Telomere Instability in Lynch Syndrome Families Leads to Some Shorter Telomeres in MSH2+/- Carriers

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Processing and Telomere Length Measurements
2.2. Statistical Analyses
3. Results
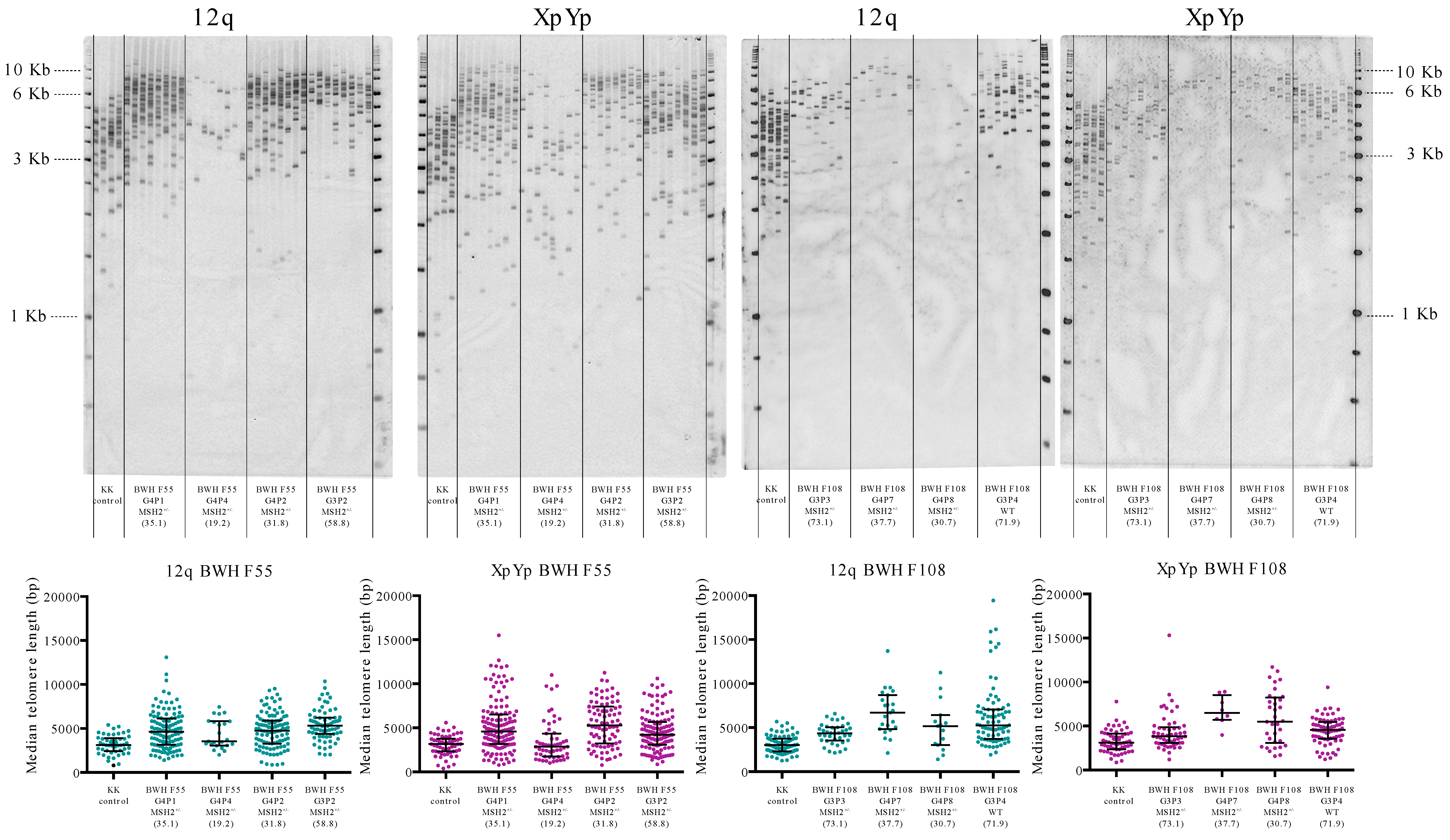
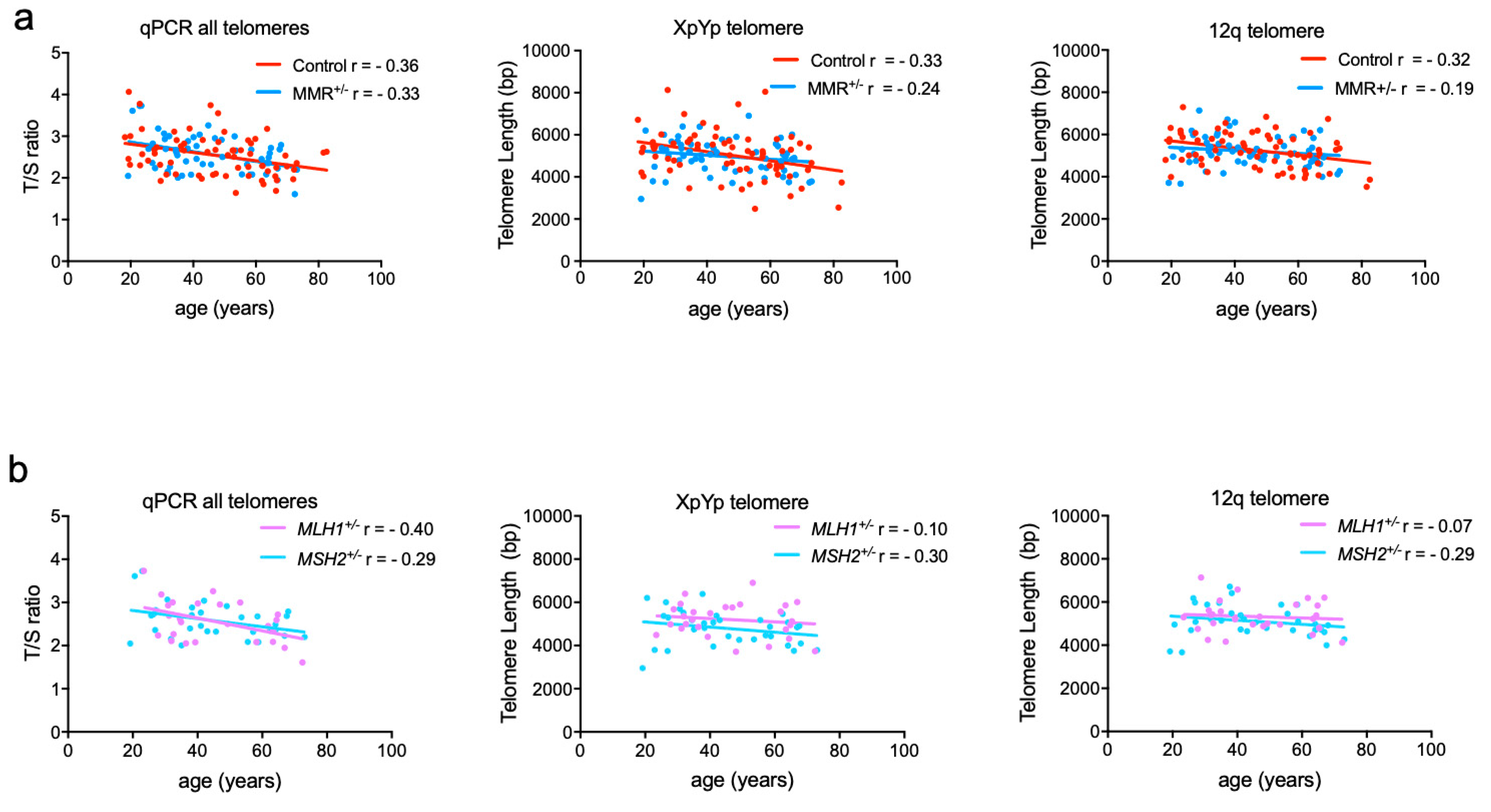
3.1. Cross-sectional Analysis of Total Telomeric DNA and Individual Telomere Lengths in LS and Controls
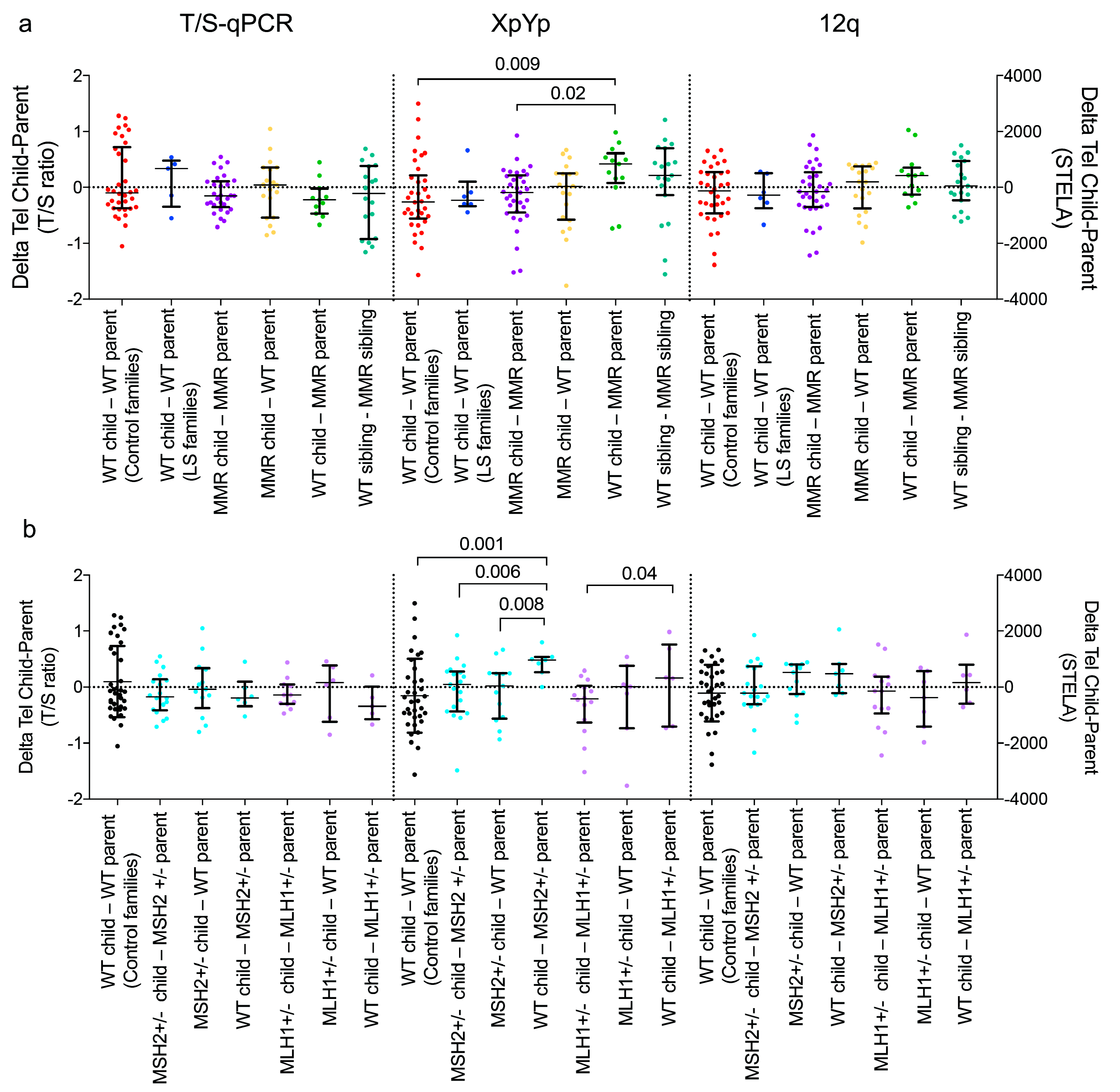
3.2. Comparison of Age-adjusted of Telomere Lengths within Families
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stanley, S.E.; Armanios, M. The short and long telomere syndromes: Paired paradigms for molecular medicine. Curr. Opin. Genet. Dev. 2015, 33, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opresko, P.L.; Shay, J.W. Telomere-associated aging disorders. Ageing Res. Rev. 2017, 33, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Maciejowski, J.; de Lange, T. Telomeres in cancer: Tumour suppression and genome instability. Nat. Rev. Mol. Cell Biol. 2017, 18, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, H.T.; Lynch, P.M.; Lanspa, S.J.; Snyder, C.L.; Lynch, J.F.; Boland, C.R. Review of the Lynch syndrome: History, molecular genetics, screening, differential diagnosis, and medicolegal ramifications. Clin. Genet. 2009, 76, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Alazzouzi, H.; Domingo, E.; Gonzalez, S.; Blanco, I.; Armengol, M.; Espin, E.; Plaja, A.; Schwartz, S.; Capella, G.; Schwartz, S., Jr. Low levels of microsatellite instability characterize MLH1 and MSH2 HNPCC carriers before tumor diagnosis. Hum. Mol. Genet. 2005, 14, 235–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coolbaugh-Murphy, M.I.; Xu, J.P.; Ramagli, L.S.; Ramagli, B.C.; Brown, B.W.; Lynch, P.M.; Hamilton, S.R.; Frazier, M.L.; Siciliano, M.J. Microsatellite instability in the peripheral blood leukocytes of HNPCC patients. Hum. Mutat. 2010, 31, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Kansikas, M.; Kasela, M.; Kantelinen, J.; Nystrom, M. Assessing how reduced expression levels of the mismatch repair genes MLH1, MSH2, and MSH6 affect repair efficiency. Hum. Mutat. 2014, 35, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Baird, D.M.; Jeffreys, A.J.; Royle, N.J. Mechanisms underlying telomere repeat turnover, revealed by hypervariable variant repeat distribution patterns in the human Xp/Yp telomere. EMBO J. 1995, 14, 5433–5443. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellite mutations in the germline: Implications for evolutionary inference. Trends Genet. 2000, 16, 551–558. [Google Scholar] [CrossRef]
- de Lange, T. Shelterin-Mediated Telomere Protection. Annu. Rev. Genet. 2018, 52, 223–247. [Google Scholar] [CrossRef]
- Pickett, H.A.; Baird, D.M.; Hoff-Olsen, P.; Meling, G.I.; Rognum, T.O.; Shaw, J.; West, K.P.; Royle, N.J. Telomere instability detected in sporadic colon cancers, some showing mutations in a mismatch repair gene. Oncogene 2004, 23, 3434–3443. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Bermudez, A.; Hills, M.; Pickett, H.A.; Phan, A.T.; Mergny, J.L.; Riou, J.F.; Royle, N.J. Human telomeres that contain (CTAGGG)n repeats show replication dependent instability in somatic cells and the male germline. Nucleic Acids Res. 2009, 37, 6225–6238. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Bermudez, A.; Royle, N.J. Deficiency in DNA mismatch repair increases the rate of telomere shortening in normal human cells. Hum. Mutat. 2011, 32, 939–946. [Google Scholar] [CrossRef]
- Ponti, G.; Ruini, C.; Tomasi, A. Mismatch repair gene deficiency and genetic anticipation in Lynch syndrome: Myth or reality? Dis. Colon Rectum 2015, 58, 141–142. [Google Scholar] [CrossRef] [PubMed]
- von Salome, J.; Boonstra, P.S.; Karimi, M.; Silander, G.; Stenmark-Askmalm, M.; Gebre-Medhin, S.; Aravidis, C.; Nilbert, M.; Lindblom, A.; Lagerstedt-Robinson, K. Genetic anticipation in Swedish Lynch syndrome families. PLoS Genet. 2017, 13, e1007012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ten Broeke, S.W.; Rodriguez-Girondo, M.; Suerink, M.; Aretz, S.; Bernstein, I.; Capella, G.; Engel, C.; Gomez-Garcia, E.B.; van Hest, L.P.; von Knebel Doeberitz, M.; et al. The Apparent Genetic Anticipation in PMS2-Associated Lynch Syndrome Families Is Explained by Birth-cohort Effect. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1010–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vulliamy, T.; Marrone, A.; Szydlo, R.; Walne, A.; Mason, P.J.; Dokal, I. Disease anticipation is associated with progressive telomere shortening in families with dyskeratosis congenita due to mutations in TERC. Nat. Genet. 2004, 36, 447–449. [Google Scholar] [CrossRef] [Green Version]
- Holohan, B.; Wright, W.E.; Shay, J.W. Cell biology of disease: Telomeropathies: An emerging spectrum disorder. J. Cell Biol. 2014, 205, 289–299. [Google Scholar] [CrossRef]
- Segui, N.; Pineda, M.; Guino, E.; Borras, E.; Navarro, M.; Bellido, F.; Moreno, V.; Lazaro, C.; Blanco, I.; Capella, G.; et al. Telomere length and genetic anticipation in Lynch syndrome. PLoS ONE 2013, 8, e61286. [Google Scholar] [CrossRef] [Green Version]
- Jeyapalan, J.N.; Mendez-Bermudez, A.; Zaffaroni, N.; Dubrova, Y.E.; Royle, N.J. Evidence for alternative lengthening of telomeres in liposarcomas in the absence of ALT-associated PML bodies. Int. J. Cancer 2008, 122, 2414–2421. [Google Scholar] [CrossRef]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of seven loci affecting mean telomere length and their association with disease. Nat. Genet. 2013, 45, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Lai, T.P.; Wright, W.E.; Shay, J.W. Comparison of telomere length measurement methods. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Kimura, M.; Kim, S.; Cao, X.; Srinivasan, S.R.; Berenson, G.S.; Kark, J.D.; Aviv, A. Longitudinal versus cross-sectional evaluations of leukocyte telomere length dynamics: Age-dependent telomere shortening is the rule. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniali, L.; Benetos, A.; Susser, E.; Kark, J.D.; Labat, C.; Kimura, M.; Desai, K.; Granick, M.; Aviv, A. Telomeres shorten at equivalent rates in somatic tissues of adults. Nat. Commun. 2013, 4, 1597. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.; Baird, D.M.; Royle, N.J. The plasticity of human telomeres demonstrated by a hypervariable telomere repeat array that is located on some copies of 16p and 16q. Hum. Mol. Genet. 1999, 8, 1637–1646. [Google Scholar] [CrossRef] [Green Version]
- Baird, D.M.; Coleman, J.; Rosser, Z.H.; Royle, N.J. High levels of sequence polymorphism and linkage disequilibrium at the telomere of 12q: Implications for telomere biology and human evolution. Am. J. Hum. Genet. 2000, 66, 235–250. [Google Scholar] [CrossRef] [Green Version]
- Nersisyan, L.; Hopp, L.; Loeffler-Wirth, H.; Galle, J.; Loeffler, M.; Arakelyan, A.; Binder, H. Telomere Length Maintenance and Its Transcriptional Regulation in Lynch Syndrome and Sporadic Colorectal Carcinoma. Front. Oncol. 2019, 9, 1172. [Google Scholar] [CrossRef] [Green Version]
- Aubert, G.; Hills, M.; Lansdorp, P.M. Telomere length measurement-caveats and a critical assessment of the available technologies and tools. Mutat. Res. 2012, 730, 59–67. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| LS Families (N = 24) | Control Families (N = 10) | |||||||
|---|---|---|---|---|---|---|---|---|
| MMR Mutation-Carriers (N = 60) | Non-Carriers (N = 31) | Controls (N = 37) | ||||||
| Cancer | Cancer-free | Cancer | Cancer-free | Total 91 | Cancer | Cancer-free | Total 37 | |
| n | 32 | 28 | 2 | 29 | 3 | 34 | ||
| Parent: N (%) | 24 (75) | b 3 (11) | 2 (100) | c 15 (52) | 44 (48) | 3 (100) | 15 (44) | 18 (49) |
| Children: N (%) | 8 (25) | 25 (89) | 0 (0) | 14 (48) | 47 (52) | 0 (0) | 19 (56) | 19 (51) |
| MSH2+/−: N (%) | 15 (47) | 18 (64) | - | - | 33 (36) | - | - | |
| MLH1+/−: N (%) | 17 (53) | 10 (36) | - | - | 27 (30) | - | - | |
| a MMR+/+: N (%) | - | - | 2 (100) | 29 (100) | 31 (34) | 3 (100) | 34 (100) | 37 (100) |
| Sex: N (%) | F: 14 (44) | F: 17 (61) | F: 2 (100) | F: 16 (55) | F: 49 (54) | F: 2 (67) | F: 21 (62) | 23 (62) |
| M: 18 (56) | M: 11 (39) | M: 0 (0) | M: 13 (45) | M: 42 (46) | M: 1 (33) | M: 13 (38) | 14 (38) | |
| Mean age at sample ±SD parents | 60 ± 8 | 66 ± 2 | 65 ± 11 | 63 ± 7 | 62 ± 8 | 56 ± 3 | 62 ± 10 | 61 ± 10 |
| Mean age at sample ±SD children | 35 ± 8 | 33 ± 7 | - | 35 ± 8 | 34 ± 7 | - | 31 ± 11 | 31 ± 11 |
| Mean age 1st cancer ±SD parents | 45 ± 11 | - | 59 ± 9 | - | - | 44 ± 8 | - | - |
| Mean age 1st cancer ±SD children | 24 ± 7 | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido-Navas, M.C.; Tippins, F.; Barwell, J.; Hoffman, J.; Codd, V.; Royle, N.J. Telomere Instability in Lynch Syndrome Families Leads to Some Shorter Telomeres in MSH2+/- Carriers. Life 2020, 10, 265. https://doi.org/10.3390/life10110265
Garrido-Navas MC, Tippins F, Barwell J, Hoffman J, Codd V, Royle NJ. Telomere Instability in Lynch Syndrome Families Leads to Some Shorter Telomeres in MSH2+/- Carriers. Life. 2020; 10(11):265. https://doi.org/10.3390/life10110265
Chicago/Turabian StyleGarrido-Navas, M. Carmen, Frances Tippins, Julian Barwell, Jonathan Hoffman, Veryan Codd, and Nicola J. Royle. 2020. "Telomere Instability in Lynch Syndrome Families Leads to Some Shorter Telomeres in MSH2+/- Carriers" Life 10, no. 11: 265. https://doi.org/10.3390/life10110265