
Gut Microbiome Composition in Patients with Chronic Urticaria: A Review of Current Evidence and Data
 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
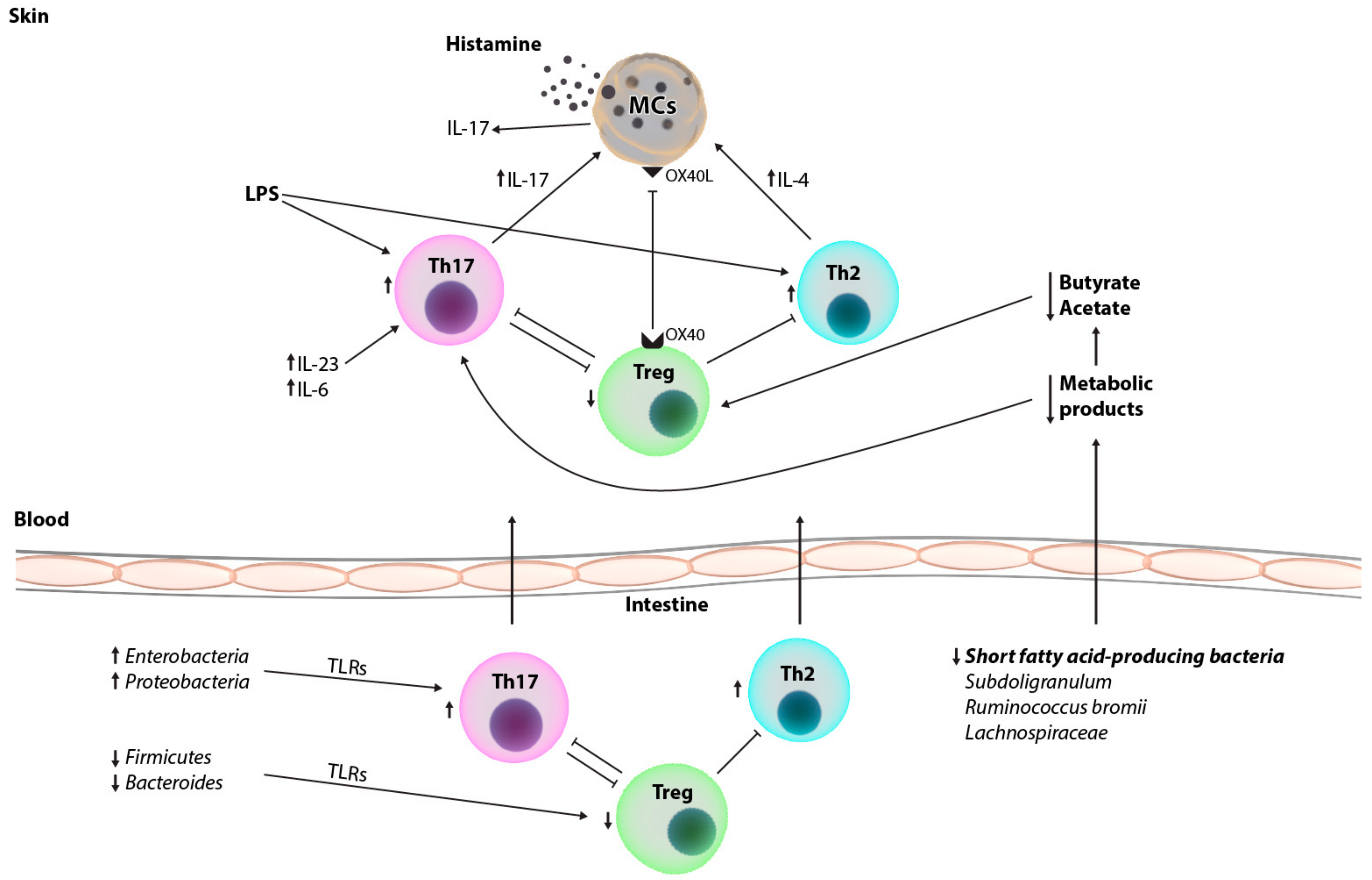
2. Immunopathogenesis of Chronic Urticarial and Immune Changes in the Peripheral Blood and Skin of Patients
3. Characteristics of the Gut Microbiome of Healthy Individuals
4. Approaches to Gut Microbiota Analysis
5. Gut Microbiome in Patients with Chronic Spontaneous Urticaria
6. Therapeutic Implications
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zuberbier, T.; Abdul Latiff, A.H.; Abuzakouk, M.; Aquilina, S.; Asero, R.; Baker, D.; Ballmer-Weber, B.; Bangert, C.; Ben-Shoshan, M.; Bernstein, J.A.; et al. The international EAACI/GA2LEN/EuroGuiDerm/APAAACI guideline for the definition, classification, diagnosis, and management of urticaria. Allergy 2022, 77, 34–766. [Google Scholar] [CrossRef] [PubMed]
- Zuberbier, T.; Aberer, W.; Asero, R.; Abdul Latiff, A.H.; Baker, D.; Ballmer-Weber, B.; Bernstein, J.A.; Bindslev-Jensen, C.; Brzoza, Z.; Buense Bedrikow, R.; et al. The EAACI/GA2LEN/EDF/WAO Guideline for the definition, classification, diagnosis and management of urticaria. Allergy 2018, 73, 1393–1414. [Google Scholar] [CrossRef] [PubMed]
- Gonçalo, M.; Gimenéz-Arnau, A.; Al-Ahmad, M.; Ben-Shoshan, M.; Bernstein, J.A.; Ensina, L.F.; Bernstein, J.A.; Bindslev-Jensen, C.; Brzoza, Z.; Buense Bedrikow, R.; et al. The global burden of chronic urticaria for the patient and society. Br. J. Dermatol. 2021, 184, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Itakura, A.; Tani, Y.; Kaneko, N.; Hide, M. Impact of chronic urticaria on quality of life and work in Japan: Results of a Real-World study. J. Dermatol. 2018, 45, 963–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabelo-Filardi, R.; de Oliveira, R.D.; Campos, R.A. Parameters associated with chronic spontaneous urticaria duration and severity: A systematic review. Int. Arch. Allergy Immunol. 2013, 161, 197–204. [Google Scholar] [CrossRef]
- Guillén-Aguinaga, S.; Jáuregui Presa, I.; Aguinaga-Ontoso, E.; Guillén-Grima, F.; Ferrer, M. Updosing nonsedating antihistamines in patients with chronic spontaneous urticaria: A systematic review and meta-analysis. Br. J. Dermatol. 2016, 175, 1153–1165. [Google Scholar] [CrossRef]
- Joshi, S.; Khan, D. The expanding field of biologics in the management of chronic urticaria. J. Allergy Clin. Immunol. In Pract. 2017, 5, 1489–1499. [Google Scholar] [CrossRef]
- Maurer, M.; Staubach, P.; Raap, U.; Richter-Huhn, G.; Bauer, A.; Ruëff, F.; Jakob, T.; Yazdi, A.S.; Mahler, V.; Wagner, N.; et al. H1-antihistamine-refractory chronic spontaneous urticaria: It’s worse than we thought—First results of the multicenter real-life AWARE study. Clin. Exp. Allergy 2017, 47, 684–692. [Google Scholar] [CrossRef]
- Min, T.K.; Saini, S.S. Emerging therapies in chronic spontaneous urticaria. Allergy Asthma Immunol. Res. 2019, 11, 470–481. [Google Scholar] [CrossRef]
- Amin, P.; Levin, L.; Holmes, S.; Picard, J.; Bernstein, J. Investigation of patient-specific characteristics associated with treatment outcomes for chronic urticaria. J. Allergy Clin. Immunol. In Pract. 2015, 3, 400–407. [Google Scholar] [CrossRef]
- Lu, T.; Chen, Y.; Guo, Y.; Sun, J.; Shen, W.; Yuan, M.; Zhang, S.; He, P.; Jiao, X. Altered gut microbiota diversity and composition in chronic urticaria. Dis. Markers 2019, 2019, 6417471. [Google Scholar] [CrossRef] [Green Version]
- Nabizadeh, E.; Jazani, N.H.; Bagheri, M.; Shahabi, S. Association of altered gut microbiota composition with chronic urticaria. Ann. Allergy Asthma Immunol. 2017, 119, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Guo, S.; He, H.; Gong, L.; Cui, H. Gut microbiome and serum metabolome analyses identify unsaturated fatty acids and butanoate metabolism induced by gut microbiota in patients with chronic spontaneous urticaria. Front. Cell Infect. Microbiol. 2020, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The gut microbiota and inflammation: An overview. Int. J. Environ. Res. Public Health 2020, 17, 7618. [Google Scholar] [CrossRef]
- Chen, Q.; Zhong, H.; Chen, W.C.; Zhai, Z.; Zhou, Z.; Song, Z.; Hao, F. Different expression patterns of plasma Th1-, Th2-, Th17- and Th22-related cytokines correlate with serum autoreactivity and allergen sensitivity in chronic spontaneous urticaria. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 441–448. [Google Scholar] [CrossRef]
- Bansal, C.J.; Bansal, A.S. Stress, pseudoallergens, autoimmunity, infection and inflammation in chronic spontaneous urticaria. Allergy Asthma Clin. Immunol. 2019, 15, 56. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yi, W.; He, L.; Luo, S.; Wang, J.; Jiang, L.; Long, H.; Zhao, M.; Lu, Q. Abnormalities in gut microbiota and metabolism in patients with chronic spontaneous urticaria. Front. Immunol. 2021, 12, 691304. [Google Scholar] [CrossRef]
- Bracken, S.J.; Abraham, S.; MacLeod, A.S. Autoimmune theories of chronic spontaneous urticaria. Front. Immunol. 2019, 10, 627. [Google Scholar] [CrossRef]
- Auyeung, P.; Mittag, D.; Hodgkin, P.D.; Harrison, L.C. Autoreactive T cells in chronic spontaneous urticaria target the IgE Fc receptor Iα subunit. J. Allergy Clin. Immunol. 2016, 138, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Arnau, A.M.; DeMontojoye, L.; Asero, R.; Cugno, M.; Kulthanan, K.; Yanase, Y.; Hide, M.; Kaplan, A.P. The pathogenesis of chronic spontaneous urticaria: The role of infiltrating cells. J. Allergy Clin. Immunol. Pract. 2021, 9, 2195–2208. [Google Scholar] [CrossRef]
- Zhou, B.; Li, J.; Liu, R.; Zhu, L.; Peng, C. The role of crosstalk of immune cells in pathogenesis of chronic spontaneous urticaria. Front. Immunol. 2022, 13, 879754. [Google Scholar] [CrossRef] [PubMed]
- Prosty, C.; Gabrielli, S.; Ben-Shoshan, M.; Le, M.; Giménez-Arnau, A.M.; Litvinov, I.V.; Lefrançois, P.; Netchiporouk, E. In silico identification of immune cell-types and pathways involved in chronic spontaneous urticaria. Front. Med. 2022, 9, 926753. [Google Scholar] [CrossRef] [PubMed]
- Atwa, M.; Emara, A.; Youssef, N.; Bayoumy, N. Serum concentration of IL-17, IL-23 and TNF-α among patients with chronic spontaneous urticaria: Association with disease activity and autologous serum skin test. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Sabag, D.A.; Matanes, L.; Bejar, J.; Toubi, E.; Maurer, M.; Vadasz, Z. Interleukin-17 is a potential player and treatment target in severe chronic spontaneous urticaria. Clin. Exp. Allergy 2020, 50, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Toubi, E.; Vadasz, Z. The emerging role of IL-17 in the immune-pathogenesis of chronic spontaneous urticaria. Immunotargets Ther. 2020, 9, 217–223. [Google Scholar] [CrossRef]
- Sun, R.S.; Sui, J.F.; Chen, X.H.; Ran, X.Z.; Yang, Z.F.; Guan, W.D.; Yang, T. Detection of CD4+ CD25+ FOXP3+ regulatory T cells in peripheral blood of patients with chronic autoimmune urticaria. Aust. J. Dermatol. 2011, 52, e15–e18. [Google Scholar] [CrossRef]
- Arshi, S.; Babaie, D.; Nabavi, M.; Tebianian, M.; Ghalehbaghi, B.; Jalali, F.; Ahmadvand, A.; Gholami, R. Circulating level of CD4+ CD25+ FOXP3+ T cells in patients with chronic urticaria. Int. J. Dermatol. 2014, 53, e561–e566. [Google Scholar] [CrossRef]
- Gri, G.; Piconese, S.; Frossi, B.; Manfroi, V.; Merluzzi, S.; Tripodo, C.; Viola, A.; Odom, S.; Rivera, J.; Colombo, M.P.; et al. CD4+CD25+ regulatory T cells suppress mast cell degranulation and allergic responses through OX40-OX40L interaction. Immunity 2008, 29, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Piconese, S.; Gri, G.; Tripodo, C.; Musio, S.; Gorzanelli, A.; Frossi, B.; Pedotti, R.; Pucillo, C.E.; Colombo, M.P. Mast cells counteract regulatory T-cell suppression through interleukin-6 and OX40/OX40L axis toward Th17-cell differentiation. Blood 2009, 114, 2639–2648. [Google Scholar] [CrossRef]
- Egert, M.; Simmering, R.; Riedel, C.U. The association of the skin microbiota with health, immunity, and disease. Clin. Pharmacol. Ther. 2017, 102, 62–69. [Google Scholar] [CrossRef]
- Ferček, I.; Lugović-Mihić, L.; Tambić-Andrašević, A.; Ćesić, D.; Grginić, A.G.; Bešlić, I.; Mravak-Stipetić, M.; Mihatov-Štefanović, I.; Buntić, A.M.; Čivljak, R. Features of the skin microbiota in common inflammatory skin diseases. Life 2021, 11, 962. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 15, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Polkowska-Pruszyńska, B.; Gerkowicz, A.; Krasowska, D. The gut microbiome alterations in allergic and inflammatory skin diseases—An update. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 455–464. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Burgueño, J.F.; Abreu, M.T. Epithelial Toll-like receptors and their role in gut homeostasis and disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 263–278. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 9th ed.; Elsevier Health Sciences: Philadelphia, PA, USA, 2016. [Google Scholar]
- Rosenbaum, J.T.; Silverman, G.J. The microbiome and systemic lupus erythematosus. N. Engl. J. Med. 2018, 378, 2236–2237. [Google Scholar] [CrossRef]
- Coit, P.; Sawalha, A.H. The human microbiome in rheumatic autoimmune diseases: A comprehensive review. Clin. Immunol. 2016, 170, 70–79. [Google Scholar] [CrossRef]
- Abdel-Gadir, A.; Stephen-Victor, E.; Gerber, G.K.; Noval, R.M.; Wang, S.; Harb, H.; Wang, L.; Li, N.; Crestani, E.; Spielman, S.; et al. Microbiota therapy acts via a regulatory T cell Myd88/Rorγt pathway to suppress food allergy. Nat. Med. 2019, 25, 1164–1174. [Google Scholar] [CrossRef]
- Stefanovic, N.; Flohr, C.; Irvine, A.D. The exposome in atopic dermatitis. Allergy 2020, 75, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; O’mahony, L. The importance of social networks-an ecological and evolutionary framework to explain the role of microbes in the aetiology of allergy and asthma. Allergy 2019, 74, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Chu, Z.; Shi, L.; Geng, S.; Guo, K. Gut microbiome alterations and functional prediction in chronic spontaneous urticaria patients. J. Microbiol. Biotechnol. 2021, 31, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Godlewska, U.; Brzoza, P.; Kwiecień, K.; Kwitniewski, M.; Cichy, J. Metagenomic studies in inflammatory skin diseases. Curr. Microbiol. 2020, 77, 3201–3212. [Google Scholar] [CrossRef] [PubMed]
- Wilmanski, T.; Rappaport, N.; Diener, C.; Gibbons, S.M.; Price, N.D. From taxonomy to metabolic output: What factors define gut microbiome health? Gut Microbes 2021, 13, 1907270. [Google Scholar] [CrossRef]
- De Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Wang, J.W.; Kuo, C.H.; Kuo, F.C.; Wang, Y.K.; Hsu, W.H.; Yu, F.J.; Hu, H.M.; Hsu, P.I.; Wang, J.Y.; Wu, D.C. Fecal microbiota transplantation: Review and update. J. Formos. Med. Assoc. 2019, 118, S23–S31. [Google Scholar] [CrossRef]
- Yang, Z.D.; Guo, Y.S.; Huang, J.S.; Gao, Y.F.; Peng, F.; Xu, R.Y.; Su, H.H.; Zhang, P.J. Isomaltulose exhibits prebiotic activity, and modulates gut microbiota, the production of short chain fatty acids, and secondary bile acids in rats. Molecules 2021, 26, 2464. [Google Scholar] [CrossRef]
- Horvat, S.; Rupnik, M. Interactions between Clostridioides difficile and fecal microbiota in in vitro batch model: Growth, sporulation, and microbiota changes. Front. Microbiol. 2018, 9, 1633. [Google Scholar] [CrossRef]
- Dekkers, K.F.; Sayols-Baixeras, S.; Baldanzi, G.; Nowak, C.; Hammar, U.; Nguyen, D.; Varotsis, G.; Brunkwall, L.; Nielsen, N.; Eklund, A.C.; et al. An online atlas of human plasma metabolite signatures of gut microbiome composition. Nat. Commun. 2022, 13, 5370. [Google Scholar] [CrossRef]
- Putignani, L.; Del Chierico, F.; Vernocchi, P.; Cicala, M.; Cucchiara, S.; Dallapiccola, B.; Dysbiotrack Study Group. Gut microbiota dysbiosis as risk and premorbid factors of IBD and IBS along the childhood-adulthood transition. Inflamm. Bowel. Dis. 2016, 22, 487–504. [Google Scholar] [CrossRef] [Green Version]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut microbiota profiling: Metabolomics based approach to unravel compounds affecting human health. Front. Microbiol. 2016, 7, 1144. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.X.; Wang, S.Y.; Kuo, C.H.; Tsai, I.L. Metabolome analysis for investigating host-gut microbiota interactions. J. Formos. Med. Assoc. 2019, 118, S10–S22. [Google Scholar] [CrossRef]
- Mojsak, P.; Rey-Stolle, F.; Parfieniuk, E.; Kretowski, A.; Ciborowski, M. The role of gut microbiota (GM) and GM-related metabolites in diabetes and obesity. A review of analytical methods used to measure GM-related metabolites in fecal samples with a focus on metabolites’ derivatization step. J. Pharm. Biomed. Anal. 2020, 191, 113617. [Google Scholar] [CrossRef]
- Liu, R.; Peng, C.; Jing, D.; Xiao, Y.; Zhu, W.; Zhao, S.; Zhang, J.; Chen, X.; Li, J. Biomarkers of gut microbiota in chronic spontaneous urticaria and symptomatic dermographism. Front. Cell Infect. Microbiol. 2021, 11, 703126. [Google Scholar] [CrossRef]
- Kemp, P.F.; Aller, J.Y. Bacterial diversity in aquatic and other environments: What 16S rDNA libraries can tell us. FEMS Microbiol. Ecol. 2004, 47, 161–177. [Google Scholar] [CrossRef]
- Hilty, M.; Burke, C.; Pedro, H.; Cardenas, P.; Bush, A.; Bossley, C.; Davies, J.; Ervine, A.; Poulter, L.; Pachter, L.; et al. Disordered microbial communities in asthmatic airways. PLoS ONE 2010, 5, e8578. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.J.; Nelson, C.E.; Brodie, E.L.; Desantis, T.Z.; Baek, M.S.; Liu, J.; Woyke, T.; Allgaier, M.; Bristow, J.; Wiener-Kronish, J.P.; et al. Airway microbiota and bronchial hyperresponsiveness in patients with suboptimally controlled asthma. J. Allergy Clin. Immunol. 2011, 127, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Loo, E.X.L.; Chew, L.J.M.; Zulkifli, A.B.; Ta, L.D.H.; Kuo, I.C.; Goh, A.; Teoh, O.H.; Van Bever, H.; Gluckman, P.D.; Yap, F.; et al. Comparison of microbiota and allergen profile in house dust from homes of allergic and non-allergic subjects- results from the GUSTO study. World Allergy Organ. J. 2018, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.; Turnbaugh, P.; Klein, S.; Gordon, J. Microbial Ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Turnbaugh, P.; Ley, R.; Mahowald, M.; Magrini, V.; Mardis, E.; Gordon, J. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Berry, D. The emerging view of firmicutes as key fibre degraders in the human gut. Environ. Microbiol. 2016, 18, 2081–2083. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.; Wexler, A.; Harding, B.; Whitney, J.; Bohn, A.; Goo, Y.; Tran, B.Q.; Barry, N.A.; Zheng, H.; Peterson, S.B. A type VI secretion-related pathway in bacteroidetes mediates interbacterial antagonism. Cell Host Microbe 2014, 16, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezazadeh, A.; Shahabi, S.; Bagheri, M.; Nabizadeh, E.; Jazani, N.H. The protective effect of Lactobacillus and Bifidobacterium as the gut microbiota members against chronic urticaria. Int. Immunopharmacol. 2018, 59, 168–173. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Schmetzer, O.; Lakin, E.; Topal, F.A.; Preusse, P.; Freier, D.; Church, M.K.; Maurer, M. IL-24 is a common and specific autoantigen of IgE in patients with chronic spontaneous urticaria. J. Allergy Clin. Immunol. 2018, 142, 876–882. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, Y.; Inokuchi, R.; Harada, S.; Aoki, K.; Ishii, Y.; Shinohara, K. Bacterial meningitis caused by hypervirulent Klebsiella pneumoniae capsular genotype K54 with development of granuloma-like nodal enhancement in the brain during the subacute phase. Intern. Med. 2017, 56, 373–376. [Google Scholar] [CrossRef] [Green Version]
- Bao, R.; Hesser, L.; He, Z.; Zhou, X.; Nadeau, K.; Nagler, C. Fecal microbiome and metabolome differ in healthy and food-allergic twins. J. Clin. Investig. 2021, 131, e141935. [Google Scholar] [CrossRef]
- Song, Y.; Dan, K.; Yao, Z.; Yang, X.; Chen, B.; Hao, F. Altered gut microbiota in H1-antihistamine-resistant chronic spontaneous urticaria associates with systemic inflammation. Front. Cell Infect. Microbiol. 2022, 12, 831489. [Google Scholar] [CrossRef]
- Liu, C.W.; Cheng, Y.C.; Yu, Y.T.; Huang, Y.C. Roles of gut-microbiota and probiotics in chronic urticaria: A systematic review and meta-analysis. Australas. J. Dermatol. 2022, 63, e166–e170. [Google Scholar] [CrossRef]
- Liu, R.; Peng, C.; Jing, D.; Xiao, Y.; Zhu, W.; Zhao, S.; Zhang, J.; Chen, X.; Li, J. Lachnospira is a signature of antihistamine efficacy in chronic spontaneous urticaria. Exp. Dermatol. 2022, 31, 242–247. [Google Scholar] [CrossRef]
- Meehan, C.J.; Beiko, R.G. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhang, S.; Liu, X.; Huo, Y.; Su, B.; Li, X. Effects of a probiotic intervention on Escherichia Coli and high-fat diet-induced intestinal microbiota imbalance. Appl. Microbiol. Biotechnol. 2020, 104, 1243–1257. [Google Scholar] [CrossRef]
- Widhiati, S.; Purnomosari, D.; Wibawa, T.; Soebono, H. The role of gut microbiome in inflammatory skin disorders: A systematic review. Dermatol. Rep. 2021, 14, 9188. [Google Scholar] [CrossRef]
- Kim, S.K.; Guevarra, R.B.; Kim, Y.T.; Kwon, J.; Kim, H.; Cho, J.H.; Kim, H.B.; Lee, J.H. Role of probiotics in human gut microbiome-associated diseases. J. Microbiol. Biotechnol. 2019, 29, 1335–1340. [Google Scholar] [CrossRef]
- Atefi, N.; Fallahpour, M.; Sharifi, S.; Ghassemi, M.; Roohaninasab, M.; Goodarzi, A. Probiotic as an adjuvant therapy in chronic urticaria: A blinded randomized controlled clinical trial. Eur. Ann. Allergy Clin. Immunol. 2022, 54, 123–130. [Google Scholar] [CrossRef]
- Chen, B.; Sun, L.; Zhang, X. Integration of microbiome and epigenome to decipher the pathogenesis of autoimmune diseases. J. Autoimmun. 2017, 83, 31–42. [Google Scholar] [CrossRef]
- Podder, I.; Jaiswal, S.; Das, A. Dietary strategies for chronic spontaneous urticaria: An evidence-based review. Int. J. Dermatol. 2021, 26. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Kirjavainen, P.V.; Apostolou, E.; Arvola, T.; Salminen, S.J.; Gibson, G.R.; Isolauri, E. Characterizing the composition of intestinal microflora as a prospective treatment target in infant allergic disease. FEMS Immunol. Med. Microbiol. 2001, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fuchs-Tarlovsky, V.; Marquez-Barba, M.F.; Sriram, K. Probiotics in dermatologic practice. Nutrition 2016, 32, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Matszaki, T.; Chin, J. Modulating immune responses with probiotic bacteria. Immunol. Cell Biol. 2000, 78, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Von der Weid, T.; Bulliard, C.; Schirin, E.J. Induction by a lactic acid bacterium of a population of CD4+ T cells with low proliferative capacity that produce transforming growth factor b and interleukin-10. Clin. Diagn. Lab. Immunol. 2001, 8, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Nettis, E.; Di Leo, E.; Pastore, A.; Distaso, M.; Zaza, I.; Vacca, M.; Macchia, L.; Vacca, A. Probiotics and refractory chronic spontaneous urticaria. Eur. Ann. Allergy Clin. Immunol. 2016, 48, 182–187. [Google Scholar]
- Drago, L.; Nicola, L.; Iemoli, E.; Banfi, G.; De Vecchi, E. Strain-dependent release of cytokines modulated by Lactobacillus salivarius human isolates in an in vitro model. BMC Res. Notes 2010, 3, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Ćesić, D.; Lugović-Mihić, L.; Ferček, I.; Gverić Grginić, A.; Jelić, M.; Bešlić, I.; Tambić Andrašević, A. Salivary microbiota is significantly less diverse in patients with chronic spontaneous urticaria compared to healthy controls: Preliminary results. Life 2021, 11, 1329. [Google Scholar] [CrossRef]
- Bi, X.D.; Lu, B.Z.; Pan, X.X.; Liu, S.; Wang, J.Y. Adjunct therapy with probiotics for chronic urticaria in children: Randomised placebo-controlled trial. Allergy Asthma Clin. Immunol. 2021, 17, 39. [Google Scholar] [CrossRef]


{kind=link}
| Examinees | Methods | Key Results | CU Patients Versus Healthy Controls (HCs) | Limitations | References | |
|---|---|---|---|---|---|---|
| Increase | Decrease | |||||
| 20 CU patients compared to 20 age- and sex-matched HCs | Bacterial genomic DNA extracted from stool samples and sequenced by PCR; bacterial amounts determined by qPCR. | The frequency and relative amounts of A. muciniphila, C. leptum, and F. prausnitzii were significantly higher in HCs than in those with CU. | Family Enterobacteriaceae | Species Akkermansia muciniphila, Clostridium leptum, Faecalibacterium prausnitzii (p < 0.001, p < 0.01, and p < 0.05) | Small number of examinees, more sophisticated analysing methods not included | [12] Nabizadeh et al., 2017. |
| 20 CU patients compared to 20 age- and sex-matched HCs | Fecal samples analysed by PCR for frequency and bacterial load of Lactobacillus and Bifidobacterium genera. | The relative amounts of Lactobacillus and Bifidobacterium were significantly higher in fecal samples from HCs compared to CU patients. | Genus Lactobacillus, Bifidobacterium (p = 0.038 and 0.039) | Small number of examinees, more sophisticated analysing methods not included | [57] Rezazadeh et al. 2018. | |
| 10 CU patients compared to 10 HCs | Intestinal microbiome was analysed using 16S rRNA sequencing. | Significantly different microbial composition was observed between CU patients and HCs at the genus level. | Phylum Proteobacteria, Actinobacteria Order Enterobacteriales, Lactobacillales, Pseudomonadales; Genus Veillonella, Sutterella, Streptococcus, Clostridium, and Escherichia; Species E.coli | Phylum Bacteroidetes Genus Faecalibacterium, Prevotella, Lachnobacterium; Species Faecalibacterium prausnitzii, Prevotella copri, Bacteroides fragilis, Bacteroides plebeius | Very small number of examinees | [11] Lu et al., 2019. |
| 100 CSU patients compared to 100 HCs | Fecal and blood samples were analysed using 16S rRNA gene sequencing and untargeted metabolomics, respectively. | The CSU group exhibited differences at the phyla and genera levels with decreased alpha diversity. The serum metabolome analysis revealed changes in unsaturated fatty acids and the butanoate metabolism pathway. | Family Enterobacteriaceae | Phylum Firmicutes; Genus Bacteroides, Faecalibacterium, Bifidobacterium, Lactobacillus, Ruminococcaceae | Small number of examinees | [13] Wang et al., 2020. |
| 39 CSU patients compared to 40 HCs | Fecal samples were analysed using 16S rRNA gene sequencing and untargeted metabolomics. | A significant difference in beta diversity with no significant differences in alpha diversity between CSU patients and HCs (p < 0.05). Gut metabolomics showed decreased isobutyric acid in CSU patients. | Phylum Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria Genus Lactobacillus, Turicibacter, Lachnobacterium | Genus Phascolarctobacterium | Single-center study, small number of examinees, confounding factors (age and diet) not taken into account | [17] Wang et al., 2021. |
| 20 CSU patients compared to 20 age- and sex-matched HCs | Fecal microbial composition was analysed using 16S rRNA sequencing. | Beta diversity significantly differed between the two groups while alpha diversity did not. | Phylum Firmicutes, Bacteroidetes, Proteobacteria (p = 0.03), Verrucomicrobia Class Bacilli (p = 0.04) Order Enterobacterales (p = 0.03) Family Enterobacteriaceae (p = 0.03) | Genus Megamonas, Megasphaera, Dialister (all p < 0.05) | Small number of examinees | [44] Zhang et al., 2021. |
| 25 CSD patients compared to 25 age- and sex-matched HCs | Fecal samples were analysed by 16S rRNA sequencing, additionally verified by qPCR. | There were significant differences in both alpha and beta diversity in different indices with Subdoligranulum and Ruminococcus bromii (short chain fatty acid producing bacteria) as the gut microbiota biomarkers in CSD. | Phylum Fusobacteria Class Gammaproteobacteria, Fusobacteria Order Enterobacteria, Fusobacteria Family Enterobacteriaceae, Fusobacteraceae, Peptostreptococcaceae, Streptococcaceae Genus Klebsiella | Phylum Firmicutes Class Clostridia, Alphaproteobacterial, Deltaproteobacteria Order Clostridiales, Rhodospirillales, Caulobacterales, Desulfovibrionales Family Ruminococceae, Rikenellaceae, Muribaculaceae, Christensenellaceae, Caulobacteraceae Genus/species Subdoligranulum, Ruminococcus bromii | Small number of examinees | [56] Liu et al., 2021. |
| 25 CSU patients with nsAH resistance, 19 CSU patients without nsAH resistance and 19 HCs | 16S rRNA sequencing of the intestinal microbiome. | Much lower alpha diversity and evenness was observed in CSU patients with nsAH resistance than in those without (p < 0.05), who together with HCs showed almost no change in genera bacterium. | Genus Prevotella, Megamonas, Escherichia, Succinivibrio, Klebsiella, Colidextribacter (all p < 0.05) (in nsAH-resistant patients) | Genus Blautia, Alistipes, Anaerostipes, Lachnospira (all p < 0.05) (in nsAH-resistant patients) | Single-center study, small number of examinees, nsAHs other than levocetirizine not included | [58] Song et al., 2022. |
| 15 responders and 15 non-responders to nsAH with CSU, with an extended cohort of another 30 responders and 30 non-responders | Gut microbiota (fecal samples) were analysed using 16S rDNA sequencing (verified by qPCR). | The abundance of Lachnospira in responders was higher than in non-responders with possible prediction of the AH efficacy in CSU patients. | Family Lachnospiraceae Genus Lachnospira | Small number of examinees | [59] Liu et al., 2022. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krišto, M.; Lugović-Mihić, L.; Muñoz, M.; Rupnik, M.; Mahnic, A.; Ozretić, P.; Jaganjac, M.; Ćesić, D.; Kuna, M. Gut Microbiome Composition in Patients with Chronic Urticaria: A Review of Current Evidence and Data. Life 2023, 13, 152. https://doi.org/10.3390/life13010152
Krišto M, Lugović-Mihić L, Muñoz M, Rupnik M, Mahnic A, Ozretić P, Jaganjac M, Ćesić D, Kuna M. Gut Microbiome Composition in Patients with Chronic Urticaria: A Review of Current Evidence and Data. Life. 2023; 13(1):152. https://doi.org/10.3390/life13010152
Chicago/Turabian StyleKrišto, Mirela, Liborija Lugović-Mihić, Melba Muñoz, Maja Rupnik, Aleksander Mahnic, Petar Ozretić, Morana Jaganjac, Diana Ćesić, and Matea Kuna. 2023. "Gut Microbiome Composition in Patients with Chronic Urticaria: A Review of Current Evidence and Data" Life 13, no. 1: 152. https://doi.org/10.3390/life13010152