
Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides
by
 , , and
, , and
Elham Amin
1,2,
Mohamed Sadek Abdel-Bakky
3,4,
Hamdoon A. Mohammed
1,5 and
and
Marwa H. A. Hassan
2,* 1
Department of Medicinal Chemistry and Pharmacognosy, College of Pharmacy, Qassim University, Buraydah 51452, Saudi Arabia
2
Department of Pharmacognosy, Faculty of Pharmacy, Beni-Suef University, Beni-Suef 62514, Egypt
3
Department of Pharmacology and Toxicology, College of Pharmacy, Qassim University, Buraydah 51452, Saudi Arabia
4
Department of Pharmacology and Toxicology, Faculty of Pharmacy, Al-Azhar University, Cairo 11751, Egypt
5
Department of Pharmacognosy and Medicinal Plants, Faculty of Pharmacy, Al-Azhar University, Cairo 11751, Egypt
*
Author to whom correspondence should be addressed.
Life 2022, 12(11), 1852; https://doi.org/10.3390/life12111852
Submission received: 18 October 2022
/
Revised: 2 November 2022
/
Accepted: 7 November 2022
/
Published: 11 November 2022
(This article belongs to the Special Issue Plants as a Promising Biofactory for Bioactive Compounds)
Abstract
:Natural products continue to provide inspiring chemical moieties that represent a key stone in the drug discovery process. As per our previous research, the halophyte Agathophora alopecuroides was noted as a potential antidiabetic plant. However, the chemical profiling and highlighting the metabolite(s) responsible for the observed antidiabetic activity still need to be investigated. Accordingly, the present study presents the chemical profiling of this species using the LC-HRMS/MS technique followed by a study of the ligand–protein interaction using the molecular docking method. LC-HRMS/MS results detected twenty-seven compounds in A. alopecuroides extract (AAE) belonging to variable chemical classes. Among the detected compounds, alkaloids, flavonoids, lignans, and iridoids were the most prevailing. In order to highlight the bioactive compounds in AAE, the molecular docking technique was adopted. Results suggested that the two alkaloids (Eburnamonine and Isochondrodendrine) as well as the four flavonoids (Narirutin, Pelargonidin 3-O-rutinoside, Sophora isoflavanone A, and Dracorubin) were responsible for the observed antidiabetic activity. It is worth mentioning that this is the first report for the metabolomic profiling of A. alopecuroides as well as the antidiabetic potential of Isochondrodendrine, Sophora isoflavanone A, and Dracorubin that could be a promising target for an antidiabetic drug.
1. Introduction
Diabetes mellitus is a chronic metabolic disease recognized by an increase in blood glucose levels which develops from a deficiency in insulin secretion, action, or both of them [1]. Type 2 diabetes mellitus (T2DM) is the most common health problem and accounts for about 90% of diabetes cases with 4.9 million mortalities throughout the world [2]. Inhibiting the digestion of dietary carbohydrates is one of the effective procedures for the management of postprandial hyperglycemia in T2DM. One of the essential digestive enzymes is pancreatic α-amylase which converts dietary carbohydrates such as starch into smaller oligosaccharides mixture that are further broken down into glucose by α-glucosidase, another important metabolic enzyme. Upon the absorption of glucose, it enters the bloodstream and causes postprandial elevation in blood glucose levels. Therefore, blocking the enzymes α-amylase and α-glucosidase can inhibit the digestion of carbohydrates, postpone glucose uptake, and subsequently lower blood sugar levels [3]. Recently, medicinal plants proved their great therapeutic potential and negligible side effects in the treatment of T2DM. For instance, several medicinal herbs have been reported to exhibit strong glucosidase and amylase inhibitory properties [4,5,6].
Halophytes are salt-tolerating plants noted for their ability to produce variable secondary metabolites, such as alkaloids, glycosides, and terpenes. Hence, they could be considered as promising sources for bioactive metabolites that could be used for the treatment of various diseases such as diabetes [7,8,9]. A. alopecuroides is a halophytic species prevalent in the deserts of Saudi Arabia and was reported to exhibit a strong in vitro and in vivo antidiabetic activity [7]; nevertheless, its chemical profile remained to be investigated [10].
Dereplication, defined as the rapid identification of known compounds from natural product extracts, represents an important step in drug discovery programs. This approach combines the benefits of different analytical techniques, modern spectroscopic methods, as well database searches for the prompt characterization of an active compound during the drug discovery process. Recently, advances in technology have provided what is called tandem analytical techniques, such as LC-MS, LC-MS/MS, LC-NMR, HPLC-PDA, and LC-NMR/MS [11]. Tandem mass spectrometry (MS/MS) is a powerful technique for the characterization of target phytoconstituents in complex plant extracts. The high sensitivity, selectivity, and fast screening abilities of the LC-MS/MS technique, compared to other dereplication techniques, rationalized the privilege of this technique for the online identification of secondary metabolites in plant extracts [11].
Furthermore, advances in computational biology have had a great impact on reducing the time, cost, and effort spent while screening the biological activity of natural products. Molecular docking is now widely adopted for predicting the binding mode and binding affinity of a drug-like molecule into the active site of the receptor. A huge number of natural and synthetic compounds could be virtually screened for activity against a wide array of targets, thus reducing the time and effort and giving a rapid expectation for the most promising candidates [12].
Based on this concept and in continuation of our previous research on the halophytes with promising antidiabetic potential [7], which recorded the strong in vitro and in vivo antidiabetic potential of AAE, the current study investigated the metabolic content of AAE using LC-MS/MS followed by the screening of the annonated compounds for the enzyme inhibitory potential against the two carbohydrate-metabolizing enzymes—α-amylase and α-glucosidase—using the molecular docking technique.
2. Materials and Methods
2.1. Plant Materials
Agathophora alopecuroides var. papillosa was collected from the Qassim region in the northcentral Saudi Arabia during October 2021. The taxonomic identity of the plant was confirmed by Ibrahim Aldakhil, botanical expert, Qassim Area, and avoucher sample number QPP-101 was kept at the College of Pharmacy, Qassim University (Buraydah, Qassim, Saudi Arabia). The aerial parts of the plant were carefully cleaned then dried in the shade for two weeks. The dried powdered plant material (500 g) was extracted by maceration in 80% methanol (3 times × 1000 mL) at room temperature with frequent shaking. The methanolic extract was then concentrated by a vacuum rotary evaporator. The dried extract was then kept in an amber-colored vial at 4 °C till further use.
2.2. Metabolites Profiling of the Methanolic Extract of A. alopecuroides
AAE was reconstituted in HPLC grade methanol, filtered through a 0.22 µm PTFE membrane, and then separation was performed adopting Thermo Scientific C18 column (AcclaimTM Polar Advantage II, 3 × 150 mm, 3 µm particle size) on an UltiMate 3000 UHPLC system (Dionex). Gradient elution was performed at a flow rate of 0.4 mL/min and a column temperature of 40 °C, using H2O + 0.1% Formic Acid (A) and 100% Acetonitrile (B) with a 22 min total run time. The injection volume of the sample was 3 µL. The gradient started at 5% B (0–3 min), 80% B (3–10 min), 80% B (10–15 min), and 5% B (15–22 min). High-resolution mass spectrometry was carried out using a MicroTOF QIII (Bruker Daltonic, Bremen, Germany) using an ESI positive ionization and adjusting the following settings: capillary voltage: 4500 V; nebulizer pressure: 2.0 bar; drying gas: 8 L/min at 300 °C. The mass range was 50–1000 m/z. The accurate mass data of the molecular ions, provided by the TOF analyzer, were processed by Compass Data Analysis software (Bruker Daltonik GmbH).
2.3. Molecular Docking Study
AutoDock Vina software was used in all molecular docking experiments [13]. All dereplicated compounds were docked against the active sites of both human α-amylase and human α-glucosidase crystal structure (PDB codes: 4W93 and 3L4W, respectively) [14,15]. The binding site was determined according to the enzyme’s co-crystallized ligands (Montbretin A and Miglitol, respectively). The co-ordinates of the grid boxes were (x = −9.682; y = 4.274; z = −23.145 and x = 45.424; y = 92.375; z = 34.811). The size of the grid box was set to 20 Å. Exhaustiveness was set to 24. Ten poses were generated for each docking experiment. Docking poses were analyzed and visualized using Pymol software [13]. The full method is provided in the Supplementary Materials.
3. Results
3.1. In Vitro Testing of the Antidibetic Activity of A. alopecuroides (AAE)
This research represents an extension of our previous work on halophytic plants with potential antidiabetic activity. In the previous publication, in vitro testing revealed the strong inhibitory activity of the hydroalcoholic extract (AAE) against α-glucosidase and α-amylase with IC50 values 117.9 and 90.9 µg/mL, respectively, compared to 191.4 and 53.3 µg/mL of the standard drug Acarbose [7].
3.2. Metabolomic Profiling of the Methanolic Extract of A. alopecuroides (AAE)
The metabolic profiling using LC-HRMS/MS of AAE led to the annonation of 27 metabolites with variable chemical structures. The annonated metabolites could be classified according to their chemical classes into: nine alkaloids, five flavonoids, four lignans, two iridoid glycosides, two acids, one anthraquinone, one benzisochroman, one furanochomarine, one triterpenoid saponin, and one benzofuran (Table 1, Figure 1 and Figure 2). Using the metabolomic data, the nine alkaloids were annotated from the mass ion peaks at m/z 275.152, 294.173, 361.153, 285.290, 328.155, 313.131, 301.131, 594.273, and 356.319, which were in agreement with the molecular formulas C16H21NO3, C19H22N2O, C19H23NO6, C16H15NO4, C19H22NO4, C18H19NO4, C17H19NO4, C36H38N2O6, and C24H40N2, respectively. These alkaloids were dereplicated as Epinorlycoramine (1), Eburnamonine (2), 3-Acetylnerbowdine (3), Arborinine (4), 1,2-Dehydroreticuline (5), N-Feruloyltyramine (16), Powelline (19), Isochondrodendrine (24), and Conessine (26), respectively. While the five flavonoids were dereplicated from the mass ion peaks at m/z 580.179, 579.171, 446.121, 370.142, and 488.162 that matched with the molecular formulas C27H32O14, C27H31O14, C22H22O10, C21H22O6, and C32H24O5, respectively. These metabolites were characterized as Narirutin (9), Pelargonidin 3-O-rutinoside (10), Biochanin A-β-D-glucoside (12), Sophora isoflavanone A (14), and Dracorubin (23), respectively. The four lignans were dereplicated from the molecular ion peaks at m/z 372.194, 520.194, 312.121, and 342.131 corresponding to the suggested formulas C22H28O5, C26H32O11, C15H20O7, and C16H22O8, respectively, as Veraguensin (8), Pinoresinol glucoside (12), 4-Hydroxycinnamyl alcohol 4-D-glucoside (17), and Coniferin (18), respectively. The mass ion peaks at m/z 344.147 and 388.137 were in agreement with the molecular formulas C16H24O8 and C17H24O10 that were characterized as the two iridoid glycosides Boschnaloside (11) and Geniposide (15). Additionally, seven mass ion peaks at m/z 402.095, 288.136, 300.136, 314.152, 628.304, 278.225, and 634.408, compatible with the molecular formulae C20H18O9, C17H20O4, C18H20O4, C19H22O4, C38H44O8, C18H30O2, and C36H58O9, respectively, were dereplicated as Versiconol acetate (6), Karwinaphthol B (7), Toxyl angelate (20), Heliettin (21), Gambogic acid (22) Punicic acid (25), and Soyasapogenol B 3-O-d-glucuronide (27), respectively.
3.3. Molecular Docking Analysis
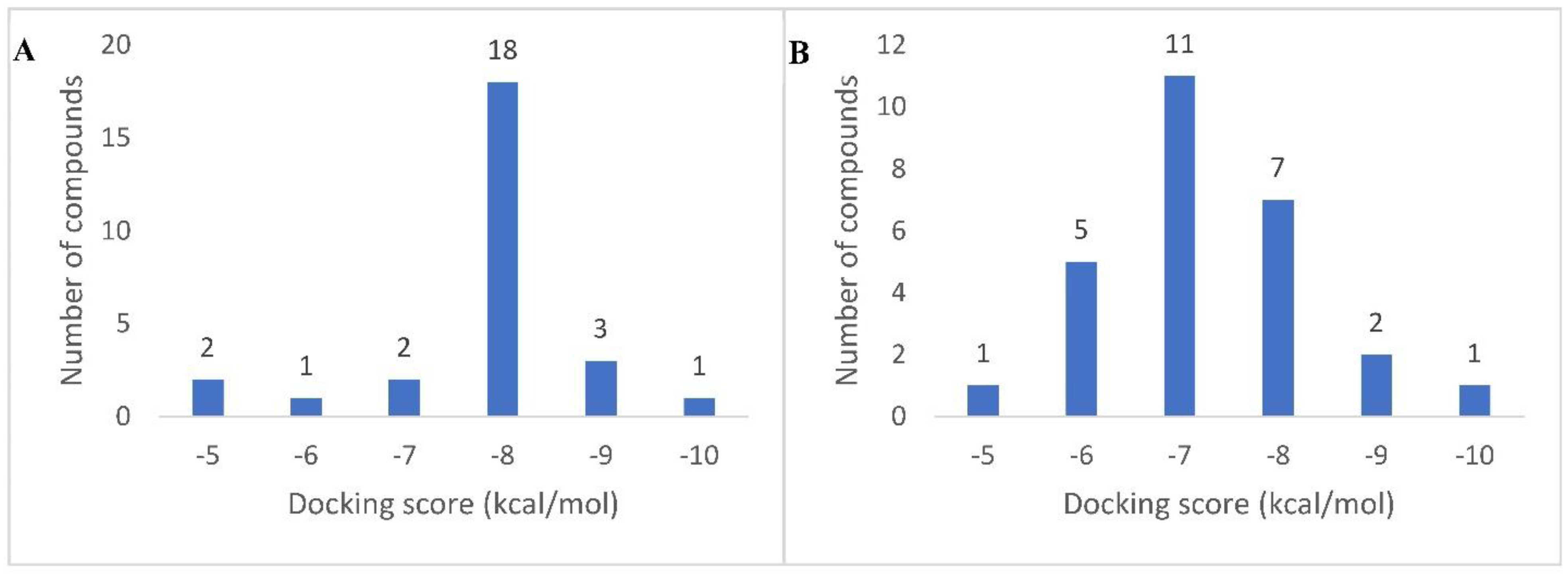
All the dereplicated compounds were subjected to molecular docking against both human α-amylase and human α-glucosidase enzymes (PDB codes: 4W93 and 3L4W, respectively). As shown in (Table 2 and Figure 3) and (Supplementary Materials, Figures S1 and S2), all dereplicated compounds received scores ranging from ~−4.5 to −9.1 kcal/mol against both enzymes. Most of the compounds (21 compounds in the case of α-amylase, and 23 compounds in the case of α-glucosidase) received scores ranging between −5 and −8 kcal/mol. The compounds that achieved the best score (<−8 kcal/mol) against α-amylase enzyme were Eburnamonine (2), Narirutin (9), Pelargonidin 3-O-rutinoside (10), and Isochondrodendrine (24). Considering α-glucosidase enzyme inhibition, the best scoring compounds (<−8 kcal/mol) were Pelargonidin 3-O-rutinoside (10), Sophora isoflavanone A (14), and Dracorubin (23). The two co-crystallized compounds—Montbretin A with amylase and Miglitol with glucosidase—achieved binding scores of −8.1 and −8.0 kcal/mol, respectively, and hence −8.0 kcal/mol was chosen as a cut-off value to select the best scoring compounds with each enzyme.
4. Discussion
This study is a continuation of our previous research dealing with the potential antidiabetic agents from halophytes. Previously, AAE was acknowledged for potent antidiabetic activity through both in vitro and in vivo investigations. The current study adds more information about the metabolic profiling of AAE and highlights the most promising metabolites expected to be responsible for the recorded antidiabetic activity.
- Metabolomic profiling of the methanolic extract of A. alopecuroides
For tentative identification of the components of AAE, we adopted a dereplication strategy using the hyphenated technique: liquid chromatography coupled with tandem mass spectrometry (LC-HRMS/MS) (Figure 1 and Figure 2). The dereplication process led to the recognition of 27 compounds for the first time from AAE. The detected metabolites belong to different chemical classes: alkaloids, flavonoids, lignans, iridoid glycosides, anthraquinones, benzisochromans, furanochomarine, triterpenoid saponins, acids, and benzofurans (Table 1, Figure 2). Amid the detected metabolites, alkaloids represent the most prevailing chemical class. These alkaloids belong to different types, and among them the most abundant is the isoquinoline-type alkaloids (Figure 1). Epinorlycoramine (1) is a galanthamine-type alkaloid previously isolated from Narcissus leonensis whole plant [16], while Eburnamonine (2), an eburnan-type alkaloid, was isolated from Kopsia larutensis leaves [17]. From the crinine-type alkaloids, two alkaloids were detected: 3-Acetylnerbowdine (3), which was reported in Nerine bowdenii bulbs using GC-MS analysis [18,19], and Powelline (19), reported in Crinum latifolium leaves using GC-MS analysis [40]. Arborinine (4), a nitrogen-containing drug similar to anthracene classified as acridone alkaloid, was previously isolated from the ethyl acetate extract of the Glycosmis parva plant [20]. The quaternary isoquinoline alkaloid 1,2-Dehydroreticuline (5), previously isolated from Xylopia parviflora root and bark [21,22], was also identified. N-Feruloyltyramine (16) is a phenolic amide alkaloid previously detected in Lycium barbarum fruits and A. alopecuroides [10,35]. Isochondrodendrine (24) is bis benzyli-soquinolinic alkaloids previously reported in the Cissampelos pareira plant [46,51]. Finally, Conessine (26), a pentacyclic steroidal alkaloid, has been isolated from Holarrhena floribunda G. Don. [48].
Another biologically important and widely prevailing chemical class is the flavonoids that were represented by five compounds from different flavonoid subclasses. Narirutin (9) is a flavanone common in the citrus family and reported for potent antidiabetic activity using in vitro and docking studies [27]. Pelargonidin 3-O-rutinoside (10), an anthocyanin with potent antidiabetic activity depicted through the inhibition of α-glucosidase and α-amylase enzymes, was isolated from strawberries [28]. Biochanin A-β-d-glucoside (12) is an isoflavone previously isolated from Trifolium pratense L. [31], while Sophora isoflavanone A (14) is a pterocarpan previously identified from Sophora tomentosa [33]. Finally, Dracorubin (23) was recognized as the major red coloring matter in the tree Dracaena draco resin [45].
LC-HRMS/MS results also characterized four lignans. Veraguensin (8) is a lignan compound previously identified in N. turbacensis (Kunth) Nees leaves and root bark [25,26]. Pinoresinol glucoside (12) was isolated from Prunus domestica [32] and was stated to exhibit potent antioxidant activity and powerful in vitro antihyperglycemic and hepatoprotective effects. 4-Hydroxycinnamyl alcohol 4-d-glucoside “4-O-β-d-glucopyranosyl-p-coumaric acid” (17) is a phenolic acid derivative that was isolated and identified in the flaxseed phenolic rich fraction [36,37]. Coniferin (18), is a phenolic glycoside previously isolated from the bark of Paulownia tomentosa [38,39].
Furthermore, the two iridoid glycosides Boschnaloside (11), previously reported from Euphrasia pectinata aerial parts [29,30], and Geniposide (15), isolated from Gardenia jasminoides Ellis fruit [34], were detected in AAE.
An additional seven compounds from variable secondary metabolites classes were also detected in AAE and dereplicated as: the anthraquinone compound Versiconol acetate (6), previously recognized in the cultures of Aspergillus parasiticus after using the insecticide dichlorvos [52,53]; the dimethyl benzisochroman compound Karwinaphthol B (7), isolated from Karwinskia humboldtiana roots [24]; the natural benzofuran Toxyl angelate (20), previously isolated from Isocoma wrightii plant [41]; the furanocoumarine compound Heliettin (21), previously isolated from the stem bark of Helietta longifolia Britt and Helietta apiculata [42,43]; Gambogic acid (22), previously isolated from Garcinia hanburyi plant [44]; the unsaturated fatty acid Punicic acid (25), previously isolated from pomegranate seed oil [47]; and finally, the triterpenoid saponin Soyasapogenol B 3-O-d-glucuronide (27), previously isolated from aerial parts of Lathylus palustris L. [49,50].
- Molecular docking study of A. alopecuroides metabolites for inhibition of α-amylase and α-glucosidase enzymes
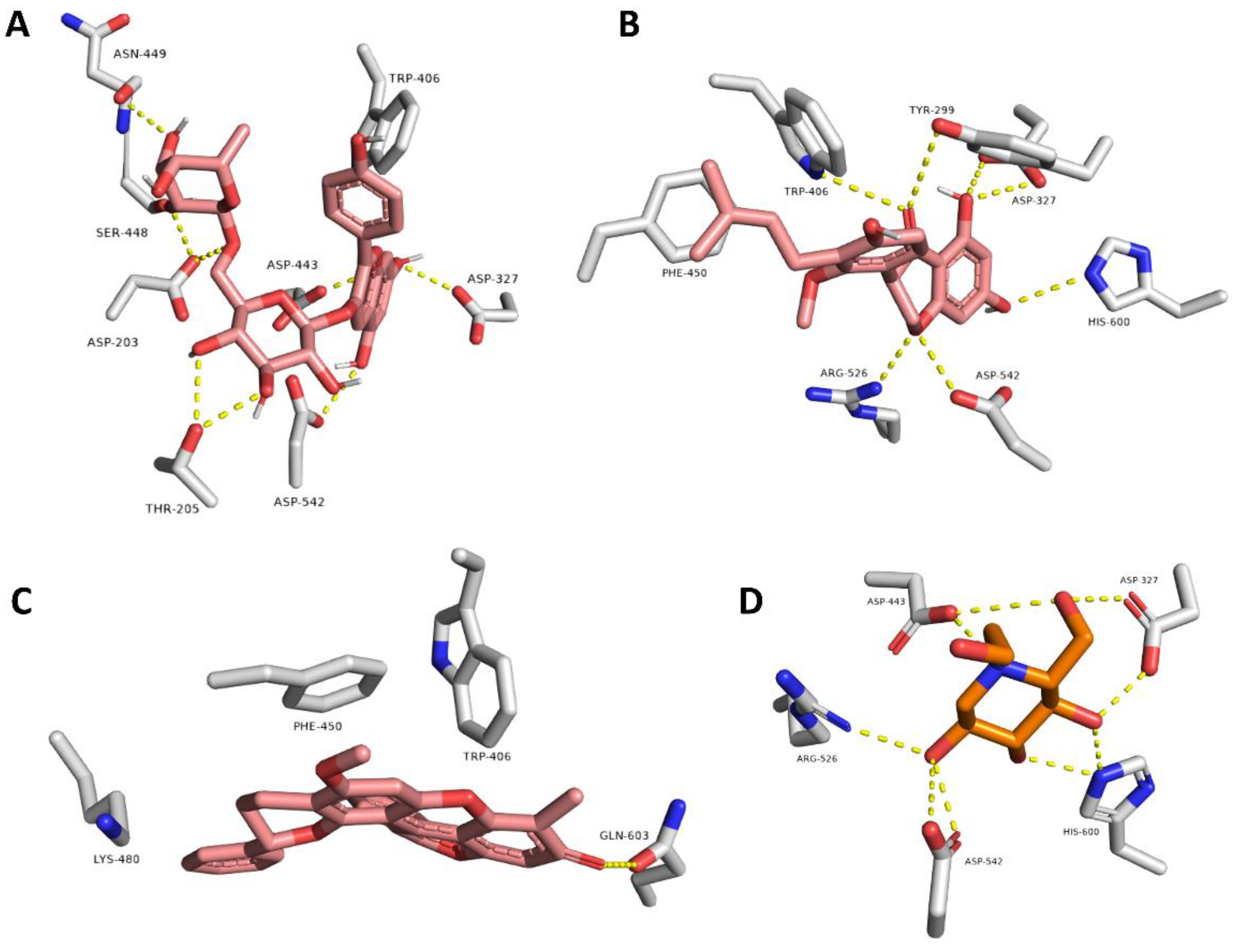
In order to highlight the probably bioactive metabolites in AAE, all the dereplicated compounds were subjected to molecular docking study against both human α-amylase and α-glucosidase enzymes. All the dereplicated compounds displayed binding energies within the range of −4.5 to −9.1 Kcal/mol with the two enzymes (Table 2). The two alkaloids Eburnamonine (2) and Isochondrodendrine (24) as well as the two flavonoids Narirutin (9) and Pelargonidin 3-O-rutinoside (10) achieved the best binding scores with α-amylase enzyme. These compounds showed various binding modes inside the enzyme active site. Narirutin and Pelargonidin 3-O-rutinoside binding poses were comparable with that of the co-crystalized inhibitor Montbretin A (Figure 4B,C,E), where they established multiple H-bonds with TYR-151, ASP-197, HIS-201, GLU-233, HiS-299, and ASP-300. In addition, Narirutin established further hydrophobic interactions with TRP-58 and TRP-59. Narirutin was previously reported to have a potent role in diabetes management and control of its complications. This effect was confirmed via in vitro and docking studies against eight target proteins including α-amylase and α-glucosidase. In this report, Narirutin displayed hydrogen bonding interactions with both enzymes [13,15,27]. Moreover, variable flavonoids were previously tested for α-amylase and α-glucosidase inhibitory activity using in vitro testing and molecular docking approaches. The ligand–enzyme complexes for these compounds were studied, and it was concluded that the interactions occur mainly through H-bonding [54,55].
On the other side, the alkaloid Eburnamonine (2) established four hydrophobic interactions only inside the enzyme’s active site with TRP-58 TRP-59, TYR-62, and LEU-165 without any H-bonds (Figure 4A). Eburnamonine is an alkaloid that was previously isolated from several Vinica species and stated to contribute to the recorded antidiabetic effect of the total extract, via increasing hepatic utilization of glucose, suppressing the gluconeogenic enzymes, and regulation of insulin secretion, glucose, and lipid metabolism [17,56,57]. One more alkaloid, the isoquinoline alkaloid Isochondrodendrine (24), showed a remarkable result where it achieved the highest docking score (−9.1 kcal/mol) among all tested compounds. It established two H-bonds with THR-163 and ASP-300 together with a single hydrophobic interaction with TRP-59 (Figure 4D). Notably, this is the first report for the α-amylase enzyme inhibitory potential of Isochondrodendrine (24). However, other alkaloids, e.g., Topetecan and Cathine, were previously studied, and docking results concluded potent inhibitory activity against α-amylase enzyme [58].
Regarding the α-glucosidase enzyme, the three flavonoids Pelargonidin 3-O-rutinoside (10), Sophora isoflavanone A (14), and Dracorubin (23) achieved the best scores for binding affinity. They showed different binding interactions inside the enzyme’s active site (Figure 5). Pelargonidin 3-O-rutinoside and Sophora isoflavanone A established interactions highly similar to that of the co-crystalized inhibitor Miglitol, where H-bonds were the predominant, e.g., with ASP-203, ASP-327, TRP-406, ASP-443, ASN-449, ARG-526, ASP-542, and HIS-600 (Figure 5A,B). On the other hand, Dracorubin’s major interactions were hydrophobic (e.g., with TRP-406, PHE-450, and LYS-480) in addition to a single H-bond with GLN-603 (Figure 5C). It is worth mentioning that this is the first report on the anti-enzyme activity of both compounds Sophora isoflavanone A and Dracorubin. On the other side, the anthocyanin compound, Pelargonidin 3-O-rutinoside, was previously reported to be a potent novel α-glucosidase inhibitor that can improve postprandial hyperglycemia [28,59]. Herein, Pelargonidin 3-O-rutinoside showed a promising dual inhibitory activity against both enzymes α-amylase and α-glucosidase, expressed as binding energy (−8.5 and −8.4 kcal/mol, respectively), which was better than that of both co-crystalized inhibitors of the two corresponding enzymes (−8.1 and −8.0 kcal, mol, respectively). The current results augmented the previous finding for α-glucosidase inhibitory activity in addition to providing further proof of the α-amylase inhibitory effect. Accordingly, this study nominated Pelargonidin 3-O-rutinoside as a potential antidiabetic agent.
- Bridging the metabolomic profiling of A. alopecuroides with its biological activity.
In a research program dedicated to investigation of the biological potential and the phytochemical content of halophytes, A. alopecuroides was acknowledged for its characteristic antidiabetic activity [7]. Hence, it was crucial to explore the phytoconstituents in this species that might be responsible for such characteristic activity. In order to achieve this goal, the LC-HRMS/MS technique was employed. The current results addressed the richness of AAE with a wide variety of secondary metabolite classes. Among them, alkaloids and flavonoids are the most characteristic. Afterwards, the molecular docking technique was used to assess the antidiabetic potential of all dereplicated compounds. Docking results indicated the probable potential of most of the annonated compounds (binding energies ranging from −5 to −9); however, the most characteristic results were recorded by alkaloid and flavonoid constituents. Some of these constituents, e.g., Eburnamonine, Narirutin, and Pelargonidin 3-O-rutinoside, were previously reported for such activity, thus giving an interpretation for the observed antidiabetic activity of the crude extract. Other metabolites, such as Isochondrodendrine (−9.1 Kcal/mol, α-amylase), Sophora isoflavanone A (−9.1 Kcal/mol, α-glucosidase), and Dracorubin (−8.3 Kcal/mol, α-amylase), were noted for the first time as potential antidiabetic compounds. This finding adds more explanation for the observed activity of the total extract. Among the remaining dereplicated compounds, Pinoresinol glucoside was previously stated as a potent antidiabetic natural entity [32]. Herein, Pinoresinol glucoside displayed good inhibitory activity against α-amylase enzyme (−7.9 Kcal/mol) and α-glucosidase (−6.0 Kcal/mol). Other compounds such as Heliettin, 1,2-Dehydroreticuline, Epinorlycoramine, N-Feruloyltyramine, and Veraguensin also displayed good activity, expressed as binding energy in the range of −7.5 to −7.9 Kcal/mol. In conclusion, AAE contains a powerful mixture of phytoconstituents that could be considered, either individually or collectively, as a probable antidiabetic agents.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/life12111852/s1. Detailed material and method of docking study. References [60,61,62,63] are cited in the supplementary materials. Figure S1: 2D diagram of the binding interactions of the twenty-seven compounds detected in A. alopecuroides against α-Glucosidase enzyme. Figure S2: 2D diagram of the binding interactions of the twenty-seven compounds detected in A. alopecuroides against α-Amylase enzyme.
Author Contributions
Conceptualization, E.A.; investigation, E.A. and M.S.A.-B.; methodology, E.A. and M.H.A.H.; software, M.S.A.-B. and H.A.M.; writing—original draft preparation, E.A.; writing—review and editing, E.A. and M.H.A.H.; supervision, E.A. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the Deanship of Scientific research, Qassim University, grant number (10122-pharmacy-2020-1-3-I).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All Data are contained within the article and Supplementary Materials.
Acknowledgments
The authors gratefully acknowledge Qassim University, represented by the Deanship of Scientific research, for financial support of this research under the number (10122-pharmacy-2020-1-3-I) during the academic year 1441AH-2021 AD.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Belayneh, Y.M.; Birru, E.M. Antidiabetic activities of hydromethanolic leaf extract of Calpurnia aurea (Ait.) Benth. Subspecies aurea (Fabaceae) in mice. Evid. Based Complement. Altern. Med. 2018, 2018, 3509073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alqahtani, A.S.; Hidayathulla, S.; Rehman, T.; ElGamal, A.A.; Al-Massarani, S.; Razmovski-Naumovski, V.; Alqahtani, M.S.; El Dib, R.A.; AlAjmi, M.F. Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia. Biomolecules 2019, 10, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajaria, D.; Tiwari, S.; Tripathi, J.; Tripathi, Y.; Ranjana. In-vitro α amylase and glycosidase inhibitory effect of ethanolic extract of antiasthmatic drug—Shirishadi. J. Am. Pharm. Technol. Res. 2013, 4, 206. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Zhu, L.; Chen, Y.; Tian, J.; Wang, Y. In vivo and in vitro antioxidant activity and α-glucosidase, α-amylase inhibitory effects of flavonoids from Cichorium glandulosum seeds. Food Chem. 2013, 139, 59–66. [Google Scholar] [CrossRef]
- Nair, S.S.; Kavrekar, V.; Mishra, A. In vitro studies on alpha amylase and alpha glucosidase inhibitory activities of selected plant extracts. Eur. J. Exp. Biol. 2013, 3, 128–132. [Google Scholar]
- Mumtaz, M.W.; Al-Zuaidy, M.H.; Hamid, A.A.; Danish, M.; Akhtar, M.T.; Mukhtar, H. Metabolite profiling and inhibitory properties of leaf extracts of Ficus benjamina towards α-glucosidase and α-amylase. Int. J. Food Prop. 2018, 21, 1560–1574. [Google Scholar] [CrossRef] [Green Version]
- Amin, E.; Abdel-Bakky, M.S.; Darwish, M.A.; Mohammed, H.A.; Chigurupati, S.; Qureshi, K.A.; Hassan, M.H. The Glycemic Control Potential of Some Amaranthaceae Plants, with Particular Reference to In Vivo Antidiabetic Potential of Agathophora alopecuroides. Molecules 2022, 27, 973. [Google Scholar] [CrossRef]
- Amin, E.; Abdel-Bakky, M.S.; Mohammed, H.A.; Chigrupati, S.; Qureshi, K.A.; Hassan, M.H.A. Phytochemical Analysis and Evaluation of the Antioxidant and Antimicrobial Activities of Five Halophytes from Qassim Flora. Pol. J. Environ. Stud. 2022, 31, 3005–3012. [Google Scholar] [CrossRef]
- Chikhi, I.; Allali, H.; Dib, M.E.A.; Medjdoub, H.; Tabti, B. Antidiabetic activity of aqueous leaf extract of Atriplex halimus L. (Chenopodiaceae) in streptozotocin–induced diabetic rats. Asian Pac. J. Trop. Dis. 2014, 4, 181–184. [Google Scholar] [CrossRef]
- Othman, A.; Sayed, A.M.; Amen, Y.; Shimizu, K. Possible neuroprotective effects of amide alkaloids from Bassia indica and Agathophora alopecuroides: In vitro and in silico investigations. RSC Adv. 2022, 12, 18746–18758. [Google Scholar] [CrossRef]
- Sashidhara, K.V.; Rosaiah, J.N. Various Dereplication Strategies Using LC-MS for Rapid Natural Product Lead Identification and Drug Discovery. Nat. Prod. Commun. 2007, 2, 193–202. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, L.; Jayakanthan, K.; Mohan, S.; Nasi, R.; Johnston, B.D.; Pinto, B.M.; Rose, D.R. New Glucosidase Inhibitors from an Ayurvedic Herbal Treatment for Type 2 Diabetes: Structures and Inhibition of Human Intestinal Maltase-Glucoamylase with Compounds from Salacia reticulata. Biochemistry 2009, 49, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.K.; Zhang, X.; Caner, S.; Tysoe, C.; Nguyen, N.T.; Wicki, J.; Williams, D.E.; Coleman, J.; McNeill, J.H.; Yuen, V.; et al. The amylase inhibitor montbretin A reveals a new glycosidase inhibition motif. Nat. Chem. Biol. 2015, 11, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Bastida, J.; Viladomat, F.; Bergoñon, S.; Fernandez, J.M.; Codina, C.; Rubiralta, M.; Quirion, J.-C. Alkaloids from Narcissus leonensis. Phytochemistry 1993, 34, 1656–1658. [Google Scholar] [CrossRef]
- Kam, T.-S.; Tan, P.-S.; Chuah, C.-H. Alkaloids from leaves of Kopsia larutensis. Phytochemistry 1992, 31, 2936–2938. [Google Scholar] [CrossRef]
- Cahlíková, L.; Vaněčková, N.; Šafratová, M.; Breiterová, K.; Blunden, G.; Hulcová, D.; Opletal, L. The Genus Nerine Herb. (Amaryllidaceae): Ethnobotany, Phytochemistry, and Biological Activity. Molecules 2019, 24, 4238. [Google Scholar] [CrossRef] [Green Version]
- Cahlíková, L.; Zavadil, S.; Macáková, K.; Valterova, I.; Kulhánková, A.; Hostalkova, A.; Kuneš, J.; Opletal, L. Isolation and Cholinesterase Activity of Amaryllidaceae Alkaloids from Nerine bowdenii. Nat. Prod. Commun. 2011, 6, 1827–1830. [Google Scholar] [CrossRef] [Green Version]
- Piboonprai, K.; Khumkhrong, P.; Khongkow, M.; Yata, T.; Ruangrungsi, N.; Chansriniyom, C.; Iempridee, T. Anticancer activity of arborinine from Glycosmis parva leaf extract in human cervical cancer cells. Biochem. Biophys. Res. Commun. 2018, 500, 866–872. [Google Scholar] [CrossRef]
- Hagel, J.M.; Facchini, P.J. Benzylisoquinoline Alkaloid Metabolism: A Century of Discovery and a Brave New World. Plant Cell Physiol. 2013, 54, 647–672. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, Y.; Moriyasu, M.; Ichimaru, M.; Iwasa, K.; Kato, A.; Mathenge, S.G.; Mutiso, P.B.C.; Juma, F.D. Quaternary isoquinoline alkaloids from Xylopia parviflora. Phytochemistry 2004, 65, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Steyn, P.S.; Vleggaar, R.; Wessels, P.L.; Cole, R.J.; Scott, D.B. Structure and carbon-13 nuclear magnetic resonance assignments of versiconal acetate, versiconol acetate, and versiconol, metabolites from cultures of Aspergillus parasiticus treated with dichlorvos. J. Chem. Soc. Perkin Trans. 1 1979, 451–459. [Google Scholar] [CrossRef]
- Mitscher, L.A.; Gollapudi, S.R.; Oburn, D.S.; Drake, S. Antimicrobial agents from higher plants: Two dimethylbenzisochromans from Karwinskia humboldtiana. Phytochemistry 1985, 24, 1681–1683. [Google Scholar] [CrossRef]
- Li, Y.; Xie, S.; Ying, J.; Wei, W.; Gao, K. Chemical Structures of Lignans and Neolignans Isolated from Lauraceae. Molecules 2018, 23, 3164. [Google Scholar] [CrossRef] [Green Version]
- Macías-Villamizar, V.; Cuca-Suárez, L.; González, F.V.; Rodríguez, S. Lignoids Isolated from Nectandra turbacensis (Kunth) Nees (Lauraceae). Rec. Nat. Prod. 2016, 10, 654. [Google Scholar]
- Qurtam, A.A.; Mechchate, H.; Es-Safi, I.; Al-Zharani, M.; Nasr, F.A.; Noman, O.M.; Aleissa, M.; Imtara, H.; Aleissa, A.M.; Bouhrim, M.; et al. Citrus Flavanone Narirutin, In Vitro and In Silico Mechanistic Antidiabetic Potential. Pharmaceutics 2021, 13, 1818. [Google Scholar] [CrossRef]
- Xu, Y.; Xie, L.; Xie, J.; Liu, Y.; Chen, W. Pelargonidin-3-O-rutinoside as a novel α-glucosidase inhibitor for improving postprandial hyperglycemia. Chem. Commun. 2018, 55, 39–42. [Google Scholar] [CrossRef]
- Viljoen, A.; Mncwangi, N.; Vermaak, I. Anti-Inflammatory Iridoids of Botanical Origin. Curr. Med. Chem. 2012, 19, 2104–2127. [Google Scholar] [CrossRef] [Green Version]
- Ersöz, T.; Berkman, M.; Taşdemir, D.; Çaliş, İ.; Ireland, C.M. Iridoid and phenylethanoid glycosides from Euphrasia pectinata. Turk. J. Chem. 2002, 26, 178–188. [Google Scholar]
- Toebes, A.H.W.; de Boer, V.; Verkleij, J.A.C.; Lingeman, H.; Ernst, W.H.O. Extraction of Isoflavone Malonylglucosides from Trifolium pratense L. J. Agric. Food Chem. 2005, 53, 4660–4666. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Ashour, M.L.; El-Beshbishy, H.A.; Ahmed Hamza, A.; Singab, A.N.B.; Wink, M. Pinoresinol-4-O-β-D-glucopyranoside: A lignan from prunes (Prunus domestica) attenuates oxidative stress, hyperglycaemia and hepatic toxicity in vitro and in vivo. J. Pharm. Pharmacol. 2020, 72, 1830–1839. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Ichinose, K.; Takahashi, C.; Ho, F.-C.; Wu, J.-B.; Sankawa, U. Chemical studies on Sophora tomentosa: The isolation of a new class of isoflavonoid. Chem. Pharm. Bull. 1990, 38, 2756–2759. [Google Scholar] [CrossRef]
- Liang, Z.; Yang, M.; Xu, X.; Xie, Z.; Huang, J.; Li, X.; Yang, D. Isolation and purification of geniposide, crocin-1, and geniposidic acid from the fruit of Gardenia jasminoides Ellis by high-speed counter-current chromatography. Sep. Sci. Technol. 2014, 49, 1427–1433. [Google Scholar] [CrossRef]
- Forino, M.; Tartaglione, L.; Dell’Aversano, C.; Ciminiello, P. NMR-based identification of the phenolic profile of fruits of Lycium barbarum (goji berries). Isolation and structural determination of a novel N-feruloyl tyramine dimer as the most abundant antioxidant polyphenol of goji berries. Food Chem. 2016, 194, 1254–1259. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef]
- Johnsson, P.; Peerlkamp, N.; Kamal-Eldin, A.; Andersson, R.E.; Andersson, R.; Lundgren, L.N.; Åman, P. Polymeric fractions containing phenol glucosides in flaxseed. Food Chem. 2002, 76, 207–212. [Google Scholar] [CrossRef]
- Terazawa, M.; Okuyama, H.; Miyake, M. Isolation of coniferin and syringin from the cambial tissue and inner-bark of some angiospermous woods. J. Jpn. Wood Res. Soc. 1984, 30, 409–412. [Google Scholar]
- Sticher, O.; Lahloub, M.F. Phenolic Glycosides of Paulownia tomentosa Bark. Planta Med. 1982, 46, 145–148. [Google Scholar] [CrossRef]
- Tram, N.T.N.; Mitova, M.; Bankova, V.; Handjieva, N.; Popov, S.S. GC-MS of Crinum latifolium L. alkaloids. Z. Nat. C 2002, 57, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Zalkow, L.; Ekpo, B.; Gelbaum, L.; Harris III, R.; Keinan, E.; Novak Jr, J.; Ramming, C.; Van Derveer, D. The benzofurans of Isocoma wrightii. Structure and stereochemistry. J. Nat. Prod. 1979, 42, 203–219. [Google Scholar] [CrossRef] [PubMed]
- De Moura, N.F.; Simionatto, E.; Porto, C.; Hoelzel, S.C.S.; Dessoy, E.C.S.; Zanatta, N.; Morel, A.F. Quinoline Alkaloids, Coumarins and Volatile Constituents of Helietta longifoliata. Planta Med. 2002, 68, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.E.; de Arias, A.R.; Yaluff, G.; de Bilbao, N.V.; Nakayama, H.; Torres, S.; Schinini, A.; Guy, I.; Heinzen, H.; Fournet, A. Antileishmanial activity of furoquinolines and coumarins from Helietta apiculata. Phytomedicine 2010, 17, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.N.H.; Ho, W.S. Antiproliferative activity of gambogic acid isolated from Garcinia hanburyi in Hep3B and Huh7 cancer cells. Oncol. Rep. 2013, 29, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.J.; Sousa, M.; Parola, A.J.; de Melo, J.S.S.; Catarino, F.; Marçalo, J.; Pina, F. Identification of 7, 4′-Dihydroxy-5-methoxyflavylium in “Dragon’s Blood”: To Be or Not to Be an Anthocyanin. Chem. Eur. J. 2007, 13, 1417–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Li, F.; Yan, X.; Cheng, Q.; Xue, R.; Yu, H.; Li, Z.; Wang, C. Phytochemical Constituents and Biological Activities of Plants from the Genus cissampelos. Chem. Biodivers. 2021, 18, e2100358. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, A.; Wójcicki, K.; Klensporf-Pawlik, D.; Marzec, M.; Lucarini, M.; Durazzo, A.; Fonseca, J.; Santini, A.; Nowak, I.; Souto, E.B. Cold-Pressed Pomegranate Seed Oil: Study of Punicic Acid Properties by Coupling of GC/FID and FTIR. Molecules 2022, 27, 5863. [Google Scholar] [CrossRef]
- Lannang, A.M.; Anjum, S.; Tangmouo, J.G.; Krohn, K.; Choudhary, M.I. Conessine isolated from Holarrhena floribunda. Acta Crystallogr. Sect. E Struct. Rep. Online 2007, 63, o4398. [Google Scholar] [CrossRef]
- Ikeda, T.; Udayama, M.; Okawa, M.; Arao, T.; Kinjo, J.; Nohara, T. Partial Hydrolysis of Soyasaponin I and the Hepatoprotective Effects of the Hydrolytic Products. Studies on the Hepatoprotective Drugs. Part IV. (Studies on the Constituents of the Leguminous Plants. Part LVII.) Study of the Structure-Hepatoprotective Relationship of Soyasapogenol B Analogs. Chem. Pharm. Bull. 1998, 46, 359–361. [Google Scholar] [CrossRef] [Green Version]
- Udayama, M.; Ohkawa, M.; Yoshida, N.; Kinjo, J.; Nohara, T. Structures of Three New Oleanene Glucuronides Isolated from Lathyrus palustris var. pilosus and Hepatoprotective Activity. Chem. Pharm. Bull. 1998, 46, 1412–1415. [Google Scholar] [CrossRef] [Green Version]
- Dwuma-Badu, D.; Ayim, J.S.; Mingle, C.; Tackie, A.; Slatkin, D.; Knapp, J.; Schiff, P., Jr. Alkaloids of Cissampelos pareira. Phytochemistry 1975, 14, 2520–2521. [Google Scholar] [CrossRef]
- Steyn, P.S.; Vleggaar, R.; Wessels, P.L.; Scott, D.B. Biosynthesis of versiconal acetate, versiconol acetate, and versiconol, metabolites from cultures of Aspergillus parasiticus treated with dichlorvos. The role of versiconal acetate in aflatoxin biosynthesis. J. Chem. Soc. Perkin Trans. 1 1979, 460–463. [Google Scholar] [CrossRef]
- Yao, R.C.; Hsieh, D.P. Step of dichlorvos inhibition in the pathway of aflatoxin biosynthesis. Appl. Microbiol. 1974, 28, 52–57. [Google Scholar] [CrossRef]
- Jadalla, B.M.I.S.; Moser, J.J.; Sharma, R.; Etsassala, N.G.E.R.; Egieyeh, S.A.; Badmus, J.A.; Marnewick, J.L.; Beukes, D.; Cupido, C.N.; Hussein, A.A. In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents. Separations 2022, 9, 190. [Google Scholar] [CrossRef]
- Ahmed, S.; Ali, C.; Ruma, R.A.; Mahmud, S.; Paul, G.K.; Saleh, A.; Alshahrani, M.M.; Obaidullah, A.J.; Biswas, S.K.; Rahman, M.; et al. Molecular Docking and Dynamics Simulation of Natural Compounds from Betel Leaves (Piper betle L.) for Investigating the Potential Inhibition of Alpha-Amylase and Alpha-Glucosidase of Type 2 Diabetes. Molecules 2022, 27, 4526. [Google Scholar] [CrossRef]
- Chikezie, P.C.; Ojiako, O.A.; Nwufo, K.C. Overview of anti-diabetic medicinal plants: The Nigerian research experience. J. Diabetes Metab. 2015, 6, 546. [Google Scholar] [CrossRef]
- Pereira, A.S.; Haan, H.D.; Peña-García, J.; Moreno, M.M.; Pérez-Sánchez, H.; Apostolides, Z. Exploring African Medicinal Plants for Potential Anti-Diabetic Compounds with the DIA-DB Inverse Virtual Screening Web Server. Molecules 2019, 24, 2002. [Google Scholar] [CrossRef] [Green Version]
- Akshatha, J.V.; SantoshKumar, H.S.; Prakash, H.S.; Nalini, M.S. In silico docking studies of α-amylase inhibitors from the anti-diabetic plant Leucas ciliata Benth. and an endophyte, Streptomyces longisporoflavus. 3 Biotech 2021, 11, 51. [Google Scholar] [CrossRef]
- Oliveira, H.; Fernandes, A.; Brás, N.F.; Mateus, N.; de Freitas, V.; Fernandes, I. Anthocyanins as antidiabetic agents—In vitro and in silico approaches of preventive and therapeutic effects. Molecules 2020, 25, 3813. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. Cheminform. J. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastman, P.; Friedrichs, M.S.; Chodera, J.D.; Radmer, R.J.; Bruns, C.M.; Ku, J.P.; Beauchamp, K.A.; Lane, T.J.; Wang, L.; Shukla, D.; et al. OpenMM 4: A Reusable, Extensible, Hardware Independent Library for High Performance Molecular Simulation. J. Chem. Theory Comput. 2013, 9, 461. [Google Scholar] [CrossRef] [PubMed]
- Dallakyan, S.; Olson, A.J. Chemical Biology; Hempel, J.E., Williams, C.H., Hong, C.C., Eds.; Springer: New York, NY, USA, 2015; pp. 243–250. [Google Scholar] [CrossRef]
Figure 1.
LC-HRMS chromatogram of the dereplicated metabolites of A. alopecuroides methanolic extract (positive mode).
Figure 1.
LC-HRMS chromatogram of the dereplicated metabolites of A. alopecuroides methanolic extract (positive mode).

Figure 2.
Structures of the dereplicated compounds from A. alopecuroides methanolic extract by LC-HRMS/MS.
Figure 2.
Structures of the dereplicated compounds from A. alopecuroides methanolic extract by LC-HRMS/MS.

Figure 3.
Docking score distribution of the dereplicated metabolites in AAE against both human α-amylase (A) and α-glucosidase (B) (PDB: 4W93 and 3L4W, respectively).
Figure 3.
Docking score distribution of the dereplicated metabolites in AAE against both human α-amylase (A) and α-glucosidase (B) (PDB: 4W93 and 3L4W, respectively).

Figure 4.
Docking poses of compounds with docking scores <−8.0 kcal/mol (i.e., Eburnamonine, Narirutin, Pelargonidin 3-O-rutinoside, and Isochondrodendrine along with the co-crystalized inhibitor Montbretin A) inside the human α-amylase (A–E, respectively).
Figure 4.
Docking poses of compounds with docking scores <−8.0 kcal/mol (i.e., Eburnamonine, Narirutin, Pelargonidin 3-O-rutinoside, and Isochondrodendrine along with the co-crystalized inhibitor Montbretin A) inside the human α-amylase (A–E, respectively).

Figure 5.
Docking poses of compounds with docking scores <−8.0 kcal/mol (i.e., Pelargonidin 3-O-rutinoside, Sophora isoflavanone A, and Dracorubin along with the co-crystalized inhibitor miglitol) inside the human α-glucosidase (A–D, respectively).
Figure 5.
Docking poses of compounds with docking scores <−8.0 kcal/mol (i.e., Pelargonidin 3-O-rutinoside, Sophora isoflavanone A, and Dracorubin along with the co-crystalized inhibitor miglitol) inside the human α-glucosidase (A–D, respectively).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The LC-HRMS/MS dereplication results of A. alopecuroides methanolic extract.
| No. | Tentative Identification | Rt | m/z | Formula | Ms/Ms-Fragmnets | Biological Source | Nature of Compounds | References |
|---|---|---|---|---|---|---|---|---|
| 1 | Epinorlycoramine | 1.2 | 275.152 | C16H21NO3 | 112.0874-124.0875-154.0978-190.098-202.0982-220.1087 | Narcissus leonensis plant | Alkaloid | [16] |
| 2 | Eburnamonine | 1.6 | 294.173 | C19H22N2O | 114.1028-145.0519 | Leaves of Kopsia zarutensi | Alkaloid | [17] |
| 3 | 3-Acetylnerbowdine | 2.0 | 361.153 | C19H23NO6 | 151.0774 | Bulbs of Nerine bowdenii | Alkaloid | [18,19] |
| 4 | Arborinine | 3.3 | 285.290 | C16H15NO4 | 110.0375-135.0299-153.0546 | Leaves of Glycosmis parva | Alkaloid | [20] |
| 5 | 1,2-Dehydroreticuline | 4.1 | 328.155 | C19H22NO4 | 120.0817-132.0821-166.0880-178.0875 | Bark and the root of Xylopia parviflora | Alkaloid | [21,22] |
| 6 | Versiconol acetate | 6.7 | 402.095 | C20H18O9 | 143.0711 | Culture of Aspergillus parasiticus | Anthraquinone | [23] |
| 7 | Karwinaphthol B | 7.0 | 288.136 | C17H20O4 | 112.0879-117.0333-138.0664-145.0289 | Roots of Karwinskia humboldtiana | Benzisochromans | [24] |
| 8 | Veraguensin | 7.4 | 372.194 | C22H28O5 | 177.0550-196.1444-222.1249 | Leaves and root bark of Nectandra turbacensis (Kunth) Nees | Lignan | [25,26] |
| 9 | Narirutin | 8.6 | 580.179 | C27H32O14 | 313.0713-415.1029-433.1137 | Citrus fruits | Flavanone glycoside | [27] |
| 10 | Pelargonidin 3-O-rutinoside | 8.6 | 579.171 | C27H31O14 | 313.0713-397.0918 415.1029-433.1137 | Strawberries | Anthocyanin | [28] |
| 11 | Boschnaloside | 8.8 | 344.147 | C16H24O8 | 151.0742-177.0546-186.0527 | Boschniakia rossica plant | Iridoid glycoside | [29,30] |
| 12 | Biochanin A-β-d-glucoside | 9.0 | 446.121 | C22H22O10 | 145.0288-175.0619-177.0551 | Trifolium pratense L. plant | Isoflavone glycoside | [31] |
| 13 | Pinoresinol glucoside | 9.1 | 520.194 | C26H32O11 | 177.0552-184.0717-186.0523-191.0710 | Prunes of Prunus domestica L. | Lignan | [32] |
| 14 | Sophora isoflavanone A | 9.2 | 370.142 | C21H22O6 | 145.0285-177.0553-284.0695 | Sophora tomentosa L. plant | Isoflavone | [33] |
| 15 | Geniposide | 9.2 | 388.137 | C17H24O10 | 149.0614-151.0390-177.0552-186.0541 | Fruit of Gardenia jasminoides Ellis | Iridoid glycoside | [34] |
| 16 | N-Feruloyltyramine | 9.3 | 313.131 | C18H19NO4 | 117.0339-145.0293-149.0607-162.0539 | Fruits of Lycium barbarum (goji berries) Bassia indica and A. alopecuroides plants | Alkaloid | [10,35] |
| 17 | 4-Hydroxycinnamyl alcohol 4-d-glucoside | 9.3 | 312.121 | C15H20O7 | 117.0339-145.0293-149.0607- | Linum usitatissimum, Linn. plant | Lignan | [36,37] |
| 18 | Coniferin | 9.4 | 342.131 | C16H22O8 | 137.0600-175.0763-177.0546-218.0794 | Paulownia tomentosa bark | Lignan | [38,39] |
| 19 | Powelline | 9.7 | 301.131 | C17H19NO4 | 121.0652-135.0448-163.0395-180.0660 | Leaves from Crinum latifolium L. | Alkaloid | [40] |
| 20 | Toxyl angelate | 9.7 | 300.136 | C18H20O4 | 121.0652-135.0448-145.0286-163.0395- | Isocoma wrightii plant | Banzofuran | [41] |
| 21 | Heliettin | 10.1 | 314.152 | C19H22O4 | 121.0657-145.0289-177.0553 | Stem bark of Helietta longifoliata Britt | Furanochomarine | [42,43] |
| 22 | Gambogic acid | 10.2 | 628.304 | C38H44O8 | 121.0655-177.0549-201.0549-297.1123 | Garcinia hanburyi plant | Phenolic acid (Xanthoid derivative) | [44] |
| 23 | Dracorubin | 10.9 | 488.162 | C32H24O5 | 121.0654-177.0565-201.0545-323.0921 | Resin extracted from the tree Dracaena draco | Proanthocyanidine | [45] |
| 24 | Isochondrodendrine | 12.1 | 594.273 | C36H38N2O6 | 565.2679 | Cissampelos mucronate and Cissampelos pareira plants | Alkaloids | [46] |
| 25 | Punicic acid | 13.0 | 278.225 | C18H30O2 | 107.0864-121.1017-133.1021-135.1160-149.1334 | Pomegranate Seed Oil | Unsaturated fatty acid | [47] |
| 26 | Conessine | 13.3 | 356.319 | C24H40N2 | 121.1015-123.1154-135.1162-149.1330 | Holarrhena floribunda G. Don. plant | Alkaloid | [48] |
| 27 | Soyasapogenol B 3-O-d-glucuronide | 13.5 | 634.408 | C36H58O9 | 133.0863-177.1134-247.2054-291.2317 | Aerial parts of Lathylus palustris L. | Triterpenoid saponin | [49,50] |
Table 2.
Docking sores and estimated absolute binding free energies (in kcal/mol) of the dereplicated structures, along with those of the reported co-crystalized inhibitors.
Table 2.
Docking sores and estimated absolute binding free energies (in kcal/mol) of the dereplicated structures, along with those of the reported co-crystalized inhibitors.
| No. | Tentative Identification | Binding Energy (kcal/mol) | |
|---|---|---|---|
| α-Amylase | α-Glucosidase | ||
| 1 | Epinorlycoramine | −7.6 | −6.2 |
| 2 | Eburnamonine | −8.7 | −6.4 |
| 3 | 3-Acetylnerbowdine | −7.2 | −6.6 |
| 4 | Arborinine | −7.5 | −6.0 |
| 5 | 1,2-Dehydroreticuline | −7.4 | −7.4 |
| 6 | Versiconol acetate | −7.5 | −7.1 |
| 7 | Karwinaphthol B | −7.4 | −5.9 |
| 8 | Veraguensin | −7.5 | −6.7 |
| 9 | Narirutin | −8.5 | −7.9 |
| 10 | Pelargonidin 3-O-rutinoside | −8.5 | −8.4 |
| 11 | Boschnaloside | −7.1 | −5.8 |
| 12 | Biochanin A-β-D-glucoside | −7.2 | −6.3 |
| 13 | Pinoresinol glucoside | −7.9 | −6.0 |
| 14 | Sophora isoflavanone A | −7.4 | −9.1 |
| 15 | Geniposide | −7.2 | −6.5 |
| 16 | N-Feruloyltyramine | −7.3 | −7.6 |
| 17 | 4-Hydroxycinnamyl alcohol 4-D-glucoside | −7.3 | −6.8 |
| 18 | Coniferin | −6.9 | −5.7 |
| 19 | Powelline | −6.7 | −6.5 |
| 20 | Toxyl angelate | −7.2 | −6.4 |
| 21 | Heliettin | −7.2 | −7.1 |
| 22 | Gambogic acid | −7.2 | −7.1 |
| 23 | Dracorubin | −7.8 | −8.3 |
| 24 | Isochondrodendrine | −9.1 | −7.0 |
| 25 | Punicic acid | −5.6 | −5.7 |
| 26 | Conessine | −4.8 | −5.6 |
| 27 | Soyasapogenol B 3-O-D-glucuronide | −4.5 | −4.7 |
| STD | Montbretin A | −8.1 | …. |
| Miglitol | …. | −8.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Amin, E.; Abdel-Bakky, M.S.; Mohammed, H.A.; Hassan, M.H.A. Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides. Life 2022, 12, 1852. https://doi.org/10.3390/life12111852
AMA Style
Amin E, Abdel-Bakky MS, Mohammed HA, Hassan MHA. Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides. Life. 2022; 12(11):1852. https://doi.org/10.3390/life12111852
Chicago/Turabian StyleAmin, Elham, Mohamed Sadek Abdel-Bakky, Hamdoon A. Mohammed, and Marwa H. A. Hassan. 2022. "Chemical Profiling and Molecular Docking Study of Agathophora alopecuroides" Life 12, no. 11: 1852. https://doi.org/10.3390/life12111852
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.