Erythromycin Treatment of Brassica campestris Seedlings Impacts the Photosynthetic and Protein Synthesis Pathways


Abstract
:1. Introduction
2. Materials and Methods
2.1. Erythromycin Treatment and Measurement of Plant Growth and Phyiological Parameters
2.2. Detection of Erythromycin Residue in Brassica campestris Seedlings
2.3. Total Protein Extraction and One-Dimemnsional Gel Electrophoresis
2.4. In-Gel Digestion
2.5. LC-MS/MS Analysis
2.6. Data Analysis
2.7. RNA Extraction and qRT-PCR
3. Results
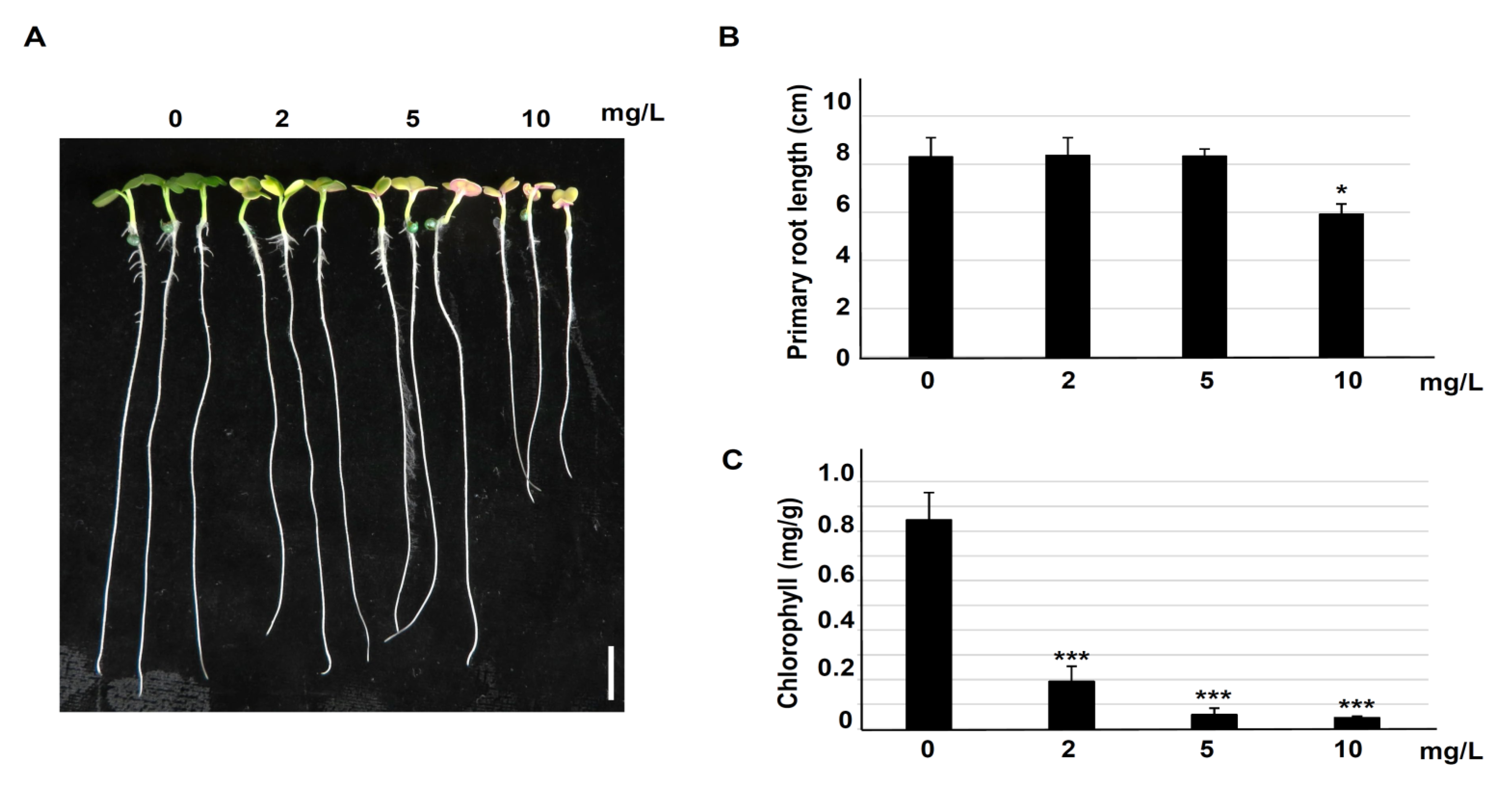
3.1. Erythromycin Inhibits Brassia campestris Seedling Growth
3.2. Erythromycin Accumulation in Vegetative Tissue
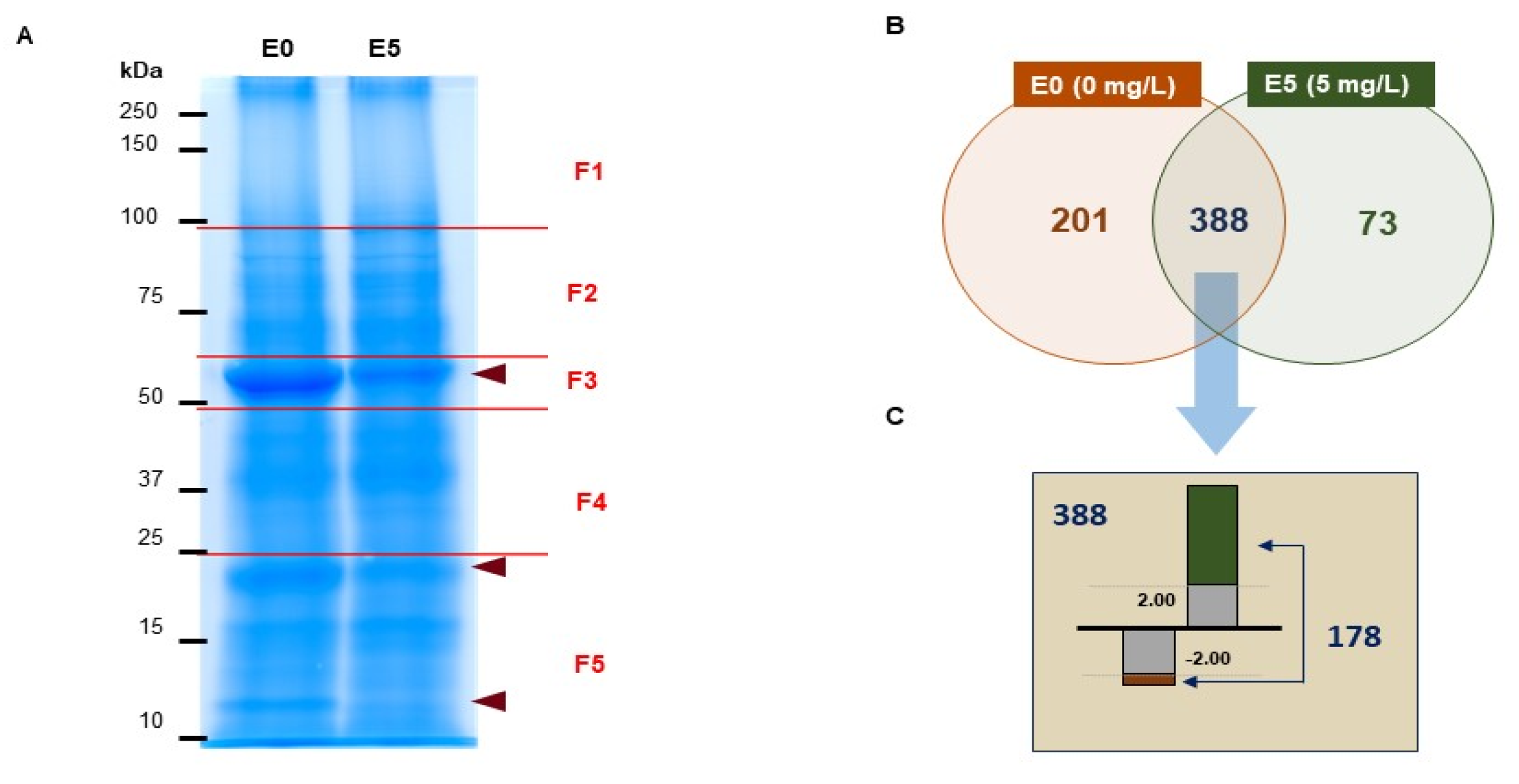
3.3. Differential Proteomic Analysis of Brassica campestris Seedlings
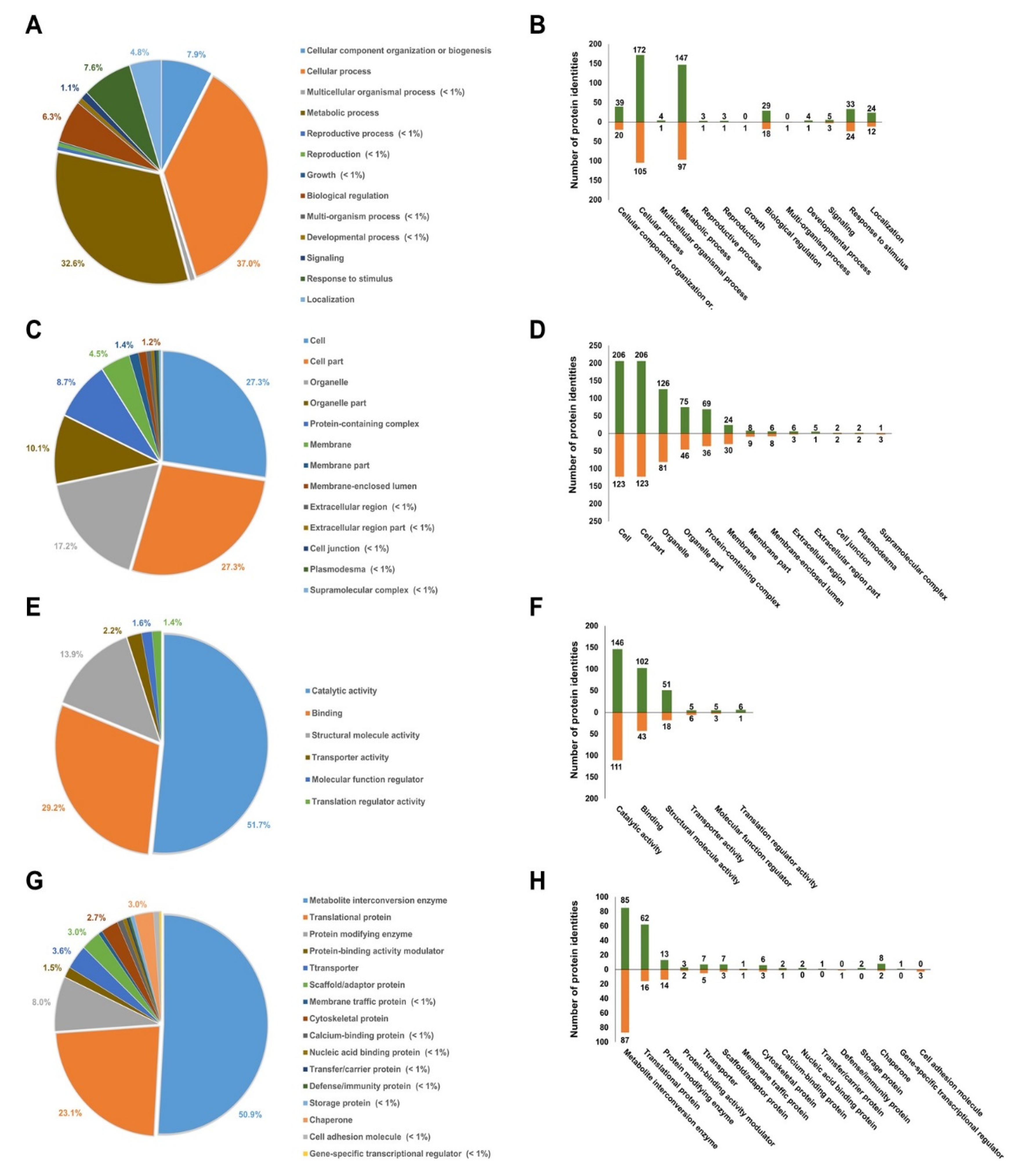
3.4. Gene Ontology Analysis of Differentially Expressed Proteins
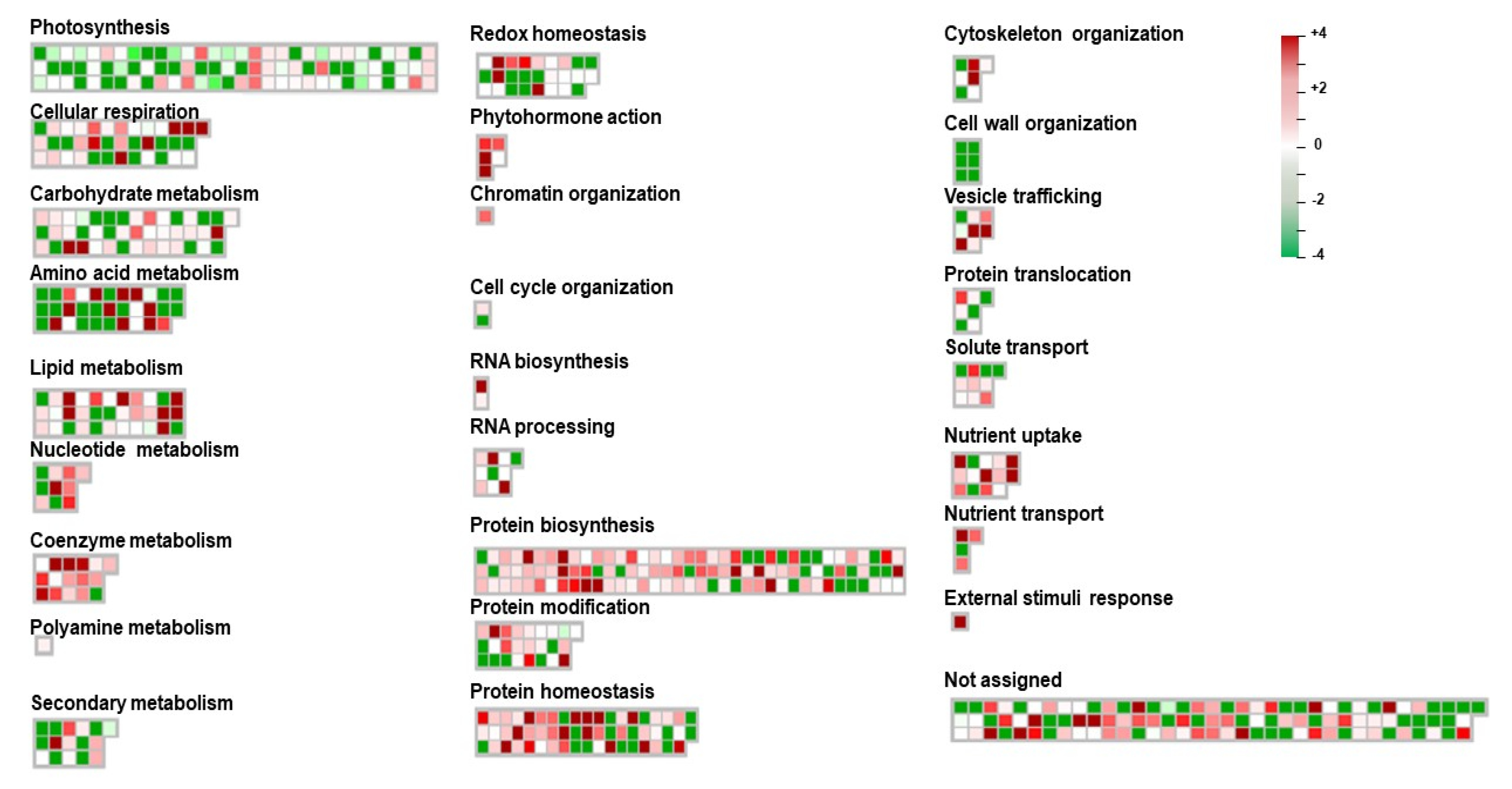
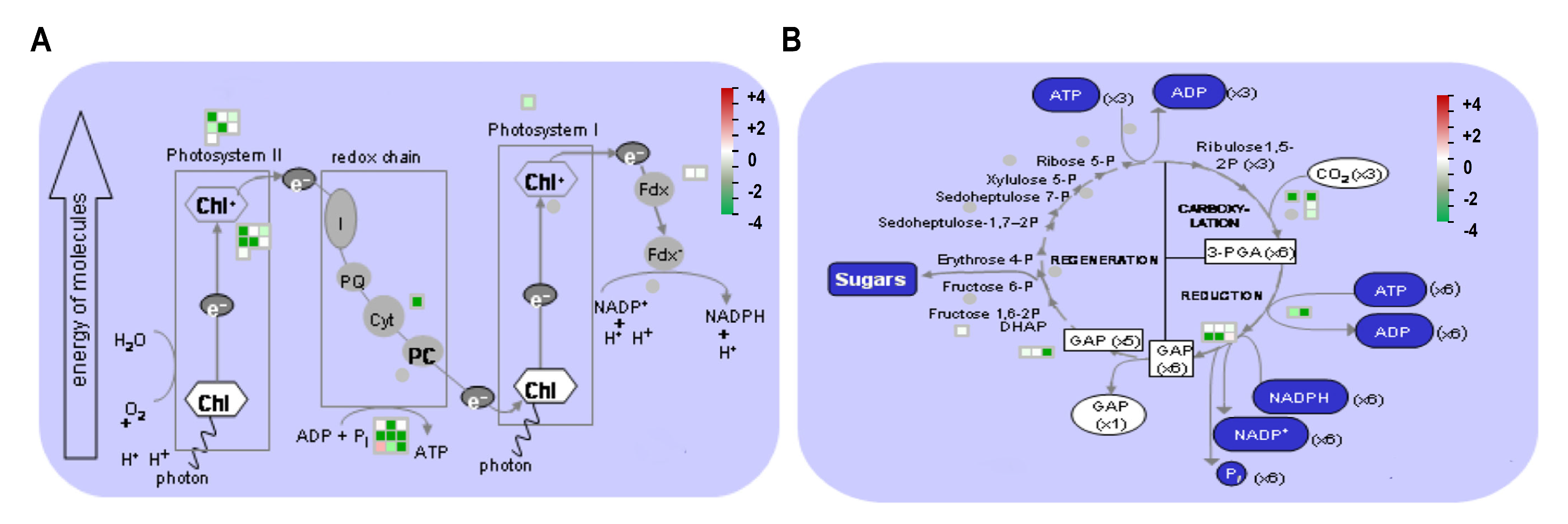
3.5. MapMan Pathway Analysis of Differentially Expressed Proteins
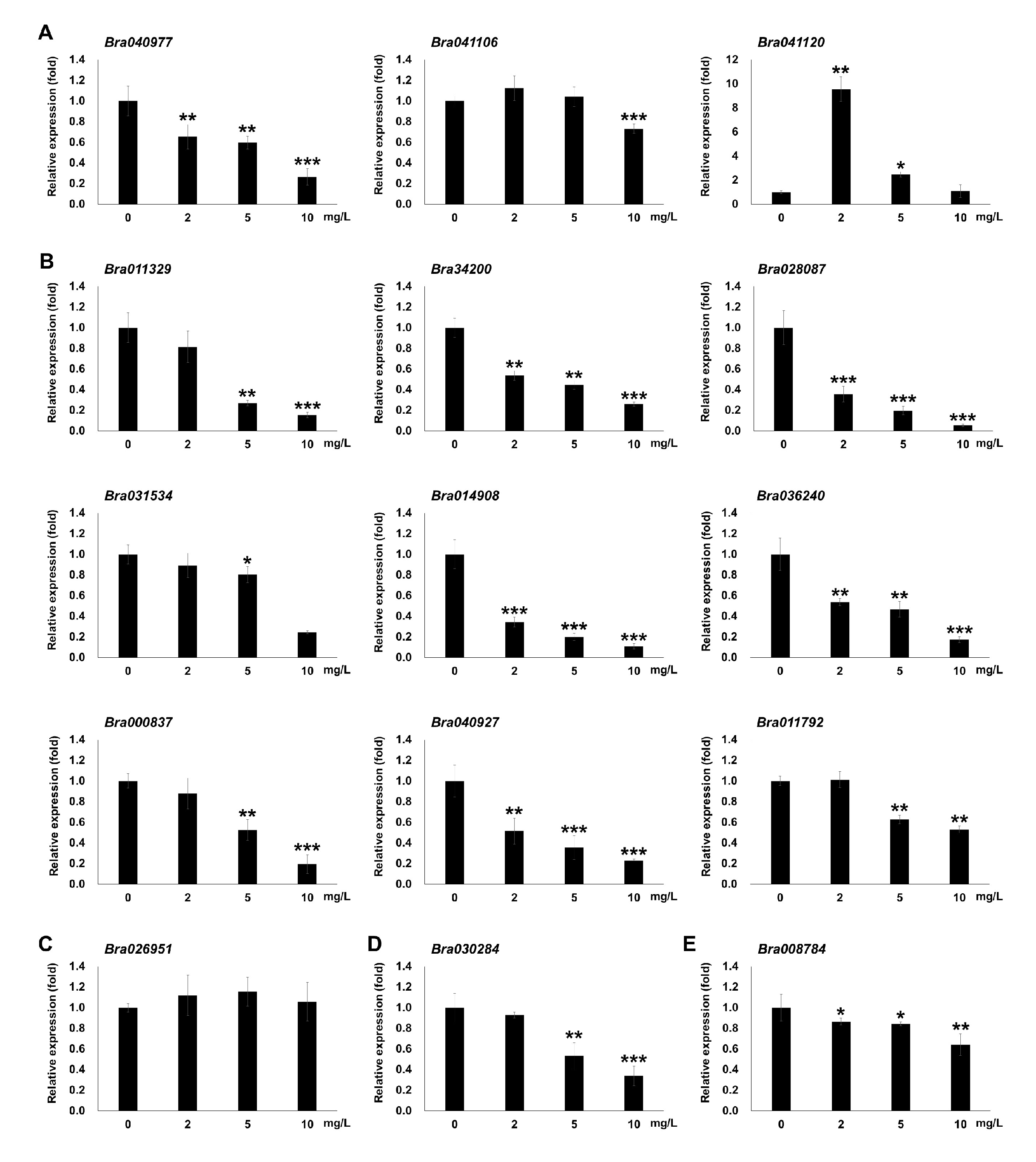
3.6. DAPs in Photosynthesis and qRT-PCR Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lv, J.; Deng, S.; Zhang, L. A review of artificial intelligence applications for antimicrobial resistance. Biosaf. Health 2020. [Google Scholar] [CrossRef]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezzariai, A.; Hafidi, M.; Khadra, A.; Aemig, Q.; El Fels, L.; Barret, M.; Merlina, G.; Patureau, D.; Pinelli, E. Human and veterinary antibiotics during composting of sludge or manure: Global perspectives on persistence, degradation, and resistance genes. J. Hazard. Mater. 2018, 359, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2014, 34, 473–492. [Google Scholar] [CrossRef] [Green Version]
- Boxall, A.B.A.; Fogg, L.A.; Blackwell, P.A.; Kay, P.; Pemberton, E.J.; Croxford, A. Veterinary medicines in the environment. Rev. Environ. Contam. Toxicol. 2004. [Google Scholar] [CrossRef]
- Ding, C.; He, J. Effect of antibiotics in the environment on microbial populations. Appl. Microbiol. Biotechnol. 2010, 87, 925–941. [Google Scholar] [CrossRef]
- Cheong, M.S.; Seo, K.H.; Chohra, H.; Yoon, Y.E.; Choe, H.; Kantharaj, V.; Lee, Y.B. Influence of sulfonamide contamination derived from veterinary antibiotics on plant growth and development. Antibiotics 2020, 9, 456. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K. Plant Environmental Stress Responses for Survival and Biomass Enhancement. Clim. Chang. Plant Abiotic Stress Toler. 2013, 79–108. [Google Scholar] [CrossRef]
- Pessarakli, M. Plant Responses under Environmental Stress Conditions. Adv. Plants Agric. Res. 2015, 2, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.; Finck, A.; Blair, G.; Tandon, H. Nutrient management guidelines for some major field crops. Plant Nutr. Food Secur. 2006, 235–348. [Google Scholar]
- Power, A.G. Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Misra, R.V.; Roy, R.N.; Hiraoka, H. On-Farm Composting Methods, Composting Methods and Techniques; UN-FAO: Rome, Italy, 2003; pp. 1–26. [Google Scholar]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef] [PubMed]
- Minden, V.; Deloy, A.; Volkert, A.M.; Leonhardt, S.D.; Pufal, G. Antibiotics impact plant traits, even at small concentrations. AoB Plants 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carla, E.; Francini, A.; Raffaelli, A.; Sebastiani, L. Science of the Total Environment Morpho-physiological response of Populus alba to erythromycin: A timeline of the health status of the plant. Sci. Total Environ. 2016, 569–570, 540–547. [Google Scholar] [CrossRef]
- Kura-hotta, M.; Satoh, K.; Katoh, S. Relationship between photosynthesis and chlorophyll content during leaf senescence of rice seedlings. Plant Cell Physiol. 1987, 28, 1321–1329. [Google Scholar] [CrossRef]
- Rochaix, J.D. Regulation of photosynthetic electron transport. Biochim. Biophys. Acta—Bioenerg. 2011, 1807, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef] [Green Version]
- Raines, C.A. The Calvin cycle revisited. Photosynth. Res. 2003, 75, 1–10. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Milo, R. The global mass and average rate of rubisco. Proc. Natl. Acad. Sci. USA 2019, 116, 4738–4743. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.C.; Sullivan, J.A.; Wang, J.H.; Jerome, C.A.; MacLean, D.; Allen, J.F.; Horner, D.S.; Howe, C.J.; Lopez-Juez, E.; Herrmann, R.G.; et al. Coordination of plastid and nuclear gene expression. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Barkan, A.; Goldschmidt-Clermont, M. Participation of nuclear genes in chloroplast gene expression. Biochimie 2000, 82, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Morales, F.; Ancín, M.; Fakhet, D.; González-Torralba, J.; Gámez, A.L.; Seminario, A.; Soba, D.; Ben Mariem, S.; Garriga, M.; Aranjuelo, I. Photosynthetic metabolism under stressful growth conditions as a bases for crop breeding and yield improvement. Plants 2020, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haak, D.C.; Fukao, T.; Grene, R.; Hua, Z.; Ivanov, R.; Perrella, G.; Li, S. Multilevel regulation of abiotic stress responses in plants. Front. Plant Sci. 2017, 8, 1564. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, S.; Liu, K.; Wang, S.; Huang, L.; Guo, L. Proteomics: A powerful tool to study plant responses to biotic stress. Plant Methods 2019, 15, 135. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant abiotic stress proteomics: The major factors determining alterations in cellular proteome. Front. Plant Sci. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shryock, T.R.; Mortensen, J.E.; Baumholtz, M. The effects of macrolides on the expression of bacterial virulence mechanisms. J. Antimicrob. Chemother. 1998, 41, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Usary, J.; Champney, W.S. Erythromycin inhibition of 50S ribosomal subunit formation in Escherichia coli cells. Mol. Microbiol. 2001, 40, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric Analysis of Chlorophylls and Carotenoids from Commonly Grown Fern Species by Using Various Extracting Solvents. Res. J. Chem. Sci. 2014, 2231, 606X. [Google Scholar]
- Wang, K.; Lin, K.; Huang, X.; Chen, M. A Simple and Fast Extraction Method for the Determination of Multiclass Antibiotics in Eggs Using LC-MS/MS. J. Agric. Food Chem. 2017, 65, 5064–5073. [Google Scholar] [CrossRef]
- Paulo, J.A. Sample preparation for proteomic analysis using a GeLC-MS/MS strategy. J. Biol. Methods 2016, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Schwacke, R.; Ponce-Soto, G.Y.; Krause, K.; Bolger, A.M.; Arsova, B.; Hallab, A.; Gruden, K.; Stitt, M.; Bolger, M.E.; Usadel, B. MapMan4: A Refined Protein Classification and Annotation Framework Applicable to Multi-Omics Data Analysis. Mol. Plant 2019, 12, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Gubernator, B.; Bartoszewski, R.; Kroliczewski, J.; Wildner, G.; Szczepaniak, A. Ribulose-1,5-bisphosphate carboxylase/oxygenase from thermophilic cyanobacterium Thermosynechococcus elongatus. Photosynth. Res. 2008, 95, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Lee, J. Leaf Proteome Analysis in Brassica rapa L. (Inbred line ‘Chiifu’) using Shotgun Proteome Approach. Plant Breed. Biotechnol. 2015, 3, 389–395. [Google Scholar] [CrossRef]
- Thompson, L.A.; Darwish, W.S. Environmental Chemical Contaminants in Food: Review of a Global Problem. J. Toxicol. 2019, 2019, 2345283. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.R.; Owens, G.; Kwon, S.I.; So, K.H.; Lee, D.B.; Ok, Y.S. Occurrence and environmental fate of veterinary antibiotics in the terrestrial environment. Water. Air Soil Pollut. 2011, 214, 163–174. [Google Scholar] [CrossRef]
- Hirsch, R.; Ternes, T.; Haberer, K.; Kratz, K.L. Occurrence of antibiotics in the aquatic environment. Sci. Total Environ. 1999, 225, 109–118. [Google Scholar] [CrossRef]
- Pan, M.; Wong, C.K.C.; Chu, L.M. Distribution of antibiotics in wastewater-irrigated soils and their accumulation in vegetable crops in the Pearl River Delta, Southern China. J. Agric. Food Chem. 2014, 62, 11062–11069. [Google Scholar] [CrossRef]
- Opris, O.; Copaciu, F.; Loredana Soran, M.; Ristoiu, D.; Niinemets, Ü.; Copolovici, L. Influence of nine antibiotics on key secondary metabolites and physiological characteristics in Triticum aestivum: Leaf volatiles as a promising new tool to assess toxicity. Ecotoxicol. Environ. Saf. 2013, 87, 70–79. [Google Scholar] [CrossRef]
- Liu, J.; Lu, G.; Ding, J.; Zhang, Z.; Wang, Y. Tissue distribution, bioconcentration, metabolism, and effects of erythromycin in crucian carp (Carassius auratus). Sci. Total Environ. 2014, 490, 914–920. [Google Scholar] [CrossRef]
- Liu, J.; Lu, G.; Cai, Y.; Wu, D.; Yan, Z.; Wang, Y. Modulation of erythromycin-induced biochemical responses in crucian carp by ketoconazole. Environ. Sci. Pollut. Res. 2017, 24, 5285–5292. [Google Scholar] [CrossRef]
- Wan, J.; Guo, P.; Peng, X.; Wen, K. Effect of erythromycin exposure on the growth, antioxidant system and photosynthesis of Microcystis flos-aquae. J. Hazard. Mater. 2015, 283, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Cundliffe, E. On the nature of antibiotic binding sites in ribosomes. Biochimie 1987, 69, 863–869. [Google Scholar] [CrossRef]
- Carol, P.; Rozier, C.; Lazaro, E.; Ballesta, J.P.G.; Mache, R. Erythromycin and 5S rRNA binding properties of the spinach chloroplast ribosomal protein CL22. Nucleic Acids Res. 1993, 21, 635–639. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.H.; Boynton, J.E.; Gillham, N.W. Chloroplast ribosomes and protein synthesis. Microbiol. Rev. 1994, 58, 700–754. [Google Scholar] [CrossRef] [PubMed]
- Mache, R. Chloroplast ribosomal proteins and their genes. Plant Sci. 1990, 72, 1–12. [Google Scholar] [CrossRef]
- Zoschke, R.; Bock, R. Chloroplast translation: Structural and functional organization, operational control, and regulation. Plant Cell 2018, 30, 745–770. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, M.-H.; Kang, Y.-H.; Lee, C.-H.; Chung, I.-K. Effects of Six Antibiotics on the Activity of the Photosynthetic Apparatus and Ammonium Uptake of Thallus of Porphyra yezoensis. Algae 2005, 20, 121–125. [Google Scholar] [CrossRef]
- Liu, B.Y.; Nie, X.P.; Liu, W.Q.; Snoeijs, P.; Guan, C.; Tsui, M.T. Toxic effects of erythromycin, ciprofloxacin and sulfamethoxazole on photosynthetic apparatus in Selenastrum capricornutum. Ecotoxicol. Environ. Saf. 2011, 74, 1027–1035. [Google Scholar] [CrossRef]
- Deng, C.N.; Zhang, D.Y.; Pan, X.L. Toxic effects of erythromycin on photosystem I and II in Microcystis aeruginosa. Photosynthetica 2014, 52, 574–580. [Google Scholar] [CrossRef]
- Liu, B.; Liu, W.; Nie, X.; Guan, C.; Yang, Y.; Wang, Z.; Liao, W. Growth response and toxic effects of three antibiotics on Selenastrum capricornutum evaluated by photosynthetic rate and chlorophyll biosynthesis. J. Environ. Sci. 2011, 23, 1558–1563. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Roháček, K. Chlorophyll Fluorescence Parameters. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Sandmann, G.; Böger, P. Inhibition of photosynthetic electron transport by amphotericin B. Physiol. Plant. 1981, 51, 326–328. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, D.; Chen, X.; Mu, G.; Li, L.; Bao, A. Effects of levofloxacin hydrochlordie on photosystem II activity and heterogeneity of Synechocystis sp. Chemosphere 2009, 77, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Quick, W.P.; Fichtner, K.; Schulze, E.D.; Wendler, R.; Leegood, R.C.; Mooney, H.; Rodermel, S.R.; Bogorad, L.; Stitt, M. Decreased ribulose-1,5-bisphosphate carboxylase-oxygenase in transgenic tobacco transformed with “antisense” rbcS—IV. Impact on photosynthesis in conditions of altered nitrogen supply. Planta 1992, 188, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Fichtner, K.; Quick, W.P.; Schulze, E.-D.; Mooney, H.A.; Rodermel, S.R.; Bogorad, L.; Stitt, M. Decreased ribulose-1,5-bisphosphate carboxylase-oxygenase in transgenic tobacco transformed with antisense rbcS. Planta 1993, 190, 1–9. [Google Scholar] [CrossRef]
- Jiang, C.Z.; Rodermel, S.R. Regulation of photosynthesis during leaf development in RbcS antisense DNA mutants of tobacco. Plant Physiol. 1995, 107, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Goh, E.B.; Yim, G.; Tsui, W.; McClure, J.A.; Surette, M.G.; Davies, J. Transcriptional modulation of bacterial gene expression by subinhibitory concentrations of antibiotics. Proc. Natl. Acad. Sci. USA 2002, 99, 17025–17030. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Molecular Weight | Quantitative Value (Normalized Total Spectra) | Brassica Accession | AGI Locus Identifier | Description | |
|---|---|---|---|---|---|---|
| E0 | E5 | |||||
| M4EHZ1 | 20 kDa | 456.8 | 0.0 | Bra028406 | AT5G38410 | Ribulose bisphosphate carboxylase small chain |
| M4EYY5 | 20 kDa | 205.0 | 0.0 | Bra034027 | AT1G67090 | Ribulose bisphosphate carboxylase small chain |
| M4EPE0 | 28 kDa | 81.5 | 0.0 | Bra030660 | AT1G06680 | PsbP domain-containing protein |
| M4DG97 | 28 kDa | 62.4 | 0.0 | Bra015520 | AT1G06680 | PsbP domain-containing protein |
| A0A249RRH7 | 56 kDa | 33.9 | 0.0 | Bra040977 | ATCG00680 | Photosystem II CP47 reaction center protein |
| A0A249RQW1 | 39 kDa | 25.1 | 0.0 | ATCG00020 | Photosystem II protein D1 (psbA) | |
| M4FBB1 | 26 kDa | 22.4 | 0.0 | Bra038377 | AT4G09650 | Uncharacterized protein |
| M4C9F8 | 26 kDa | 22.4 | 0.0 | Bra000837 | AT4G03280 | Plastoquinol–plastocyanin reductase |
| M4DNQ7 | 28 kDa | 18.3 | 0.0 | Bra018144 | AT3G47470 | Chlorophyll a–b binding protein, chloroplastic |
| M4EV39 | 23 kDa | 15.6 | 0.0 | Bra032672 | AT4G12800 | PSI subunit V |
| A0A249RPW6 | 9 kDa | 15.6 | 0.0 | Bra041107 | ATCG00580 | Cytochrome b559 subunit alpha (psbE) |
| A0A249RQ23 | 9 kDa | 14.3 | 0.0 | ATCG01060 | Photosystem I iron–sulfur center (psaC) | |
| A0A249RPV2 | 52 kDa | 12.9 | 0.0 | Bra041123 | ATCG00280 | Photosystem II CP43 reaction center protein (psbC) |
| M4DD65 | 28 kDa | 12.2 | 0.0 | Bra014433 | AT3G61470 | Chlorophyll a–b binding protein, chloroplastic |
| M4F5A2 | 24 kDa | 8.8 | 0.0 | Bra036258 | AT4G02530 | Uncharacterized protein |
| M4CLA2 | 30 kDa | 8.1 | 0.0 | Bra004989 | AT2G40100 | Chlorophyll a–b binding protein, chloroplastic |
| A0A249RRE6 | 40 kDa | 6.8 | 0.0 | ATCG00270 | Photosystem II D2 protein (psbD) | |
| M4D4C5 | 24 kDa | 6.8 | 0.0 | Bra011329 | AT4G32260 | Uncharacterized protein |
| M4CFL7 | 29 kDa | 4.8 | 0.0 | Bra002999 | AT5G54270 | Chlorophyll a–b binding protein, chloroplastic |
| A0A249RPW8 | 82 kDa | 3.4 | 0.0 | Bra041122 | ATCG00350 | Photosystem I P700 chlorophyll a apoprotein A2 (psaB) |
| M4E904 (+2) | 32 kDa | 2.0 | 0.0 | Bra025260 | AT3G27240 | Cytochrome c domain-containing protein |
| M4C8N9 | 26 kDa | 1.4 | 0.0 | Bra000567 | AT3G63540 | PsbP domain-containing protein |
| M4F584 | 31 kDa | 33.3 | 3.8 | Bra036240 | AT4G02770 | Uncharacterized protein |
| M4EZF7 | 25 kDa | 25.1 | 5.7 | Bra034200 | AT4G03280 | Plastoquinol–plastocyanin reductase |
| M4C909 | 25 kDa | 29.9 | 7.6 | Bra000687 | AT4G09650 | Uncharacterized protein |
| M4ELR3 | 31 kDa | 26.5 | 11.4 | Bra029732 | AT3G08940 | Chlorophyll a–b binding protein, chloroplastic |
| M4DEI7 | 24 kDa | 38.7 | 13.3 | Bra014908 | AT1G31330 | PSI-F |
| M4E725 | 53 kDa | 48.2 | 17.1 | Bra024580 | AT1G23310 | Aminotran_1_2 domain-containing protein |
| M4FIJ8 | 16 kDa | 5.4 | 19.0 | Bra040927 | AT2G28900 | Uncharacterized protein |
| M4EDU1 | 65 kDa | 4.1 | 19.0 | Bra026951 | AT1G12840 | V-type proton ATPase subunit C |
| A0A249RPX1 | 35 kDa | 59.7 | 26.6 | Bra041106 | ATCG00540 | Cytochrome f (petA) |
| M4ERV7 | 28 kDa | 82.1 | 38.0 | Bra031534 | AT1G06680 | PsbP domain-containing protein |
| M4D5N5 | 73 kDa | 26.5 | 53.2 | Bra011792 | AT4G37870 | Phosphoenolpyruvate carboxykinase (ATP) |
| A0A249RRD6 | 55 kDa | 292.5 | 146.2 | Bra041120 | ATCG00120 | ATP synthase subunit alpha, chloroplastic |
| M4EH22 | 53 kDa | 2700.1 | 1086.0 | Bra028087 | ATCG00490 | Ribulose bisphosphate carboxylase large chain (rbcL) |
| Protein ID | Molecular Weight | Quantitative Value (Normalized Total Spectra) | Brassica Accession | AGI Locus Identifier | Description | |
|---|---|---|---|---|---|---|
| E0 | E5 | |||||
| M4D1Y3 | 32 kDa | 9.5 | 0.0 | Bra010483 | AT5G50250 | Uncharacterized protein |
| M4CNF7 | 15 kDa | 8.8 | 0.0 | Bra005745 | AT5G02960 | 40S ribosomal protein S23 |
| M4EQE3 | 36 kDa | 8.8 | 0.0 | Bra031014 | AT1G18080 | WD_REPEATS_REGION domain-containing protein |
| M4D7B0 | 17 kDa | 7.5 | 0.0 | Bra012370 | AT1G23290 | Ribosomal_L18e/L15P domain-containing protein |
| M4DFM6 | 17 kDa | 7.5 | 0.0 | Bra015299 | AT1G04270 | Structural constituent of ribosome |
| M4CHU3 | 16 kDa | 6.8 | 0.0 | Bra003776 | AT1G74970 | Structural constituent of ribosome |
| M4DZD2 | 32 kDa | 6.8 | 0.0 | Bra021879 | AT2G33800 | S5 DRBM domain-containing protein |
| M4E880 | 45 kDa | 6.8 | 0.0 | Bra024986 | AT1G43170 | Structural constituent of ribosome |
| A0A249RQ06 | 11 kDa | 6.8 | 0.0 | Bra001922 | AT5G47320 | 30S ribosomal protein S19, chloroplastic (rps19) |
| A0A249RPV7 | 23 kDa | 6.8 | 0.0 | Bra027599 | AT5G45250 | 30S ribosomal protein S4, chloroplastic (rps4) |
| M4D384 | 26 kDa | 4.8 | 0.0 | Bra010937 | AT1G27450 | Adenine phosphoribosyltransferase |
| M4C7V7 | 29 kDa | 4.1 | 0.0 | Bra000285 | AT2G43030 | Ribosomal protein L3 family protein |
| M4DCB8 | 19 kDa | 4.1 | 0.0 | Bra014131 | AT1G48350 | Ribosomal protein L18 family protein |
| M4DJ77 | 26 kDa | 4.1 | 0.0 | Bra016555 | AT1G18540 | Ribosomal_L6e_N domain-containing protein |
| M4CPV9 | 19 kDa | 3.4 | 0.0 | Bra006248 | AT5G14320 | 30S ribosomal protein S13, chloroplast |
| A0A249RPV9 | 27 kDa | 3.4 | 0.0 | Bra013947 | AT4G26090 | 30S ribosomal protein S2, chloroplastic (rps2) |
| A0A249RR23 | 30 kDa | 3.4 | 0.0 | ATCG00830 | 50S ribosomal protein L2, chloroplastic (rpl2) | |
| M4DDG7 | 13 kDa | 2.7 | 0.0 | Bra014535 | AT2G43460 | 60S ribosomal protein L38 |
| M4F6Q3 | 22 kDa | 2.7 | 0.0 | Bra036763 | AT1G35680 | 50S ribosomal protein L21, chloroplastic |
| M4CX77 | 24 kDa | 2.0 | 0.0 | Bra008824 | AT5G13510 | Ribosomal protein L10 family protein |
| A0A249RRI7 | 14 kDa | 2.0 | 0.0 | Bra040980 | ATCG00780 | 50S ribosomal protein L14, chloroplastic (rpl14) |
| M4DXP8 | 36 kDa | 1.4 | 0.0 | Bra021294 | AT3G18130 | WD_REPEATS_REGION domain-containing protein |
| A0A249RQJ6 | 18 kDa | 1.4 | 0.0 | ATCG00810 | 50S ribosomal protein L22, chloroplastic (rpl22) | |
| M4ENY9 | 25 kDa | 0.0 | 3.8 | Bra030509 | AT1G02780 | Ribosomal protein L19 |
| M4EZX0 | 11 kDa | 1.4 | 5.7 | Bra034363 | AT2G27710 | 60S acidic ribosomal protein P2 |
| M4D4S2 | 6 kDa | 0.0 | 5.7 | Bra011477 | AT3G44010 | 40S ribosomal protein S29 |
| M4CVX6 | 27 kDa | 2.0 | 7.6 | Bra008373 | AT1G78630 | Structural constituent of ribosome |
| M4D857 | 24 kDa | 3.4 | 9.5 | Bra012667 | AT4G16720 | Ribosomal protein L15 |
| M4DLG0 | 30 kDa | 3.4 | 9.5 | Bra017341 | AT3G25920 | Ribosomal_L18e/L15P domain-containing protein |
| M4CCC7 | 16 kDa | 0.0 | 9.5 | Bra001857 | AT4G15000 | 60S ribosomal protein L27 |
| M4CHH9 | 12 kDa | 4.8 | 11.4 | Bra003662 | AT1G77940 | Ribosomal_L7Ae domain-containing protein |
| M4EMA8 | 18 kDa | 5.4 | 13.3 | Bra029928 | AT5G23740 | Ribosomal_S17_N domain-containing protein |
| M4FD44 | 16 kDa | 0.0 | 13.3 | Bra039014 | AT2G19730 | Ribosomal_L28e domain-containing protein |
| M4CIF0 | 21 kDa | 7.5 | 15.2 | Bra003984 | AT1G66580 | Ribosomal_L16 domain-containing protein |
| M4DVY7 | 24 kDa | 6.8 | 15.2 | Bra020681 | AT5G48760 | 60S ribosomal protein L13A |
| M4C811 | 84 kDa | 5.4 | 15.2 | Bra000339 | AT2G44060 | 6,7-dimethyl-8-ribityllumazine synthase |
| M4D8V6 | 17 kDa | 3.4 | 15.2 | Bra012916 | AT3G49910 | KOW domain-containing protein |
| M4DU07 | 19 kDa | 3.4 | 15.2 | Bra020000 | AT1G09590 | 60S ribosomal protein L21 |
| M4DW28 | 10 kDa | 3.4 | 15.2 | Bra020722 | AT3G61110 | 40S ribosomal protein S27 |
| M4EE11 | 85 kDa | 0.0 | 15.2 | Bra027021 | AT1G62750 | Elongation factor G, chloroplastic |
| M4E3F1 | 23 kDa | 2.7 | 17.1 | Bra023302 | AT1G32990 | Structural constituent of ribosome |
| M4DQ40 | 25 kDa | 0.0 | 17.1 | Bra018631 | AT2G27530 | Ribosomal protein |
| M4DWX7 | 16 kDa | 8.8 | 19.0 | Bra012616 | AT4G18100 | 60S ribosomal protein L32 |
| M4DVR1 | 47 kDa | 8.8 | 19.0 | Bra020605 | AT5G28020 | Cysteine synthase |
| M4CUN4 | 16 kDa | 6.8 | 19.0 | Bra007929 | AT1G70600 | Ribosomal_L18e/L15P domain-containing protein |
| M4CFK1 | 13 kDa | 4.8 | 19.0 | Bra002983 | AT5G04800 | 40S ribosomal protein S17 |
| M4EBR8 | 73 kDa | 10.2 | 22.8 | Bra026227 | AT1G30580 | Obg-like ATPase 1 |
| M4DND0 | 24 kDa | 9.5 | 22.8 | Bra018017 | AT3G49010 | 60S ribosomal protein L13 |
| M4CAC5 | 14 kDa | 12.2 | 24.7 | Bra001154 | AT3G05560 | 60S ribosomal protein L22-2 |
| M4D389 | 23 kDa | 12.2 | 24.7 | Bra010943 | AT1G27400 | 60S ribosomal protein L17 |
| M4CQ00 | 23 kDa | 8.8 | 24.7 | Bra006289 | AT5G15200 | S4 RNA-binding domain-containing protein |
| M4FGB7 | 17 kDa | 8.1 | 28.5 | Bra040145 | AT5G18380 | 40S ribosomal protein S16 |
| M4C7F7 | 17 kDa | 5.4 | 28.5 | Bra000135 | AT2G39460 | Ribosomal_L23eN domain-containing protein |
| M4D1R2 | 16 kDa | 8.1 | 30.4 | Bra010412 | AT4G27090 | Ribosomal_L14e domain-containing protein |
| M4EBD3 | 13 kDa | 7.5 | 30.4 | Bra026092 | AT2G32060 | 40S ribosomal protein S12 |
| M4FH70 | 37 kDa | 13.6 | 32.3 | Bra040448 | AT3G63490 | Ribosomal protein |
| M4D5W4 | 12 kDa | 12.9 | 32.3 | Bra010696 | AT4G39200 | 40S ribosomal protein S25 |
| M4CGU4 | 17 kDa | 17.0 | 36.1 | Bra003427 | AT4G00100 | Ribosomal_S13_N domain-containing protein |
| M4D7X0 | 21 kDa | 15.6 | 38.0 | Bra012580 | AT4G18730 | Structural constituent of ribosome |
| M4EHJ8 | 14 kDa | 10.9 | 38.0 | Bra028263 | AT5G62300 | Ribosomal_S10 domain-containing protein |
| M4FGG9 | 105 kDa | 10.9 | 39.9 | Bra040197 | AT3G04380 | Histone-lysine N-methyltransferase |
| P51423 | 15 kDa | 17.6 | 41.8 | Bra029570 | AT4G02890 | Ubiquitin-60S ribosomal protein L40 (RL40_BRARP) |
| M4DL77 | 37 kDa | 12.2 | 41.8 | Bra017258 | AT2G36145 | Uncharacterized protein |
| M4CS06 | 18 kDa | 20.4 | 45.6 | Bra006998 | AT3G53430 | 60S ribosomal protein L12 |
| M4DIF5 | 17 kDa | 12.2 | 53.2 | Bra016282 | AT1G26630 | Eukaryotic translation initiation factor 5A |
| M4D178 | 28 kDa | 21.0 | 57.0 | Bra010227 | AT4G31700 | 40S ribosomal protein S6 |
| M4EGW3 | 25 kDa | 27.8 | 62.7 | Bra028028 | AT1G34030 | 40S ribosomal protein S18 |
| M4EZ44 | 38 kDa | 30.5 | 64.6 | Bra034087 | AT3G09820 | Adenosine kinase |
| M4CUY5 | 32 kDa | 29.2 | 64.6 | Bra008030 | AT1G72370 | 40S ribosomal protein SA |
| M4DLT0 | 28 kDa | 31.2 | 77.8 | Bra017461 | AT2G01250 | 60S ribosomal protein L7 |
| M4ELW0 | 45 kDa | 34.6 | 79.7 | Bra029780 | AT3G09630 | Ribos_L4_asso_C domain-containing protein |
| M4CEM1 | 236 kDa | 30.5 | 85.4 | Bra002652 | AT5G58410 | RING-type E3 ubiquitin transferase |
| M4D611 | 65 kDa | 48.2 | 163.3 | Bra011919 | AT1G55490 | ATP binding/protein binding |
| M4EPY7 | 74 kDa | 46.8 | 172.8 | Bra030858 | AT1G55490 | ATP binding/protein binding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, Y.-E.; Cho, H.M.; Bae, D.-w.; Lee, S.J.; Choe, H.; Kim, M.C.; Cheong, M.S.; Lee, Y.B. Erythromycin Treatment of Brassica campestris Seedlings Impacts the Photosynthetic and Protein Synthesis Pathways. Life 2020, 10, 311. https://doi.org/10.3390/life10120311
Yoon Y-E, Cho HM, Bae D-w, Lee SJ, Choe H, Kim MC, Cheong MS, Lee YB. Erythromycin Treatment of Brassica campestris Seedlings Impacts the Photosynthetic and Protein Synthesis Pathways. Life. 2020; 10(12):311. https://doi.org/10.3390/life10120311
Chicago/Turabian StyleYoon, Young-Eun, Hyun Min Cho, Dong-won Bae, Sung Joong Lee, Hyeonji Choe, Min Chul Kim, Mi Sun Cheong, and Yong Bok Lee. 2020. "Erythromycin Treatment of Brassica campestris Seedlings Impacts the Photosynthetic and Protein Synthesis Pathways" Life 10, no. 12: 311. https://doi.org/10.3390/life10120311