

Microbiota of the Colonic Diverticula in the Complicated Form of Diverticulitis: A Case Report
by
 ,
,
Dina Yarullina
1,*,
Yuliia Pankratova
2,
Olga Karaseva
1,
Tatiana Grigoryeva
1 and
Oleg Karpukhin
2
1
Institute of Fundamental Medicine and Biology, Kazan Federal University, 420008 Kazan, Russia
2
Department of Surgical Diseases, Kazan State Medical University, 420012 Kazan, Russia
*
Author to whom correspondence should be addressed.
Life 2022, 12(12), 2129; https://doi.org/10.3390/life12122129
Submission received: 7 November 2022
/
Revised: 12 December 2022
/
Accepted: 14 December 2022
/
Published: 16 December 2022
(This article belongs to the Special Issue Interactions between Microorganisms, Their Environment and Host)
Abstract
:Intestinal microbiota appears to be implicated in the pathogenesis of diverticular disease. We present the case of a patient with diverticular colon disease complicated by a pelvic abscess. During the successful surgical treatment, two specimens were taken from the resected colon segment for the microbiota analysis: an inflamed and perforated diverticulum and a diverticulum without signs of inflammation. Culturing and 16S rRNA gene sequencing revealed significant changes in the microbial community structure and composition associated with the acute inflammation and perforation of the colonic diverticulum. The characteristics that are usually associated with the inflammatory process in the gut, namely reduced microbial diversity and richness, decreased Firmicutes-to-Bacteroidetes (F/B) ratio, depletion of butyrate-producing bacteria, and Enterobacteriaceae blooming, were more pronounced in the non-inflamed diverticulum rather than in the adjacent inflamed and perforated one. This is the first study of the intraluminal microbiota of the diverticular pockets, which is more relevant to the etiology of diverticular disease than mucosa-associated microbiota via biopsies and luminal microbiota via fecal samples.
1. Introduction
The incidence of diverticular disease (DD) is steadily increasing in the 21st century. In recent times, exacerbation of DD has become a frequent gastrointestinal complication of COVID-19 [1]. The clinical symptoms of DD arise as a result of inflammation in the intestinal mucosa of the diverticula, which are small outpouchings of the colon wall [2]. This inflammation may be induced by the local ischemia consequent of the compression of the vessels in the submucosal layer of the diverticula as well as microperforations of the diverticula walls injured by the dense fecal stones [3]. Changes in the intestinal microbiota composition may be implicated in the persistence of the inflammatory process and hence the progression of DD [4]. There have been few reports on the association between microbiota and diverticulosis with contradictory results [5]. The possible reason for the vagueness about the role of the intestinal microbiota in DD is that these studies were performed on patients with different disease severity (asymptomatic diverticulosis, Symptomatic Uncomplicated Diverticular Disease (SUDD), and diverticulitis or Segmental Colitis Associated with Diverticulosis (SCAD)) and assessed different kinds of microbiota (mucosa-associated microbiota via biopsies and luminal microbiota via fecal samples). Most of the studies declared no substantial differences in the gut microbiota composition between patients with diverticulosis and controls [6,7,8] except for non-significant changes in the relative abundance of some taxa: Proteobacteria and Comamonadaceae [7], Akkermansia muciniphila [8], and Clostridium cluster IV [9]. Interestingly, the mucosal microbiota was significantly different between “diseased” (affected by diverticulitis) and adjacent “healthy” tissue. The microbiota associated with a region of the sigmoid colon chronically affected by diverticulitis exhibited a relative overabundance of Microbacteriaceae and Ascomycota [10]. In patients with SUDD biopsies taken in the gut segment affected by diverticula, the biopsies were characterized by a higher abundance of Enterobacteriaceae and a lower abundance of Bacteroides/Prevotella group and Akkermansia compared with biopsies taken from the distant site. Notably, in control subjects without colonic diverticula and patients with asymptomatic diverticulosis, no differences between the mucosal microbiota of the diverticular or non-diverticular regions were detected [9]. These findings are evidence that we should consider diverticula as a unique niche that selectively promote the development of specific microbial communities, which may play a role in the progression of diverticulosis towards symptomatic forms [11]. The aim of this study is to characterize the juxta-mucosal microbiota in the lumens of diverticula from a patient who has undergone surgery for a pelvic abscess and diverticular perforation. Herein, using a conventional culture method and 16S rRNA-based sequencing approach, we assessed the microbiota composition of the inflamed perforated diverticulum (ID) and an adjacent non-inflamed diverticulum (NID) from the same colonic segment.
2. Case Report
A 52-year-old man was admitted at the Emergency Department of our hospital with complaints of pain in the lower abdomen, fever, chills, liquid stool 2–3 times per day, and general weakness over the previous week. The patient used rifaximin (400 mg orally twice daily) and metronidazole (500 mg orally three times daily) for self-medication but did not feel improvement. His medical history revealed an episode of acute diverticulitis with subsequent conservative treatment five years before this admission. The patient also had a history of psoriasis and hypertension (stage 2, risk 2).
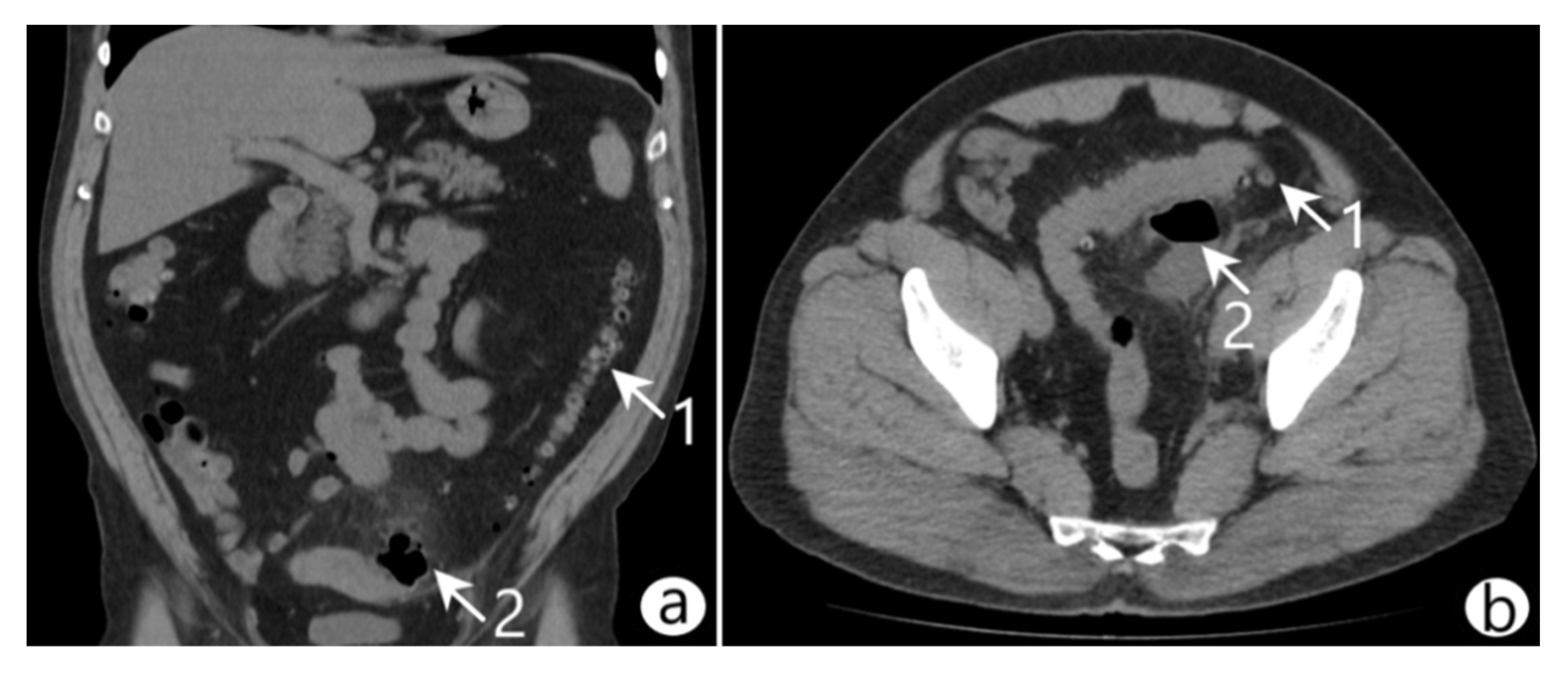
On admission, vital signs were as follows: body temperature 37.8 °C, blood pressure 110/70 mmHg, heart rate 86 beats/min, and respiratory rate 20/min. Laboratory blood tests showed the following: white blood cell (WBC) count 14.1 × 109/L with neutrophilic granulocyte percentage 74.5%, red blood cell (RBC) count 4.3 × 1012/L, platelets 21.8 × 1010/L, hemoglobin (HGB) 11.2 g/dL, and hematocrit (HCT) 43.3%. The abdominal physical examination revealed symmetry, softness in palpation, no swelling, pain in the left side of the mesogastrium, and no symptoms of peritoneal irritation. The abdominal ultrasound indicated edema and thickening of the sigmoidal wall and a pericolic abscess. Computed tomography (CT) with intravenous contrast revealed an inflamed diverticulum in the sigmoid colon, a pelvic abscess (45 × 35 × 56 mm) containing gas and fluid, and infiltration of the surrounding adipose tissue (Figure 1). A diagnosis of diverticular colon disease complicated by the formation of a pelvic abscess (Hinchey II) was made.
The treatment strategy was determined according to the European Clinical Guidelines for Diverticular Disease [12]. Drainage of the abscess under ultrasound control and thus minimally invasive surgery was technically impossible due to the risk of bowel perforation since the intestine loops formed the walls of the abscess. The patient was offered surgical treatment, to which he gave consent. The preoperative examination revealed no contraindications. The preoperative preparation included bowel preparation, antibiotic prophylaxis, and prevention of venous thromboembolic complications.
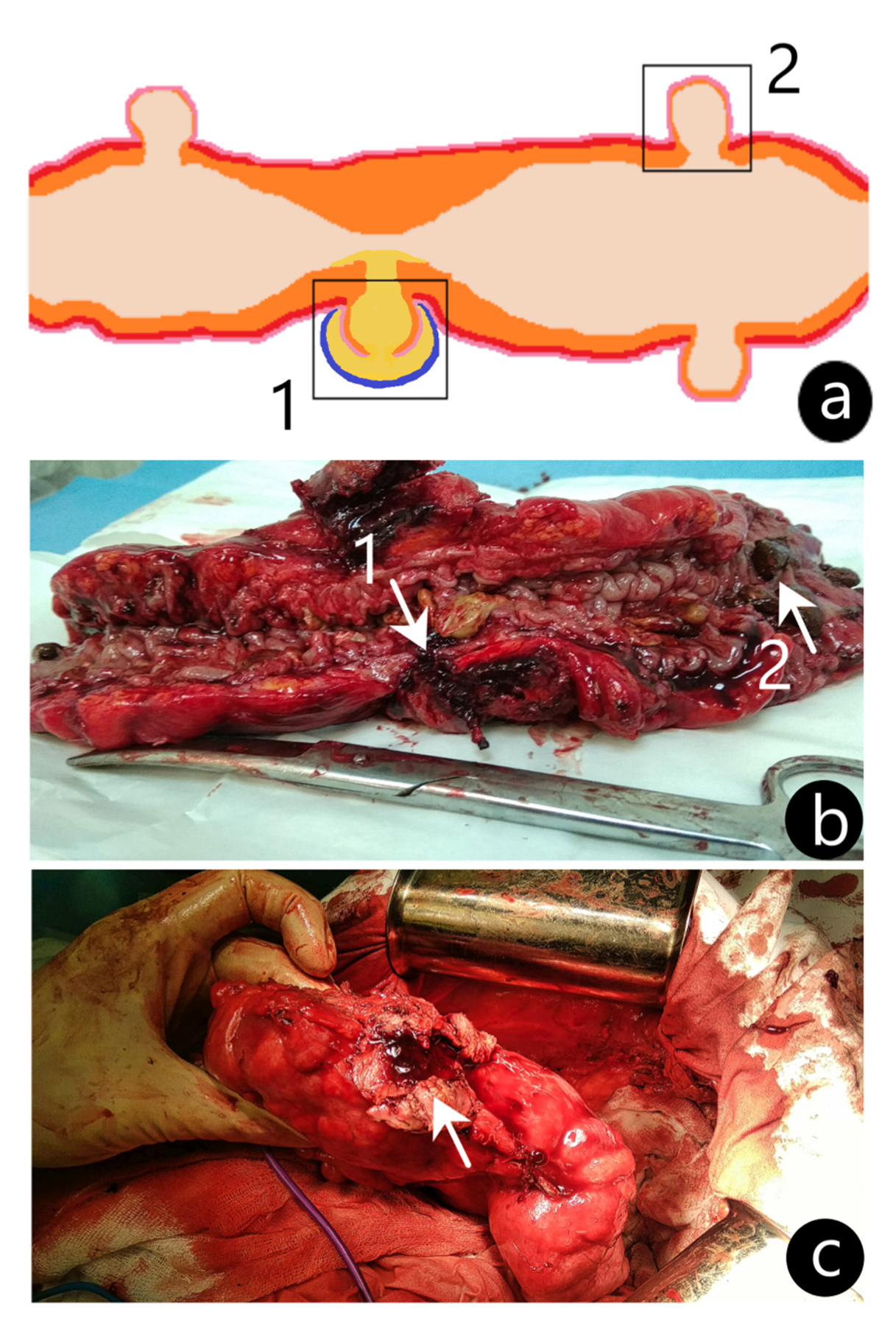
The operation was performed by the laparotomic approach. The refusal of laparoscopic intervention was due to the presence of a dense inflammatory conglomerate consisting of the abdominal wall, sigmoid colon, omentum, and loops of the small intestine with a large (45 × 35 × 56 mm) pericolic abscess in the center. During mobilization, the pericolic abscess was opened, and the contents were taken for a bacteriological examination, which further revealed microbial cell counts of Escherichia coli as high as 103 CFU/mL. A section of the sigmoid colon affected by the perforated diverticulum was resected; then, intestinal continuity was restored by an invaginating type of colorectal anastomosis. From the resected colon segment, two specimens were taken for the microbiota analysis: an inflamed and perforated diverticulum (ID) and a diverticulum without signs of inflammation (NID) (Figure 2). A routine histological examination of the specimens was performed in the Pathological Anatomy Department of the Republican Clinical Hospital (Kazan, Republic of Tatarstan). Both specimens were stained with hematoxylin and eosin and examined under the microscope (Zeiss Axio Scope A1, Goettingen, Germany). In the ID, a swollen intestinal wall with lymphocytic infiltration was revealed, while the NID showed no signs of acute inflammation.
We used a conventional culture method (Table 1) and 16S rRNA-based sequencing approach (Table 2) to assess the microbiota composition of both diverticula (for methods, see [13]). After trimming low-quality reads, the dereplicated reads were used for amplicon sequence variants (ASVs) inference. A total of 26,781 16S rRNA sequence reads was obtained following quality filtering, equating to 13,390 ± 3939 (mean ± SEM) reads per sample. After the removal of chimeras (1.75%) and non-bacterial sequences (9.38%), the number of mapped sequence reads per sample ranged from 8796 to 15,012 (mean ± SEM of 60,886 ± 4034). Following the removal of rare OTUs, defined as OTUs with 2 or less sequences across all the samples, rarefaction analysis demonstrated sufficient sequencing depth for a comparison analysis between the samples. The alpha-diversity analysis revealed an increased species richness and evenness in ID compared to NID (Table 2). These data were in opposition to the results of culturing, which showed nearly equal total bacterial growth (the culturable microbiota portion) in both diverticula (Table 1). The four phyla, Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria, constituted the main bulk of microbiota in both diverticula, but in ID, the relative abundance of the Firmicutes phylum was significantly increased, and the Proteobacteria decreased as compared to NID (Table 3). The Firmicutes-to-Bacteroidetes (F/B) ratio is widely used to describe the gut microbiota composition. A decreased or an increased F/B ratio is regarded as an indicator of dysbiosis [14]. In ID, we found a F/B ratio of 69.3%/16.6%, which is significantly higher than in NID (Firmicutes 43.2%/Bacteroidetes 17.3%). Although the total load of bacteria of the Bacteroidetes phylum was similar between the two diverticula, we observed notable differences in the content of the two most abundant genera assigned to this phylum. In ID, we detected an increased abundance of Bacteroides (8%), while the Prevotella content was significantly depleted (5.4%). Conversely, NID showed an increase in the abundance of the genus Prevotella (9.6%) and a decrease in the abundance of the genus Bacteroides (5.4%) (Table 3).
We revealed a significant difference in the population of Enterobacteriaceae between the two samples (relative abundance of 5.4% in ID vs. 29.3% in NID) (Table 3). In this family, the 16S rRNA-based sequencing approach is characterized by the limited resolution at the genus and species levels [15]. We attempted to analyze enterobacteria using culture methods but obtained contradictory data regarding their amount in the two samples (Table 1), which may originate from the prevalence of uncultivated microorganisms in NID that could not be detected by the culture method.
Surprisingly, ID was characterized by the preponderant presence of the genera Blautia, Coprococcus, Roseburia, and Faecalibacterium, which are the butyrate-producing bacteria with anti-inflammatory properties. NID showed an increase in the abundance of the genera Streptococcus and Ralstonia and a decrease in the abundance of Collinsella as compared with ID (Table 3).
The postoperative period proceeded without complications, and the patient was discharged on the sixth day after surgical treatment to a rehabilitation facility. Six months later, he was seen in the clinic and reported no further clinical symptoms of diverticulitis; laboratory tests and control sonography results revealed no signs of the disease.
3. Discussion
Several lines of evidence suggest a role for intestinal microbiota in the pathogenesis of DD. Substantial changes in the gut microbial community structure have been detected in the patients suffering from DD [4,5]. Probiotics are able to restore changes in microbiota composition and, therefore, are potentially applicable in the treatment of this disease [16]. The beneficial effects of rifaximin administration in SUDD also support the involvement of bacteria in the pathophysiological process [17]. In line with the possible etiological role of microbiota in diverticulitis, we assume that disease-causing infectious agents might be enriched at sites of active inflammation relative to comparatively unaffected mucosa. We have, therefore, used a conventional culture-based method and bacterial 16S rRNA gene sequencing technology to compare the mucosa-associated microbiota from ID and NID from the same colonic segment. It has repeatedly been shown that DD generally affects only limited segments of the colon [11], and thus diverticular pockets are more relevant samples than feces or mucosal biopsies.
The microbiota profiles differed substantially between ID and adjacent NID. Inflammation and perforation of the diverticulum were accompanied by an increased phylogenetic diversity and total amount of bacteria, an abundance of Firmicutes, a depletion of Proteobacteria, and an advanced F/B ratio in the gut microbiota as compared to NID. The most drastic difference between the two samples was in reference to the overabundance of uncultivated Enterobacteriaceae family in NID, as follows from the results of sequencing and culture methods. Different from our study, it has been well documented that intestinal inflammation is usually associated with a reduction in microbial richness and diversity, decreased F/B ratio, and increased abundance of potential proinflammatory Proteobacteria [18]. Fecal microbiota in diverticulosis and SUDD was reported to be deficient in Clostridium cluster IV, which is now classified as several genera in the family Ruminococcaceae, including the anti-inflammatory and butyrate-producing species Faecalibacterium prausnitzii [9]. In contrast, ID in our study preponderated over NID in the relative abundance of Ruminococcaceae and F. prausnitzii in 1.6 and 1.9 times, respectively. The other butyrate producers, such as Blautia, Coprococcus, Roseburia, and Faecalibacterium, were significantly increased in ID as compared to NID. Overall, the characteristics that are usually associated with the inflammatory process in the gut, namely reduced microbial diversity and richness, decreased F/B ratio, depletion of butyrate-producing bacteria, and Enterobacteriaceae blooming, were more pronounced in NID rather than in ID. Of these two studied samples, the microbiota of NID corresponded to the literature data on the fecal or mucosal microbiota of patients with SUDD [9] or acute diverticulitis [19].
We found an inverse relationship in enrichment between Prevotella and Bacteroides in both ID and NID. The Prevotella-to-Bacteroides ratios (P/B ratios) in the studied diverticula were 0.67 and 1.79, respectively. These two genera of the Bacteroidales order have previously been suggested as the main determinants of two of the three enterotypes. Enterotype 1 is characterized by a relatively high abundance of Bacteroides spp., enterotype 2 by Prevotella spp., and enterotype 3 by Ruminococcus [20]. The understanding of the biological significance of enterotypes is still limited. Enterotype 1 has previously been associated with the Western diet that has a high intake of animal protein and saturated fat, whereas enterotype 2 has been reported to be predominant in individuals that consume carbohydrate- and fiber-rich diets [21]. Nevertheless, individuals belonging to enterotype 2 had significantly higher plasma concentrations of cholesterol and trimethylamine-N-oxide (TMAO), a proatherogenic metabolite, than individuals belonging to enterotype 1 [22,23]. Our data suggest that inflammation in the diverticula is associated with a shift in the P/B ratio towards a predominance of Bacteroides.
The other taxa whose abundance differed significantly in ID and NID were Collinsella aerofaciens, Streptococcus luteciae, and Ralstonia spp. C. aerofaciens was increased in ID compared to NID (2.3 vs. 0.4%, respectively), while two other taxa were preponderant in NID compared to ID (5.5 vs. 1.4% and 4.3 vs. 1.0%, respectively). Although the association of these known intestinal commensals with some diseases has been reported [24,25,26], the mechanism of their probable pathophysiological activity remains obscure.
Thus, our results indicate several pronounced changes in the intraluminal mucosal microbiota of the colonic diverticulum injured by acute inflammation and perforation. This pilot study presents only a snapshot of the intestinal microbiota within the diverticula confined space of which provides persistence of normal and pathogenic microbiota. Observed changes in the microbiota may reflect location-associated differences and warrant further microbiota studies throughout the length of the colon. Another important limitation of this study is that gut microbiota varies greatly between individuals, and the results obtained for one patient may not be widely generalizable. Further studies should help to elucidate the role of microbiota in the pathogenesis of DD and its progression towards more severe forms. The prediction of diverticulitis risks based on microbiota biomarkers opens the perspective of prophylaxis of disease relapses, more timely treatment, and prevention of complications, including the deliberate use of antibiotics for diverticulitis treatment and application of probiotics and fecal microbiota transplantation (FMT) for specific modulation of intestinal microbiota.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/life12122129/s1, Table S1: Taxonomic profiling of the diverticula by 16S ribosomal RNA gene sequencing.
Author Contributions
Conceptualization: D.Y. and O.K. (Oleg Karpukhin); Methodology: D.Y., T.G., and O.K. (Oleg Karpukhin); Formal analysis and investigation: Y.P. and O.K. (Olga Karaseva); Writing—original draft preparation: D.Y. and O.K. (Oleg Karpukhin); Writing—review and editing: D.Y., T.G. and O.K. (Oleg Karpukhin); Funding acquisition: D.Y.; Supervision: O.K. (Oleg Karpukhin). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation (grant N 22-16-00040). The work was performed in frames of the Kazan Federal University Strategic Academic Leadership Program (PRIORITY-2030).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Local Ethics Committee of Kazan State Medical University (Protocol No. 4, date 26 April 2022).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patient to publish this paper.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Karpukhin, O.Y.; Pankratova, Y.S.; Shakirov, R.R.; Cherkashina, M.I. The influence of the COVID-19 pandemic on the diverticular disease complications treatment. Bull. Contemp. Clin. Med. 2022, 15, 33–39. [Google Scholar] [CrossRef]
- Tursi, A.; Scarpignato, C.; Strate, L.L.; Lanas, A.; Kruis, W.; Lahat, A.; Danese, S. Colonic diverticular disease. Nat. Rev. Dis. Prim. 2020, 6, 1–23. [Google Scholar] [CrossRef]
- Wedel, T.; Barrenschee, M.; Lange, C.; Cossais, F.; Böttner, M. Morphologic Basis for Developing Diverticular Disease, Diverticulitis, and Diverticular Bleeding. Visc. Med. 2015, 31, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Ticinesi, A.; Nouvenne, A.; Corrente, V.; Tana, C.; Di Mario, F.; Meschi, T. Diverticular Disease: A Gut Microbiota Perspective. J. Gastrointest. Liver Dis. 2019, 28, 327–337. [Google Scholar] [CrossRef]
- Reitano, E.; Francone, E.; Bona, E.; Follenzi, A.; Gentilli, S. Gut Microbiota Association with Diverticular Disease Pathogenesis and Progression: A Systematic Review. Am. J. Dig. Dis. 2022, 1–9. [Google Scholar] [CrossRef]
- van Rossen, T.M.; Ooijevaar, R.E.; Kuyvenhoven, J.P.; Eck, A.; Bril, H.; Buijsman, R.; Boermeester, M.A.; Stockmann, H.B.A.C.; de Korte, N.; Budding, A.E. Microbiota composition and mucosal immunity in patients with asymptomatic diverticulosis and controls. PLoS ONE 2021, 16, e0256657. [Google Scholar] [CrossRef]
- Jones, R.B.; Fodor, A.A.; Peery, A.F.; Tsilimigras, M.C.B.; Winglee, K.; McCoy, A.; Sioda, M.; Sandler, R.S.; Keku, T.O. An Aberrant Microbiota is not Strongly Associated with Incidental Colonic Diverticulosis. Sci. Rep. 2018, 8, 4951. [Google Scholar] [CrossRef] [Green Version]
- Tursi, A.; Mastromarino, P.; Capobianco, D.; Elisei, W.; Miccheli, A.; Capuani, G.; Tomassini, A.; Campagna, G.; Picchio, M.; Giorgetti, G.; et al. Assessment of Fecal Microbiota and Fecal Metabolome in Symptomatic Uncomplicated Diverticular Disease of the Colon. J. Clin. Gastroenterol. 2016, 50 (Suppl. 1), S9–S12. [Google Scholar] [CrossRef] [Green Version]
- Barbara, G.; Scaioli, E.; Barbaro, M.R.; Biagi, E.; Laghi, L.; Cremon, C.; Marasco, G.; Colecchia, A.; Picone, G.; Salfi, N.; et al. Gut microbiota, metabolome and immune signatures in patients with uncomplicated diverticular disease. Gut 2016, 66, 1252–1261. [Google Scholar] [CrossRef]
- Schieffer, K.M.; Sabey, K.; Wright, J.R.; Toole, D.R.; Drucker, R.; Tokarev, V.; Harris, L.R.; Deiling, S.; Eshelman, M.A.; Hegarty, J.P.; et al. The Microbial Ecosystem Distinguishes Chronically Diseased Tissue from Adjacent Tissue in the Sigmoid Colon of Chronic, Recurrent Diverticulitis Patients. Sci. Rep. 2017, 7, 8467. [Google Scholar] [CrossRef]
- Spiller, R.C.; Sloan, T.J. Do diverticula provide a unique niche for microbiota which can lead to activation of the innate immune system? Gut 2016, 66, 1175–1176. [Google Scholar] [CrossRef]
- Schultz, J.K.; Azhar, N.; Binda, G.A.; Barbara, G.; Biondo, S.; Boermeester, M.A.; Chabok, A.; Consten, E.C.J.; Van Dijk, S.T.; Johanssen, A.; et al. European Society of Coloproctology: Guidelines for the management of diverticular disease of the colon. Color. Dis. 2020, 22, 5–28. [Google Scholar] [CrossRef]
- Yarullina, D.R.; Shafigullin, M.U.; Sakulin, K.A.; Arzamastseva, A.A.; Shaidullov, I.F.; Markelova, M.I.; Grigoryeva, T.V.; Karpukhin, O.Y.; Sitdikova, G.F. Characterization of gut contractility and microbiota in patients with severe chronic constipation. PLoS ONE 2020, 15, e0235985. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- McLean, K.; Rosenthal, C.A.; Sengupta, D.; Owens, J.; Cookson, B.T.; Hoffman, N.G.; Salipante, S.J. Improved Species-Level Clinical Identification of Enterobacteriaceae through Broad-Range dnaJ PCR and Sequencing. J. Clin. Microbiol. 2019, 57, e00986-19. [Google Scholar] [CrossRef]
- Piccioni, A.; Franza, L.; Brigida, M.; Zanza, C.; Torelli, E.; Petrucci, M.; Nicolò, R.; Covino, M.; Candelli, M.; Saviano, A.; et al. Gut Microbiota and Acute Diverticulitis: Role of Probiotics in Management of this Delicate Pathophysiological Balance. J. Pers. Med. 2021, 11, 298. [Google Scholar] [CrossRef]
- Moniuszko, A.; Rydzewska, G. The effect of cyclic rifaximin therapy on symptoms of diverticular disease from the perspective of the gastroenterology outpatient clinic: A “real-life” study. Gastroenterol. Rev. 2017, 12, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Caruso, R.; Lo, B.C.; Núñez, G. Host–microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef]
- MJ, O.G.; Turner, G.A.; Frizelle, F.A. Distinct changes in the colonic microbiome associated with acute diverticulitis. Color. Dis. 2022, 1–11. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roager, H.M.; Licht, T.R.; Poulsen, S.K.; Larsen, T.M.; Bahl, M.I. Microbial Enterotypes, Inferred by the Prevotella-to-Bacteroides Ratio, Remained Stable during a 6-Month Randomized Controlled Diet Intervention with the New Nordic Diet. Appl. Environ. Microbiol. 2014, 80, 1142–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.P.; Adley, C.C. Ralstonia spp.: Emerging global opportunistic pathogens. Eur. J. Clin. Microbiol. 2013, 33, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, M.; Chow, C.-E.T.; Ryan, C.N.; Chan, L.S.; Dufour, J.; Aye, P.P.; Blanchard, J.; Moehs, C.P.; Sestak, K. Dietary Gluten-Induced Gut Dysbiosis Is Accompanied by Selective Upregulation of microRNAs with Intestinal Tight Junction and Bacteria-Binding Motifs in Rhesus Macaque Model of Celiac Disease. Nutrients 2016, 8, 684. [Google Scholar] [CrossRef]
Figure 1.
Coronal (a) and axial (b) abdominal CT scans revealed diverticulitis (arrow 1) of the sigmoid colon complicated by the formation of a pelvic abscess (arrow 2).
Figure 1.
Coronal (a) and axial (b) abdominal CT scans revealed diverticulitis (arrow 1) of the sigmoid colon complicated by the formation of a pelvic abscess (arrow 2).

Figure 2.
Visualization of the colonic specimens with diverticula. (a) An illustrated representation of the sigmoid colon with an inflamed perforated diverticulum (ID) (1) and non-inflamed diverticulum (NID) (2) obtained for analysis shown in boxes. (b) Postoperative image of the sigmoid colon resection with ID (arrow 1) and NID (arrow 2). (c) Intraoperative image of the sigmoid colon with capsule fragment of diverticular pericolic abscess (arrow).
Figure 2.
Visualization of the colonic specimens with diverticula. (a) An illustrated representation of the sigmoid colon with an inflamed perforated diverticulum (ID) (1) and non-inflamed diverticulum (NID) (2) obtained for analysis shown in boxes. (b) Postoperative image of the sigmoid colon resection with ID (arrow 1) and NID (arrow 2). (c) Intraoperative image of the sigmoid colon with capsule fragment of diverticular pericolic abscess (arrow).


{kind=link}
{kind=link}
Table 1.
Microbiota of diverticula assessed by culture method.
| Groups of Microorganisms, lg CFU/g | Inflamed Diverticulum (ID) | Non-Inflamed Diverticulum (NID) |
|---|---|---|
| Total bacterial growth of aerobic bacteria | 13.45 | 13.37 |
| Total bacterial growth of anaerobic bacteria | 11.90 | 12.18 |
| Lactobacilli | 12.15 | 12.29 |
| Bifidobacterium spp. | 11.99 | nd |
| Enterobacteriaceae | 12.27 | 7.81 |
| Salmonella spp., Shigella spp. | nd | nd |
nd—not determined.
Table 2.
Number of reads and alpha-diversity indices of the diverticula.
| Inflamed Diverticulum (ID) | Non-Inflamed Diverticulum (NID) | |
|---|---|---|
| Raw reads | 16,176 | 10,605 |
| Non-chimeric reads | 15,765 | 10,587 |
| Reads with OTU | 15,012 | 8796 |
| Alpha-diversity indices | ||
| Observed OTUs | 665 | 458 |
| Phylogenetic diversity | 41.0 | 32.6 |
| Chao1 | 670.4 | 496.6 |
| Shannon | 7.3 | 6.1 |
| Simpson | 0.98 | 0.92 |
Table 3.
Major taxa detected in diverticula by 16S ribosomal RNA gene sequencing *.
| Taxonomy | Relative Abundance, % | |
|---|---|---|
| Inflamed Diverticulum (ID) | Non-Inflamed Diverticulum (NID) | |
| Actinobacteria | 3.6 | 3.2 |
| Actinomycetales | 0.3 | 2.0 |
| Coriobacteriales | 3.3 | 0.6 |
| Coriobacteriaceae | 3.3 | 0.6 |
| Collinsella | 2.3 | 0.4 |
| Bacteroidetes | 16.6 | 17.3 |
| Bacteroidales | 16.6 | 17.3 |
| Bacteroidaceae | 8.0 | 5.4 |
| Bacteroides | 8.0 | 5.4 |
| Prevotellaceae | 5.4 | 9.6 |
| Prevotella | 5.4 | 9.6 |
| Firmicutes | 69.3 | 43.2 |
| Lactobacillales | 2.5 | 5.8 |
| Streptococcaceae | 1.6 | 5.5 |
| Streptococcus | 1.4 | 5.5 |
| Clostridiales | 66.3 | 36.2 |
| Lachnospiraceae | 29.0 | 14.8 |
| Blautia | 8.7 | 4.4 |
| Coprococcus | 4.7 | 1.4 |
| Roseburia | 4.7 | 1.4 |
| Ruminococcaceae | 23.2 | 14.3 |
| Faecalibacterium | 9.2 | 4.9 |
| Ruminococcus | 3.0 | 2.1 |
| Unknown Ruminococcaceae | 9.9 | 7.0 |
| Proteobacteria | 9.5 | 36.0 |
| Burkholderiales | 1.3 | 6.3 |
| Oxalobacteraceae | 1.0 | 4.3 |
| Ralstonia | 1.0 | 4.3 |
| Enterobacteriales | 5.4 | 29.3 |
| Enterobacteriaceae | 5.4 | 29.3 |
| Unknown Enterobacteriaceae | 5.4 | 29.3 |
| Pseudomonadales | 1.4 | 0.0 |
| Other phyla | 1 | 0.3 |
* See Table S1 for the complete list of taxa detected in the diverticula by 16S rRNA gene sequencing.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yarullina, D.; Pankratova, Y.; Karaseva, O.; Grigoryeva, T.; Karpukhin, O. Microbiota of the Colonic Diverticula in the Complicated Form of Diverticulitis: A Case Report. Life 2022, 12, 2129. https://doi.org/10.3390/life12122129
AMA Style
Yarullina D, Pankratova Y, Karaseva O, Grigoryeva T, Karpukhin O. Microbiota of the Colonic Diverticula in the Complicated Form of Diverticulitis: A Case Report. Life. 2022; 12(12):2129. https://doi.org/10.3390/life12122129
Chicago/Turabian StyleYarullina, Dina, Yuliia Pankratova, Olga Karaseva, Tatiana Grigoryeva, and Oleg Karpukhin. 2022. "Microbiota of the Colonic Diverticula in the Complicated Form of Diverticulitis: A Case Report" Life 12, no. 12: 2129. https://doi.org/10.3390/life12122129
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.