
Overexpression of p75NTR in Human Seminoma: A New Biomarker?

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Human Tissues
2.3. Tissue Microarray (TMA) Construction and Validation
2.4. Histopathological Analysis
2.5. Immunohistochemistry
2.6. Scoring System
2.7. Statistical Analysis
3. Results
3.1. Morphology Analysis in Control Testis and Seminoma
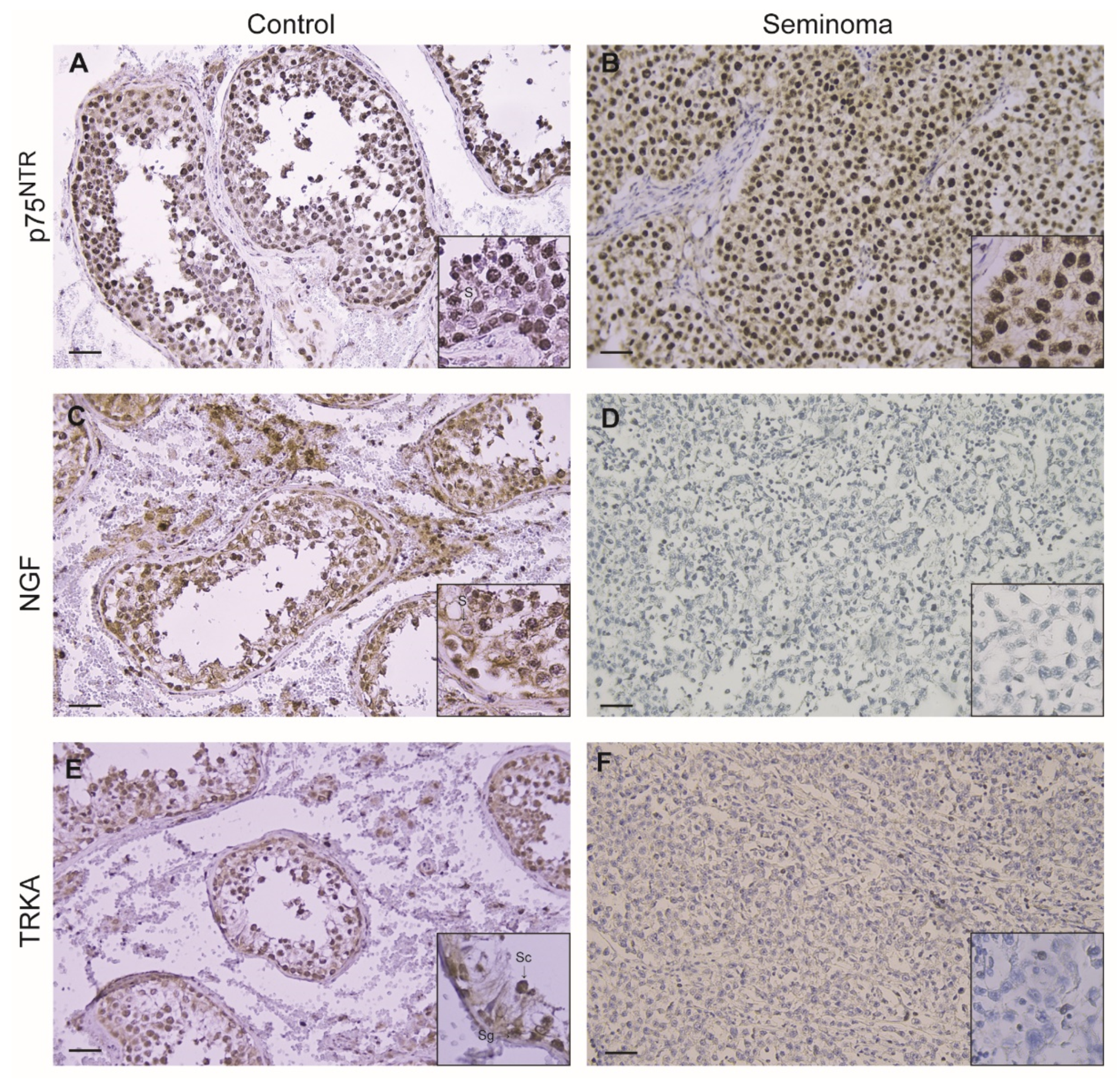
3.2. Immunohistochemical Localization of p75NTR, NGF and TrkA in Control Testis and TGCC
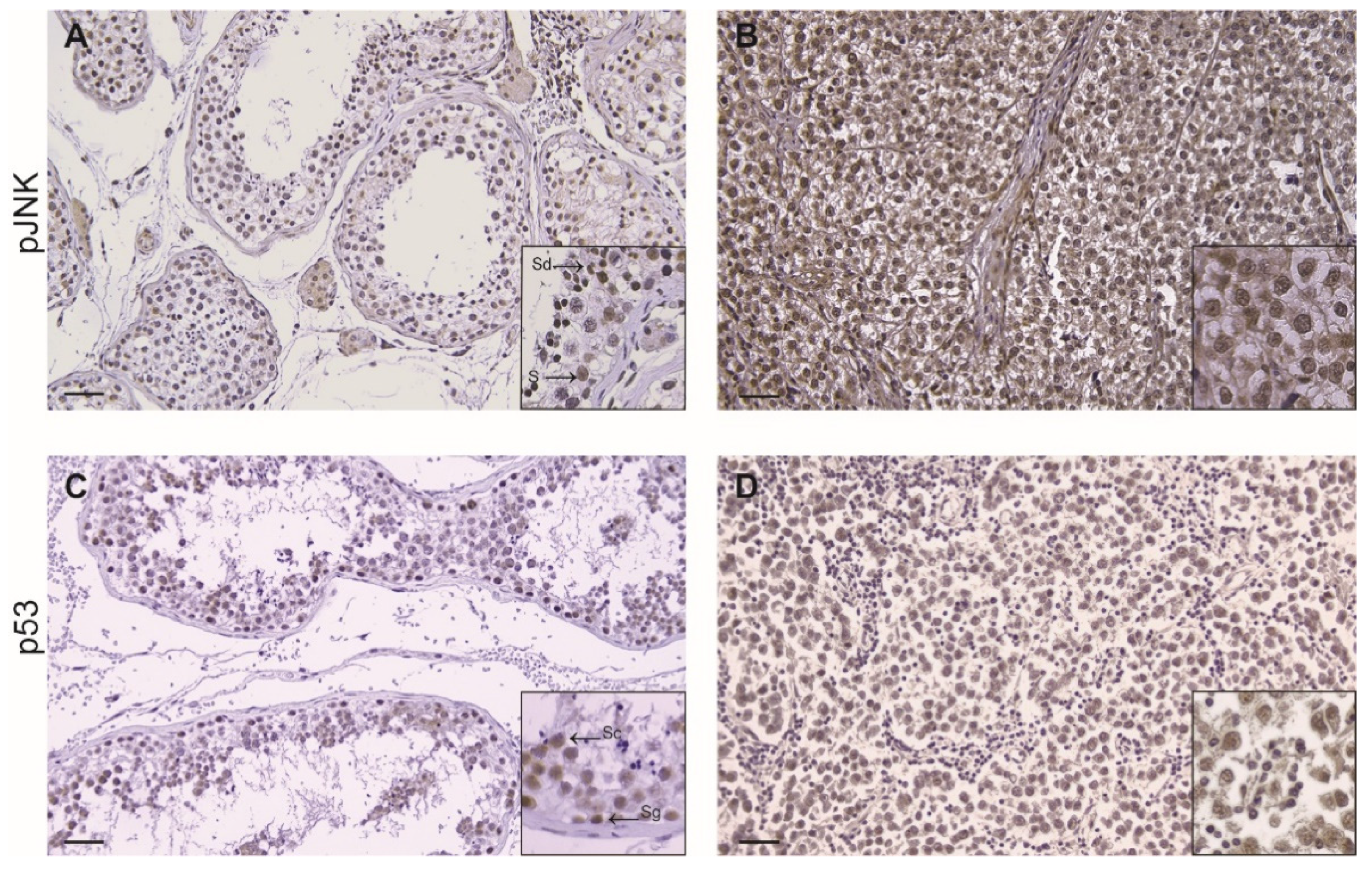
3.3. Immunohistochemistry of p53 in TGCC
3.4. Validation of TMA Method
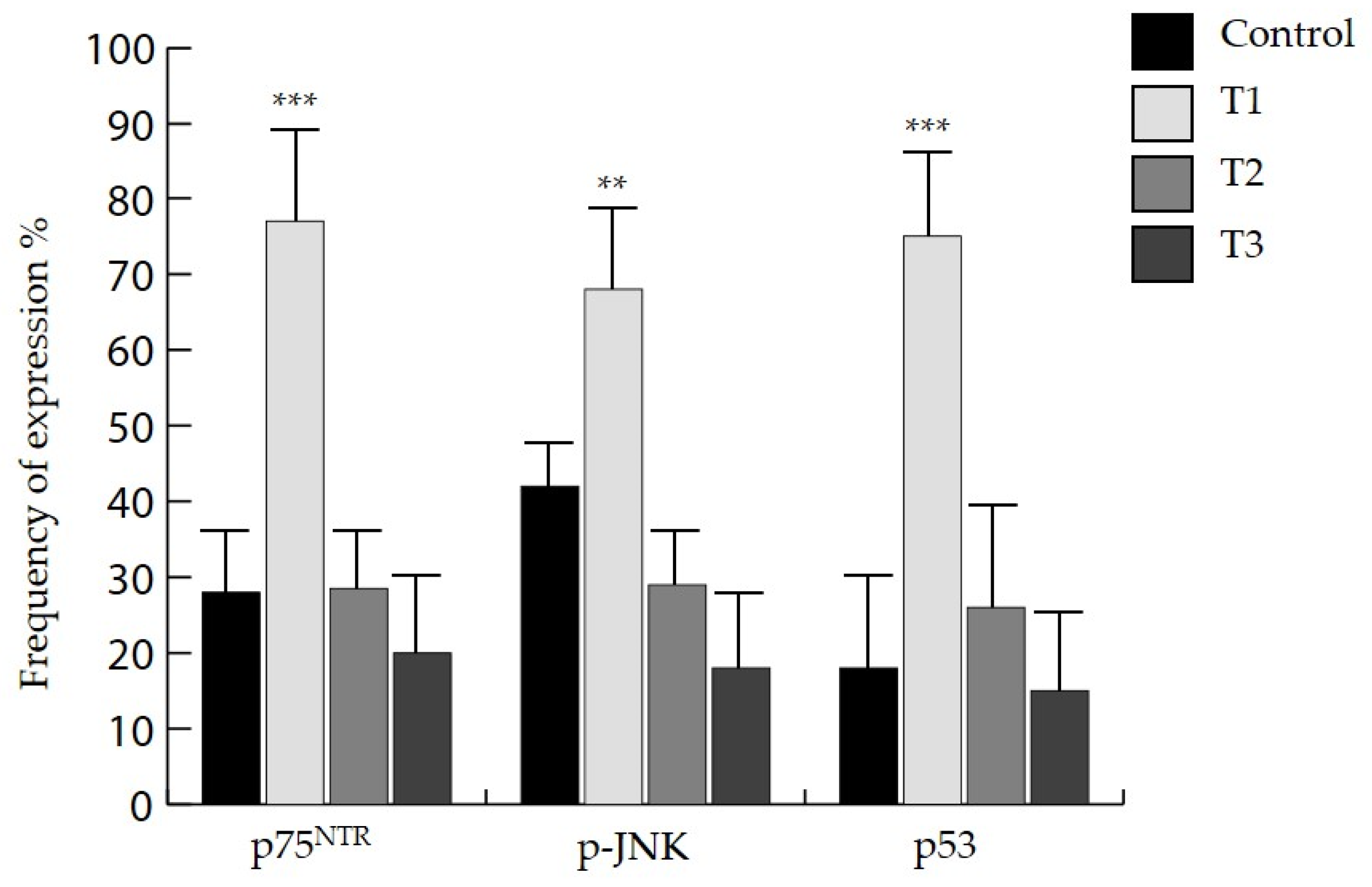
3.5. Clinico-Pathological Significance of p75NTR, p-JNK and p53
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghazarian, A.A.; Kelly, S.P.; Altekruse, S.F.; Rosenberg, P.S.; McGlynn, K.A. Future of Testicular Germ Cell Tumor Incidence in the United States: Forecast through 2026. Cancer 2017, 123, 2320–2328. [Google Scholar] [CrossRef] [Green Version]
- Bode, P.K.; Thielken, A.; Brandt, S.; Barghorn, A.; Lohe, B.; Knuth, A.; Moch, H. Cancer Testis Antigen Expression in Testicular Germ Cell Tumorigenesis. Mod. Pathol. 2014, 27, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Cornet, C.; Lortet-Tieulent, J.; Forman, D.; Béranger, R.; Flechon, A.; Fervers, B.; Schüz, J.; Bray, F. Testicular Cancer Incidence to Rise by 25% by 2025 in Europe? Model-Based Predictions in 40 Countries Using Population-Based Registry Data. Eur. J. Cancer 2014, 50, 831–839. [Google Scholar] [CrossRef]
- Milardi, D.; Grande, G.; Vincenzoni, F.; Pierconti, F.; Pontecorvi, A. Proteomics for the Identification of Biomarkers in Testicular Cancer-Review. Front. Endocrinol. 2019, 10, 462. [Google Scholar] [CrossRef] [Green Version]
- Batool, A.; Karimi, N.; Wu, X.N.; Chen, S.R.; Liu, Y.X. Testicular Germ Cell Tumor: A Comprehensive Review. Cell. Mol. Life Sci. 2019, 76, 1713–1727. [Google Scholar] [CrossRef]
- Skakkebaek, N.E. Possible Carcinoma-in-situ of the Testis. Lancet Lond. Engl. 1972, 2, 516–517. [Google Scholar] [CrossRef]
- Sheikine, Y.; Genega, E.; Melamed, J.; Lee, P.; Reuter, V.E.; Ye, H. Molecular Genetics of Testicular Germ Cell Tumors. Am. J. Cancer Res. 2012, 2, 153–167. [Google Scholar] [PubMed]
- Chen, S.R.; Liu, Y.X. Regulation of Spermatogonial Stem Cell Self-Renewal and Spermatocyte Meiosis by Sertoli Cell Signaling. Reprod. Camb. Engl. 2015, 149, R159–R167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.R.; Zheng, Q.S.; Zhang, Y.; Gao, F.; Liu, Y.X. Disruption of Genital Ridge Development Causes Aberrant Primordial Germ Cell Proliferation but Does Not Affect Their Directional Migration. BMC Biol. 2013, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Rebourcet, D.; O’Shaughnessy, P.J.; Pitetti, J.L.; Monteiro, A.; O’Hara, L.; Milne, L.; Tsai, Y.T.; Cruickshanks, L.; Riethmacher, D.; Guillou, F.; et al. Sertoli Cells Control Peritubular Myoid Cell Fate and Support Adult Leydig Cell Development in the Prepubertal Testis. Dev. Camb. Engl. 2014, 141, 2139–2149. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Q.; Batool, A.; Chen, S.R.; Liu, Y.X. GATA4 Is a Negative Regulator of Contractility in Mouse Testicular Peritubular Myoid Cells. Reprod. Camb. Engl. 2018, 156, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Persson, H.; Ayer-Le Lievre, C.; Söder, O.; Villar, M.J.; Metsis, M.; Olson, L.; Ritzen, M.; Hökfelt, T. Expression of Beta-Nerve Growth Factor Receptor MRNA in Sertoli Cells Downregulated by Testosterone. Science 1990, 247, 704–707. [Google Scholar] [CrossRef]
- Müller, D.; Davidoff, M.S.; Bargheer, O.; Paust, H.J.; Pusch, W.; Koeva, Y.; Jezek, D.; Holstein, A.F.; Middendorff, R. The Expression of Neurotrophins and Their Receptors in the Prenatal and Adult Human Testis: Evidence for Functions in Leydig Cells. Histochem. Cell Biol. 2006, 126, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.A.; Odorisio, T.; Fradeani, A.; Rienzi, L.; De Felici, M.; Cattaneo, A.; Siracusa, G. Low-Affinity Nerve Growth Factor Receptor Is Expressed during Testicular Morphogenesis and in Germ Cells at Specific Stages of Spermatogenesis. Mol. Reprod. Dev. 1994, 37, 157–166. [Google Scholar] [CrossRef]
- Djakiew, D.; Pflug, B.; Dionne, C.; Onoda, M. Postnatal Expression of Nerve Growth Factor Receptors in the Rat Testis. Biol. Reprod. 1994, 51, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Zhou, X. The Potential Roles of Neurotrophins in Male Reproduction. Reprod. Camb. Engl. 2013, 145, R89–R95. [Google Scholar] [CrossRef] [Green Version]
- Griffin, N.; Faulkner, S.; Jobling, P.; Hondermarck, H. Targeting Neurotrophin Signaling in Cancer: The Renaissance. Pharmacol. Res. 2018, 135, 12–17. [Google Scholar] [CrossRef] [PubMed]
- AIOM—Associazione Italiana di Oncologia Medica. Tumore del Testicolo: Linee Guida; AIOM: Milan, Italy, 2018. [Google Scholar]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and Predictive Factors in Breast Cancer by Immunohistochemical Analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar]
- Jin, H.; Pan, Y.; He, L.; Zhai, H.; Li, X.; Zhao, L.; Sun, L.; Liu, J.; Hong, L.; Song, J.; et al. P75 Neurotrophin Receptor Inhibits Invasion and Metastasis of Gastric Cancer. Mol. Cancer Res. 2007, 5, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Khwaja, F.S.; Quann, E.J.; Pattabiraman, N.; Wynne, S.; Djakiew, D. Carprofen Induction of P75NTR-Dependent Apoptosis via the P38 Mitogen-Activated Protein Kinase Pathway in Prostate Cancer Cells. Mol. Cancer Ther. 2008, 7, 3539–3545. [Google Scholar] [CrossRef] [Green Version]
- Gravina, G.L.; Marampon, F.; Sanità, P.; Mancini, A.; Colapietro, A.; Scarsella, L.; Jitariuc, A.; Biordi, L.; Ficorella, C.; Festuccia, C. Increased Expression and Activity of P75NTR Are Crucial Events in Azacitidine-Induced Cell Death in Prostate Cancer. Oncol. Rep. 2016, 36, 125–130. [Google Scholar] [CrossRef]
- Marchetti, D.; Aucoin, R.; Blust, J.; Murry, B.; Greiter-Wilke, A. P75 Neurotrophin Receptor Functions as a Survival Receptor in Brain-Metastatic Melanoma Cells. J. Cell. Biochem. 2004, 91, 206–215. [Google Scholar] [CrossRef]
- Wang, L.; Rahn, J.J.; Lun, X.; Sun, B.; Kelly, J.J.; Weiss, S.; Robbins, S.M.; Forsyth, P.A.; Senger, D.L. Gamma-Secretase Represents a Therapeutic Target for the Treatment of Invasive Glioma Mediated by the P75 Neurotrophin Receptor. PLoS Biol. 2008, 6, e289. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, S.; Tomellini, E.; Dhamani, F.; Meignan, S.; Adriaenssens, E.; Xuefen, L.B. Extracellular Cleavage of the P75 Neurotrophin Receptor Is Implicated in Its Pro-Survival Effect in Breast Cancer Cells. FEBS Lett. 2013, 587, 2591–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieckmann, K.P.; Richter-Simonsen, H.; Kulejewski, M.; Ikogho, R.; Zecha, H.; Anheuser, P.; Pichlmeier, U.; Isbarn, H. Testicular Germ-Cell Tumours: A Descriptive Analysis of Clinical Characteristics at First Presentation. Urol. Int. 2018, 100, 409–419. [Google Scholar] [CrossRef]
- Pierconti, F.; Milardi, D.; Martini, M.; Grande, G.; Cenci, T.; Gulino, G.; Larocca, L.M.; Rindi, G.; Pontecorvi, A.; De Marinis, L. Pituitary-Tumour-Transforming-Gene 1 Expression in Testicular Cancer. Andrologia 2015, 47, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Grande, G.; Milardi, D.; Martini, M.; Cenci, T.; Gulino, G.; Mancini, F.; Bianchi, A.; Pontecorvi, A.; Pierconti, F. Protein Expression of PTTG-1, OCT-4, and KLF-4 in Seminoma: A Pilot Study. Front. Endocrinol. 2019, 10, 619. [Google Scholar] [CrossRef] [PubMed]
- Rabizadeh, S.; Oh, J.; Zhong, L.T.; Yang, J.; Bitler, C.M.; Butcher, L.L.; Bredesen, D.E. Induction of Apoptosis by the Low-Affinity NGF Receptor. Science 1993, 261, 345–348. [Google Scholar] [CrossRef]
- Gentry, J.J.; Casaccia-Bonnefil, P.; Carter, B.D. Nerve Growth Factor Activation of Nuclear Factor KappaB through Its P75 Receptor Is an Anti-Apoptotic Signal in RN22 Schwannoma Cells. J. Biol. Chem. 2000, 275, 7558–7565. [Google Scholar] [CrossRef] [Green Version]
- Micera, A.; Lambiase, A.; Stampachiacchiere, B.; Bonini, S.; Bonini, S.; Levi-Schaffer, F. Nerve Growth Factor and Tissue Repair Remodeling: TrkA(NGFR) and P75(NTR), Two Receptors One Fate. Cytokine Growth Factor Rev. 2007, 18, 245–256. [Google Scholar] [CrossRef]
- Sperger, J.M.; Chen, X.; Draper, J.S.; Antosiewicz, J.E.; Chon, C.H.; Jones, S.B.; Brooks, J.D.; Andrews, P.W.; Brown, P.O.; Thomson, J.A. Gene Expression Patterns in Human Embryonic Stem Cells and Human Pluripotent Germ Cell Tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 13350–13355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajpert-De Meyts, E. Developmental Model for the Pathogenesis of Testicular Carcinoma in Situ: Genetic and Environmental Aspects. Hum. Reprod. Update 2006, 12, 303–323. [Google Scholar] [CrossRef] [Green Version]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective Identification of Tumorigenic Breast Cancer Cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of Human Brain Tumour Initiating Cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous Stem Cells Can Arise from Pediatric Brain Tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef] [Green Version]
- Dean, M.; Fojo, T.; Bates, S. Tumour Stem Cells and Drug Resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar] [CrossRef]
- Tomellini, E.; Lagadec, C.; Polakowska, R.; Le Bourhis, X. Role of P75 Neurotrophin Receptor in Stem Cell Biology: More than Just a Marker. Cell. Mol. Life Sci. 2014, 71, 2467–2481. [Google Scholar] [CrossRef]
- Okumura, T.; Tsunoda, S.; Mori, Y.; Ito, T.; Kikuchi, K.; Wang, T.C.; Yasumoto, S.; Shimada, Y. The Biological Role of the Low-Affinity P75 Neurotrophin Receptor in Esophageal Squamous Cell Carcinoma. Clin. Cancer Res. 2006, 12, 5096–5103. [Google Scholar] [CrossRef] [Green Version]
- Boiko, A.D.; Razorenova, O.V.; van de Rijn, M.; Swetter, S.M.; Johnson, D.L.; Ly, D.P.; Butler, P.D.; Yang, G.P.; Joshua, B.; Kaplan, M.J.; et al. Human Melanoma-Initiating Cells Express Neural Crest Nerve Growth Factor Receptor CD271. Nature 2010, 466, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Herrera, S.C.; Bach, E.A. JNK Signaling Triggers Spermatogonial Dedifferentiation during Chronic Stress to Maintain the Germline Stem Cell Pool in the Drosophila Testis. eLife 2018, 7, e36095. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Banerjee, P.; Nikolakaki, E.; Dai, T.; Rubie, E.A.; Ahmad, M.F.; Avruch, J.; Woodgett, J.R. The Stress-Activated Protein Kinase Subfamily of c-Jun Kinases. Nature 1994, 369, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Bubici, C.; Papa, S. JNK Signalling in Cancer: In Need of New, Smarter Therapeutic Targets. Br. J. Pharmacol. 2014, 171, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Bhakar, A.L.; Kennedy, T.E.; Barker, P.A. The P75 Neurotrophin Receptor Activates Akt (Protein Kinase B) through a Phosphatidylinositol 3-Kinase-Dependent Pathway. J. Biol. Chem. 2001, 276, 23097–23104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pflug, B.R.; Onoda, M.; Lynch, J.H.; Djakiew, D. Reduced Expression of the Low Affinity Nerve Growth Factor Receptor in Benign and Malignant Human Prostate Tissue and Loss of Expression in Four Human Metastatic Prostate Tumor Cell Lines. Cancer Res. 1992, 52, 5403–5406. [Google Scholar] [PubMed]
- Krygier, S.; Djakiew, D. Neurotrophin Receptor P75(NTR) Suppresses Growth and Nerve Growth Factor-Mediated Metastasis of Human Prostate Cancer Cells. Int. J. Cancer 2002, 98, 1–7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 4) | Seminoma (n = 26) | |
|---|---|---|
| p75NTR | 4 (3–4) | 8 (7–8) *** |
| NGF | 2 (2–3) | 0 (0–0) |
| TrkA | 3 (2–4) | 0 (0–0) |
| Marker | Cutoff | Sensitivity (%) | Specificity (%) | PPV | NPV | AUC (95% CI) |
|---|---|---|---|---|---|---|
| p75NTR | ≥7 | 67.8% | 59.0% | 21.4% | 76.0% | 0.80 (0.75–0.98) |
| Marker | Control (n = 4) | Seminoma (n = 26) |
|---|---|---|
| p-JNK | 3 (2–3) | 7 (6–8) *** |
| P53 | 3 (2–3) | 7 (6–8) *** |
| nr | NGF | TrkA | p75NTR | ||||
|---|---|---|---|---|---|---|---|
| Positive (%) | p | Positive (%) | p | Positive (%) | p | ||
| T stage | 0.91 | 0.40 | 0.001 | ||||
| T1 | 27 | 1 (0.03) | 5 (5.9) | 21 (77.7) | |||
| T2 | 7 | 0 (0.0) | 0 (0.0) | 2 (28.5) | |||
| T3 | 5 | 0 (0.0) | 0 (0.0) | 1 (20.0) | |||
| T4 | 1 | 0 (0.0) | 0 (0.0) | 0 (0.0) | |||
| Vascular invasion | ns | ns | 0.59 | ||||
| No | 34 | 0 (0.0) | 0 (0.0) | 13 (38.2) | |||
| Yes | 6 | 0 (0.0) | 0 (0.0) | 3 (64.3) | |||
| Histology | ns | ns | 0.0001 | ||||
| Seminoma | 26 | 0 (0.0) | 0 (0.0) | 25 (96.1) | |||
| Non-seminomatous | 14 | 0 (0.0) | 0 (0.0) | 4 (28.5) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perri, A.; Rago, V.; Malivindi, R.; Maltese, L.; Lofaro, D.; Greco, E.A.; Tucci, L.; Bonofiglio, R.; Vergine, M.; La Vignera, S.; et al. Overexpression of p75NTR in Human Seminoma: A New Biomarker? Life 2021, 11, 629. https://doi.org/10.3390/life11070629
Perri A, Rago V, Malivindi R, Maltese L, Lofaro D, Greco EA, Tucci L, Bonofiglio R, Vergine M, La Vignera S, et al. Overexpression of p75NTR in Human Seminoma: A New Biomarker? Life. 2021; 11(7):629. https://doi.org/10.3390/life11070629
Chicago/Turabian StylePerri, Anna, Vittoria Rago, Rocco Malivindi, Lorenza Maltese, Danilo Lofaro, Emanuela Alessandra Greco, Luigi Tucci, Renzo Bonofiglio, Margherita Vergine, Sandro La Vignera, and et al. 2021. "Overexpression of p75NTR in Human Seminoma: A New Biomarker?" Life 11, no. 7: 629. https://doi.org/10.3390/life11070629