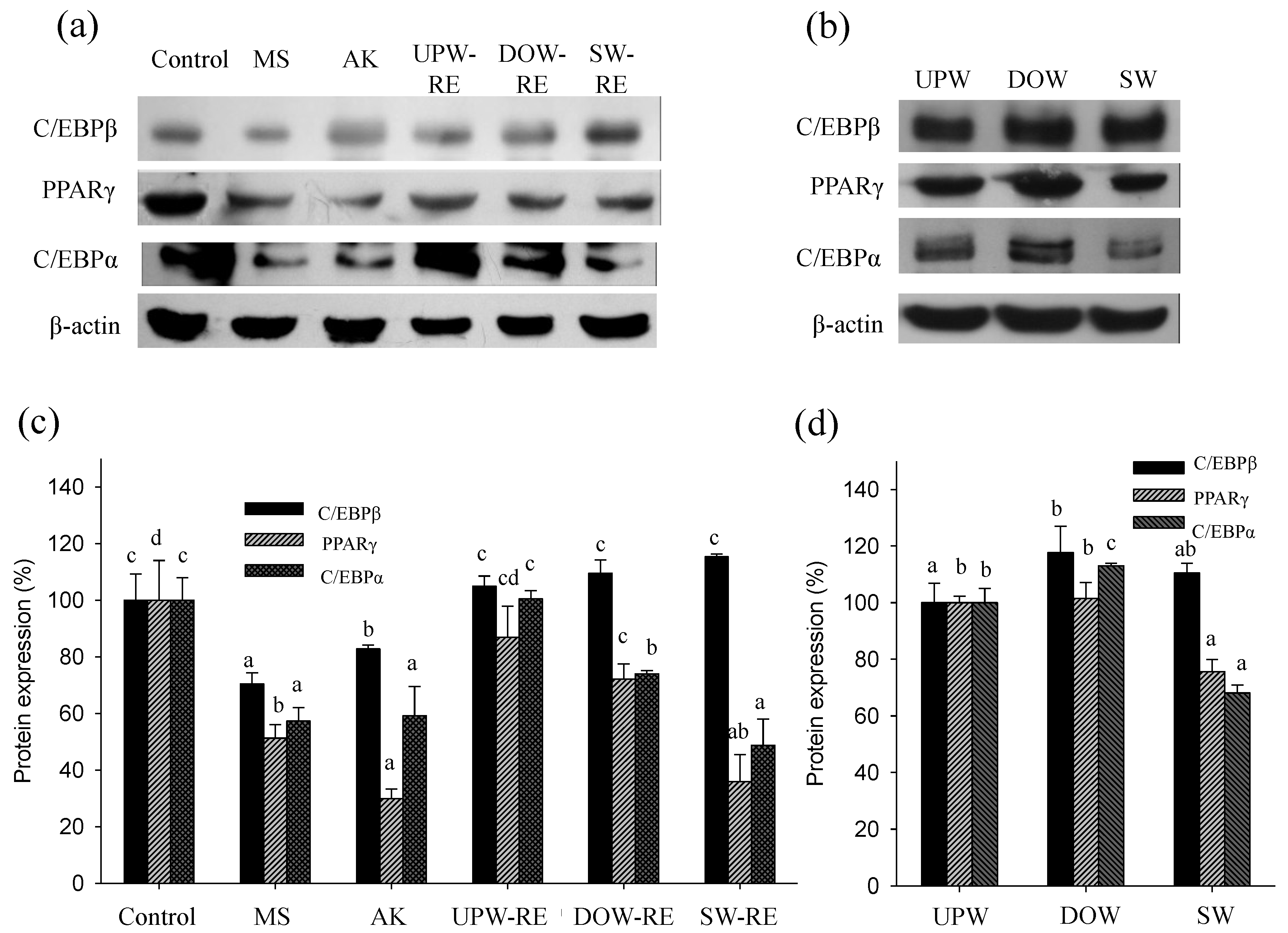
The other factor may be attributed to the DOW-stimulated functional metabolites production in the RMD. In the previous study, monascin and ankaflavin were observed to inhibit differentiation of 3T3-L1 pre-adipocytes. Specifically, the expressions of C/EBPα, C/EBPβ, C/EBPδ, and PPARγ proteins were inhibited, which also reduced the accumulation of intracellular TG. Moreover, monascin and ankaflavin enhanced the decomposition of intracellular lipid droplets in mature adipocytes lipogenesis to release glycerol and reduce extracellular LPL activity, which subsequently reduces intracellular TG synthesis [
5]. In addition to inhibiting mature adipocyte lipogenesis by lowering HR-LPL activity and increasing the lipolysis effect, monascin and ankaflavin were also observed to inhibit pre-adipocyte differentiation in animals by inhibiting the expression of C/EBPα, C/EBPβ, and PPARγ proteins [
5].
In the
in vivo animal test, the results showed that DOW-RMD demonstrated better anti-obesity ability. The ions in DOW may influence the regulation of adipocytes. Therefore, we further simulated SW that contains the major ions in a DOW. In metabolite production, DOW-RMD and SW-RMD are RMDs cultured in DOW and SW. The two products contained similar amount of monascin, but the ankaflavin level in SW-RMD was less than that in DOW-RMD. Thus, the ions in SW are crucial to enhancing monascin production; however, other effective ions in DOW may be present to facilitate an increase in ankaflavin. The levels of monascin and ankaflavin may influence the anti-obesity ability such as differentiation and lipogenesis. The
in vitro results showed that the inhibitory effect of DOW-RE and SW-RE on PPARγ and C/EBPα protein expressions was substantially better than that of UPW-RE. The inhibitory effect of monascin and ankaflavin on PPARγ and C/EBPα expressions was also evident. Therefore, DOW-RE and SW-RE, compared to UPW-RE, were able to significantly inhibit differentiation possibly because of the relatively high levels of monascin and ankaflavin. Although previous studies have shown that DOW inhibited the expression of C/EBPβ, PPARγ, and C/EBPα [
25], the inhibitory effect of DOW on transcription factor expressions was not identified in this study. However, the six ions in SW possessed the ability to inhibit PPARγ and C/EBPα expressions, indicating that these ions are effective in inhibiting differentiation. Besides the six ions contained in DOW, additional ions may exist in small quantities. These additional ions may stimulate PPARγ and C/EBPα expressions. Consequently, although SW-RE contains fewer amounts of monascin and ankaflavin than DOW-RE, its ability to inhibit PPARγ and C/EBPα expressions is superior to DOW-RE. This phenomenon suggests that SW-RMD may have absorbed the ions in SW, which subsequently increased the inhibitory effect on PPARγ and C/EBPα expressions.
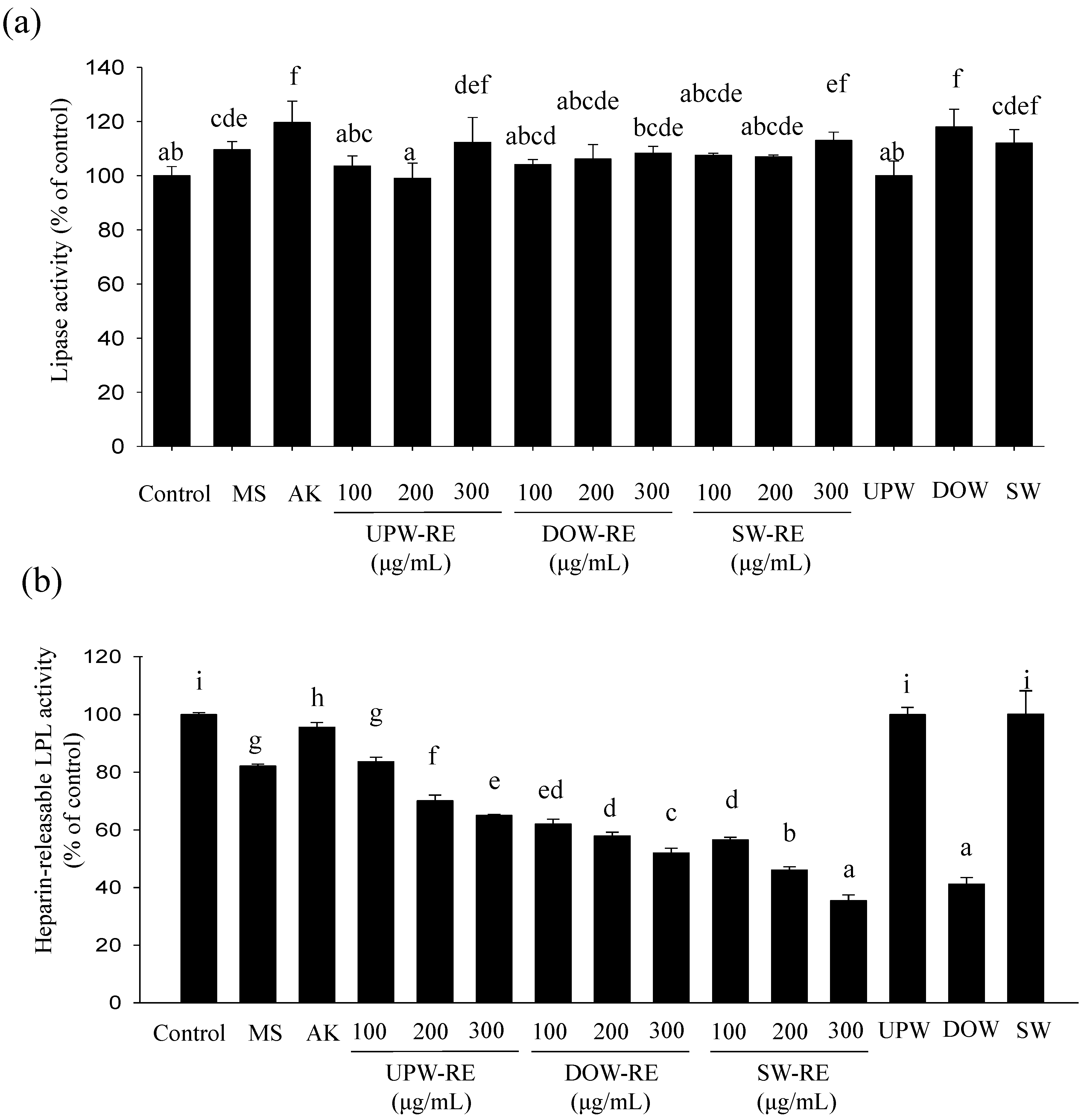
In the process of forming lipid droplets in mature adipocytes, the results showed that the lipolysis effect of RMD extract improves with increasing concentration to inhibit lipogenesis. The lipolysis effect of UPW-RE, DOW-RE, and SW-RE demonstrated no significant differences. This result was similar to that obtained in animal testing, where no significant differences were observed between the lipolysis effect of DOW-RMD and UPW-RMD. This observation showed that DOW cannot significantly enhance the lipolysis effect in RMD. This research and previous studies have verified that the lipolysis ability of monascin and ankaflavin [
6]. However, increases in monascin and ankaflavin cannot reflect the enhancement of the lipolysis effect in DOW-RMD in this study, possibly because insufficient amounts of monascin and ankaflavin were available to perform lipolysis effect or because monascin and ankaflavin have reached maximum utility that increasing the level does not significantly influence the lipolysis effect. In this study, we identified that significant lipolysis effect was performed by DOW treatment but not DOW-RMD because the amount of DOW absorbed in DOW-RMD may be not more enough to achieve the lipolysis effect.
In this study, as the concentration increased, the RMD extract effectively inhibited LPL activity, thereby inhibiting lipid droplet formation. Compared to the UPW-RE, the DOW-RE was shown to perform a better inhibitory effect on LPL activity. This result corresponded to that of the animal testing, in which DOW-RMD presented a better inhibitory effect on LPL activity. LPL expression is mediated by the activation of PPARγ by cognate ligands, as LPL is a downstream gene of PPARγ. The PPARγ/RXR complex binds to the PPRE present in the promoter region of the LPL gene and increases LPL gene expression [
26]. The induction of lipoprotein lipase synthesis by PPARγ is mainly in the mature adipocytes in order to increase local generation of free fatty acids [
27,
28]. However, monascin is proven to inhibit PPARγ expression and HR-LPL activity in this study and our previous study [
6]. Therefore, this could be one of the primary factors of monascin- and ankaflavin-mediated inhibition of lipogenesis. According to the
in vivo and
in vitro tests results, more potent LPL activity may be contributed from the higher monascin levels, as well as the absorption of DOW in DOW-RMD. Furthermore, SW-RE but not SW demonstrated an inhibitory effect on LPL activity, suggesting that the LPL activity-lowering effect of SW-RE should be contributed form the increased monascin levels or other compound but not the ions of SW. However, besides the six ions in SW, additional ions in DOW may be the functional ions for the LPL activity-lowering effect.
SW contains Mg2+, Ca2+, Na+, K+, Fe2+, and Zn2+, all of which are the main ions in DOW. In addition, the concentrations of these ions in SW were adjusted to that in DOW. Although SW contains fewer types of ions than DOW (which comprises more than ten types of ions), it still enhanced the rate of monascin and ankaflavin production in RMD. However, the ability of SW to increase the ankaflavin level is less effective than that of DOW. Despite having several similarities with DOW in monascin production, the ability of DOW and SW in the regulation of adipogenesis exhibits numerous differences. SW is able to inhibit PPARγ and C/EBPα expressions and is able to slightly increase lipolysis effect but not inhibit the HR-LPL activity. Conversely, DOW cannot inhibit PPARγ and C/EBPα expressions but is able to enhance the lipolysis effect and inhibit HR-LPL activity. However, both DOW-RMD and SW-RMD are able to inhibit PPARγ and C/EBPα expressions and improve HR-LPL activity because of the different reason as follow: DOW-RMD contains higher monascin and ankaflavin levels as well as DOW accumulation; and SW-RMD contains higher level of monascin and accumulates six types of effective ions.
Regarding the reduction of blood lipids, in comparison to UPW-RMD, DOW-RMD was able to reduce the level of TC and LDL-C in the serum. This result corresponded to that obtained in our previous results [
14]. DOW-RMD performed better blood lipid-reduction effect because it contained higher levels of blood lipid-reducing substances monascin and ankaflavin. Furthermore, DOW comprised minerals and trace elements such as Mg
2+, Ca
2+, and K
+. Numerous studies have indicated that a higher Mg/Ca ratio facilitates the prevention of cardiovascular disease [
29]. Cohen
et al. (2002) showed that an intake of 8.3 g Mg salt per day significantly reduced TC and TG levels in the blood [
29]. Shahkhalili
et al. (2001) [
30] identified that supplementing participants with 0.9 mg/day Ca reduced the level of LDL-C in the blood. DOW-RMD was used as the test substance in this study. DOW including Mg
2+, Ca
2+, Zn
2+, Fe
2+, K
+, and
etc. was supplemented daily to culture RMD, accumulated gradually in DOW-RMD; thus, the hypolipidemic effect of was enhanced.
4. Experimental Section
4.1. Chemicals
LC grade acetonitrile, chloroform, methanol, and dimethyl sulfoxide (DMSO) were purchased from Merck Co. (Darmstadat, Germany). Tryptone, yeast extract, peptone, malt extract, potato dextrose agar (PDA), and Bacto-agar were purchased from Difco Co. (Detroit, MI, USA). Monoclonal C/EBPα antibody was purchased from GeneTex Co (Irvine, CA, USA). Monoclonal C/EBPβ antibody and polyclonal PPARγ antibody were purchased from Novus Biological (Littleton, CO, USA). Dulbecco’s modified Eagle’s medium and fetal bovine serum were purchased from Invitrogen Life Technologies (Carlsbad, CA, USA). Dexamethasone, isobutylmethylxanthine, insulin, oil-red O, heparin, p-nitrophenyl butyrate were purchased from Sigma Chemical Co. (St Louis, MO, USA). Trypan blue stain was purchased from Gibco BRL Life Technologies Inc. (Gaithersburg, MD, USA).
4.2. The Source of DOW and the Preparation of SW
The DOW purchased from the Taiwan Yes Deep Ocean Water Co. (Hualien, Taiwan) was pumped from a depth of 670 m in the Pacific Ocean near the Eastern Taiwan and processed though the electrodeionization. The concentrations of the trace elements and minerals in DOW including Al, Cu, Zn, As, Ba, Cd, Cr, Pb, Hg, Se, Ag, Ca, Mg, K, Na, Sb, Tl, Be, Fluoride, Nitrate, Sulfate, Chloramines, and Chlorine have been measured and published in our previous study [
14]. SW was prepared by mixing the main ions of DOW including 20.65 mg/L Mg, 5.02 mg/L Ca, 7.71 mg/L Na, 0.22 mg/L K, 0.0062 mg/L Fe, and 0.019 mg/L Zn ion with equal concentrations to that in DOW.
4.3. Preparation of UPW-RMD, DOW-RMD, and SW-RMD
Monascus purpureus NTU 568 fermented product has been proven to perform a potent hypolipidemic effect in our previous study [
31,
32]. The culture strain was maintained on PDA slant at 10 °C and transferred monthly. The
Dioscorea root (
Dioscorea batatas Dence) purchased from a local supermarket in Taiwan was used to produce RMD using the method of solid-state culture. UPW, DOW, and SW were used as all of the water used in the production of UPW-RMD, DOW-RMD, and SW-RMD, respectively. Five hundred grams
Dioscorea substrates were, respectively, soaked in distilled water for 8 and 1 h. After that, excess water was removed with a sieve. The substrate was autoclaved for 20 min at 121 °C in a “koji-dish” (the koji-dish is made of wood with the dimension of 30 × 20 × 5 cm). After being cooled, the substrate was inoculated with a 5% (v/w) spore suspension (10
7 spores/mL). The inoculated substrate was cultured at 28 °C for 10 days. In addition, during the culturing stage, 100 mL of water is added once every 12 h at a total of three times and the addition of water starts on the fifth day of culture. After fermentation, the crushed and dried product with the mold was used for the experiments [
14,
33].
4.4. Animal Experiments
Animal experiments protocol is refer to our previous study involved the anti-obesity evaluation of RMR [
6]. Male Sprague Dawley (SD) rats at 6–8 weeks of age were purchased from the BioLasco Co. (Taipei, Taiwan). The animals were housed individually and allowed free access to a standard laboratory chow (Ralston Purina, St Louis, MO, USA) and water. Three weeks later, the rats were randomly assigned to one of the following diets for 8 weeks: standard chow (control group, NOR; 4.5% fat, 3.34 kcal/g), high-fat (HF) diet consisting of 26.7% butter powder (Gene Asia Biotech Co., Ltd., Nang-Tou, Taiwan) in standard chow (HF group; 30% fat, 4.17 kcal/g), HF diet plus 27.81 mg/day 100 g bw UPW-RMD powder (UPW-R-1X group), HF diet plus 27.81 mg/day 100 g bw DOW-RMD powder (DOW-R-1X group), HF diet plus 55.62 mg/day 100 g bw DOW-RMD powder (DOW-R-2X, group), HF diet plus 0.152 mg/day mg/day 100 g bw monascin (MS group), HF diet plus 0.162 mg/day 100 g bw ankaflavin (AK group), The recommendation dosage of DOW-RMD or UPW-RMD for anti-obesity effect is suggested as 2 g/day for human in our previous study [
24]. The dosage of monascin and ankaflavin in the MS and AK groups were equal to that in the DOW-R-1X group. The doses of the test substances used in this study were calculated according to Boyd’s formula for body surface area for adult humans (weight: 65 kg; height: 170 cm). Each sample was orally administrated to the rats by stomach tube in each group [
24].
Food consumption and body weight were recorded weekly. At the end of the study, the rats were deprived of food for 16 h before being scarified by CO2 asphyxiation. Blood samples were collected from the posterior vena cava and centrifuged at 700× g for 10 min; the serum was stored at −20 °C until analyzed. Perirenal and epididymal fat pads were removed and weighed. Portions of the adipose tissue were immersed in 10% formaldehyde for histological inspection; other portions were frozen immediately in liquid nitrogen and stored at −80 °C for analysis of lipolysis and HR-LPL activity. Liver was excised and stored at −20 °C for the measurement of lipids. The experiment was reviewed and approved by the Animal Care and Research Ethics Committee of the National Taitung University.
4.5. Biochemical Analyse
The serum total cholesterol (TC), triglyceride (TG), low density lipoprotein cholesterol (LDL-C), high density lipoprotein cholesterol (HDL-C), creatinine, uric acid, Na, K, ketone body (hydroxybutyrate) concentrations, and aspartate aminotransferase (AST), alanine aminotransferase (ALT), and creatine kinase (CK) activities were measured using the commercial kits (Randox Laboratories Ltd., Antrim, UK). Lipolysis effect and HR-LPL activity assay followed the method of our previous studies [
6].
4.6. Adipose Tissue Histology
The adipose tissue samples were fixed in formaldehyde, embedded in paraffin, cut into 5-mm sections and stained with hematoxylin and eosin. Cross-sectional areas of the adipocytes were calculated from the histogram according to Chen and Farese [
34]. For the estimation of fat pads cell number, the lipid content of 0.3 g of fat tissue was extracted by using the method of Folch
et al. [
35]. The total cell number in the fat pads was calculated by dividing the lipid content of the fat pad by the mean weight of cell lipids. The lipid weight of the average fat cell was calculated from the mean cell volume, assuming a lipid density of 0.915 (density of triolein).
4.7. Cell Culture
3T3-L1 preadipocytes purchased from the Bioresource Collection and Research Center (Hsinchu, Taiwan) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum) at 37 °C in 5% CO2. To induce differentiation, 2-day postconfluent 3T3-L1 preadipocytes (day 0) were stimulated for 48 h with 0.5 mM isobutylmethylxanthine, 1 mM dexamethasone and 10 mg/mL insulin (MDI) added to basal medium. On day 2, the MDI medium was replaced with basal medium containing insulin only. On day 4 and thereafter, the cells were cultured in basal medium, which was freshly changed every 2 days until the cells were analyzed.
DOW-RE, SW-RE, and UPW-RE were prepared using the extraction of DOW-RMD, SW-RMD, and UPW-RMD with 10-fold volume of 95% ethanol at 37 °C for 24 h, respectively. DOW-RE, SW-RE, and UPW-RE were diluted to various concentrations with DMEM medium, and further used as the treatment medium in the cell experiments. The vehicle control was 0.3% ethanol in culture medium, which was equal to the ethanol concentration in all RMD extract treatments.
4.8. Oil-Red O Staining
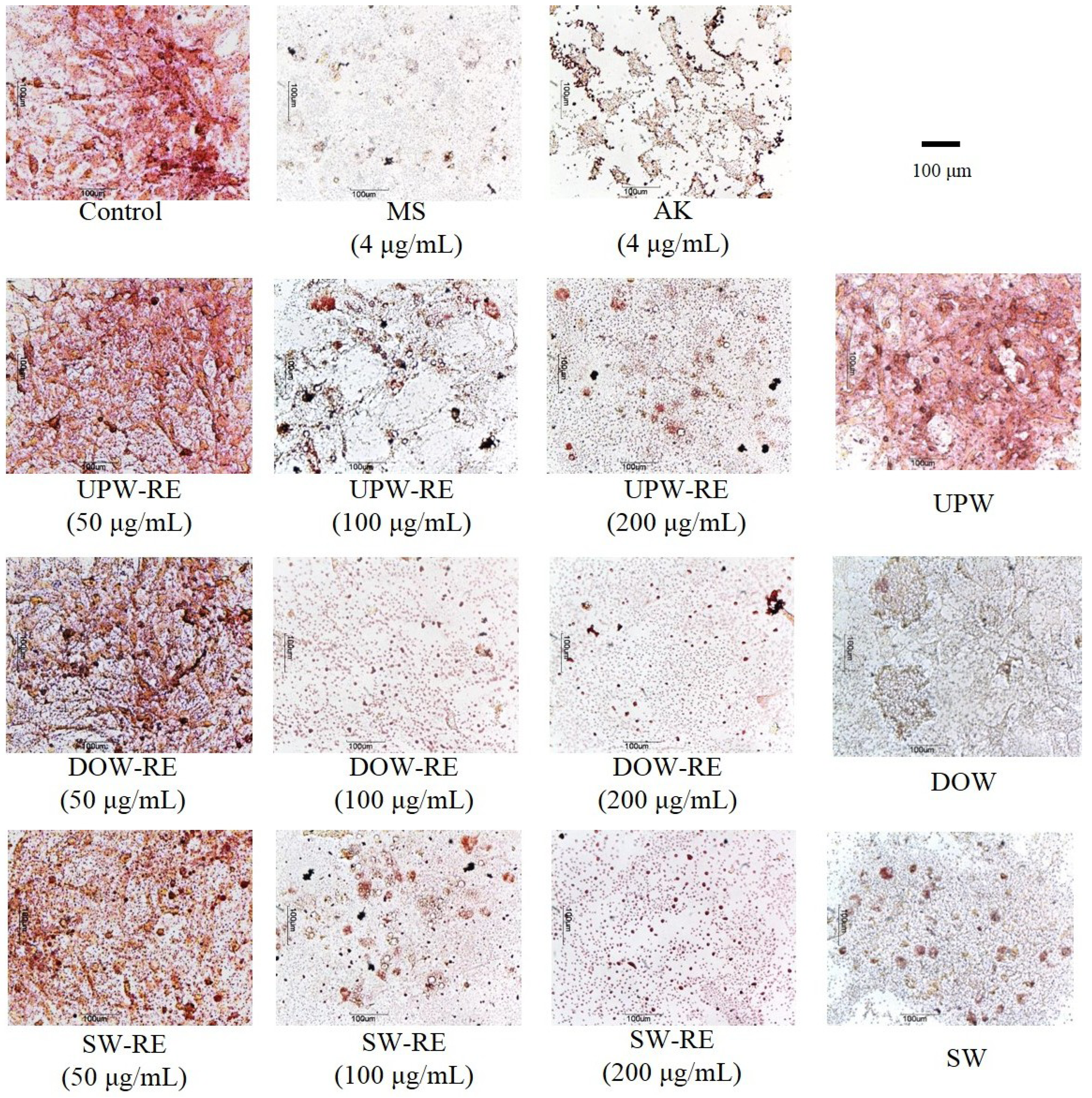
Differentiated 3T3-L1 cells on day 8 were fixed with 10% formaldehyde and then stained with oil-red O. Pictures were taken using a microscope (ECLIPSE TS100; Nikon Co., Tokyo, Japan) [
36].
4.9. Lipolysis Assay
The fully differentiated 3T3-L1 adipocytes (days 8–12 after differentiation induction) were treated with test substances in Krebs Ringer bicarbonate (KRB) buffer (20 mM NaCl, 4.7 mM KCl, 2.2 mM CaCl
2, 1.2 mM MgSO
4 7H
2O, 1.2 mM KH
2PO
4, 25 mM NaHCO
3 and 2% BSA; pH 7.4) for 24 h. Adipose explants (0.1 g) of perirenal and epididymal fat pads from experimental rats were incubated in 1 mL of KRB buffer at 37 °C for 1 h [
37]. Glycerol was determined enzymatically from the supernatant by using a Randox kit.
4.10. Heparin-Releasable Lipoprotein Lipase (HR-LPL) Activity Assay
After incubation of the 3T3-L1 mature adipocytes with the experimental medium for 24 h, the medium was discarded. The cells were rinsed with KRB buffer and then cultured in heparin-KRB (10 U/mL heparin) at 37 °C for 1 h. The conditioned heparin-KRB was collected from each well for the assay of HR-LPL activity. In the animal study, a sample of perirenal and epididymal adipose tissue weighing 0.1 g was placed in 1 ml of KRB buffer supplemented with 10 U/mL heparin at 37 °C for 1 h. LPL activity was measured according to the previous study on the basis of its esterase property using
p-nitrophenyl butyrate as a substrate [
36].
The TG hydrolase activity of LPL with synthetic TG substrates is inhibited by molar sodium chloride, and this property has been used to distinguish LPL activity from the activities of other lipases in plasma. Thus, HR-LPL activity was calculated from the productivity of
p-nitrophenol using the following equation [
36].
where
A400 (0.15 M NaCl) and
A400 (1 M NaCl) were the absorbances of released
p-nitrophenol at 400 nm in 0.15 M and in 1 M NaCl assay buffer, respectively, and 0.012 is the micromolar extinction coefficient of
p-nitrophenol.
4.11. Immunoblotting
Protein concentration was determined by bicinchoninic acid (BCA) method. A total of 40 μg of total protein from each sample was applied for Western blot representative of three independent experiments according to the previous studies [
38,
39]. The samples were separated on 10% SDS-PAGE gels and transferred to polyvinylidene fluoride membranes. After blocking in a gelatin-NET solution, blots were incubated with monoclonal C/EBPα antibody (1:5000), monoclonal C/EBPβ antibody (1:2000), polyclonal PPARγ antibody (1:1000), polyclonal PPARγ antibody (1:1000) at room temperature for 1 h. Then, bands were incubated with specific horse radish peroxidase (HRP)-conjugated secondary antibodies (1:100,000) at room temperature for 1 h and visualized by enhanced chemiluminescence (ECL) substrate with UVP AutoChemi Image system (UVP Inc., Upland, CA, USA). Protein loading was evaluated by anti-actin antibody (1:5000).
4.12. Statistics
Data are expressed as means ± standard deviation. Analysis of variance by Duncan’s test and Pearson’s product-moment correlation coefficient test were determined using SPSS version 10.0 software (SPSS Institute, Inc., Chicago, IL, USA). Differences with p < 0.05 were considered statistically significant.




{kind=link}
{kind=link}
{kind=link}


