
Label-Free Quantification (LFQ) of Fecal Proteins for Potential Pregnancy Detection in Polar Bears

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Use Statement
2.2. Subjects and Sample Selection
2.3. Sample Preparation
2.4. LFQ Mass Spectrometry Workflow
2.5. Data Analysis
3. Results
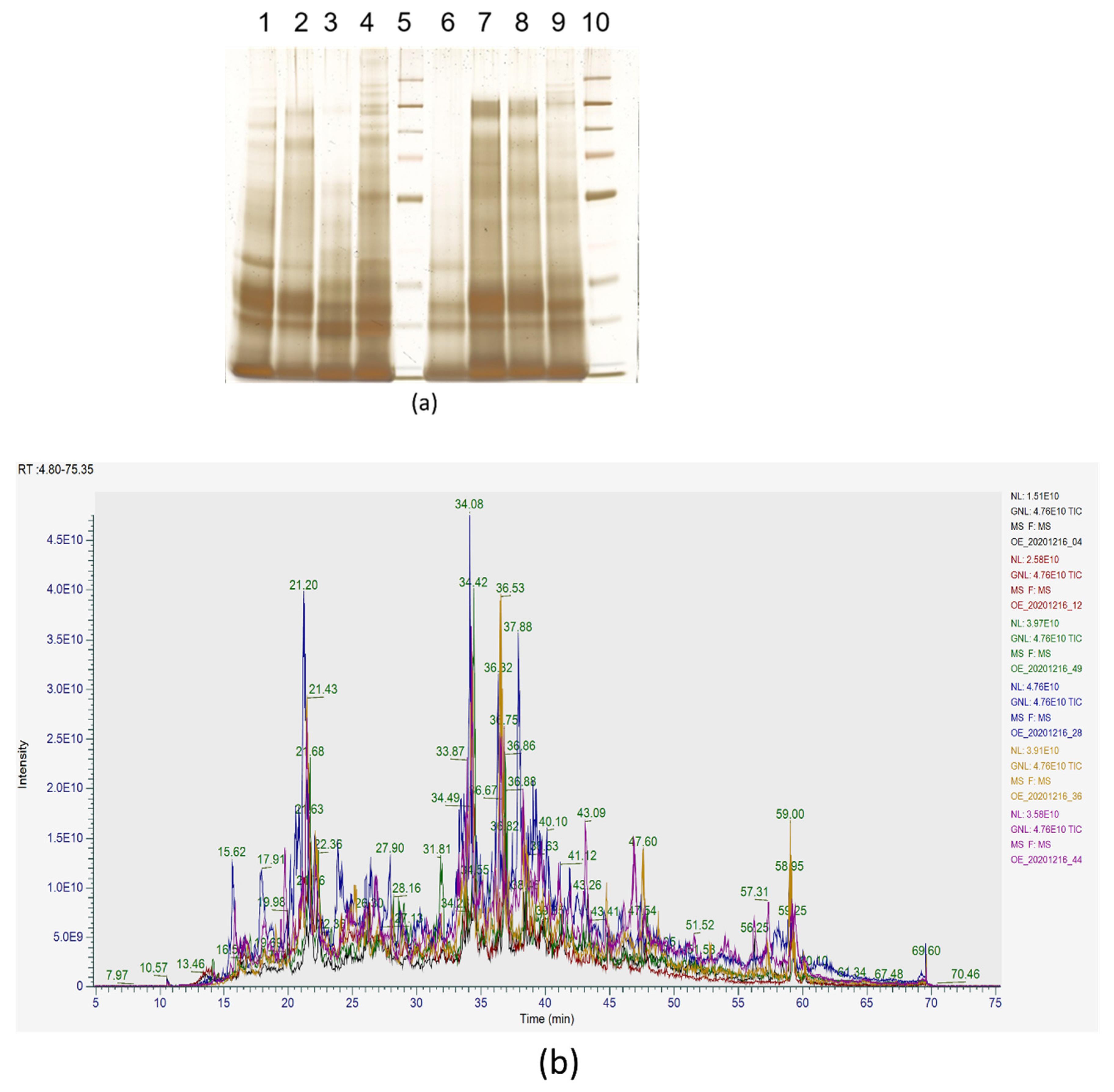
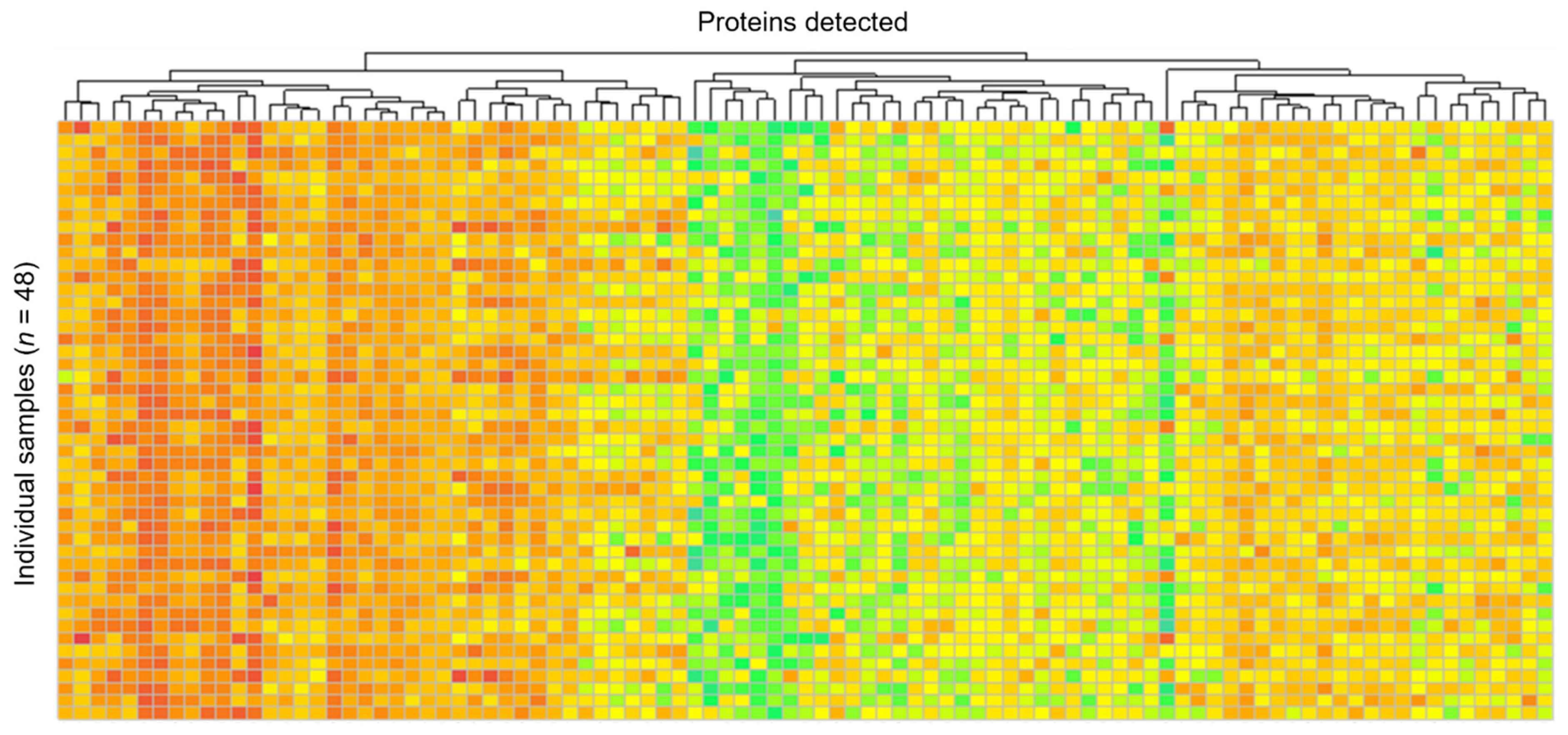
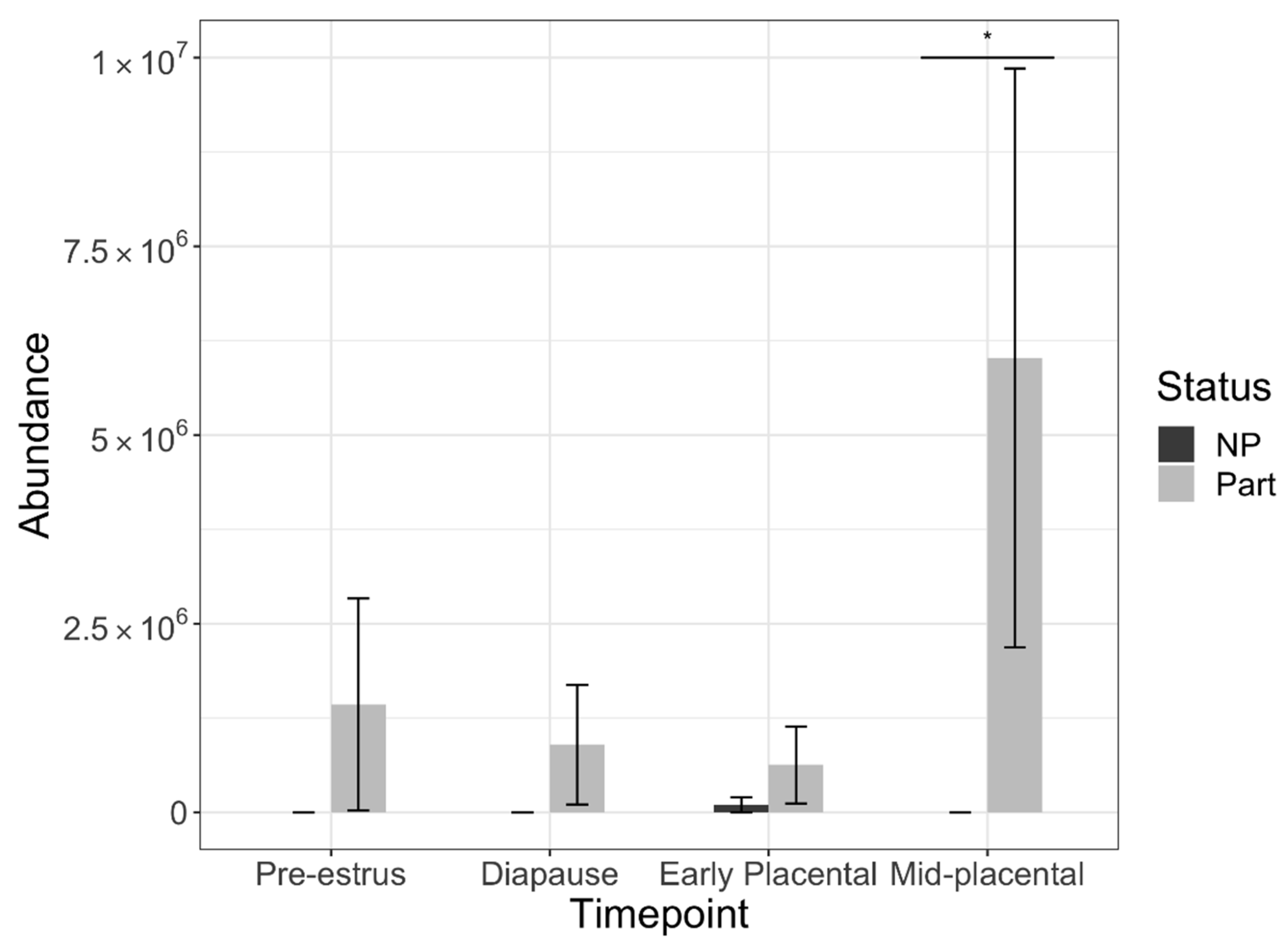
3.1. Protein Extraction and Abundance
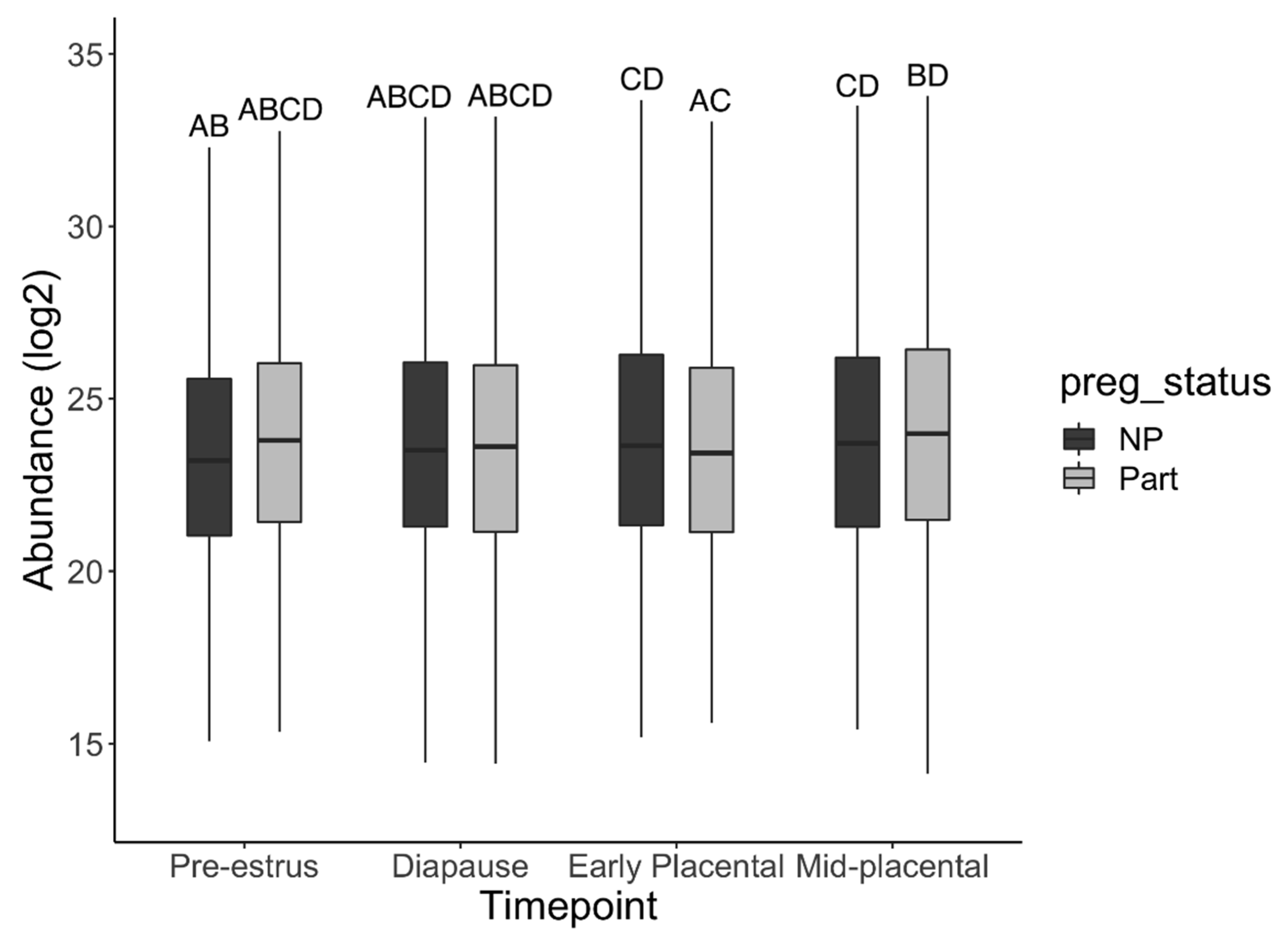
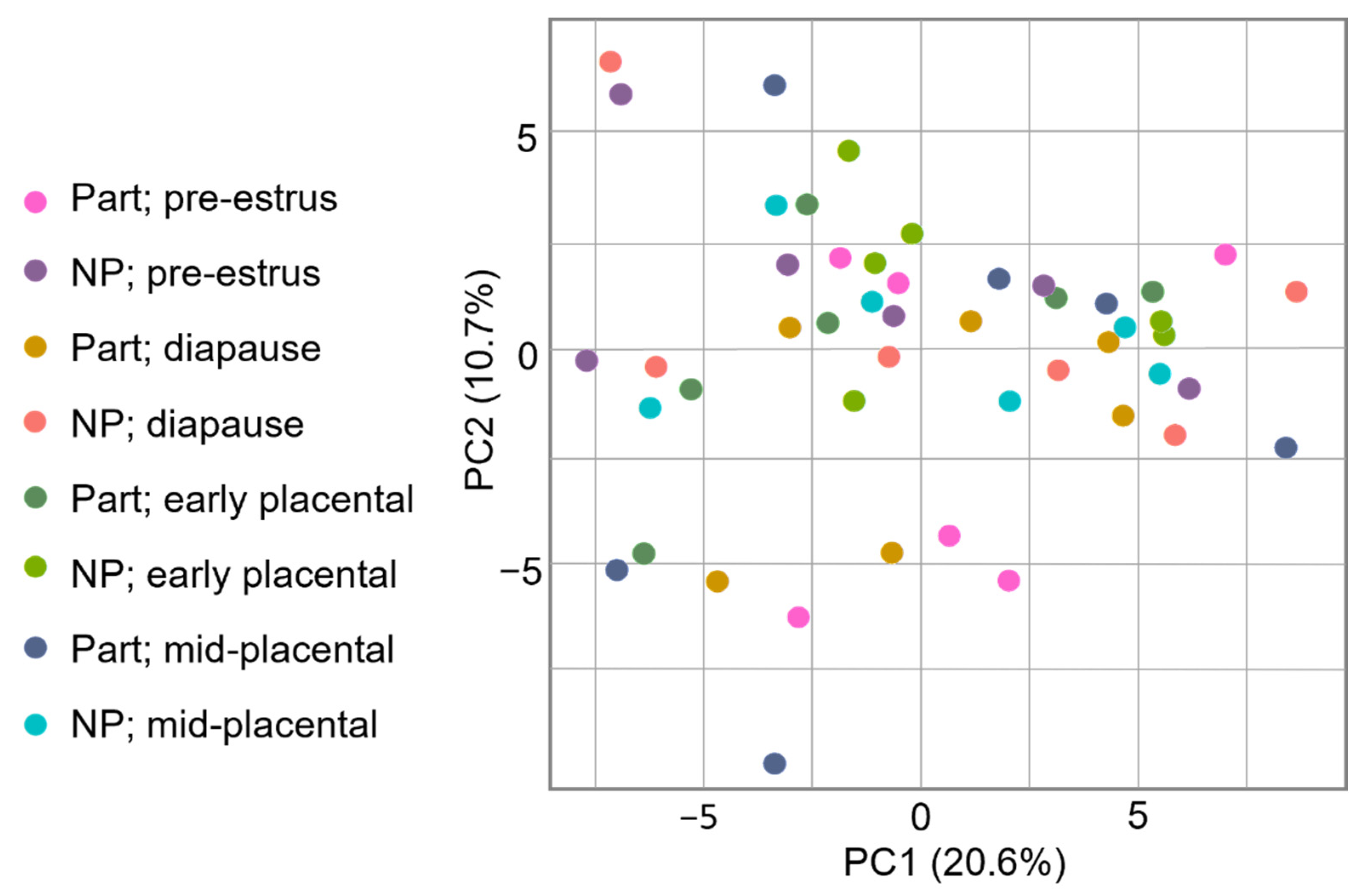
3.2. Protein Identification and Quantification
3.3. Gene Ontology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Radcliffe, R.W. Ultrasonographic Characterization of Ovarian Events and Fetal Gestational Parameters in Two Southern Black Rhinoceros and Correlation to Fecal Progesterone. Theriogenology 2001, 55, 1033–1049. [Google Scholar] [CrossRef]
- Robeck, T.R.; Gili, C.; Doescher, B.M.; Sweeney, J.; De Laender, P.; Van Elk, C.E.; O’Brien, J.K. Altrenogest and Progesterone Therapy during Pregnancy in Bottlenose Dolphins (Tursiops Truncatus) with Progesterone Insufficiency. J. Zoo Wildl. Med. 2012, 43, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Curry, E.; Browning, L.J.; Reinhart, P.; Roth, T.L. Integrating Trans-Abdominal Ultrasonography with Fecal Steroid Metabolite Monitoring to Accurately Diagnose Pregnancy and Predict the Timing of Parturition in the Red Panda (Ailurus Fulgens Styani). Zoo Biol. 2017, 36, 193–200. [Google Scholar] [CrossRef]
- Roth, T.L.; Bateman, H.L.; Kroll, J.L.; Steinetz, B.G.; Reinhart, P.R. Endocrine and Ultrasonographic Characterization of a Successful Pregnancy in a Sumatran Rhinoceros (Dicerorhinus Sumatrensis) Supplemented with a Synthetic Progestin. Zoo Biol. 2004, 23, 219–238. [Google Scholar] [CrossRef]
- Wojtusik, J.; Brandicourt, I.M.C.; Rice, W.; Roth, T.L. Reproductive Cycle and Pregnancy Monitoring in the Common Hippopotamus (Hippopotamus Amphibius) through Salivary Steroid Analyses and Transabdominal Ultrasonography. J. Zoo Aqua. Res. 2020, 8, 181–187. [Google Scholar] [CrossRef]
- Fieß, M.; Heistermann, M.; Hodges, J.K. Patterns of Urinary and Fecal Steroid Excretion during the Ovarian Cycle and Pregnancy in the African Elephant (Loxodonta Africana). Gen. Comp. Endocrinol. 1999, 115, 76–89. [Google Scholar] [CrossRef]
- Schwarzenberger, F. The Many Uses of Non-Invasive Faecal Steroid Monitoring in Zoo and Wildlife Species. Int. Zoo Yearb. 2007, 41, 52–74. [Google Scholar] [CrossRef]
- Carlson, D.A.; Gese, E.M. Relaxin as a Diagnostic Tool for Pregnancy in the Coyote (Canis Latrans). Anim. Reprod. Sci. 2007, 101, 304–312. [Google Scholar] [CrossRef] [Green Version]
- Bauman, J.E.; Clifford, D.L.; Asa, C.S. Pregnancy Diagnosis in Wild Canids Using a Commercially Available Relaxin Assay. Zoo Biol. 2008, 27, 406–413. [Google Scholar] [CrossRef]
- Dehnhard, M.; Kumar, V.; Chandrasekhar, M.; Jewgenow, K.; Umapathy, G. Non-Invasive Pregnancy Diagnosis in Big Cats Using the PGFM (13,14-Dihydro-15-Keto-PGF2α) Assay. PLoS ONE 2015, 10, e0143958. [Google Scholar] [CrossRef] [Green Version]
- Finkenwirth, C.; Jewgenow, K.; Meyer, H.H.D.; Vargas, A.; Dehnhard, M. PGFM (13,14-Dihydro-15-Keto-PGF2α) in Pregnant and Pseudo-Pregnant Iberian Lynx: A New Noninvasive Pregnancy Marker for Felid Species. Theriogenology 2010, 73, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.M.; Brown, J.L.; Kersey, D.C.; Snyder, R.J.; Durrant, B.S.; Kouba, A.J. Use of Urinary 13,14, Dihydro-15-Keto-Prostaglandin F2α (PGFM) Concentrations to Diagnose Pregnancy and Predict Parturition in the Giant Panda (Ailuropoda Melanolecua). PLoS ONE 2018, 13, e0195599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvornicky-Raymond, Z.T.; Donnelly, K.A.; Emerson, J.A.; Penfold, L.M.; Citino, S.B. Evaluation of a Visual Enzyme-Linked Immunosorbent Assay (ELISA) for Pregnancy Detection in Four Ungulate Species. J. Zoo Wildl. Med. 2021, 51, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.M.; Pohler, K.G.; Smith, M.F.; Green, J.A. Placental PAGs: Gene Origins, Expression Patterns, and Use as Markers of Pregnancy. Reproduction 2015, 149, R115–R126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, S.S.; Nelson, R.A.; Ramsay, M.A.; Stirling, I.; Bahr, J.M. Annual Changes in Serum Sex Steroids in Male and Female Black (Ursus Americanus) and Polar (Ursus Maritimus) Bears. Biol. Reprod. 1988, 38, 1044–1050. [Google Scholar] [CrossRef]
- Boone, W.R.; Keck, B.B.; Catlin, J.C.; Casey, K.J.; Boone, E.T.; Dye, P.S.; Schuett, R.J.; Tsubota, T.; Bahr, J.C. Evidence That Bears Are Induced Ovulators. Theriogenology 2004, 61, 1163–1169. [Google Scholar] [CrossRef]
- Van de Velde, F.; Stirling, I.; Richardson, E. Polar Bear (Ursus Maritimus) Denning in the Area of the Simpson Peninsula, Nunavut. Arctic 2003, 56, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Stoops, M.A.; MacKinnon, K.M.; Roth, T.L. Longitudinal Fecal Hormone Analysis for Monitoring Reproductive Activity in the Female Polar Bear (Ursus Maritimus). Theriogenology 2012, 78, 1977–1986. [Google Scholar] [CrossRef]
- Curry, E.; Roth, T.; MacKinnon, K.; Stoops, M. Factors Influencing Annual Fecal Testosterone Metabolite Profiles in Captive Male Polar Bears (Ursus Maritimus). Reprod. Domest. Anim. 2012, 47, 222–225. [Google Scholar] [CrossRef]
- Meyerson, R.; Long, S. Polar Bear (Ursus Maritimus) Population Analysis & Breeding and Transfer Plan; Association of Zoos and Aquariums Population Management Center: Chicago, IL, USA, 2019. [Google Scholar]
- Curry, E.; Safayi, S.; Meyerson, R.; Roth, T.L. Reproductive Trends of Captive Polar Bears in North American Zoos: A Historical Analysis. J. Zoo Aquar. Res. 2015, 3, 99–106. [Google Scholar]
- Andrews, C.J.; Thomas, D.G.; Welch, M.V.; Yapura, J.; Potter, M.A. Monitoring Ovarian Function and Detecting Pregnancy in Felids: A Review. Theriogenology 2020, 157, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenberger, F.; Rietschel, W.; Vahala, J.; Holeckova, D.; Thomas, P.; Maltzan, J.; Baumgartner, K.; Schaftenaar, W. Fecal Progesterone, Estrogen, and Androgen Metabolites for Noninvasive Monitoring of Reproductive Function in the Female Indian Rhinoceros, Rhinoceros Unicornis. Gen. Comp. Endocrinol. 2000, 119, 300–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzenberger, F.; Fredriksson, G.; Schaller, K.; Kolter, L. Fecal Steroid Analysis for Monitoring Reproduction in the Sun Bear (Helarctos Malayanus). Theriogenology 2004, 62, 1677–1692. [Google Scholar] [CrossRef] [PubMed]
- Dehnhard, M.; Jewgenow, K. Measurement of Faecal Prostaglandins in Felids and Three Ursid Species. Wien Tierarztl 2013, 100, 227–237. [Google Scholar]
- Schulz, L.; Nelson, R.A.; Pyter, L.M.; Bahr, J.M. Induction of Pseudopregnancy in the American Black Bear (Ursus Americanus). J. Exp. Zool. 2003, 298A, 162–166. [Google Scholar] [CrossRef]
- Mead, R.A.; Bowles, M.; Starypan, G.; Jones, M. Evidence for Pseudopregnancy and Induced Ovulation in Captive Wolverines (Gulo Gulo). Zoo Biol. 1993, 12, 353–358. [Google Scholar] [CrossRef]
- Curry, E.; Stoops, M.A.; Roth, T.L. Non-Invasive Detection of Candidate Pregnancy Protein Biomarkers in the Feces of Captive Polar Bears (Ursus Maritimus). Theriogenology 2012, 78, 308–314. [Google Scholar] [CrossRef]
- Koester, D.C.; Wildt, D.E.; Maly, M.; Comizzoli, P.; Crosier, A.E. Non-Invasive Identification of Protein Biomarkers for Early Pregnancy Diagnosis in the Cheetah (Acinonyx Jubatus). PLoS ONE 2017, 12, e0188575. [Google Scholar] [CrossRef] [Green Version]
- Byron, M.J.; Koester, D.C.; Edwards, K.L.; Mozdziak, P.E.; Farin, C.E.; Crosier, A.E. Immunoglobulin J Chain as a Non-Invasive Indicator of Pregnancy in the Cheetah (Acinonyx Jubatus). PLoS ONE 2020, 15, e0225354. [Google Scholar] [CrossRef] [Green Version]
- Curry, E.; Easley, J.; Wojtusik, J.; Roth, T. Identification of Mink (Neovison Vison) Fecal Proteins during Embryonic Diapause and Placental Pregnancy for Non-Invasive Pregnancy Diagnosis in Wildlife. In Proceedings of the III International Symposium on Embryonic Diapause, Ascona, Switzerland, 2–6 June 2020. [Google Scholar] [CrossRef]
- Kennedy, V.; Roth, T.L.; Donelan, E.; Curry, E. Western Blot Evaluation of Fecal Transthyretin (TTR) for Use as a Pregnancy Test in Polar Bears (Ursus Maritimus). In Proceedings of the 51st Annual Conference of the Society for the Study of Reproduction, New Orleans, LA, USA, 10–13 July 2018. [Google Scholar]
- Wu, W.W.; Wang, G.; Baek, S.J.; Shen, R.-F. Comparative Study of Three Proteomic Quantitative Methods, DIGE, CICAT, and ITRAQ, Using 2D Gel- or LC−MALDI TOF/TOF. J. Proteome. Res. 2006, 5, 651–658. [Google Scholar] [CrossRef]
- Giard, D.J. Routine Heat Inactivation of Serum Reduces Its Capacity to Promote Cell Attachment. In Vitro Cell Dev. Biol. 1987, 23, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Hollaender, A. Effects of Ultraviolet Radiation. Annu. Rev. Physiol. 1946, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Guerra-Moreno, A.; Ang, J.; Micoogullari, Y. Protein Degradation and the Pathologic Basis of Disease. Am. J. Pathol. 2019, 189, 94–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hock, B.; Rahman, M.; Rauchalles, S.; Dankwardt, A.; Seifert, M.; Haindl, S.; Kramer, K. Stabilisation of Immunoassays and Receptor Assays. J. Mol. Catal. B Enzym. 1999, 7, 115–124. [Google Scholar] [CrossRef]
- Zhao, L.; Cong, X.; Zhai, L.; Hu, H.; Xu, J.-Y.; Zhao, W.; Zhu, M.; Tan, M.; Ye, B.-C. Comparative Evaluation of Label-Free Quantification Strategies. J. Proteom. 2020, 215, 103669. [Google Scholar] [CrossRef]
- Chauvin, A.; Boisvert, F.-M. Clinical Proteomics in Colorectal Cancer, a Promising Tool for Improving Personalised Medicine. Proteomes 2018, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Geyer, P.E.; Kulak, N.A.; Pichler, G.; Holdt, L.M.; Teupser, D.; Mann, M. Plasma Proteome Profiling to Assess Human Health and Disease. Cell Syst. 2016, 2, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Sandow, J.J.; Rainczuk, A.; Infusini, G.; Makanji, M.; Bilandzic, M.; Wilson, A.L.; Fairweather, N.; Stanton, P.G.; Garama, D.; Gough, D.; et al. Discovery and Validation of Novel Protein Biomarkers in Ovarian Cancer Patient Urine. Prot. Clin. Appl. 2018, 12, 1700135. [Google Scholar] [CrossRef]
- Yu, Y.; Suh, M.-J.; Sikorski, P.; Kwon, K.; Nelson, K.E.; Pieper, R. Urine Sample Preparation in 96-Well Filter Plates for Quantitative Clinical Proteomics. Anal. Chem. 2014, 86, 5470–5477. [Google Scholar] [CrossRef]
- Titz, B.; Gadaleta, R.; Lo Sasso, G.; Elamin, A.; Ekroos, K.; Ivanov, N.; Peitsch, M.; Hoeng, J. Proteomics and Lipidomics in Inflammatory Bowel Disease Research: From Mechanistic Insights to Biomarker Identification. Int. J. Mol. Sci. 2018, 19, 2775. [Google Scholar] [CrossRef] [Green Version]
- Rode, K.D.; Robbins, C.T.; Stricker, C.A.; Taras, B.D.; Tollefson, T.N. Energetic and Health Effects of Protein Overconsumption Constrain Dietary Adaptation in an Apex Predator. Sci. Rep. 2021, 11, 15309. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.T.; Tollefson, T.N.; Rode, K.D.; Erlenbach, J.A.; Ardente, A.J. New Insights into Dietary Management of Polar Bears (Ursus Maritimus) and Brown Bears (U. Arctos). Zoo Biol. 2022, 41, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Eismann, T.; Huber, N.; Shin, T.; Kuboki, S.; Galloway, E.; Wyder, M.; Edwards, M.J.; Greis, K.D.; Shertzer, H.G.; Fisher, A.B.; et al. Peroxiredoxin-6 Protects against Mitochondrial Dysfunction and Liver Injury during Ischemia-Reperfusion in Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G266–G274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2015, arXiv:1406.5823. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Pack. Version 2021, 1, 3. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Use R! Springer International Publishing: Houston, TX, USA, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, 13. [Google Scholar] [CrossRef]
- The UniProt Consortium; Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; et al. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Valdés, A.; Holst, B.S.; Lindersson, S.; Ramström, M. Development of MS-Based Methods for Identification and Quantification of Proteins Altered during Early Pregnancy in Dogs. J. Proteom. 2019, 192, 223–232. [Google Scholar] [CrossRef]
- Heck, M.; Neely, B.A. Proteomics in Non-Model Organisms: A New Analytical Frontier. J. Proteome Res. 2020, 19, 3595–3606. [Google Scholar] [CrossRef]
- Goldstein, B.; King, N. The Future of Cell Biology: Emerging Model Organisms. Trends Cell Biol. 2016, 26, 818–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callier, V. Solving Peto’s Paradox to Better Understand Cancer. Proc. Natl. Acad. Sci. USA. 2019, 116, 1825–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, J.M.; Sulak, M.; Chigurupati, S.; Lynch, V.J. A Zombie LIF Gene in Elephants Is Upregulated by TP53 to Induce Apoptosis in Response to DNA Damage. Cell Rep. 2018, 24, 1765–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of Cancer Resistance in Long-Lived Mammals. Nat. Rev. Cancer 2018, 18, 433–441. [Google Scholar] [CrossRef]
- Grabek, K.R.; Cooke, T.F.; Epperson, L.E.; Spees, K.K.; Cabral, G.F.; Sutton, S.C.; Merriman, D.K.; Martin, S.L.; Bustamante, C.D. Genetic Variation Drives Seasonal Onset of Hibernation in the 13-Lined Ground Squirrel. Commun. Biol. 2019, 2, 478. [Google Scholar] [CrossRef] [Green Version]
- Jansen, H.T.; Leise, T.; Stenhouse, G.; Pigeon, K.; Kasworm, W.; Teisberg, J.; Radandt, T.; Dallmann, R.; Brown, S.; Robbins, C.T. The Bear Circadian Clock Doesn’t ‘Sleep’ during Winter Dormancy. Front. Zool. 2016, 13, 42. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Haines, S.; Coates, D. Proteomic Profiling of Stem Cell Tissues during Regeneration of Deer Antler: A Model of Mammalian Organ Regeneration. J. Proteome Res. 2020, 19, 1760–1775. [Google Scholar] [CrossRef]
- Li, C.; Zhao, H.; Liu, Z.; McMahon, C. Deer Antler—A Novel Model for Studying Organ Regeneration in Mammals. Int. J. Biochem. Cell Biol. 2014, 56, 111–122. [Google Scholar] [CrossRef]
- Wetterstrand, K. DNA Sequencing Costs: Data from the NHGRI Genome Sequencing Program (GSP); NIH: Bethesda, MD, USA, 2021. [Google Scholar]
- Pongracz, J.D.; Paetkau, D.; Branigan, M.; Richardson, E. Recent Hybridization between a Polar Bear and Grizzly Bears in the Canadian Arctic. Arctic 2017, 70, 151. [Google Scholar] [CrossRef]
- Wilson, A.E.; Michaud, S.A.; Jackson, A.M.; Stenhouse, G.; McClelland, C.J.R.; Coops, N.C.; Janz, D.M. Protein Biomarkers in Serum as a Conservation Tool to Assess Reproduction: A Case Study on Brown Bears (Ursus Arctos). Conserv. Physiol. 2021, 9, coab091. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProtID | Protein | Mean Raw Abundance |
|---|---|---|
| A0A452SML2 | Carboxypeptidase A1 | 14,329,861,669 |
| A0A452SXJ4 | Uncharacterized protein | 13,577,232,635 |
| A0A452UQI9 | Ig-like domain-containing protein | 6,370,583,108 |
| A0A452UQ85 | IGv domain-containing protein | 6,192,069,758 |
| A0A3Q7UBB8 | IgGFc-binding protein | 5,536,351,190 |
| A0A3Q7W0E2 | Titin isoform X2 | 4,528,398,405 |
| A0A3Q7VT65 | Alkaline phosphatase | 2,394,052,435 |
| A0A384BSZ9 | Dipeptidyl peptidase 4 | 2,189,880,724 |
| A0A452U793 | Peptidase S1 domain-containing protein | 2,139,152,673 |
| A0A452QAI0 | Ig-like domain-containing protein | 1,425,672,788 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curry, E.; Philpott, M.E.; Wojtusik, J.; Haffey, W.D.; Wyder, M.A.; Greis, K.D.; Roth, T.L. Label-Free Quantification (LFQ) of Fecal Proteins for Potential Pregnancy Detection in Polar Bears. Life 2022, 12, 796. https://doi.org/10.3390/life12060796
Curry E, Philpott ME, Wojtusik J, Haffey WD, Wyder MA, Greis KD, Roth TL. Label-Free Quantification (LFQ) of Fecal Proteins for Potential Pregnancy Detection in Polar Bears. Life. 2022; 12(6):796. https://doi.org/10.3390/life12060796
Chicago/Turabian StyleCurry, Erin, Megan E. Philpott, Jessye Wojtusik, Wendy D. Haffey, Michael A. Wyder, Kenneth D. Greis, and Terri L. Roth. 2022. "Label-Free Quantification (LFQ) of Fecal Proteins for Potential Pregnancy Detection in Polar Bears" Life 12, no. 6: 796. https://doi.org/10.3390/life12060796