
Alpha-Mangostin Reduces Pericellular Fibronectin on Suspended Tumor Cells and Therapeutically, but Not Prophylactically, Suppresses Distant Metastasis


 and
and 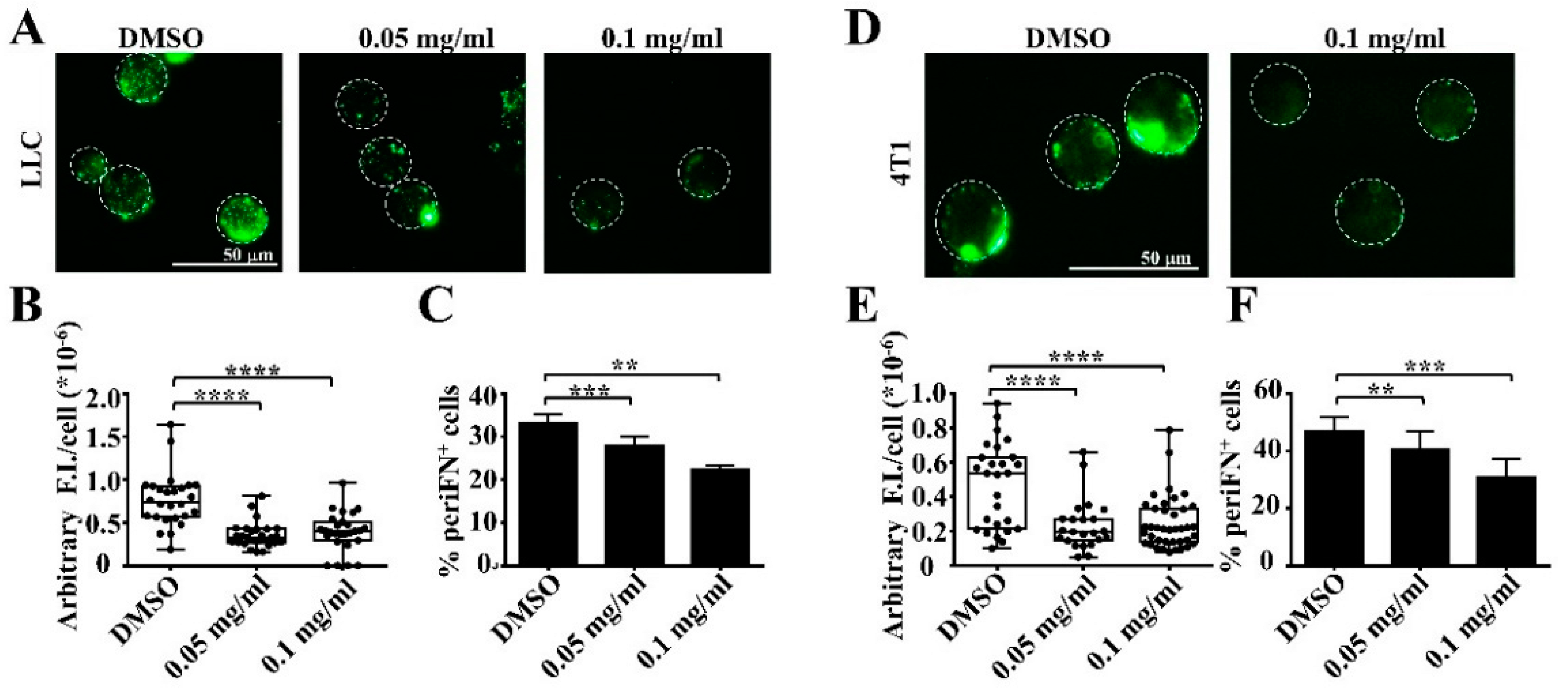{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Materials
2.3. Extraction of Mangosteen Pericarp
2.4. Immunofluorescence (IF) Staining for periFN on Suspended Tumor Cells
2.5. MTT Assay
2.6. Apoptosis Assay
2.7. Immunoblotting
2.8. Experimental Tumor Metastasis Animal Models
2.9. Statistical Analysis
3. Results
3.1. Extracts of Mangosteen Pericarps Suppress periFN Assembly on Tumor Cells in Suspension
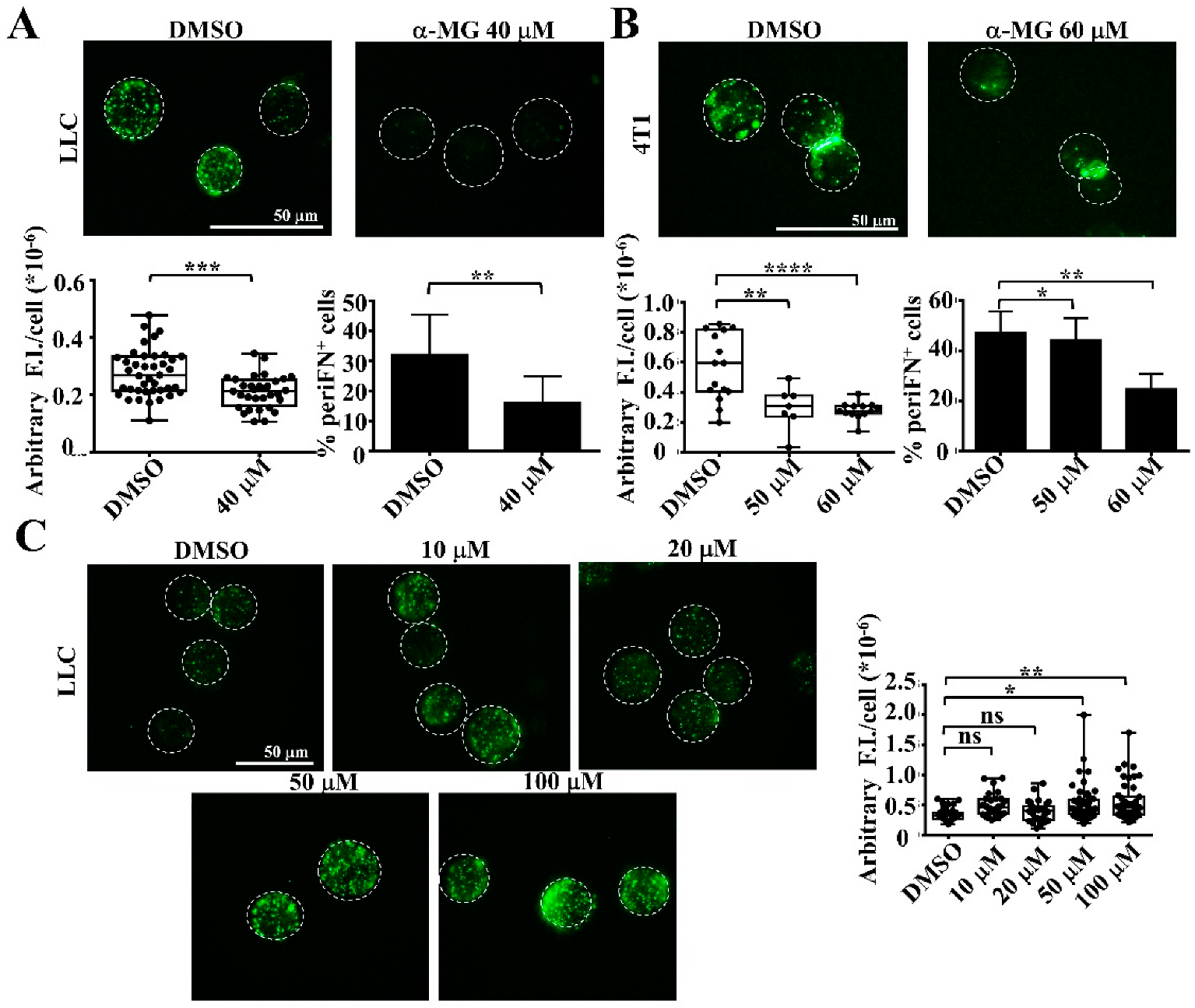
3.2. Alpha-MG, but Not Xanthone, in MP Extracts Suppresses periFN Assembly on Suspended Tumor Cells
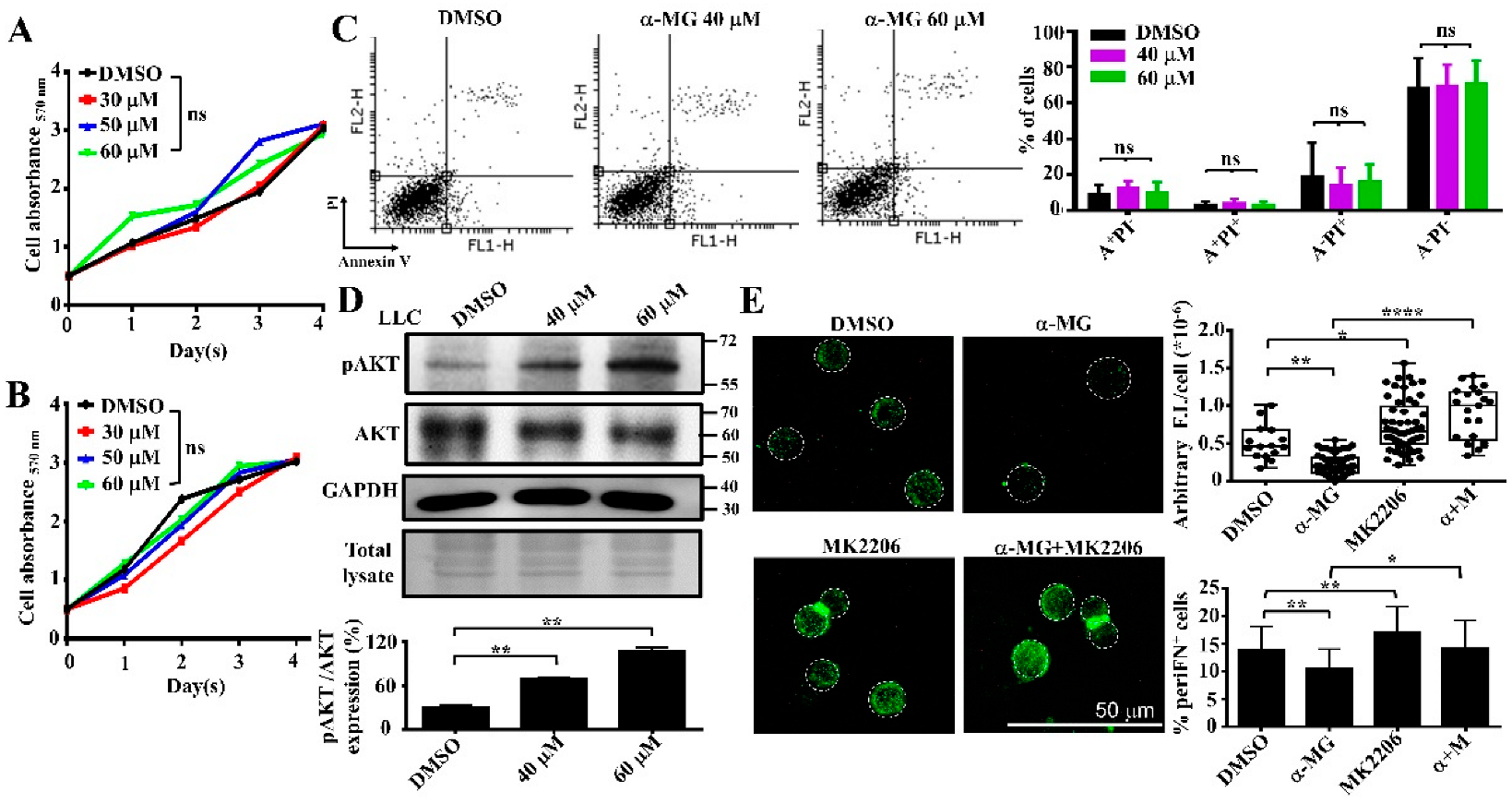
3.3. Alpha-MG Treatment Does Not Affect Tumor Cell Viability in Suspension Due to Increased AKT Activity
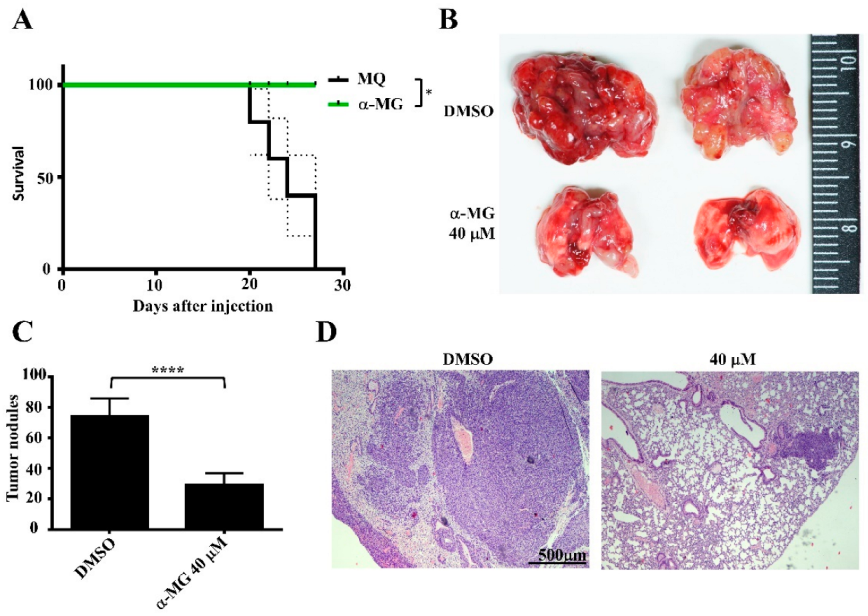
3.4. Pretreatment of Suspended Tumor Cells with α-MG Prior to Intravenous Inoculation Reduces Lung Metastasis
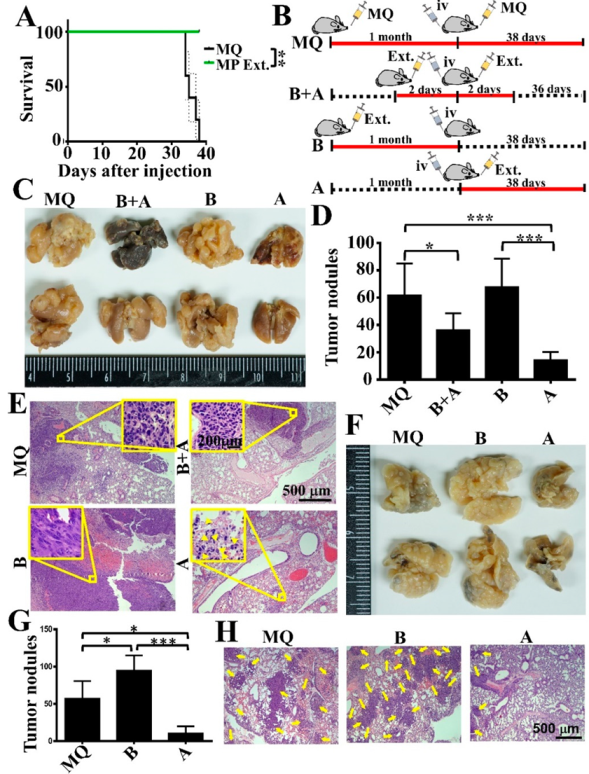
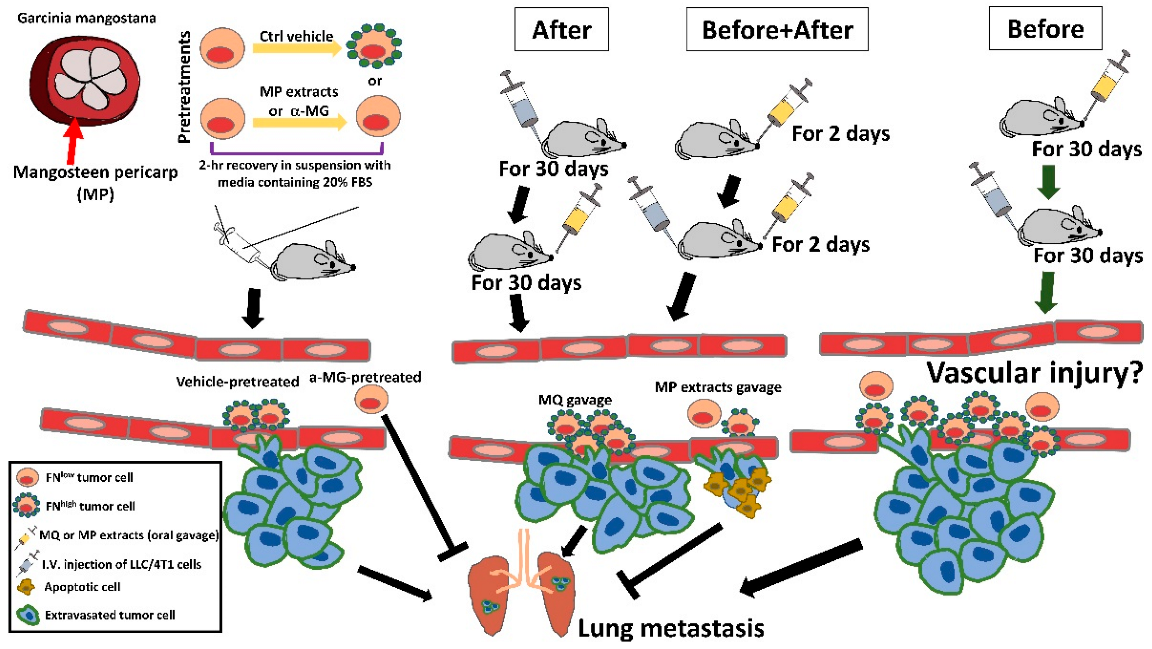
3.5. Oral Gavage with MP Extracts Therapeutically, but Not Prophylactically, Suppresses Lung Metastasis of Tumor Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gui, P.; Bivona, T.G. Evolution of metastasis: New tools and insights. Trends Cancer 2021, 8, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhang, Y.; Tian, Y.; Sun, Y.; Chen, Y.; Zhang, Y.; Zhu, Q.; Pan, J.; Qing, C. 9-Nitro-20(S)-carbonate-camptothecin (NCP4), a novel prodrug of 9-nitrocamptothecin (9-NC), exhibits potent chemotherapeutic efficacy and improved safety against hepatocarcinoma. Eur. J. Pharmacol. 2022, 923, 174898. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; Zahmatkeshan, M.; Akbarzadeh, A.; Rabiee, N.; Ahmadi, S.; Keyhanvar, P.; Rezayat, S.M.; Seifalian, A.M. Chemotherapeutic effects of Apigenin in breast cancer: Preclinical evidence and molecular mechanisms; enhanced bioavailability by nanoparticles. Biotechnol. Rep. 2022, 34, e00730. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, M.G.; Piccolo, M.; Misso, G.; Santamaria, R.; Irace, C. Bioactivity and Development of Small Non-Platinum Metal-Based Chemotherapeutics. Pharmaceutics 2022, 14, 954. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Carraher, C.; Schwarzbauer, J.E. Assembly of fibronectin extracellular matrix. Annu. Rev. Cell Dev. Biol. 2010, 26, 397–419. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-C.; Yang, C.-H.; Cheng, L.-H.; Chang, W.-T.; Lin, Y.-R.; Cheng, H.-C. Fibronectin in Cancer: Friend or Foe. Cells 2020, 9, 27. [Google Scholar] [CrossRef]
- Cheng, H.-C.; Abdel-Ghany, M.; Elble, R.C.; Pauli, B.U. Lung endothelial dipeptidyl peptidase IV promotes adhesion and metastasis of rat breast cancer cells via tumor cell surface-associated fibronectin. J. Biol. Chem. 1998, 273, 24207–24215. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-C.; Abdel-Ghany, M.; Zhang, S.; Pauli, B.U. Is the Fischer 344/CRJ rat a protein-knock-out model for dipeptidyl peptidase IV-mediated lung metastasis of breast cancer? Clin. Exp. Metastasis 1999, 17, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-C.; Abdel-Ghany, M.; Pauli, B.U. A Novel Consensus Motif in Fibronectin Mediates Dipeptidyl Peptidase IV Adhesion and Metastasis. J. Biol. Chem. 2003, 278, 24600–24607. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-J.; Russo, J.; Kohwi, Y.; Kohwi-Shigematsu, T. SATB1 reprogrammes gene expression to promote breast tumour growth and metastasis. Nature 2008, 452, 187–193. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Lee, S.-H.; Liao, I.-C.; Huang, S.-H.; Cheng, H.-C.; Liao, P.-C. Secretomic analysis identifies alpha-1 antitrypsin (A1AT) as a required protein in cancer cell migration, invasion, and pericellular fibronectin assembly for facilitating lung colonization of lung adenocarcinoma cells. Mol. Cell. Proteom. 2012, 11, 1320–1339. [Google Scholar] [CrossRef] [PubMed]
- Kenny, H.A.; Chiang, C.-Y.; White, E.A.; Schryver, E.M.; Habis, M.; Romero, I.; Ladanyi, A.; Penicka, C.V.; George, J.; Matlin, K.; et al. Mesothelial cells promote early ovarian cancer metastasis through fibronectin secretion. J. Clin. Investig. 2014, 124, 4614–4628. [Google Scholar] [CrossRef]
- Yu, M.; Ting, D.; Stott, S.L.; Wittner, B.S.; Ozsolak, F.; Paul, S.; Ciciliano, J.C.; Smas, M.E.; Winokur, D.; Gilman, A.J.; et al. RNA sequencing of pancreatic circulating tumour cells implicates WNT signalling in metastasis. Nature 2012, 487, 510–513. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Lin, J.-F.; Cheng, L.-H.; Chang, W.-T.; Kao, Y.-H.; Chang, M.-M.; Wang, B.-J.; Cheng, H.-C. Pterostilbene prevents AKT-ERK axis-mediated polymerization of surface fibronectin on suspended lung cancer cells independently of apoptosis and suppresses metastasis. J. Hematol. Oncol. 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Huang, L.; Cheng, H.-C.; Isom, R.; Chen, C.-S.; Levine, R.A.; Pauli, B.U. Protein kinase Cepsilon mediates polymeric fibronectin assembly on the surface of blood-borne rat breast cancer cells to promote pulmonary metastasis. J. Biol. Chem. 2008, 283, 7616–7627. [Google Scholar] [CrossRef] [PubMed]
- Koss-Mikołajczyk, I.; Baranowska, M.; Todorovic, V.; Albini, A.; Sansone, C.; Andreoletti, P.; Cherkaoui-Malki, M.; Lizard, G.; Noonan, D.; Sobajic, S.; et al. Prophylaxis of Non-communicable Diseases: Why Fruits and Vegetables may be Better Chemopreventive Agents than Dietary Supplements Based on Isolated Phytochemicals? Curr. Pharm. Des. 2019, 25, 1847–1860. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.Y.; Hashim, N.M.; Mariod, A.A.; Mohan, S.; Abdulla, M.A.; Abdelwahab, S.I.; Arbab, I.A. α-Mangostin from Garcinia mangostana Linn: An updated review of its pharmacological properties. Arab. J. Chem. 2016, 9, 317–329. [Google Scholar] [CrossRef]
- Rafiq, S.; Sharma, V.; Nazir, A.; Rashid, R.; Sa, S.; Nayik, F.N.A.G.A. Development of Probiotic Carrot Juice. J. Nutr. Food Sci. 2016, 6, 2. [Google Scholar] [CrossRef]
- Ames, B.N.; Gold, L.S. The Prevention of Cancer. Drug Metab. Rev. 1998, 30, 201–223. [Google Scholar] [CrossRef]
- Borek, C. Dietary Antioxidants and Human Cancer. Integr. Cancer Ther. 2004, 3, 333–341. [Google Scholar] [CrossRef]
- Jung, H.-A.; Su, B.-N.; Keller, W.J.; Mehta, R.G.; Kinghorn, A.D. Antioxidant xanthones from the pericarp of Garcinia mangostana (Mangosteen). J. Agric. Food Chem. 2006, 54, 2077–2082. [Google Scholar] [CrossRef] [PubMed]
- Peres, V.; Nagem, T.J.; de Oliveira, F.F. Tetraoxygenated naturally occurring xanthones. Phytochemistry 2000, 55, 683–710. [Google Scholar] [CrossRef]
- Zhu, X.; Li, J.; Ning, H.; Yuan, Z.; Zhong, Y.; Wu, S.; Zeng, J.-Z. α-Mangostin Induces Apoptosis and Inhibits Metastasis of Breast Cancer Cells via Regulating RXRα-AKT Signaling Pathway. Front. Pharmacol. 2021, 12, 739658. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Zhang, L.; Ji, H.; Kou, Y.; Ou, B. Bioavailability and antioxidant effects of a xanthone-rich mangosteen (Garcinia mangostana) product in humans. J. Agric. Food Chem. 2009, 57, 8788–8792. [Google Scholar] [CrossRef] [PubMed]
- Paknikar, S.K.; Kadam, S.H.; Ehrlich, A.L.; Bates, R.B. Alternate Biosynthesis of Valerenadiene and Related Sesquiterpenes. Nat. Prod. Commun. 2013, 8, 1195–1196. [Google Scholar] [CrossRef]
- Huang, L.-T.; Tsai, C.-L.; Huang, S.-H.; Chang, M.-M.; Chang, W.-T.; Cheng, L.-H.; Cheng, H.-C. Depleting RhoA/Stress Fiber-Organized Fibronectin Matrices on Tumor Cells Non-Autonomously Aggravates Fibroblast-Driven Tumor Cell Growth. Int. J. Mol. Sci. 2020, 21, 8272. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, F.; Dong, Y.; Ren, F. α-Mangostin Induces Apoptosis in Human Osteosarcoma Cells Through ROS-Mediated Endoplasmic Reticulum Stress via the WNT Pathway. Cell Transplant. 2021, 30, 9636897211035080. [Google Scholar] [CrossRef]
- Shan, T.; Ma, Q.; Guo, K.; Liu, J.; Li, W.; Wang, F.; Wu, E. Xanthones from mangosteen extracts as natural chemopreventive agents: Potential anticancer drugs. Curr. Mol. Med. 2011, 11, 666–677. [Google Scholar] [CrossRef]
- Zhang, K.-J.; Gu, Q.-L.; Yang, K.; Ming, X.-J.; Wang, J.-X. Anticarcinogenic Effects of α-Mangostin: A Review. Planta Medica 2016, 83, 188–202. [Google Scholar] [CrossRef]
- Shariati, M.; Meric-Bernstam, F. Targeting AKT for cancer therapy. Expert Opin. Investig. Drugs 2019, 28, 977–988. [Google Scholar] [CrossRef]
- Yu, T.; Huang, X.; Liu, J.; Fu, Q.; Wang, B.; Qian, Z. Polymeric nanoparticles encapsulating α-mangostin inhibit the growth and metastasis in colorectal cancer. Appl. Mater. Today 2019, 16, 351–366. [Google Scholar] [CrossRef]
- Ocaña, A.; Vera-Badillo, F.; Al-Mubarak, M.; Templeton, A.J.; Corrales-Sánchez, V.; Díez-González, L.; Cuenca-Lopez, M.D.; Seruga, B.; Pandiella, A.; Amir, E. Activation of the PI3K/mTOR/AKT pathway and survival in solid tumors: Systematic review and meta-analysis. PLoS ONE 2014, 9, e95219. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wu, L.; Chen, Y.; Li, Y.; Wang, Q.; Li, M.; Hao, K.; Zhang, W.; Jiang, S.; Wang, Z. Cytotoxic and Antiproliferative Effects of β-Mangostin on Rat C6 Glioma Cells Depend on Oxidative Stress Induction via PI3K/AKT/mTOR Pathway Inhibition. Drug Des. Dev. Ther. 2020, 14, 5315–5324. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Ma, J.; Lei, J.; Duan, W.; Sheng, L.; Chen, X.; Hu, A.; Wang, Z.; Wu, Z.; Wu, E.; et al. α-mangostin suppresses the viability and epithelial-mesenchymal transition of pancreatic cancer cells by downregulating the PI3K/Akt pathway. BioMed Res. Int. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.-J.; Kuo, H.-C.; Cheng, L.-H.; Lee, Y.-H.; Chang, W.-T.; Wang, B.-J.; Wang, Y.-J.; Cheng, H.-C. Apoptotic and Nonapoptotic Activities of Pterostilbene against Cancer. Int. J. Mol. Sci. 2018, 19, 287. [Google Scholar] [CrossRef]
- Wawszczyk, J.; Jesse, K.; Smolik, S.; Kapral, M. Mechanism of Pterostilbene-Induced Cell Death in HT-29 Colon Cancer Cells. Molecules 2022, 27, 369. [Google Scholar] [CrossRef]
- Chen, H.; Ma, J.; Liu, J.; Dou, L.; Shen, T.; Zuo, H.; Xu, F.; Zhao, L.; Tang, W.; Man, Y.; et al. Lysophosphatidylcholine disrupts cell adhesion and induces anoikis in hepatocytes. FEBS Lett. 2022, 596, 510–525. [Google Scholar] [CrossRef]
- Jain, K.; Basu, A. The Multifunctional Protein Kinase C-ε in Cancer Development and Progression. Cancers 2014, 6, 860–878. [Google Scholar] [CrossRef]
- Mary, E.R. Protein kinase C isoforms: Multi-functional regulators of cell life and death. Front. Biosci. 2009, 14, 2386–2399. [Google Scholar] [CrossRef]
- Jain, K.; Basu, A. Protein Kinase C-ε Promotes EMT in Breast Cancer. Breast Cancer Basic Clin. Res. 2014, 8, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Muscella, A.; Calabriso, N.; Vetrugno, C.; Urso, L.; Fanizzi, F.P.; De Pascali, S.A.; Marsigliante, S. Sublethal concentrations of the platinum(II) complex [Pt(O,O′-acac)(γ-acac)(DMS)] alter the motility and induce anoikis in MCF-7 cells. J. Cereb. Blood Flow Metab. 2010, 160, 1362–1377. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Montero, C.M.; Wygant, J.N.; McIntyre, B.W. PI3-K/Akt-mediated anoikis resistance of human osteosarcoma cells requires Src activation. Eur. J. Cancer 2006, 42, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Orozco, F.; Chitchumroonchokchai, C.; Lesinski, G.B.; Suksamrarn, S.; Failla, M.L. α-Mangostin: Anti-inflammatory activity and metabolism by human cells. J. Agric. Food Chem. 2013, 61, 3891–3900. [Google Scholar] [CrossRef]
- Tang, Y.-P.; Li, P.-G.; Kondo, M.; Ji, H.-P.; Kou, Y.; Ou, B. Effect of a mangosteen dietary supplement on human immune function: A randomized, double-blind, placebo-controlled trial. J. Med. Food 2009, 12, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-K.; Liu, C.-X.; Zhai, Y.-Y.; Li, L.-L.; Wang, X.-L.; Feng, W.-S. Protection effect of amentoflavone in Selaginella tamariscina against TNF-alpha-induced vascular injury of endothelial cells. Yao Xue Xue Bao 2013, 48, 1503–1509. [Google Scholar]
- Zhang, H.; Park, Y.; Wu, J.; Chen, X.P.; Lee, S.; Yang, J.; Dellsperger, K.C.; Zhang, C. Role of TNF-α in vascular dysfunction. Clin. Sci. 2009, 116, 219–230. [Google Scholar] [CrossRef]
- Abbate, A.; Toldo, S.; Marchetti, C.; Kron, J.; Van Tassell, B.W.; Dinarello, C.A. Interleukin-1 and the Inflammasome as Therapeutic Targets in Cardiovascular Disease. Circ. Res. 2020, 126, 1260–1280. [Google Scholar] [CrossRef]
- Ruan, C.-C.; Gao, P.-J. Role of Complement-Related Inflammation and Vascular Dysfunction in Hypertension. Hypertension 2019, 73, 965–971. [Google Scholar] [CrossRef]
- Zimmerman, M.A.; Flores, S.C. Autoimmune-Mediated Oxidative Stress and Endothelial Dysfunction: Implications of Accelerated Vascular Injury in Type I Diabetes. J. Surg. Res. 2009, 155, 173–178. [Google Scholar] [CrossRef]
- Bielawska-Pohl, A.; Crola, C.; Caignard, A.; Gaudin, C.; Dus, D.; Kieda, C.; Chouaib, S. Human NK cells lyse organ-specific endothelial cells: Analysis of adhesion and cytotoxic mechanisms. J. Immunol. 2005, 174, 5573–5582. [Google Scholar] [CrossRef]
- Yan, X.; Hegab, A.E.; Endo, J.; Anzai, A.; Matsuhashi, T.; Katsumata, Y.; Ito, K.; Yamamoto, T.; Betsuyaku, T.; Shinmura, K.; et al. Lung Natural Killer Cells Play a Major Counter-Regulatory Role in Pulmonary Vascular Hyperpermeability After Myocardial Infarction. Circ. Res. 2014, 114, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Umehara, H.; Bloom, E.; Okazaki, T.; Domae, N.; Imai, T. Fractalkine and vascular injury. Trends Immunol. 2001, 22, 602–607. [Google Scholar] [CrossRef]
- Kahaleh, B. Vascular Disease in Scleroderma: Mechanisms of Vascular Injury. Rheum. Dis. Clin. N. Am. 2008, 34, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Song, Z.; Shen, A.; Chen, T.; Zhang, A. Small molecules targeting the innate immune cGAS‒STING‒TBK1 signaling pathway. Acta Pharm. Sin. B 2020, 10, 2272–2298. [Google Scholar] [CrossRef]
- Zhang, H.; You, Q.-D.; Xu, X.-L. Targeting Stimulator of Interferon Genes (STING): A Medicinal Chemistry Perspective. J. Med. Chem. 2020, 63, 3785–3816. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Z.; Pei, J.; Luo, Q.; Zeng, X.; Li, Q.; Yang, Z.; Quan, J. Identification of α-Mangostin as an Agonist of Human STING. ChemMedChem 2018, 13, 2057–2064. [Google Scholar] [CrossRef]
- Van Herck, S.; Feng, B.; Tang, L. Delivery of STING agonists for adjuvanting subunit vaccines. Adv. Drug Deliv. Rev. 2021, 179, 114020. [Google Scholar] [CrossRef]
- Kamaruddin, N.A.; Abdullah, M.N.H.; Tan, J.J.; Lim, V.; Fong, L.Y.; Ghafar, S.A.A.; Yong, Y.K. Vascular Protective Effect and Its Possible Mechanism of Action on Selected Active Phytocompounds: A Review. Evid. -Based Complement. Altern. Med. 2022, 2022, 1–17. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, H.; Zhang, S.; Bao, L.; Chen, B.; Gong, H.; Zhao, Y.; Qi, R. Resveratrol Confers Vascular Protection by Suppressing TLR4/Syk/NLRP3 Signaling in Oxidized Low-Density Lipoprotein-Activated Platelets. Oxidative Med. Cell. Longev. 2021, 2021, 1–16. [Google Scholar] [CrossRef]
- Parsamanesh, N.; Asghari, A.; Sardari, S.; Tasbandi, A.; Jamialahmadi, T.; Xu, S.; Sahebkar, A. Resveratrol and endothelial function: A literature review. Pharmacol. Res. 2021, 170, 105725. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.-T.; Kuo, C.-H.; Tseng, L.; Li, Y.-S.; Cheng, L.-H.; Cheng, C.-Y.; Sheu, S.-R.; Chang, W.-T.; Chen, C.-C.; Cheng, H.-C. Alpha-Mangostin Reduces Pericellular Fibronectin on Suspended Tumor Cells and Therapeutically, but Not Prophylactically, Suppresses Distant Metastasis. Life 2022, 12, 1375. https://doi.org/10.3390/life12091375
Huang L-T, Kuo C-H, Tseng L, Li Y-S, Cheng L-H, Cheng C-Y, Sheu S-R, Chang W-T, Chen C-C, Cheng H-C. Alpha-Mangostin Reduces Pericellular Fibronectin on Suspended Tumor Cells and Therapeutically, but Not Prophylactically, Suppresses Distant Metastasis. Life. 2022; 12(9):1375. https://doi.org/10.3390/life12091375
Chicago/Turabian StyleHuang, Li-Tzu, Chin-Ho Kuo, Lin Tseng, Yi-Syuan Li, Li-Hsin Cheng, Chin-Yun Cheng, Shane-Rong Sheu, Wen-Tsan Chang, Chien-Chin Chen, and Hung-Chi Cheng. 2022. "Alpha-Mangostin Reduces Pericellular Fibronectin on Suspended Tumor Cells and Therapeutically, but Not Prophylactically, Suppresses Distant Metastasis" Life 12, no. 9: 1375. https://doi.org/10.3390/life12091375