
Comparison of the Effect of BPA and Related Bisphenols on Membrane Integrity, Mitochondrial Activity, and Steroidogenesis of H295R Cells In Vitro
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
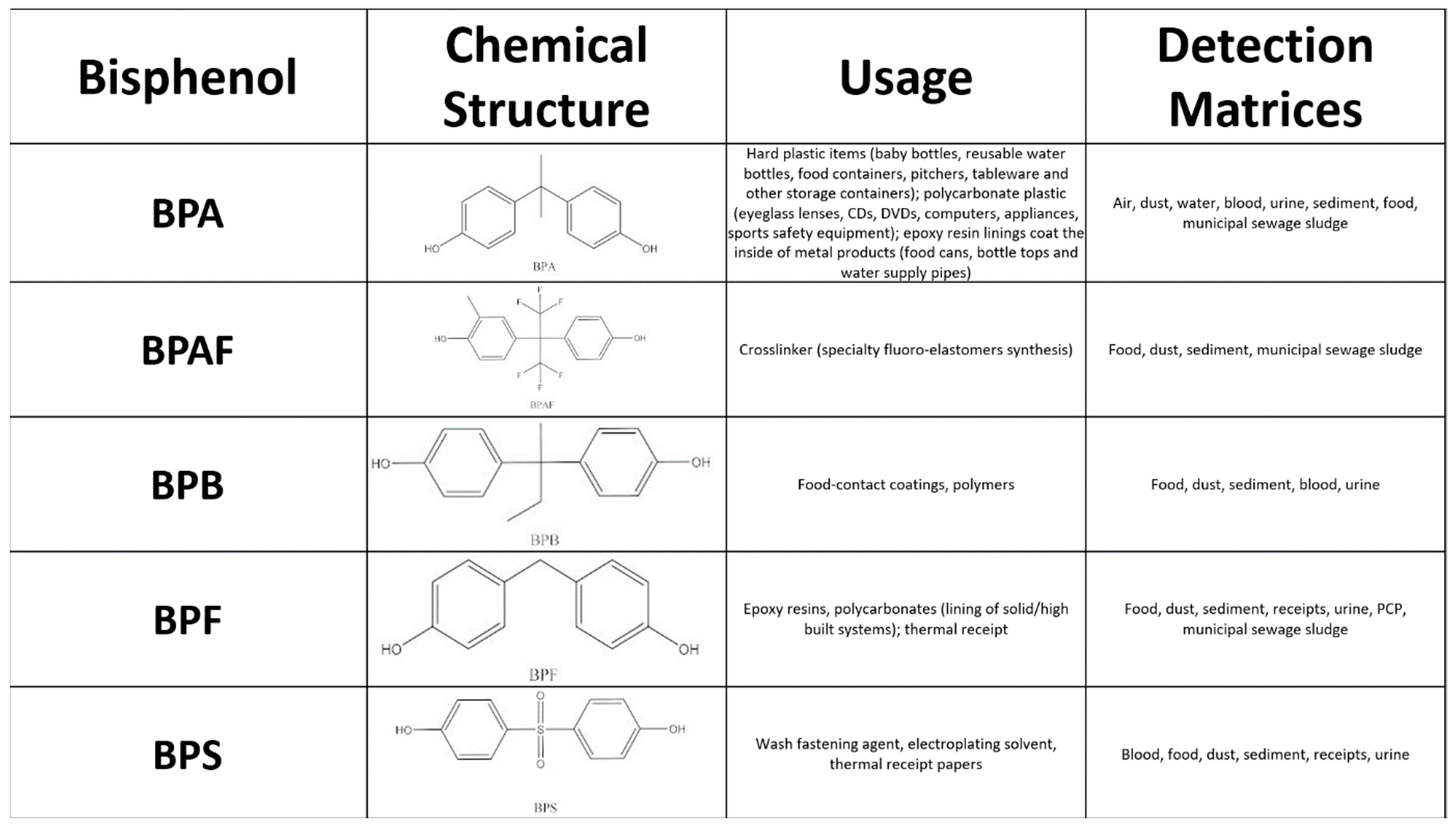
:1. Introduction
2. Materials and Methods
2.1. H295R Cell Culture and Treatment
2.2. Mitochondrial Activity Assay
2.3. CFDA-AM Assay
2.4. Assessment of Intracellular Cholesterol Concentration
2.5. Assessment of Steroid Hormone Production
2.6. Statistical Analysis
3. Results
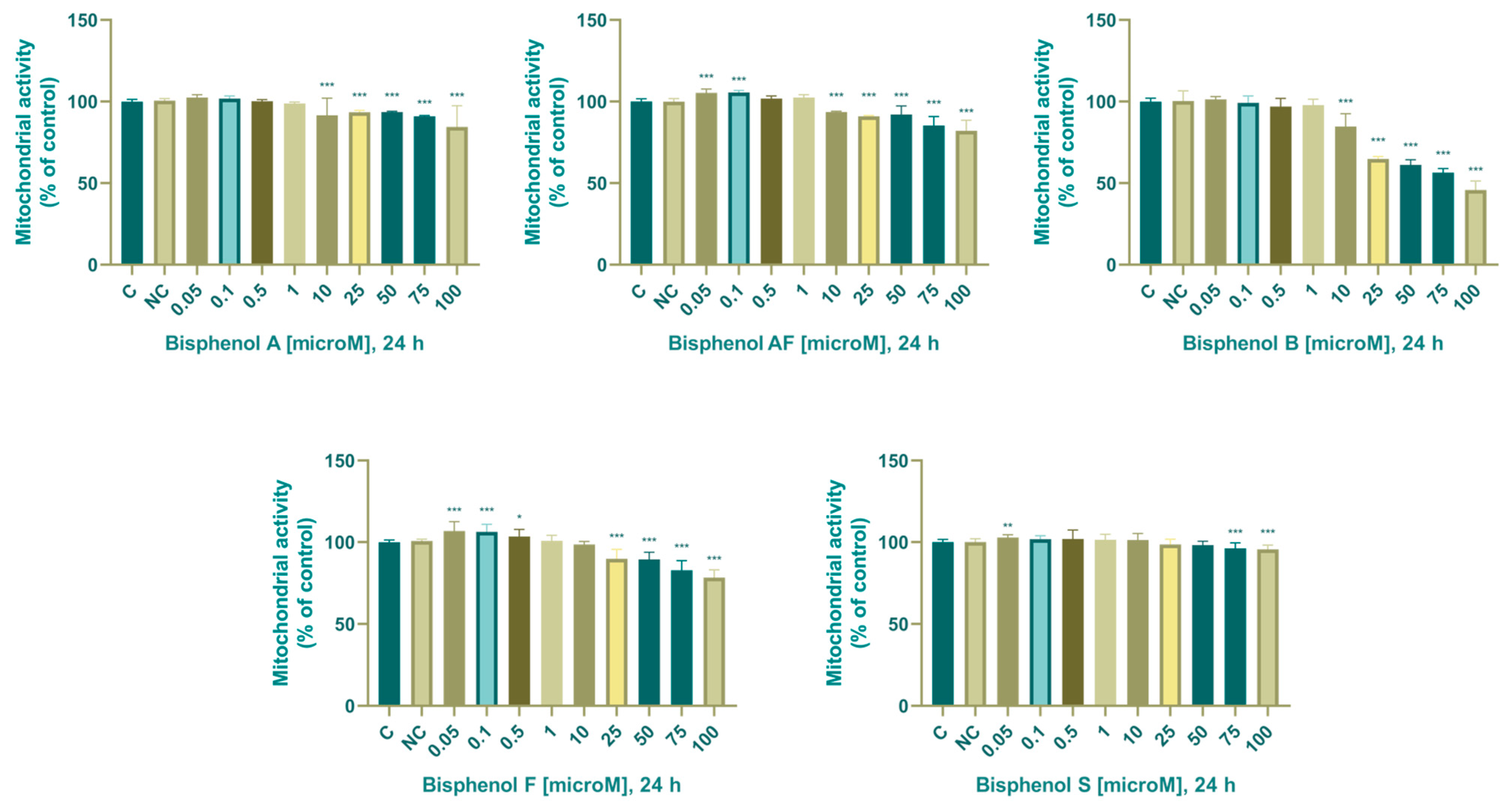
3.1. Viability Parameters
3.1.1. Mitochondrial Activity
3.1.2. Membrane Integrity
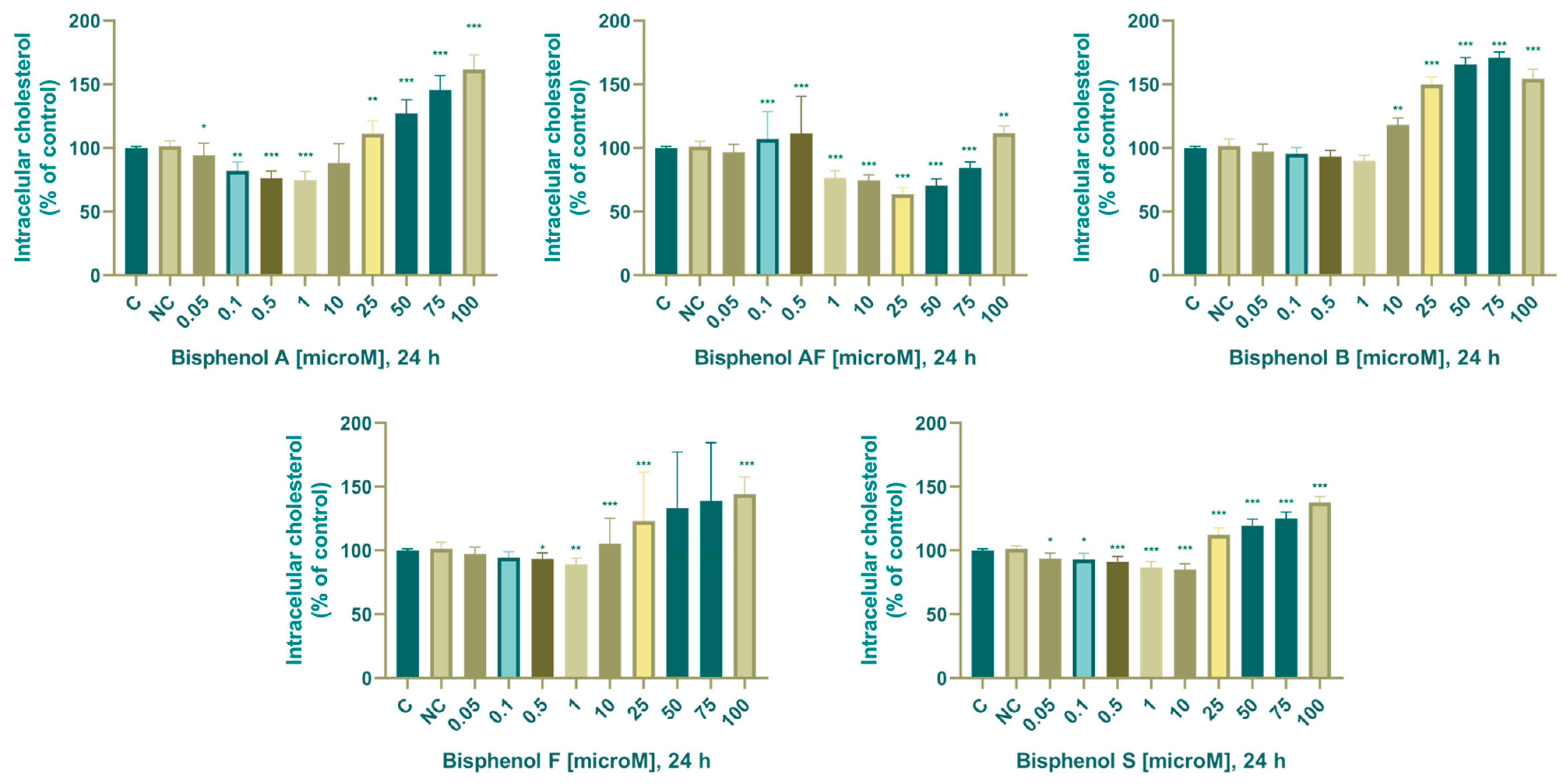
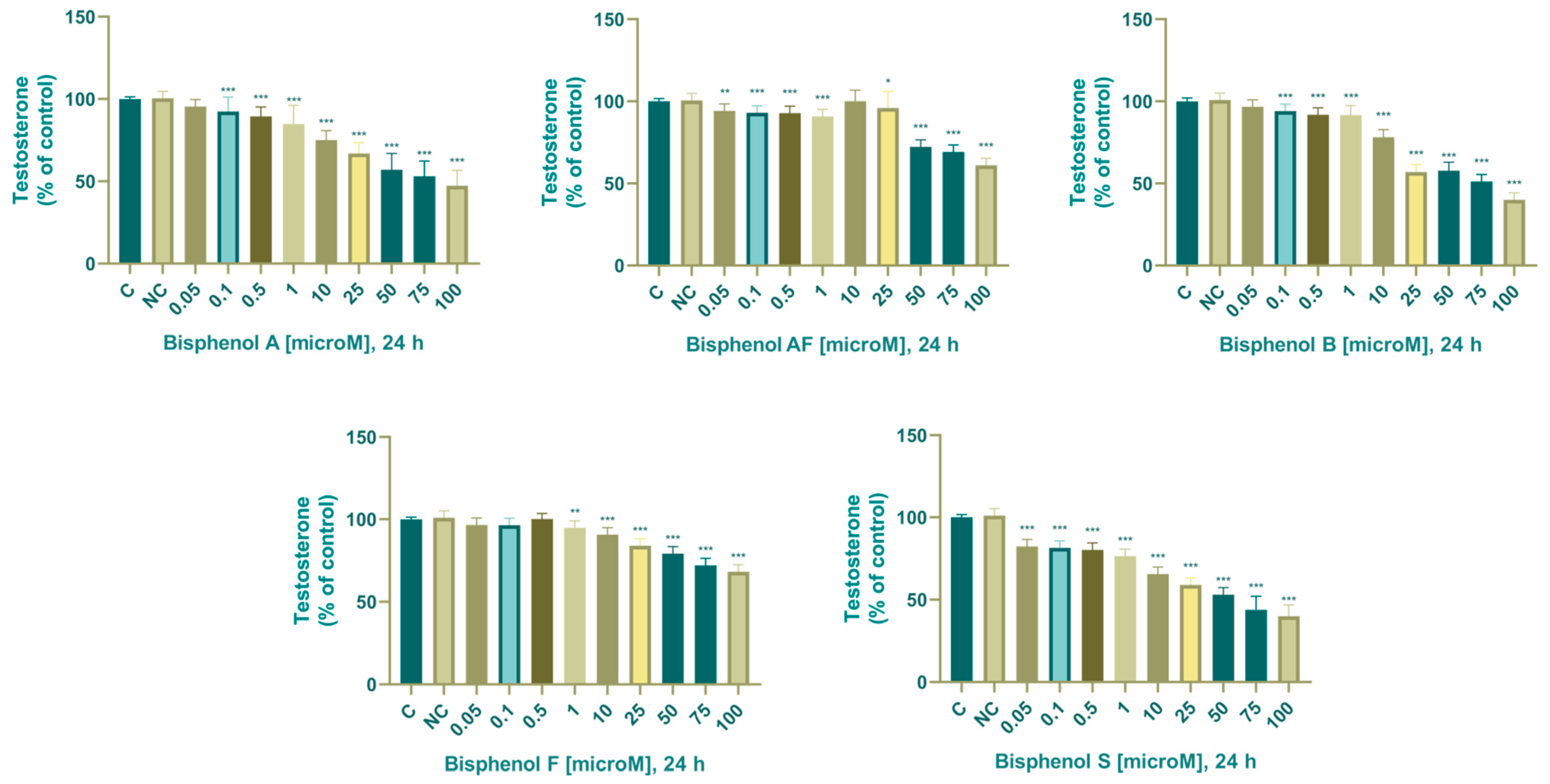
3.2. Steroidogenesis
3.2.1. Cholesterol
3.2.2. Progesterone
3.2.3. Estradiol
3.2.4. Testosterone
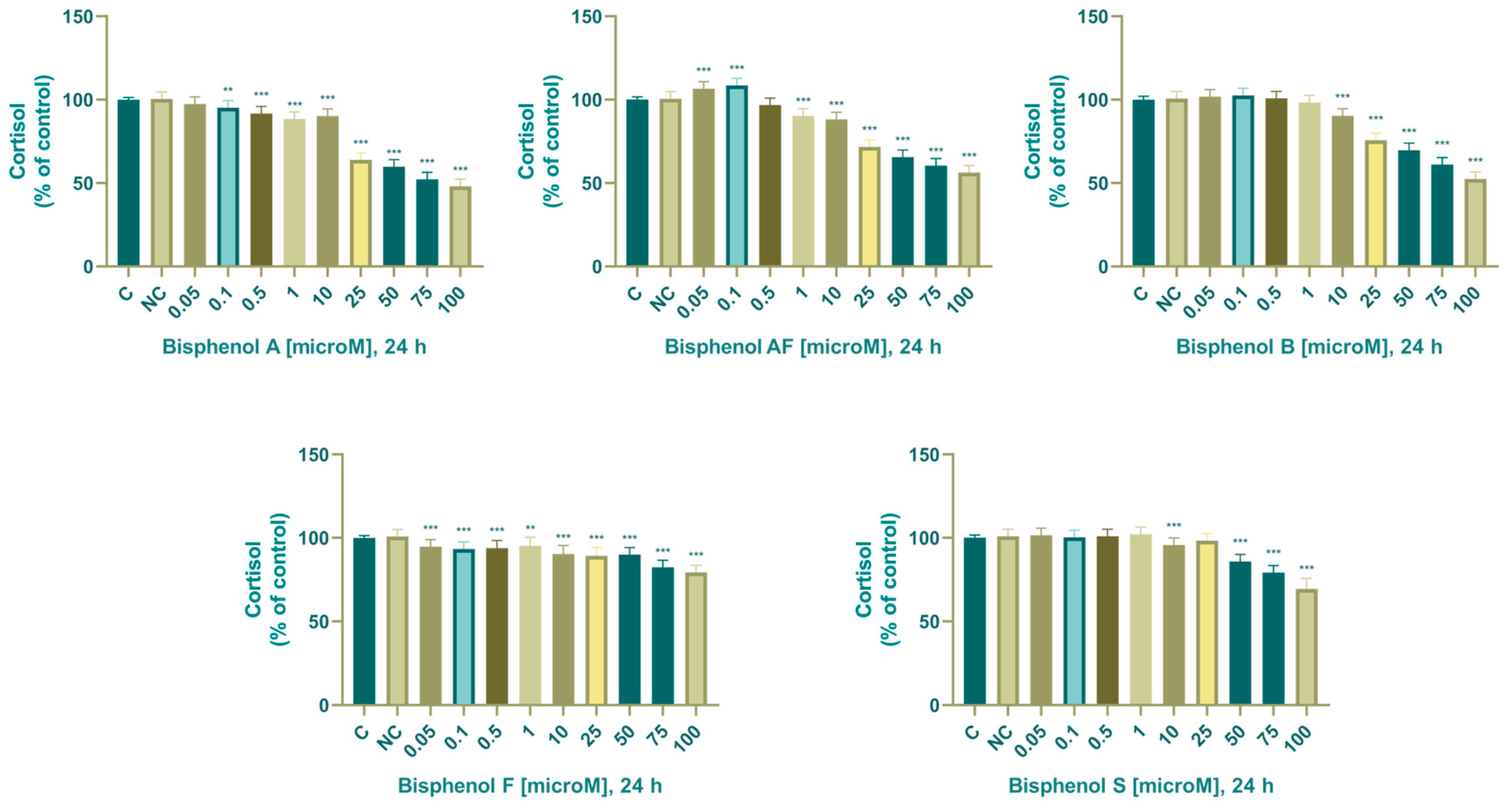
3.2.5. Cortisol
4. Discussion
4.1. BPA Alternatives Affected Cell Viability
4.2. BPA Alternatives Impaired Steroidogenesis
4.3. Intracellular Cholesterol
4.4. Progesterone
4.5. Estradiol
4.6. Testosterone
4.7. Cortisol
4.8. Future Direction and Challenges
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schug, T.T.; Janesick, A.; Blumberg, B.; Heindel, J.J. Endocrine disrupting chemicals and disease susceptibility. J. Steroid Biochem. Mol. Biol. 2011, 127, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, E.J.; Simoneau, C. Release of Bisphenol A from Polycarbonate—A Review. Crit. Rev. Food Sci. Nutr. 2013, 53, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Harnett, K.G.; Chin, A.; Schuh, S.M. BPA and BPA alternatives BPS, BPAF, and TMBPF, induce cytotoxicity and apoptosis in rat and human stem cells. Ecotoxicol. Environ. Saf. 2021, 216, 112210. [Google Scholar] [CrossRef] [PubMed]
- Geens, T.; Aerts, D.; Berthot, C.; Bourguignon, J.-P.; Goeyens, L.; Lecomte, P.; Maghuin-Rogister, G.; Pironnet, A.-M.; Pussemier, L.; Scippo, M.-L.; et al. A review of dietary and non-dietary exposure to bisphenol-A. Food Chem. Toxicol. 2012, 50, 3725–3740. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.; Kristofco, L.A.; Steele, W.B.; Yates, B.S.; Breed, C.S.; Williams, E.S.; Brooks, B.W. Global Assessment of Bisphenol A in the Environment: Review and Analysis of Its Occurrence and Bioaccumulation. Dose-Response 2015, 13, 1559325815598308. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Chakraborty, P. A review on sources and health impacts of bisphenol A. Rev. Environ. Health 2020, 35, 201–210. [Google Scholar] [CrossRef]
- Kang, J.-H.; Kondo, F.; Katayama, Y. Human exposure to bisphenol A. Toxicology 2006, 226, 79–89. [Google Scholar] [CrossRef]
- Hines, C.J.; Jackson, M.V.; Christianson, A.L.; Clark, J.C.; Arnold, J.E.; Pretty, J.R.; Deddens, J.A. Air, hand wipe, and surface wipe sampling for Bisphenol A (BPA) among workers in industries that manufacture and use BPA in the United States. J. Occup. Environ. Hyg. 2017, 14, 882–897. [Google Scholar] [CrossRef]
- Chakraborty, P.; Sampath, S.; Mukhopadhyay, M.; Selvaraj, S.; Bharat, G.K.; Nizzetto, L. Baseline investigation on plasticizers, bisphenol A, polycyclic aromatic hydrocarbons and heavy metals in the surface soil of the informal electronic waste recycling workshops and nearby open dumpsites in Indian metropolitan cities. Environ. Pollut. 2019, 248, 1036–1045. [Google Scholar] [CrossRef]
- Chakraborty, P.; Mukhopadhyay, M.; Sampath, S.; Ramaswamy, B.R.; Katsoyiannis, A.; Cincinelli, A.; Snow, D. Organic micropollutants in the surface riverine sediment along the lower stretch of the transboundary river Ganga: Occurrences, sources and ecological risk assessment. Environ. Pollut. 2018, 249, 1071–1080. [Google Scholar] [CrossRef]
- Santhi, V.; Sakai, N.; Ahmad, E.; Mustafa, A. Occurrence of bisphenol A in surface water, drinking water and plasma from Malaysia with exposure assessment from consumption of drinking water. Sci. Total Environ. 2012, 427–428, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Fabbri, E. Environmental Effects of BPA. Dose-Response 2015, 13, 1559325815598304. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Suh, S. Health risks of chemicals in consumer products: A review. Environ. Int. 2019, 123, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.H.; Han, J.H.; Lee, S.-B.; Lee, Y.-H. Inhalation Toxicity of Bisphenol A and Its Effect on Estrous Cycle, Spatial Learning, and Memory in Rats upon Whole-Body Exposure. Toxicol. Res. 2017, 33, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Lang, I.A.; Galloway, T.S.; Scarlett, A.; Henley, W.E.; Depledge, M.; Wallace, R.B.; Melzer, D. Association of Urinary Bisphenol A Concentration with Medical Disorders and Laboratory Abnormalities in Adults. JAMA 2008, 300, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.M.F.; Spritzer, P.M.; Hohl, A.; Bachega, T.A.S.S. Effects of endocrine disruptors in the development of the female reproductive tract. Arq. Bras. Endocrinol. Metabol. 2014, 58, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Osborne, G.; Rudel, R.; Schwarzman, M. Evaluating chemical effects on mammary gland development: A critical need in disease prevention. Reprod. Toxicol. 2015, 54, 148–155. [Google Scholar] [CrossRef]
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef]
- Ejaredar, M.; Lee, Y.; Roberts, D.J.; Sauve, R.; Dewey, D. Bisphenol A exposure and children’s behavior: A systematic review. J. Expo. Sci. Environ. Epidemiol. 2016, 27, 175–183. [Google Scholar] [CrossRef]
- Rubin, B.S.; Murray, M.K.; Damassa, D.A.; King, J.C.; Soto, A.M. Perinatal exposure to low doses of bisphenol A affects body weight, patterns of estrous cyclicity, and plasma LH levels. Environ. Health Perspect. 2001, 109, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Troisi, J.; Richards, S.; Scafuro, M.; Fasano, S.; Guida, M.; Pierantoni, R.; Meccariello, R. Bisphenol A in Reproduction: Epigenetic Effects. Curr. Med. Chem. 2018, 25, 748–770. [Google Scholar] [CrossRef] [PubMed]
- Mokra, K.; Kocia, M.; Michałowicz, J. Bisphenol A and its analogs exhibit different apoptotic potential in peripheral blood mononuclear cells (in vitro study). Food Chem. Toxicol. 2015, 84, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Suzuki, T.; Sanoh, S.; Kohta, R.; Jinno, N.; Sugihara, K.; Yoshihara, S.; Fujimoto, N.; Watanabe, H.; Ohta, S. Comparative Study of the Endocrine-Disrupting Activity of Bisphenol A and 19 Related Compounds. Toxicol. Sci. 2005, 84, 249–259. [Google Scholar] [CrossRef]
- Iida, H.; Maehara, K.; Doiguchi, M.; Mōri, T.; Yamada, F. Bisphenol A-induced apoptosis of cultured rat Sertoli cells. Reprod. Toxicol. 2003, 17, 457–464. [Google Scholar] [CrossRef]
- Hwang, J.K.; Min, K.H.; Choi, K.H.; Hwang, Y.C.; Jeong, I.-K.; Ahn, K.J.; Chung, H.-Y.; Chang, J.S. Bisphenol A reduces differentiation and stimulates apoptosis of osteoclasts and osteoblasts. Life Sci. 2013, 93, 367–372. [Google Scholar] [CrossRef]
- Gong, X.; Xie, H.; Li, X.; Wu, J.; Lin, Y. Bisphenol A induced apoptosis and transcriptome differences of spermatogonial stem cells in vitro. Acta Biochim. Biophys. Sin. 2017, 49, 780–791. [Google Scholar] [CrossRef]
- Zhang, X.; Chang, H.; Wiseman, S.; He, Y.; Higley, E.; Jones, P.; Wong, C.K.; Al-Khedhairy, A.; Giesy, J.P.; Hecker, M. Bisphenol A Disrupts Steroidogenesis in Human H295R Cells. Toxicol. Sci. 2011, 121, 320–327. [Google Scholar] [CrossRef]
- Bloom, M.S.; Mok-Lin, E.; Fujimoto, V.Y. Bisphenol A and ovarian steroidogenesis. Fertil. Steril. 2016, 106, 857–863. [Google Scholar] [CrossRef]
- Jambor, T.; Knížatová, N.; Lukáč, N. Men’s reproductive alterations caused by bisphenol A and its analogues: A review. Physiol. Res. 2021, 70, S643–S656. [Google Scholar] [CrossRef]
- Rathee, M.; Malik, P.; Singh, J. Bisphenol A in dental sealants and its estrogen like effect. Indian J. Endocrinol. Metab. 2012, 16, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Alomirah, H.; Cho, H.-S.; Li, Y.-F.; Liao, C.; Minh, T.B.; Mohd, M.A.; Nakata, H.; Ren, N.; Kannan, K. Urinary Bisphenol A Concentrations and Their Implications for Human Exposure in Several Asian Countries. Environ. Sci. Technol. 2011, 45, 7044–7050. [Google Scholar] [CrossRef] [PubMed]
- Longnecker, M.; Harbak, K.; Kissling, G.; Hoppin, J.; Eggesbo, M.; Jusko, T.; Eide, J.; Koch, H. The concentration of bisphenol A in urine is affected by specimen collection, a preservative, and handling. Environ. Res. 2013, 126, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.M.; Kolossa-Gehring, M.; Schröter-Kermani, C.; Angerer, J.; Brüning, T. Bisphenol A in 24 h urine and plasma samples of the German Environmental Specimen Bank from 1995 to 2009: A retrospective exposure evaluation. J. Expo. Sci. Environ. Epidemiol. 2012, 22, 610–616. [Google Scholar] [CrossRef]
- Johns, L.E.; Ferguson, K.K.; Meeker, J.D. Relationships Between Urinary Phthalate Metabolite and Bisphenol A Concentrations and Vitamin D Levels in U.S. Adults: National Health and Nutrition Examination Survey (NHANES), 2005–2010. J. Clin. Endocrinol. Metab. 2016, 101, 4062–4069. [Google Scholar] [CrossRef]
- Li, Z.; Mao, W.; Yao, L.; Zhao, N.; Zhang, Y.; Zhao, M.; Jin, H. First report on occurrence of bisphenol A isomers in human serum and whole blood. J. Hazard. Mater. 2022, 424, 127549. [Google Scholar] [CrossRef]
- Lee, J.; Choi, K.; Park, J.; Moon, H.-B.; Choi, G.; Lee, J.J.; Suh, E.; Kim, H.-J.; Eun, S.-H.; Kim, G.-H.; et al. Bisphenol A distribution in serum, urine, placenta, breast milk, and umbilical cord serum in a birth panel of mother–neonate pairs. Sci. Total Environ. 2018, 626, 1494–1501. [Google Scholar] [CrossRef]
- Aris, A. Estimation of bisphenol A (BPA) concentrations in pregnant women, fetuses and nonpregnant women in Eastern Townships of Canada. Reprod. Toxicol. 2014, 45, 8–13. [Google Scholar] [CrossRef]
- Nishikawa, M.; Iwano, H.; Yanagisawa, R.; Koike, N.; Inoue, H.; Yokota, H. Placental Transfer of Conjugated Bisphenol A and Subsequent Reactivation in the Rat Fetus. Environ. Health Perspect. 2010, 118, 1196–1203. [Google Scholar] [CrossRef]
- Khan, J.; Salhotra, S.; Goswami, P.; Akhter, J.; Jahan, S.; Gupta, S.; Sharma, S.; Banerjee, B.D.; Parvez, S.; Gupta, S.; et al. Bisphenol A triggers axonal injury and myelin degeneration with concomitant neurobehavioral toxicity in C57BL/6J male mice. Toxicology 2019, 428, 152299. [Google Scholar] [CrossRef]
- Knížatová, N.; Greifová, H.; Tokárová, K.; Jambor, T.; Lukáč, N. Evaluation of dose-dependent activity of bisphenol f on viability parameters and steroidogenesis in h295r cells. J. Microbiol. Biotechnol. Food Sci. 2021, 11, e4765. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Knížatová, N.; Greifová, H.; Tokárová, K.; Jambor, T.; Binkowski, J.; Lukáč, N. Assessment of the Effective Impact of Bisphenols on Mitochondrial Activity, Viability and Steroidogenesis in a Dose-Dependency in Human Adrenocortical Carcinoma Cells. Processes 2021, 9, 1471. [Google Scholar] [CrossRef]
- Mpatani, F.M.; Han, R.; Aryee, A.A.; Kani, A.N.; Li, Z.; Qu, L. Adsorption performance of modified agricultural waste materials for removal of emerging micro-contaminant bisphenol A: A comprehensive review. Sci. Total Environ. 2021, 780, 146629. [Google Scholar] [CrossRef] [PubMed]
- Colorado-Yohar, S.M.; Castillo-González, A.C.; Sánchez-Meca, J.; Rubio-Aparicio, M.; Sánchez-Rodríguez, D.; Salamanca-Fernández, E.; Ardanaz, E.; Amiano, P.; Fernández, M.F.; Mendiola, J.; et al. Concentrations of bisphenol-A in adults from the general population: A systematic review and meta-analysis. Sci. Total Environ. 2021, 775, 145755. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.-Y.; Chen, P.-P.; Liu, C.; Deng, Y.-L.; Miao, Y.; Zhang, M.; Cui, F.-P.; Lu, T.-T.; Shi, T.; Yang, K.-D.; et al. Bisphenol A analogues in associations with serum hormone levels among reproductive-aged Chinese men. Environ. Int. 2022, 167, 107446. [Google Scholar] [CrossRef]
- Dunder, L.; Lejonklou, M.H.; Lind, P.M.; Lind, L. Urinary bisphenol A and serum lipids: A meta-analysis of six NHANES examination cycles (2003–2014). J. Epidemiol. Community Health 2019, 73, 1012–1019. [Google Scholar] [CrossRef]
- Žalmanová, T.; Hošková, K.; Prokešová, Š.; Nevoral, J.; Ješeta, M.; Benc, M.; Yi, Y.-J.; Moravec, J.; Močáryová, B.; Martínková, S.; et al. The bisphenol S contamination level observed in human follicular fluid affects the development of porcine oocytes. Front. Cell Dev. Biol. 2023, 11, 1145182. [Google Scholar] [CrossRef]
- Huang, S.; Li, J.; Xu, S.; Zhao, H.; Li, Y.; Zhou, Y.; Fang, J.; Liao, J.; Cai, Z.; Xia, W. Bisphenol A and bisphenol S exposures during pregnancy and gestational age—A longitudinal study in China. Chemosphere 2019, 237, 124426. [Google Scholar] [CrossRef] [PubMed]
- Smarr, M.M.; Kannan, K.; Sun, L.; Honda, M.; Wang, W.; Karthikraj, R.; Chen, Z.; Weck, J.; Louis, G.M.B. Preconception seminal plasma concentrations of endocrine disrupting chemicals in relation to semen quality parameters among male partners planning for pregnancy. Environ. Res. 2018, 167, 78–86. [Google Scholar] [CrossRef]
- Beausoleil, C.; Le Magueresse-Battistoni, B.; Viguié, C.; Babajko, S.; Canivenc-Lavier, M.-C.; Chevalier, N.; Emond, C.; Habert, R.; Picard-Hagen, N.; Mhaouty-Kodja, S. Regulatory and academic studies to derive reference values for human health: The case of bisphenol S. Environ. Res. 2021, 204, 112233. [Google Scholar] [CrossRef]
- Thayer, K.A.; Taylor, K.W.; Garantziotis, S.; Schurman, S.H.; Kissling, G.E.; Hunt, D.; Herbert, B.; Church, R.; Jankowich, R.; Churchwell, M.I.; et al. Bisphenol A, Bisphenol S, and 4-HydroXyphenyl 4-IsoproOxyphenylSulfone (BPSIP) in Urine and Blood of Cashiers. Environ. Health Perspect. 2016, 124, 437–444. [Google Scholar] [CrossRef]
- Alharbi, H.F.; Algonaiman, R.; Alduwayghiri, R.; Aljutaily, T.; Algheshairy, R.M.; Almutairi, A.S.; Alharbi, R.M.; Alfurayh, L.A.; Alshahwan, A.A.; Alsadun, A.F.; et al. Exposure to Bisphenol A Substitutes, Bisphenol S and Bisphenol F, and Its Association with Developing Obesity and Diabetes Mellitus: A Narrative Review. Int. J. Environ. Res. Public Health 2022, 19, 15918. [Google Scholar] [CrossRef]
- Lehmler, H.-J.; Liu, B.; Gadogbe, M.; Bao, W. Exposure to Bisphenol A, Bisphenol F, and Bisphenol S in U.S. Adults and Children: The National Health and Nutrition Examination Survey 2013–2014. ACS Omega 2018, 3, 6523–6532. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Y.; Chen, D.; Chen, H.; Liu, X. Bisphenol A and its analogues in paired urine and house dust from South China and implications for children’s exposure. Chemosphere 2022, 294, 133701. [Google Scholar] [CrossRef]
- Blaauwendraad, S.M.; Voerman, E.; Trasande, L.; Kannan, K.; Santos, S.; Ruijter, G.J.G.; Sol, C.M.; Marchioro, L.; Shokry, E.; Koletzko, B.; et al. Associations of maternal bisphenol urine concentrations during pregnancy with neonatal metabolomic profiles. Metabolomics 2021, 17, 84. [Google Scholar] [CrossRef]
- Cobellis, L.; Colacurci, N.; Trabucco, E.; Carpentiero, C.; Grumetto, L. Measurement of bisphenol A and bisphenol B levels in human blood sera from healthy and endometriotic women. Biomed. Chromatogr. 2009, 23, 1186–1190. [Google Scholar] [CrossRef]
- Jin, H.; Zhu, J.; Chen, Z.; Hong, Y.; Cai, Z. Occurrence and Partitioning of Bisphenol Analogues in Adults’ Blood from China. Environ. Sci. Technol. 2017, 52, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Vishalakshi, G.J.; Hemshekhar, M.; Sandesha, V.D.; Prashanth, K.S.; Jagadish, S.; Paul, M.; Kemparaju, K.; Girish, K.S. Bisphenol AF elevates procoagulant platelets by inducing necroptosis via RIPK1-inflammasome axis. Toxicology 2021, 454, 152742. [Google Scholar] [CrossRef]
- Pelch, K.; Wignall, J.A.; Goldstone, A.E.; Ross, P.K.; Blain, R.B.; Shapiro, A.J.; Holmgren, S.D.; Hsieh, J.-H.; Svoboda, D.; Auerbach, S.S.; et al. A scoping review of the health and toxicological activity of bisphenol A (BPA) structural analogues and functional alternatives. Toxicology 2019, 424, 152235. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, L.; Lu, G.; Jiang, R.; Yan, Z.; Li, Y. Occurrence, toxicity and ecological risk of Bisphenol A analogues in aquatic environment—A review. Ecotoxicol. Environ. Saf. 2021, 208, 111481. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Huo, X.; Zhang, J.; Mao, Y.; Li, G.; Ye, J.; Shi, Y.; Jin, F.; Bao, S.; Zhang, J. Environmental exposure to bisphenol analogues and unexplained recurrent miscarriage: A case-control study. Environ. Res. 2022, 204, 112293. [Google Scholar] [CrossRef] [PubMed]
- Bousoumah, R.; Leso, V.; Iavicoli, I.; Huuskonen, P.; Viegas, S.; Porras, S.P.; Santonen, T.; Frery, N.; Robert, A.; Ndaw, S. Biomonitoring of occupational exposure to bisphenol A, bisphenol S and bisphenol F: A systematic review. Sci. Total Environ. 2021, 783, 146905. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Guo, M.; Yin, X.; Huang, C.; Qian, L.; Zhou, L.; Wang, Z.; Wang, L.; Shi, L.; Ji, G. A systematic comparison of neurotoxicity of bisphenol A and its derivatives in zebrafish. Sci. Total Environ. 2022, 805, 150210. [Google Scholar] [CrossRef]
- Feng, Y.; Jiao, Z.; Shi, J.; Li, M.; Guo, Q.; Shao, B. Effects of bisphenol analogues on steroidogenic gene expression and hormone synthesis in H295R cells. Chemosphere 2016, 147, 9–19. [Google Scholar] [CrossRef]
- Lin, J.; Deng, L.; Sun, M.; Wang, Y.; Lee, S.; Choi, K.; Liu, X. An in vitro investigation of endocrine disrupting potentials of ten bisphenol analogues. Steroids 2021, 169, 108826. [Google Scholar] [CrossRef]
- Gonçalves, G.D.; Semprebon, S.C.; Biazi, B.I.; Mantovani, M.S.; Fernandes, G.S.A. Bisphenol A reduces testosterone production in TM3 Leydig cells independently of its effects on cell death and mitochondrial membrane potential. Reprod. Toxicol. 2018, 76, 26–34. [Google Scholar] [CrossRef]
- Harrington, J.S.; Ryter, S.W.; Plataki, M.; Price, D.R.; Choi, A.M.K. Mitochondria in health, disease, and aging. Physiol. Rev. 2023, 103, 2349–2422. [Google Scholar] [CrossRef]
- Meyer, J.N.; Leung, M.C.K.; Rooney, J.P.; Sendoel, A.; Hengartner, M.O.; Kisby, G.E.; Bess, A.S. Mitochondria as a Target of Environmental Toxicants. Toxicol. Sci. 2013, 134, 1–17. [Google Scholar] [CrossRef]
- Goldinger, D.M.; Demierre, A.-L.; Zoller, O.; Rupp, H.; Reinhard, H.; Magnin, R.; Becker, T.W.; Bourqui-Pittet, M. Endocrine activity of alternatives to BPA found in thermal paper in Switzerland. Regul. Toxicol. Pharmacol. 2015, 71, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.J.; Bin Park, S.; Park, J.W.; Oh, S.R.; Han, M. Bisphenol A modulates inflammation and proliferation pathway in human endometrial stromal cells by inducing oxidative stress. Reprod. Toxicol. 2018, 81, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.-C.; Wu, K.-Y.; Lin, I.-W.; Yang, Z.-J.; Chang, A.-A.; Hu, M.-C. Bisphenol A Disrupts Steroidogenesis and Induces a Sex Hormone Imbalance through C-Jun Phosphorylation in Leydig Cells. Chemosphere 2017, 185, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Tyner, M.D.; Maloney, M.O.; Kelley, B.J.; Combelles, C.M. Comparing the effects of bisphenol A, C, and F on bovine theca cells in vitro. Reprod. Toxicol. 2022, 111, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Guarnotta, V.; Amodei, R.; Frasca, F.; Aversa, A.; Giordano, C. Impact of Chemical Endocrine Disruptors and Hormone Modulators on the Endocrine System. Int. J. Mol. Sci. 2022, 23, 5710. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, H.; Zou, J.; Feng, X.; Feng, D. Bisphenol A induces cholesterol biosynthesis in HepG2 cells via SREBP-2/HMGCR signaling pathway. J. Toxicol. Sci. 2019, 44, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yang, S.; Gao, R.; Deng, Y.; Liu, J.; Yuan, C.; Yao, Q.; Lv, X.; Wang, K.; Ye, X.; et al. Relationship Between the Environmental Endocrine Disruptor Bisphenol a and Dyslipidemia: A Five-Year Prospective Study. Endocr. Pract. 2020, 26, 399–406. [Google Scholar] [CrossRef]
- Wang, B.; Wang, S.; Zhao, Z.; Chen, Y.; Xu, Y.; Li, M.; Xu, M.; Wang, W.; Ning, G.; Bi, Y.; et al. Bisphenol A exposure in relation to altered lipid profile and dyslipidemia among Chinese adults: A repeated measures study. Environ. Res. 2020, 184, 109382. [Google Scholar] [CrossRef]
- Gao, Z.; Liu, S.; Tan, L.; Gao, X.; Fan, W.; Ding, C.; Li, M.; Tang, Z.; Shi, X.; Luo, Y.; et al. Testicular toxicity of bisphenol compounds: Homeostasis disruption of cholesterol/testosterone via PPARα activation. Sci. Total Environ. 2022, 836, 155628. [Google Scholar] [CrossRef]
- Samardzija, D.; Pogrmic-Majkic, K.; Fa, S.; Stanic, B.; Jasnic, J.; Andric, N. Bisphenol A decreases progesterone synthesis by disrupting cholesterol homeostasis in rat granulosa cells. Mol. Cell Endocrinol. 2018, 461, 55–63. [Google Scholar] [CrossRef]
- Maeda, N.; Fujiki, J.; Hasegawa, Y.; Ieko, T.; Miyasho, T.; Iwasaki, T.; Yokota, H. Testicular induced corticosterone synthesis in male rats under fasting stress. Steroids 2022, 177, 108947. [Google Scholar] [CrossRef] [PubMed]
- Rosenmai, A.K.; Dybdahl, M.; Pedersen, M.; van Vugt-Lussenburg, B.M.A.; Wedebye, E.B.; Taxvig, C.; Vinggaard, A.M. Are Structural Analogues to Bisphenol A Safe Alternatives? Toxicol. Sci. 2014, 139, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Mlynarcikova, A.B.; Scsukova, S. Bisphenol analogs AF, S and F: Effects on functional characteristics of porcine granulosa cells. Reprod. Toxicol. 2021, 103, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Amar, S.; Binet, A.; Téteau, O.; Desmarchais, A.; Papillier, P.; Lacroix, M.Z.; Maillard, V.; Guérif, F.; Elis, S. Bisphenol S Impaired Human Granulosa Cell Steroidogenesis in Vitro. Int. J. Mol. Sci. 2020, 21, 1821. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Shi, J.; Jiao, Z.; Duan, H.; Shao, B. Mechanism of bisphenol AF-induced progesterone inhibition in human chorionic gonadotrophin-stimulated mouse Leydig tumor cell line (mLTC-1) cells. Environ. Toxicol. 2018, 33, 670–678. [Google Scholar] [CrossRef]
- Téteau, O.; Jaubert, M.; Desmarchais, A.; Papillier, P.; Binet, A.; Maillard, V.; Elis, S. Bisphenol A and S impaired ovine granulosa cell steroidogenesis. Reproduction 2020, 159, 571–583. [Google Scholar] [CrossRef]
- Qi, J.; Liu, L.; Yang, J.; Gao, X.; Zhang, W. Bisphenol A decreases progesterone synthesis in human ovarian granulosa cells. Birth Defects Res. 2020, 112, 1843–1849. [Google Scholar] [CrossRef]
- Shoorei, H.; Seify, M.; Talebi, S.F.; Majidpoor, J.; Koohestanidehaghi, Y.; Shokoohi, M. Different types of bisphenols alter ovarian steroidogenesis: Special attention to BPA. Heliyon 2023, 9, e16848. [Google Scholar] [CrossRef]
- Mansur, A.; Adir, M.; Yerushalmi, G.; Hourvitz, A.; Gitman, H.; Yung, Y.; Orvieto, R.; Machtinger, R. Does BPA alter steroid hormone synthesis in human granulosa cells in vitro? Hum. Reprod. 2016, 31, 1562–1569. [Google Scholar] [CrossRef]
- Lin, M.; Hua, R.; Ma, J.; Zhou, Y.; Li, P.; Xu, X.; Yu, Z.; Quan, S. Bisphenol A promotes autophagy in ovarian granulosa cells by inducing AMPK/mTOR/ULK1 signalling pathway. Environ. Int. 2021, 147, 106298. [Google Scholar] [CrossRef]
- Kwintkiewicz, J.; Nishi, Y.; Yanase, T.; Giudice, L.C. Peroxisome Proliferator–Activated Receptor-γ Mediates Bisphenol A Inhibition of FSH-Stimulated IGF-1, Aromatase, and Estradiol in Human Granulosa Cells. Environ. Health Perspect. 2010, 118, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jin, H.; Kim, G.; Bae, J. A low dose of bisphenol A stimulates estradiol production by regulating β-catenin-FOXL2-CYP19A1 pathway in human ovarian granulosa cells. Biochem. Biophys. Res. Commun. 2021, 583, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Leung, L.K. Bisphenol A downregulates CYP19 transcription in JEG-3 cells. Toxicol. Lett. 2009, 189, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, X.; Ye, Y.; Li, X. Bisphenol A affects estradiol metabolism by targeting CYP1A1 and CYP19A1 in human placental JEG-3 cells. Toxicol. Vitr. 2019, 61, 104615. [Google Scholar] [CrossRef] [PubMed]
- Serra, H.; Beausoleil, C.; Habert, R.; Minier, C.; Picard-Hagen, N.; Michel, C. Evidence for Bisphenol B Endocrine Properties: Scientific and Regulatory Perspectives. Environ. Health Perspect. 2019, 127, 106001. [Google Scholar] [CrossRef] [PubMed]
- Eladak, S.; Grisin, T.; Moison, D.; Guerquin, M.-J.; N’Tumba-Byn, T.; Pozzi-Gaudin, S.; Benachi, A.; Livera, G.; Rouiller-Fabre, V.; Habert, R. A new chapter in the bisphenol A story: Bisphenol S and bisphenol F are not safe alternatives to this compound. Fertil. Steril. 2015, 103, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Turi, N.; Ullah, W.; Siddiqui, M.F.; Zakria, M.; Lodhi, K.Z.; Khan, M.M. Impact of low-dose chronic exposure to bisphenol A and its analogue bisphenol B, bisphenol F and bisphenol S on hypothalamo-pituitary-testicular activities in adult rats: A focus on the possible hormonal mode of action. Food Chem. Toxicol. 2018, 121, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Shaheen, G.; Rehman, H.; Siddiqui, M.F.; Butt, M.A. Bisphenol A and its analogs bisphenol B, bisphenol F, and bisphenol S: Comparative in vitro and in vivo studies on the sperms and testicular tissues of rats. Chemosphere 2018, 209, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Grignard, E.; Lapenna, S.; Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicol. Vitr. 2012, 26, 727–731. [Google Scholar] [CrossRef]
- Wang, Y.; Aimuzi, R.; Nian, M.; Zhang, Y.; Luo, K.; Zhang, J. Bisphenol A substitutes and sex hormones in children and adolescents. Chemosphere 2021, 278, 130396. [Google Scholar] [CrossRef]
- Feng, Y.; Yin, J.; Jiao, Z.; Shi, J.; Li, M.; Shao, B. Bisphenol AF may cause testosterone reduction by directly affecting testis function in adult male rats. Toxicol. Lett. 2012, 211, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Pirzada, M.; Afsar, T.; Razak, S.; Almajwal, A.; Jahan, S. Effect of bisphenol F, an analog of bisphenol A, on the reproductive functions of male rats. Environ. Health Prev. Med. 2019, 24, 41. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xu, H.; Gao, Z.; Li, M.; Zhu, Y.; Li, Y. Effects of bisphenol A on testis testosterone synthesis in mouse cultured in vitro. J. Hyg. Res. 2013, 42, 543–549. [Google Scholar]
- Zhang, W.; Xia, W.; Liu, W.; Li, X.; Hu, J.; Zhang, B.; Xu, S.; Zhou, Y.; Li, J.; Cai, Z.; et al. Exposure to Bisphenol a Substitutes and Gestational Diabetes Mellitus: A Prospective Cohort Study in China. Front. Endocrinol. 2019, 10, 262. [Google Scholar] [CrossRef]
- Mustieles, V.; Ocón-Hernandez, O.; Mínguez-Alarcón, L.; Dávila-Arias, C.; Pérez-Lobato, R.; Calvente, I.; Arrebola, J.P.; Vela-Soria, F.; Rubio, S.; Hauser, R.; et al. Bisphenol A and reproductive hormones and cortisol in peripubertal boys: The INMA-Granada cohort. Sci. Total Environ. 2018, 618, 1046–1053. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, J.; Wu, C. Determination of Endocrine Disruption Potential of Bisphenol A Alternatives in Food Contact Materials Using In Vitro Assays: State of the Art and Future Challenges. J. Agric. Food Chem. 2019, 67, 12613–12625. [Google Scholar] [CrossRef]
- de la Riviere, M.-E.L.; Wu, L.; Gayet, M.; Bousquet, M.; Buron, C.; Vignault, C.; Téteau, O.; Desmarchais, A.; Maillard, V.; Uzbekova, S.; et al. Cumulative and potential synergistic effects of seven different bisphenols on human granulosa cells in vitro? Environ. Pollut. 2023, 330, 121818. [Google Scholar] [CrossRef] [PubMed]
- Haggard, D.E.; Karmaus, A.L.; Martin, M.T.; Judson, R.S.; Setzer, R.W.; Friedman, K.P. High-Throughput H295R Steroidogenesis Assay: Utility as an Alternative and a Statistical Approach to Characterize Effects on Steroidogenesis. Toxicol. Sci. 2018, 162, 509–534. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štefunková, N.; Greifová, H.; Jambor, T.; Tokárová, K.; Zuščíková, L.; Bažány, D.; Massányi, P.; Capcarová, M.; Lukáč, N. Comparison of the Effect of BPA and Related Bisphenols on Membrane Integrity, Mitochondrial Activity, and Steroidogenesis of H295R Cells In Vitro. Life 2024, 14, 3. https://doi.org/10.3390/life14010003
Štefunková N, Greifová H, Jambor T, Tokárová K, Zuščíková L, Bažány D, Massányi P, Capcarová M, Lukáč N. Comparison of the Effect of BPA and Related Bisphenols on Membrane Integrity, Mitochondrial Activity, and Steroidogenesis of H295R Cells In Vitro. Life. 2024; 14(1):3. https://doi.org/10.3390/life14010003
Chicago/Turabian StyleŠtefunková, Nikola, Hana Greifová, Tomáš Jambor, Katarína Tokárová, Lucia Zuščíková, Denis Bažány, Peter Massányi, Marcela Capcarová, and Norbert Lukáč. 2024. "Comparison of the Effect of BPA and Related Bisphenols on Membrane Integrity, Mitochondrial Activity, and Steroidogenesis of H295R Cells In Vitro" Life 14, no. 1: 3. https://doi.org/10.3390/life14010003