
The Interplay between Pathophysiological Pathways in Early-Onset Severe Preeclampsia Unveiled by Metabolomics

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Baseline and Perinatal Characteristics
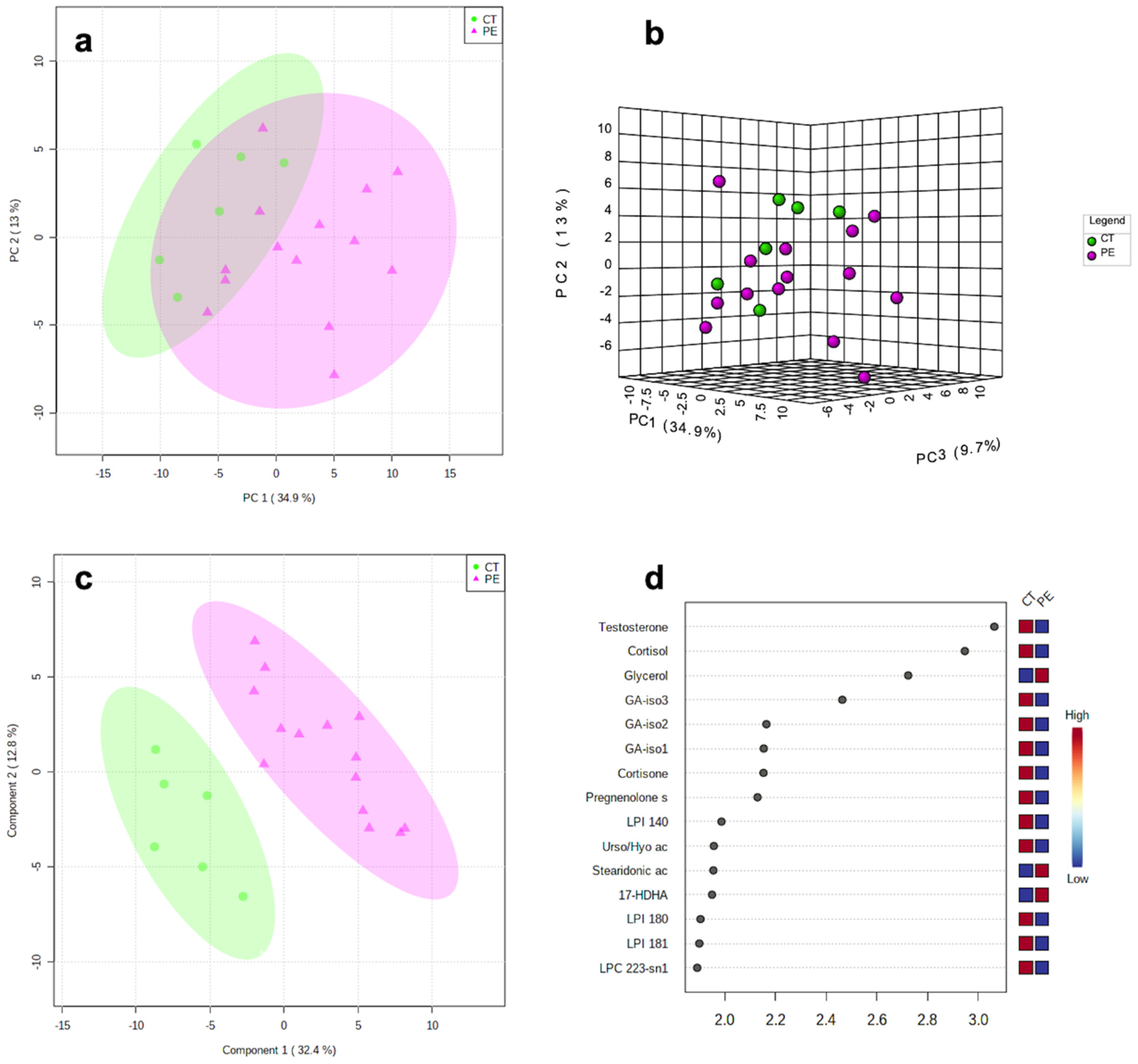
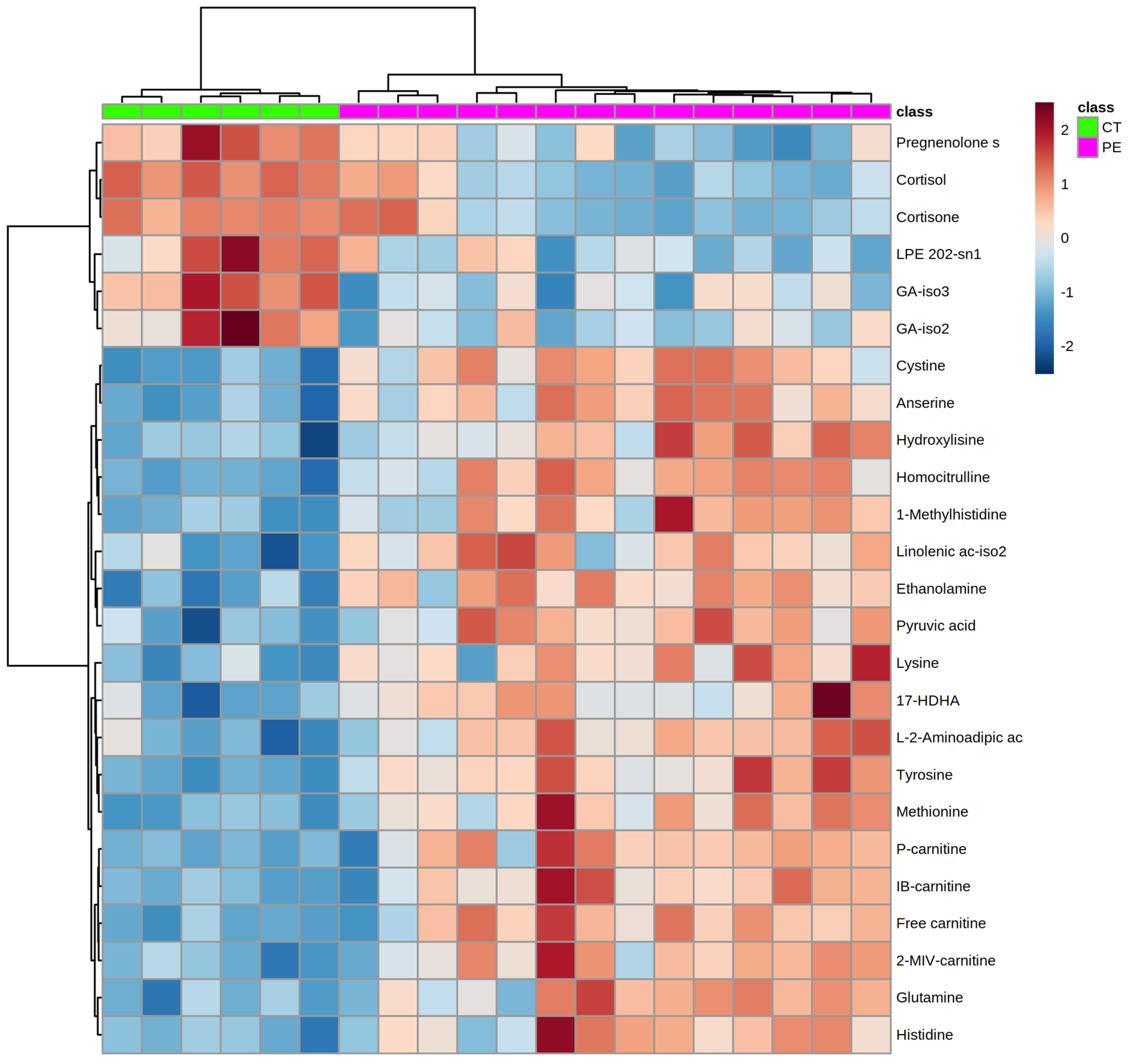
2.2. Metabolomics Results
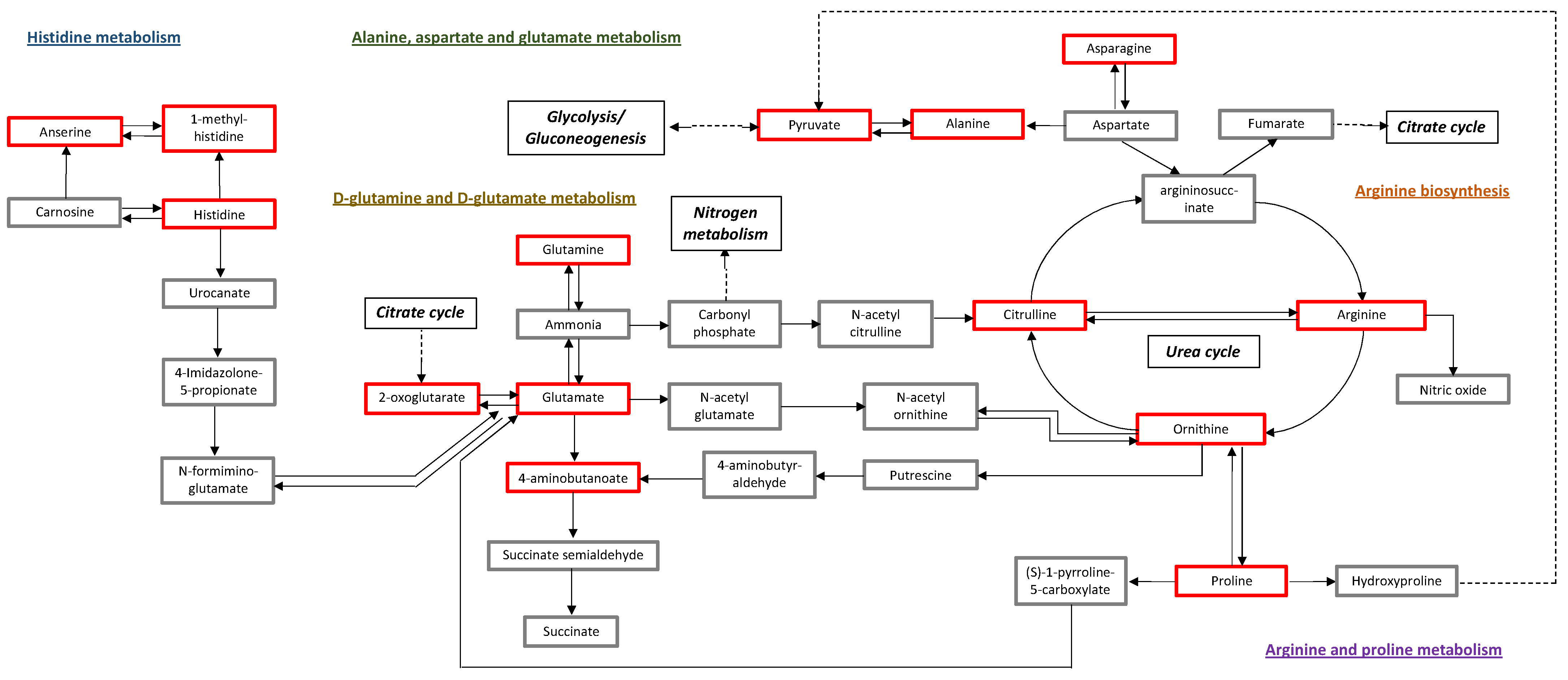
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Data Collection
4.3. Maternal Blood Sampling
4.4. Metabolomic Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Methods
Appendix A.1.1. Metabolomics Technique
Method 1: Lipidomic Analysis with Methanol Extraction

{kind=link}
{kind=link}
{kind=link}
| Standards | |
|---|---|
| 15-HETE | lysoPC 18:1 |
| arachidonic acid | lysoPE18:1 |
| Cholic acid | lysoPI 18:1 |
| Deoxycholic acid | sphingosine-1-P |
| Dehydroisoandosterone-3-sulfate | Taurocholic acid |
| Hydrocortisone (cortisol) | Carnitine Mix |
Method 2: Lipidomic Analysis with Choloform:Methanol (Folch Method) Extraction
| Standards |
|---|
| Sphingomyelin d18:1/16:0 |
| Stearoyl-rac-glycerol |
| Oelamide |
| Dilynoleyl-rac-glycerol |
| Palmitoyl-oleyl-lynoleyl-rac-glycerol |
| Dipalmitoyl-phosphoethanolamine |
| Dioleyl-phosphocholine |
| Cholesteryl linoleate |
Method 3: Aminoacids Analysis
Method 4: Polar Metabolites and Central Carbon Metabolism
| Compound | |
|---|---|
| Piruvic acid | Aconitic acid |
| Lactic acid | 3-Phosphoglyceric acid |
| Glycolic acid | Citric/isocitric acid |
| 2-Hydroxybutyric acid | Myristic acid |
| Oxalic Acid | Glucose |
| 3-Hydroxybutiric acid | Palmitic acid |
| Glycerol | Ribose-5-phosphate |
| Succinic acid | Linoleic acid |
| Glyceric acid | Oleic Acid |
| Fumaric acid | Stearic acid |
| Malic acid | Fructose-6-phosphate |
| Threonic acid | Glucose-6-phosphate |
| 2-hydroxyglutaric acid | Fructose-1,6-bisphosphate |
| alpha-Ketoglutaric acid | 6-Phosphogluconic acid |
| Phosphoenolpyruvate | Sucrose |
| Ornithine | alpha-Tocopherol |
| Glyceraldehyde-3-phosphate | Cholesterol |
Appendix A.2. Statistical Analysis
References
- Mol, B.W.J.; Roberts, C.T.; Thangaratinam, S.; Magee, L.A.; De Groot, C.J.M.; Hofmeyr, G.J. Pre-eclampsia. Lancet 2016, 387, 999–1011. [Google Scholar] [CrossRef]
- Duley, L. The Global Impact of Pre-eclampsia and Eclampsia. Semin. Perinatol. 2009, 33, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Ghulmiyyah, L.; Sibai, B. Maternal Mortality From Preeclampsia/Eclampsia. Semin. Perinatol. 2012, 36, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L.; Staff, A.C. IFPA senior award lecture: Making sense of pre-eclampsia—Two placental causes of preeclampsia? Placenta 2014, 35, S20–S25. [Google Scholar] [CrossRef] [PubMed]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myatt, L.; Roberts, J.M. Preeclampsia: Syndrome or Disease? Curr. Hypertens. Rep. 2015, 17, 83. [Google Scholar] [CrossRef] [PubMed]
- Von Dadelszen, P.; Magee, L.A.; Roberts, J.M. Subclassification of Preeclampsia. Hypertens. Pregnancy 2003, 22, 143–148. [Google Scholar] [CrossRef]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [Green Version]
- Kenny, L.C.; Broadhurst, D.; Brown, M.; Dunn, W.B.; Redman, C.W.G.; Kell, D.B.; Baker, P.N. Detection and identification of novel metabolomic biomarkers in preeclampsia. Reprod. Sci. (Thousand Oaks Calif.) 2008, 15, 591–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austdal, M.; Skrastad, R.B.; Gundersen, A.S.; Austgulen, R.; Iversen, A.C.; Bathen, T.F. Metabolomic biomarkers in serum and urine in women with preeclampsia. PLoS ONE 2014, 9, e91923. [Google Scholar] [CrossRef] [Green Version]
- Turner, E.; Brewster, J.A.; Simpson, N.A.B.; Walker, J.J.; Fisher, J. Plasma from women with preeclampsia has a low lipid and ketone body content—A nuclear magnetic resonance study. Hypertens. Pregnancy 2007, 26, 329–342. [Google Scholar] [CrossRef]
- Turner, E.; Brewster, J.A.; Simpson, N.A.; Walker, J.J.; Fisher, J. Aromatic amino acid biomarkers of preeclampsia—A nuclear magnetic resonance investigation. Hypertens. Pregnancy 2008, 27, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.; Ray, C.D.; Ray, S.; Dasgupta, S.; Chaudhury, K. Altered metabolic profile in early and late onset preeclampsia: An FTIR spectroscopic study. Pregnancy Hypertens. 2014, 4, 70–80. [Google Scholar] [CrossRef]
- Schott, S.; Hahn, J.; Kurbacher, C.; Moka, D. 31P and 1H nuclear magnetic resonance spectroscopy of blood plasma in female patients with preeclampsia. Int. J. Biomed. Sci. 2012, 8, 258–263. [Google Scholar] [PubMed]
- Liu, Y.; Zu, L.; Cai, W.; Cheng, Q.; Hua, T.; Peng, L.; Li, G.; Zhang, X. Metabolomics revealed decreased level of omega-3 PUFA-derived protective eicosanoids in pregnant women with pre-eclampsia. Clin. Exp. Pharmacol. Physiol. 2019, 46, 705–710. [Google Scholar] [CrossRef]
- Youssef, L.; Simões, R.V.; Miranda, J.; Luisa, M.; Martín, G.; Paules, C.; Crovetto, F.; Amigó, N.; Cañellas, N.; Gratacos, E. Paired maternal and fetal metabolomics reveal a differential fingerprint in preeclampsia versus fetal growth restriction. Sci. Rep. 2021, 11, 14422. [Google Scholar] [CrossRef] [PubMed]
- Baylis, C.; Beinder, E.; Sütö, T.; August, P. Recent insights into the roles of nitric oxide and reninangiotensin in the pathophysiology of preeclamptic pregnancy. Semin. Nephrol. 1998, 18, 208–230. [Google Scholar]
- Evans, R.W.; Powers, R.W.; Ness, R.B.; Cropcho, L.J.; Daftary, A.R.; Harger, G.F.; Vergona, R.; Finegold, D.N. Maternal and fetal amino acid concentrations and fetal outcomes during pre-eclampsia. Reproduction 2003, 125, 785–790. [Google Scholar] [CrossRef]
- Noris, M.; Todeschini, M.; Cassis, P.; Pasta, F.; Cappellini, A.; Bonazzola, S.; Macconi, D.; Maucci, R.; Porrati, F.; Benigni, A.; et al. L-Arginine Depletion in Preeclampsia Orients Nitric Oxide Synthase Toward Oxidant Species. Hypertension 2004, 43, 614–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorniak-Wall, T.; Grivell, R.M.; Dekker, G.A.; Hague, W.; Dodd, J.M. The role of L-arginine in the prevention and treatment of pre-eclampsia: A systematic review of randomised trials. J. Hum. Hypertens. 2014, 28, 230–235. [Google Scholar] [CrossRef]
- Cheng, S.; Rhee, E.P.; Larson, M.G.; Lewis, G.D.; McCabe, E.L.; Shen, D.; Palma, M.J.; Roberts, L.D.; Dejam, A.; Souza, A.L.; et al. Metabolite profiling identifies pathways associated with metabolic risk in humans. Circulation 2012, 125, 2222–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, A.; Stanely, C.A. Disorders of glutamate metabolism. Ment. Retard. Dev. Disabil. 2001, 295, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.E.; Brosnan, J.T. Hepatic glutamate metabolism: A tale of 2 hepatocytes. Am. J. Clin. Nutr. 2009, 90, 857–861. [Google Scholar] [CrossRef] [Green Version]
- Pouta, A.; Hartikainen, A.L.; Sovio, U.; Gissler, M.; Laitinen, J.; McCarthy, M.I.; Ruokonen, A.; Elliott, P.; Järvelin, M.R. Manifestations of Metabolic Syndrome after Hypertensive Pregnancy. Hypertension 2004, 43, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Prokopieva, V.D.; Yarygina, E.G.; Bokhan, N.A.; Ivanova, S.A. Use of Carnosine for Oxidative Stress Reduction in Different Pathologies. Oxid. Med. Cell. Longev. 2016, 2016, 2939087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, L.; Erlandsson, L.; Åkerström, B.; Miranda, J.; Paules, C.; Crovetto, F.; Crispi, F.; Gratacos, E.; Hansson, S.R. Hemopexin and α1-microglobulin heme scavengers with differential involvement in preeclampsia and fetal growth restriction. PLoS ONE 2020, 15, e0239030. [Google Scholar] [CrossRef]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef]
- Feig, D.S.; Shah, B.R.; Lipscombe, L.L.; Wu, C.F.; Ray, J.G.; Lowe, J.; Hwee, J.; Booth, G.L. Preeclampsia as a Risk Factor for Diabetes: A Population-Based Cohort Study. PLoS Med. 2013, 10, e1001425. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.L.; DerSimonian, R.; Raymond, E.; Morrow, J.D.; Roberts, L.J.; Clemens, J.D.; Hauth, J.C.; Catalano, P.; Sibai, B.; Curet, L.B.; et al. Prostacyclin and thromboxane changes predating clinical onset of preeclampsia: A multicenter prospective study. J. Am. Med. Assoc. 1999, 282, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Horvath, A.; Koletzko, B.; Szajewska, H. Effect of supplementation of women in high-risk pregnancies with long-chain polyunsaturated fatty acids on pregnancy outcomes and growth measures at birth: A meta-analysis of randomized controlled trials. Br. J. Nutr. 2007, 98, 253–259. [Google Scholar] [CrossRef]
- Rolnik, D.L.; Wright, D.; Poon, L.C.; O’Gorman, N.; Syngelaki, A.; de Paco Matallana, C.; Akolekar, R.; Cicero, S.; Janga, D.; Singh, M.; et al. Aspirin versus Placebo in Pregnancies at High Risk for Preterm Preeclampsia. N. Engl. J. Med. 2017, 377, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Serrano, N.C.; Guio-Mahecha, E.; Quintero-Lesmes, D.C.; Becerra-Bayona, S.; Paez, M.C.; Beltran, M.; Herrera, V.M.; Leon, L.J.; Williams, D.; Casas, J.P. Lipid profile, plasma apolipoproteins, and pre-eclampsia risk in the GenPE case-control study. Atherosclerosis 2018, 276, 189–194. [Google Scholar] [CrossRef]
- Arifin, R.; Kyi, W.M.; Che Yaakob, C.A.; Yaacob, N.M. Increased circulating oxidised low-density lipoprotein and antibodies to oxidised low-density lipoprotein in preeclampsia. J. Obstet. Gynaecol. (Lahore) 2017, 37, 580–584. [Google Scholar] [CrossRef]
- Youssef, L.; Miranda, J.; Blasco, M.; Paules, C.; Crovetto, F.; Palomo, M.; Torramade-Moix, S.; García-Calderó, H.; Tura-Ceide, O.; Dantas, A.P.; et al. Complement and coagulation cascades activation is the main pathophysiological pathway in early-onset severe preeclampsia revealed by maternal proteomics. Sci. Rep. 2021, 11, 3048. [Google Scholar] [CrossRef]
- Hertig, A.; Liere, P.; Chabbert-Buffet, N.; Fort, J.; Pianos, A.; Eychenne, B.; Cambourg, A.; Schumacher, M.; Berkane, N.; Lefevre, G.; et al. Steroid profiling in preeclamptic women: Evidence for aromatase deficiency. Am. J. Obstet. Gynecol. 2010, 203, 477.e1–477.e9. [Google Scholar] [CrossRef] [PubMed]
- Kosicka, K.; Siemiątkowska, A.; Szpera-Goździewicz, A.; Krzyścin, M.; Bręborowicz, G.H.; Główka, F.K. Increased cortisol metabolism in women with pregnancy-related hypertension. Endocrine 2018, 61, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Quinkler, M.; Stewart, P.M. Hypertension and the cortisol-cortisone shuttle. J. Clin. Endocrinol. Metab. 2003, 88, 2384–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steegers, E.A.P.; Von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Roberts, J.M.; Druzin, M.; August, P.A.; Gaiser, R.R.; Bakris, G.; Granger, J.P.; Barton, J.R.; Jeyabalan, A.; Bernstein, I.A.; Johnson, D.D.; et al. ACOG Guidelines: Hypertension in Pregnancy; ACOG: Washington, DC, USA, 2012; ISBN 9781934984284. [Google Scholar]
- Figueras, F.; Meler, E.; Iraola, A.; Eixarch, E.; Coll, O.; Figueras, J.; Francis, A.; Gratacos, E.; Gardosi, J. Customized birthweight standards for a Spanish population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 136, 20–24. [Google Scholar] [CrossRef]
- Robinson, H.; Fleming, J. A critical evaluation of sonar crown-rump length measurements. Br. J. Obstet. Gynaecol. 1975, 82, 702–710. [Google Scholar] [CrossRef]
- Hadlock, F.P.; Harrist, R.B.; Sharman, R.S.; Deter, R.L.; Park, S.K. Estimation of fetal weight with the use of head, body, and femur measurements—A prospective study. Am. J. Obs. Gynecol. 1985, 151, 333–337. [Google Scholar] [CrossRef]
- Bhide, A.; Acharya, G.; Bilardo, C.M.; Brezinka, C.; Cafici, D.; Hernandez-Andrade, E.; Kalache, K.; Kingdom, J.; Kiserud, T.; Lee, W.; et al. ISUOG practice guidelines: Use of Doppler ultrasonography in obstetrics. Ultrasound Obstet. Gynecol. 2013, 41, 233–239. [Google Scholar] [CrossRef]
- Gómez, O.; Figueras, F.; Fernández, S.; Bennasar, M.; Martínez, J.M.; Puerto, B.; Gratacós, E. Reference ranges for uterine artery mean pulsatility index at 11–41 weeks of gestation. Ultrasound Obstet. Gynecol. 2008, 32, 128–132. [Google Scholar] [CrossRef]
- Arduini, D.; Rizzo, G. Normal values of pulsatility index from fetal vessels: A cross-sectional study on 1556 healthy fetuses. J. Perinat. Med. 1990, 18, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Hecher, K.; Campbell, S.; Snijders, R.; Nicolaides, K. Reference ranges for fetal venous and atrioventricular blood flow parameters. Ultrasound Obstet. Gynecol. 1994, 4, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Baschat, A.A.; Gembruch, U. The cerebroplacental Doppler ratio revisited. Ultrasound Obstet. Gynecol. 2003, 21, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Comprehensive gene and pathway analysis of cervical cancer progression. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]



| Controls n = 6 | Preeclampsia n = 14 | p-Value | |
|---|---|---|---|
| Maternal characteristics | |||
| Age (years) | 36.5 ± 2.9 | 34.1 ± 3.8 | 0.28 * |
| Caucasian ethnicity | 5 (83.3) | 6 (42.9) | 0.10 ƒ |
| Pre-gestational BMI (kg/m2) | 22.2 ± 2.7 | 24.1 ± 4.1 | 0.38 * |
| Nulliparity | 5 (83.3) | 9 (64.3) | 0.39 ƒ |
| Previous preeclampsia | 0 (0) | 1 (7.1) | 0.50 ƒ |
| Chronic hypertension | 0 (0) | 1 (7.1) | 0.50 ƒ |
| Assisted reproductive technologies | 0 (0) | 3 (21.4) | 0.22 ƒ |
| Smoking during pregnancy | 1 (16.7) | 1 (7.1) | 0.52 ƒ |
| Fetal ultrasound assessment | |||
| Gestational age at ultrasound (weeks) | 26.6 ± 3.7 | 30.5 ± 2.6 | 0.013 * |
| Estimated fetal weight (g) | 994 ± 496 | 1187 ± 328 | 0.30 * |
| Estimated fetal weight centile | 34 (29–66) | 1 (0–3) | <0.001 ω |
| Uterine arteries mean PI (z score) | −0.53 ± 1.77 | 2.60 ± 2.08 | 0.002 * |
| Umbilical artery PI (z score) | −0.40 ± 0.32 | 1.27 ± 1.71 | 0.07 * |
| Middle cerebral artery PI (z score) | 0.13 ± 1.09 | −1.16 ± 0.99 | 0.027 * |
| Cerebroplacental ratio (z score) | −0.06 ± 0.89 | −1.93 ± 1.6 | 0.027 * |
| Ductus venosus PI (z score) | −1 ± 0.79 | −0.34 ± 1.29 | 0.44 * |
| Maternal biophysical and biochemical profile at the time of sampling | |||
| Gestational age at blood draw (weeks) | 30 ± 1.8 | 31.3 ± 1.8 | 0.16 * |
| Systolic blood pressure (mmHg) | 150.5 ± 18.1 | 107.7 ± 8.3 | <0.001 * |
| Diastolic blood pressure (mmHg) | 89.3 ± 8 | 70.7 ± 3.3 | <0.001 * |
| Creatinine (mg/dL) | 0.42 ± 0.07 | 0.69 ± 0.17 | 0.001 * |
| Urea (mg/dL) | 15.4 ± 4.2 | 40.4 ± 16.7 | 0.002 * |
| Glomerular filtration rate (mL/min) | 85 ± 12.2 | 62.1 ± 8 | <0.001 * |
| Sodium (mEq/L) | 138.3 ± 1.2 | 136.4 ± 2.1 | 0.048 * |
| Potassium (mEq/L) | 4.13 ± 0.21 | 4.61 ± 0.23 | 0.001 * |
| AST (u/L) | 17 (15–23) | 25 (16–48) | 0.16 ω |
| ALT (u/L) | 15 (9–21) | 27 (14–65) | 0.07 ω |
| GGT (u/L) | 10 (7–11) | 19 (15–28) | 0.013 ω |
| Uric acid (mg/dL) | 3.33 ± 0.43 | 6.09 ± 1.25 | <0.001 * |
| Glucose (mg/dL) | 75.5 ± 7.1 | 87.8 ± 19.7 | 0.13 * |
| Triglycerides (mg/dL) | 178.8 ± 67.7 | 260.6 ± 97.8 | 0.06 * |
| Total cholesterol (mg/dL) | 277.2 ± 50.1 | 241.9 ± 54 | 0.16 * |
| Fibrinogen (g/L) | 4.57 ± 0.98 | 3.44 ± 1.05 | 0.07 * |
| Platelets (103/L) | 271.8 ± 70.6 | 209.1 ± 71.9 | 0.09 * |
| Proteinuria (g/24 h) | - | 2.6 (0.6–5.8) | - |
| Protein/creatinine ratio | - | 2.2 (0.4–4.2) | - |
| Perinatal outcomes | |||
| Gestational age at delivery (weeks) | 40.3 ± 1.1 | 32 ± 1.9 | <0.001 * |
| Cesarean section | 1 (16.7) | 11 (78.6) | 0.018 ƒ |
| Female sex | 4 (66.7) | 8 (57.1) | 0.69 ƒ |
| Birthweight (g) | 3531 ± 410 | 1287 ± 267 | <0.001 * |
| Birthweight centile | 58 (41–80) | 0 (0–1) | <0.001 ω |
| APGAR score 5 min < 7 | 0 (0) | 1 (7.1) | 0.50 ƒ |
| Umbilical artery pH | 7.20 ± 0.07 | 7.20 ± 0.09 | 0.97 * |
| HMDB | Metabolite Name | Controls Mean (SD) | Preeclampsia Mean (SD) | p-Value | q-Value | Preeclampsia/Controls |
|---|---|---|---|---|---|---|
| HMDB00192 | Cystine | −0.553 (0.167) | 0.237 (0.251) | <0.0001 | < 0.0001 | Up |
| HMDB00149 | Ethanolamine | −0.397 (0.160) | 0.170 (0.174) | <0.0001 | < 0.0001 | Up |
| HMDB00158 | Tyrosine | −0.431 (0.070) | 0.185 (0.238) | <0.0001 | < 0.0001 | Up |
| HMDB00062 | Free carnitine | −0.572 (0.138) | 0.245 (0.393) | <0.0001 | < 0.0001 | Up |
| HMDB00194 | Anserine | −0.361 (0.130) | 0.155 (0.183) | <0.0001 | 0.0006 | Up |
| HMDB00824 | Propionyl-carnitine | −0.581 (0.079) | 0.249 (0.464) | <0.0001 | 0.0006 | Up |
| HMDB02013 | Iso-butyryl-carnitine/ Butyryl-carnitine | −0.565 (0.120) | 0.242 (0.465) | <0.0001 | 0.0011 | Up |
| HMDB0000138 | Glyco-cholic acid-iso3 | 0.819 (0.401) | −0.351 (0.450) | <0.0001 | 0.0014 | Down |
| HMDB00679 | Homo-citrulline | −0.614 (0.158) | 0.263 (0.318) | <0.0001 | 0.0021 | Up |
| HMDB00243 | Pyruvic acid | −0.505 (0.282) | 0.216 (0.300) | <0.0001 | 0.0034 | Up |
| HMDB00774 | Pregnenolone sulfate | 0.708 (0.395) | −0.303 (0.426) | 0.0001 | 0.0035 | Down |
| HMDB00696 | Methionine | −0.512 (0.144) | 0.219 (0.351) | 0.0001 | 0.0038 | Up |
| HMDB00510 | L-2-Aminoadipic acid | −0.561 (0.334) | 0.240 (0.339) | 0.0001 | 0.0038 | Up |
| HMDB00673 | Linolenic acid-iso2 | −0.567 (0.368) | 0.243 (0.341) | 0.0002 | 0.0044 | Up |
| HMDB00450 | Hydroxylysine | −0.490 (0.292) | 0.210 (0.354) | 0.0002 | 0.0054 | Up |
| HMDB00688 | 2-methyl-butyryl-carnitine/Isovaleryl-carnitine/Valeryl-carnitine | −0.562 (0.219) | 0.241 (0.403) | 0.0002 | 0.0060 | Up |
| HMDB0000138 | Glyco-cholic acid-iso2 | 0.719 (0.650) | −0.308 (0.374) | 0.0003 | 0.0061 | Down |
| HMDB00001 | 1-Methylhistidine | −0.524 (0.168) | 0.224 (0.390) | 0.0003 | 0.0064 | Up |
| HMDB00182 | Lysine | −0.374 (0.181) | 0.160 (0.271) | 0.0004 | 0.0069 | Up |
| HMDB06547 | Stearidonic acid | −0.650 (0.508) | 0.278 (0.534) | 0.0004 | 0.0069 | Up |
| HMDB00904 | Citrulline | −0.300 (0.080) | 0.129 (0.341) | 0.0004 | 0.0075 | Up |
| HMDB00641 | Glutamine | −0.396 (0.169) | 0.170 (0.301) | 0.0005 | 0.0075 | Up |
| HMDB11483 | LPE 20:2-sn1 | 0.616 (0.513) | −0.264 (0.380) | 0.0005 | 0.0075 | Down |
| HMDB00063 | Cortisol | 0.979 (0.168) | −0.420 (0.561) | 0.0006 | 0.0089 | Down |
| HMDB00168 | Asparagine | −0.311 (0.127) | 0.133 (0.352) | 0.0006 | 0.0089 | Up |
| HMDB00177 | Histidine | −0.372 (0.132) | 0.160 (0.299) | 0.0006 | 0.0089 | Up |
| HMDB00234 | Testosterone | 1.018 (0.787) | −0.436 (0.695) | 0.0006 | 0.0089 | Down |
| HMDB0000138 | Glyco-cholic acid-iso1 | 0.716 (0.543) | −0.307 (0.500) | 0.0007 | 0.0094 | Down |
| HMDB00687 | Leucine | −0.376 (0.195) | 0.161 (0.294) | 0.0007 | 0.0096 | Up |
| HMDB00008 | 2-Hydroxybutyric acid | −0.611 (0.395) | 0.262 (0.460) | 0.0008 | 0.0098 | Up |
| NA | LPI 18:1 | 0.631 (0.252) | −0.271 (0.740) | 0.0008 | 0.0098 | Down |
| HMDB00159 | Phenylalanine | −0.366 (0.170) | 0.157 (0.298) | 0.0009 | 0.0104 | Up |
| HMDB10383 | LPC 16:1 e | −0.294 (0.166) | 0.126 (0.235) | 0.0009 | 0.0108 | Up |
| HMDB00208 | alpha-Ketoglutaric acid | −0.368 (0.208) | 0.158 (0.306) | 0.0013 | 0.0139 | Up |
| NA | 11.13-Eicosadienoic acid | −0.530 (0.377) | 0.227 (0.418) | 0.0013 | 0.0139 | Up |
| HMDB00517 | Arginine | −0.297 (0.132) | 0.127 (0.367) | 0.0013 | 0.0139 | Up |
| NA | 17-HDHA | −0.648 (0.384) | 0.278 (0.443) | 0.0013 | 0.0139 | Up |
| HMDB00167 | Threonine | −0.415 (0.295) | 0.178 (0.353) | 0.0015 | 0.0156 | Up |
| HMDB00883 | Valine | −0.344 (0.265) | 0.147 (0.274) | 0.0016 | 0.0157 | Up |
| HMDB06528 | w3-docosapentaenoic acid | −0.524 (0.416) | 0.225 (0.435) | 0.0022 | 0.0203 | Up |
| HMDB00172 | Isoleucine | −0.309 (0.123) | 0.133 (0.289) | 0.0022 | 0.0203 | Up |
| HMDB00148 | Glutamic acid | −0.262 (0.267) | 0.112 (0.191) | 0.0022 | 0.0203 | Up |
| HMDB00214 | Ornithine | −0.439 (0.261) | 0.188 (0.394) | 0.0023 | 0.0203 | Up |
| HMDB00946 | Urso-deoxycholic/ Hyo-deoxycholic acid | 0.650 (0.394) | −0.279 (0.584) | 0.0024 | 0.0204 | Down |
| HMDB07132 HMDB07105 HMDB07051 HMDB07023 HMDB07024 | DG 34:3 | −0.411 (0.335) | 0.176 (0.344) | 0.0024 | 0.0204 | Up |
| HMDB00190 | Lactic acid | −0.295 (0.151) | 0.126 (0.273) | 0.0024 | 0.0204 | Up |
| HMDB00826 | Pentadecanoic acid | −0.440 (0.236) | 0.189 (0.407) | 0.0025 | 0.0206 | Up |
| HMDB05474 | TG 54:6 | −0.517 (0.475) | 0.221 (0.415) | 0.0026 | 0.0206 | Up |
| HMDB02231 | Eico-senoic acid | −0.521 (0.332) | 0.223 (0.474) | 0.0027 | 0.0213 | Up |
| HMDB00271 | Sarcosine | −0.481 (0.338) | 0.206 (0.436) | 0.0030 | 0.0230 | Up |
| HMDB00161 | Alanine | −0.378 (0.185) | 0.162 (0.367) | 0.0033 | 0.0243 | Up |
| HMDB12328 | Trans-palmitoleic acid | −0.497 (0.244) | 0.213 (0.607) | 0.0033 | 0.0243 | Up |
| HMDB06734 | ChoE 202 | 0.320 (0.336) | −0.137 (0.252) | 0.0034 | 0.0243 | Down |
| NA | LPC 22:3-sn1 | 0.629 (0.343) | −0.269 (0.607) | 0.0034 | 0.0243 | Down |
| HMDB05447 HMDB10490 HMDB10502 HMDB10475 HMDB10489 | TG 54:7 | −0.543 (0.565) | 0.233 (0.433) | 0.0035 | 0.0243 | Up |
| HMDB07972 | PC 34:1 e | 0.350 (0.286) | −0.150 (0.315) | 0.0037 | 0.0249 | Down |
| NA | LPI 18:2 | 0.624 (0.343) | −0.267 (0.609) | 0.0037 | 0.0249 | Down |
| HMDB04702 | 12.13-EpOME(9) | −0.461 (0.293) | 0.198 (0.443) | 0.0039 | 0.0254 | Up |
| HMDB10392 | LPC 20:2-sn2 | 0.516 (0.470) | −0.221 (0.455) | 0.0041 | 0.0264 | Down |
| HMDB0240261 | LPI 18:0 | 0.633 (0.280) | −0.271 (0.643) | 0.0042 | 0.0267 | Down |
| HMDB31654 | 3-Aminobutanoic acid | −0.415 (0.227) | 0.178 (0.414) | 0.0042 | 0.0267 | Up |
| HMDB10221 | HODE-iso1 | −0.512 (0.440) | 0.219 (0.475) | 0.0048 | 0.0295 | Up |
| NA | LPI 22:4 | 0.548 (0.358) | −0.235 (0.546) | 0.0049 | 0.0300 | Down |
| NA | LPI 20:4 | 0.590 (0.305) | −0.253 (0.613) | 0.0053 | 0.0320 | Down |
| HMDB10386 | LPC 18:2-sn2 | 0.453 (0.348) | −0.194 (0.446) | 0.0056 | 0.0330 | Down |
| HMDB00694 | 2-hydroxyglutaric acid | −0.346 (0.099) | 0.148 (0.557) | 0.0060 | 0.0349 | Up |
| HMDB08287 HMDB08257 HMDB08318 HMDB08383 HMDB08286 | PC 42:5 e | 0.339 (0.292) | −0.145 (0.333) | 0.0064 | 0.0364 | Down |
| NA | LPI 14:0 | 0.660 (0.510) | −0.283 (0.689) | 0.0077 | 0.0432 | Down |
| HMDB06547 | Stearidonic acid-iso1 | −0.405 (0.284) | 0.173 (0.431) | 0.0078 | 0.0432 | Up |
| HMDB01383 | Sphingosine-1-P | 0.402 (0.341) | −0.172 (0.414) | 0.0081 | 0.0440 | Down |
| HMDB01032 | Dehydro-epiandrost-erone sulfate | 0.607 (0.361) | −0.260 (0.670) | 0.0085 | 0.0440 | Down |
| HMDB00123 | Glycine | −0.260 (0.085) | 0.111 (0.305) | 0.0087 | 0.0440 | Up |
| HMDB00575 | Homo-cystine | −0.472 (0.373) | 0.202 (0.649) | 0.0087 | 0.0440 | Up |
| HMDB05461 | TG 54:5 | −0.324 (0.164) | 0.139 (0.614) | 0.0087 | 0.0440 | Up |
| HMDB02802 | Cortisone | 0.715 (0.138) | −0.307 (0.574) | 0.0087 | 0.0440 | Down |
| HMDB02226 | Adrenic acid | −0.421 (0.335) | 0.180 (0.447) | 0.0087 | 0.0440 | Up |
| HMDB00112 | gamma-Aminobutyric acid | −0.449 (0.609) | 0.193 (0.373) | 0.0092 | 0.0460 | Up |
| HMDB00162 | Proline | −0.308 (0.256) | 0.132 (0.329) | 0.0096 | 0.0471 | Up |
| HMDB00673 | Linolenic acid-iso1 | −0.414 (0.387) | 0.178 (0.432) | 0.0098 | 0.0474 | Up |
| HMDB10391 | LPC 20:1-sn2 | 0.526 (0.403) | −0.225 (0.578) | 0.0100 | 0.0480 | Down |
| NA | LPC 22:3-sn2 | 0.498 (0.458) | −0.214 (0.530) | 0.0105 | 0.0497 | Down |
| NA | LPI 16:0 | 0.577 (0.324) | −0.247 (0.668) | 0.0107 | 0.0498 | Down |
| Pathway | Total | Expected | Hits | Raw p | −log(10)p | Adjusted p | FDR | Impact |
|---|---|---|---|---|---|---|---|---|
| Aminoacyl-tRNA biosynthesis | 48 | 17.03 | 16 | 0.0006 × 10−9 | 12.20 | 0.05 × 10−9 | 0.05 × 10−9 | 0 |
| Arginine biosynthesis | 14 | 0.49 | 6 | 0.004 × 10−3 | 54.37 | 0.0003 | 0.0002 | 0.48 |
| Alanine, aspartate and glutamate metabolism | 28 | 0.99 | 7 | 0.03 × 10−3 | 44.95 | 0.003 | 0.0009 | 0.45 |
| Valine, leucine and isoleucine biosynthesis | 8 | 0.28 | 4 | 0.09 × 10−3 | 40.48 | 0.007 | 0.002 | 0 |
| Linoleic acid metabolism | 5 | 0.18 | 3 | 0.0004 | 33.95 | 0.032 | 0.007 | 1 |
| D-Glutamine and D-glutamate metabolism | 6 | 0.21 | 3 | 0.0008 | 31.05 | 0.06 | 0.011 | 0.50 |
| Arginine and proline metabolism | 38 | 13.48 | 6 | 0.0017 | 27.60 | 0.14 | 0.020 | 0.36 |
| Histidine metabolism | 16 | 0.57 | 4 | 0.0018 | 27.25 | 0.14 | 0.020 | 0.27 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 4 | 0.14 | 2 | 0.007 | 21.50 | 0.54 | 0.07 | 1 |
| Butanoate metabolism | 15 | 0.53 | 3 | 0.014 | 18.47 | 1 | 0.12 | 0.03 |
| Nitrogen metabolism | 6 | 0.21 | 2 | 0.017 | 17.72 | 1 | 0.13 | 0 |
| Glyoxylate and dicarboxylate metabolism | 32 | 11.36 | 4 | 0.024 | 1.61 | 1 | 0.17 | 0.11 |
| Glycine, serine and threonine metabolism | 33 | 1.17 | 4 | 0.027 | 15.68 | 1 | 0.17 | 0.34 |
| Phenylalanine metabolism | 10 | 0.35 | 2 | 0.05 | 13.34 | 1 | 0.28 | 0.36 |
| Lysine degradation | 25 | 0.89 | 3 | 0.06 | 1.25 | 1 | 0.31 | 0.14 |
| Glutathione metabolism | 28 | 0.99 | 3 | 0.07 | 11.31 | 1 | 0.37 | 0.11 |
| alpha-Linolenic acid metabolism | 13 | 0.46 | 2 | 0.08 | 11.24 | 1 | 0.37 | 0 |
| Cysteine and methionine metabolism | 33 | 1.17 | 3 | 0.11 | 0.96 | 1 | 0.51 | 0.10 |
| Glycerophospholipid metabolism | 36 | 12.77 | 3 | 0.13 | 0.88 | 1 | 0.59 | 0.13 |
| Citrate cycle (TCA cycle) | 20 | 0.71 | 2 | 0.16 | 0.81 | 1 | 0.65 | 0.10 |
| Valine, leucine and isoleucine degradation | 40 | 14.19 | 3 | 0.17 | 0.78 | 1 | 0.65 | 0 |
| beta-Alanine metabolism | 21 | 0.75 | 2 | 0.17 | 0.77 | 1 | 0.65 | 0 |
| Pyruvate metabolism | 22 | 0.78 | 2 | 0.18 | 0.74 | 1 | 0.67 | 0.29 |
| Ubiquinone and other terpenoid-quinone biosynthesis | 9 | 0.32 | 1 | 0.28 | 0.56 | 1 | 0.97 | 0 |
| Porphyrin and chlorophyll metabolism | 30 | 10.65 | 2 | 0.29 | 0.54 | 1 | 0.97 | 0 |
| Biotin metabolism | 10 | 0.35 | 1 | 0.30 | 0.52 | 1 | 0.98 | 0 |
| Steroid hormone biosynthesis | 85 | 30.16 | 4 | 0.36 | 0.45 | 1 | 1 | 0.08 |
| Glycero-lipid metabolism | 16 | 0.57 | 1 | 0.44 | 0.36 | 1 | 1 | 0.01 |
| Tyrosine metabolism | 42 | 14.90 | 2 | 0.44 | 0.35 | 1 | 1 | 0.14 |
| Primary bile acid biosynthesis | 46 | 16.32 | 2 | 0.49 | 0.31 | 1 | 1 | 0.02 |
| Pantothenate and CoA biosynthesis | 19 | 0.67 | 1 | 0.50 | 0.30 | 1 | 1 | 0 |
| Seleno-compound metabolism | 20 | 0.71 | 1 | 0.52 | 0.29 | 1 | 1 | 0 |
| Sphingolipid metabolism | 21 | 0.75 | 1 | 0.53 | 0.27 | 1 | 1 | 0.02 |
| Propanoate metabolism | 23 | 0.82 | 1 | 0.57 | 0.25 | 1 | 1 | 0 |
| Glycolysis/Gluconeogenesis | 26 | 0.92 | 1 | 0.61 | 0.21 | 1 | 1 | 0.10 |
| Arachidonic acid metabolism | 36 | 12.77 | 1 | 0.73 | 0.14 | 1 | 1 | 0 |
| Biosynthesis of unsaturated fatty acids | 36 | 12.77 | 1 | 0.73 | 0.14 | 1 | 1 | 0 |
| Pyrimidine metabolism | 39 | 13.84 | 1 | 0.76 | 0.12 | 1 | 1 | 0 |
| Steroid biosynthesis | 42 | 14.90 | 1 | 0.79 | 0.11 | 1 | 1 | 0 |
| Purine metabolism | 65 | 23.07 | 1 | 0.91 | 0.04 | 1 | 1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, L.; Crovetto, F.; Simoes, R.V.; Miranda, J.; Paules, C.; Blasco, M.; Palomo, M.; García-Calderó, H.; Tura-Ceide, O.; Dantas, A.P.; et al. The Interplay between Pathophysiological Pathways in Early-Onset Severe Preeclampsia Unveiled by Metabolomics. Life 2022, 12, 86. https://doi.org/10.3390/life12010086
Youssef L, Crovetto F, Simoes RV, Miranda J, Paules C, Blasco M, Palomo M, García-Calderó H, Tura-Ceide O, Dantas AP, et al. The Interplay between Pathophysiological Pathways in Early-Onset Severe Preeclampsia Unveiled by Metabolomics. Life. 2022; 12(1):86. https://doi.org/10.3390/life12010086
Chicago/Turabian StyleYoussef, Lina, Francesca Crovetto, Rui Vasco Simoes, Jezid Miranda, Cristina Paules, Miquel Blasco, Marta Palomo, Héctor García-Calderó, Olga Tura-Ceide, Ana Paula Dantas, and et al. 2022. "The Interplay between Pathophysiological Pathways in Early-Onset Severe Preeclampsia Unveiled by Metabolomics" Life 12, no. 1: 86. https://doi.org/10.3390/life12010086