Extracellular Alpha-Synuclein Promotes a Neuroinhibitory Secretory Phenotype in Astrocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Methods
2.1. Immunohistochemistry
2.2. Cell Culture and Immunocytochemistry
2.3. Confocal Microscopy
2.4. Western Blotting
2.5. ELISA Assays
2.6. Statistical Analysis
3. Results
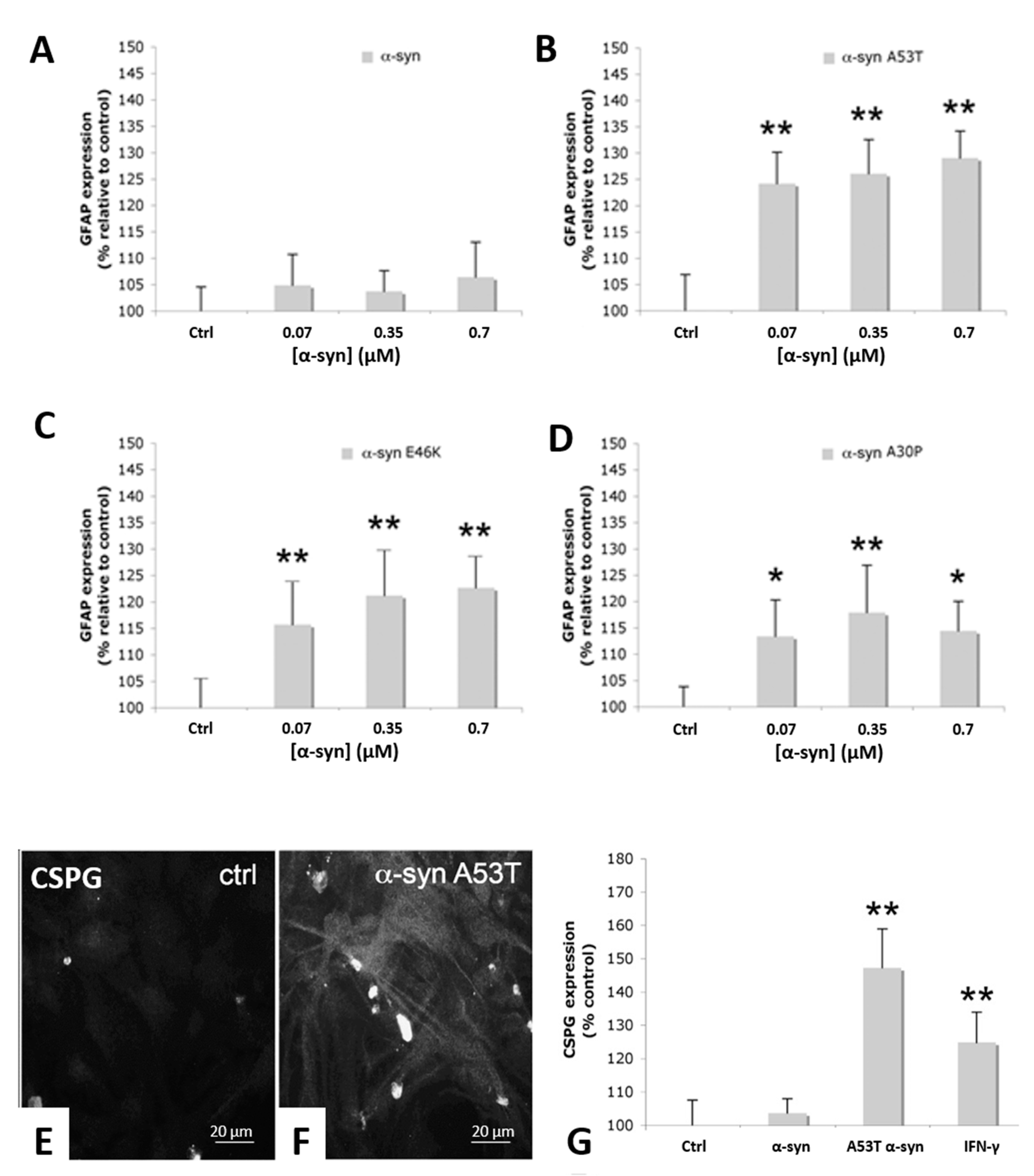
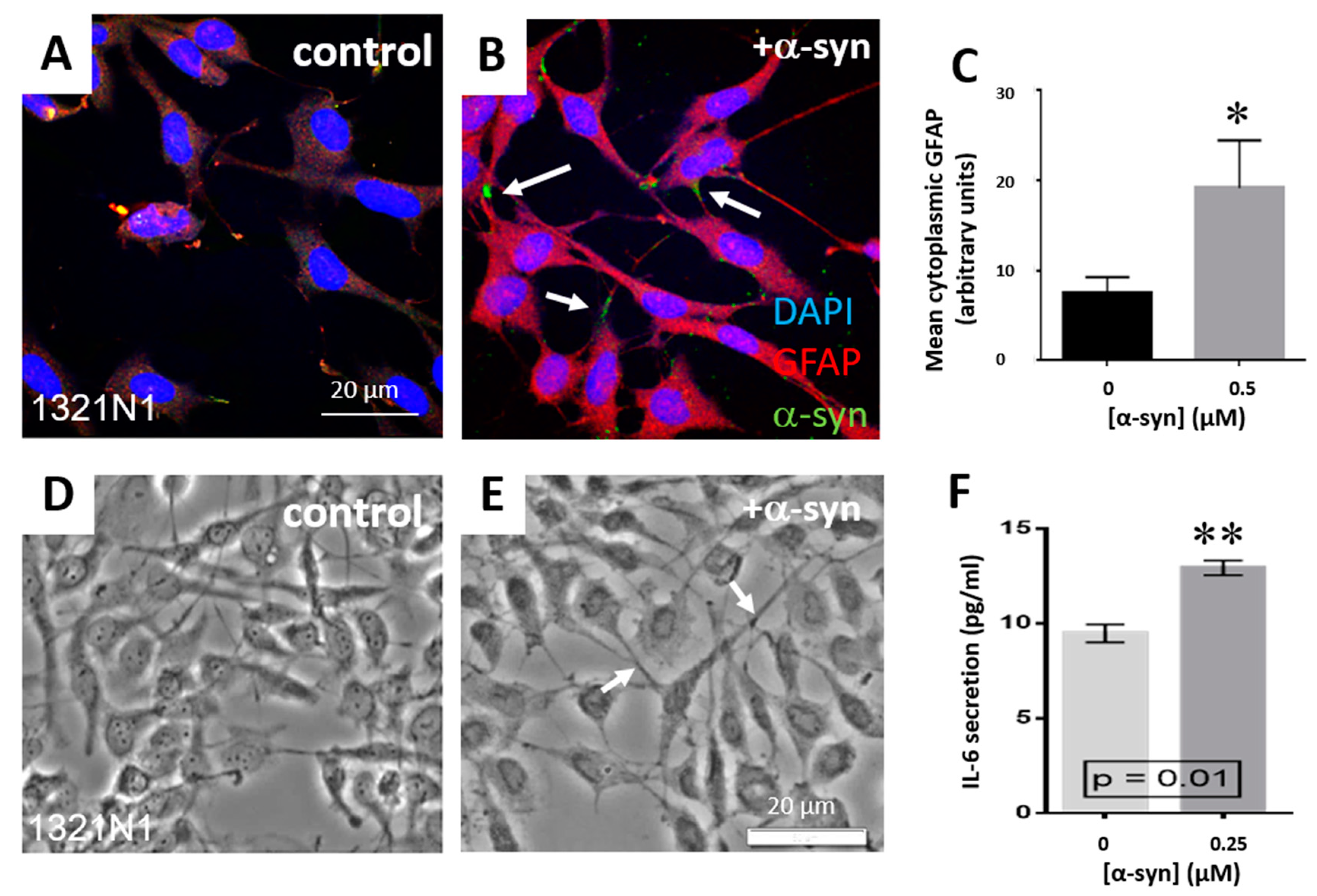
3.1. α-Syn Treatment Promotes Increased Astrocyte Expression of Glial Fibrillary Acidic Protein and Chondroitin-Sulphate Proteoglycans and IL-6 Secretion
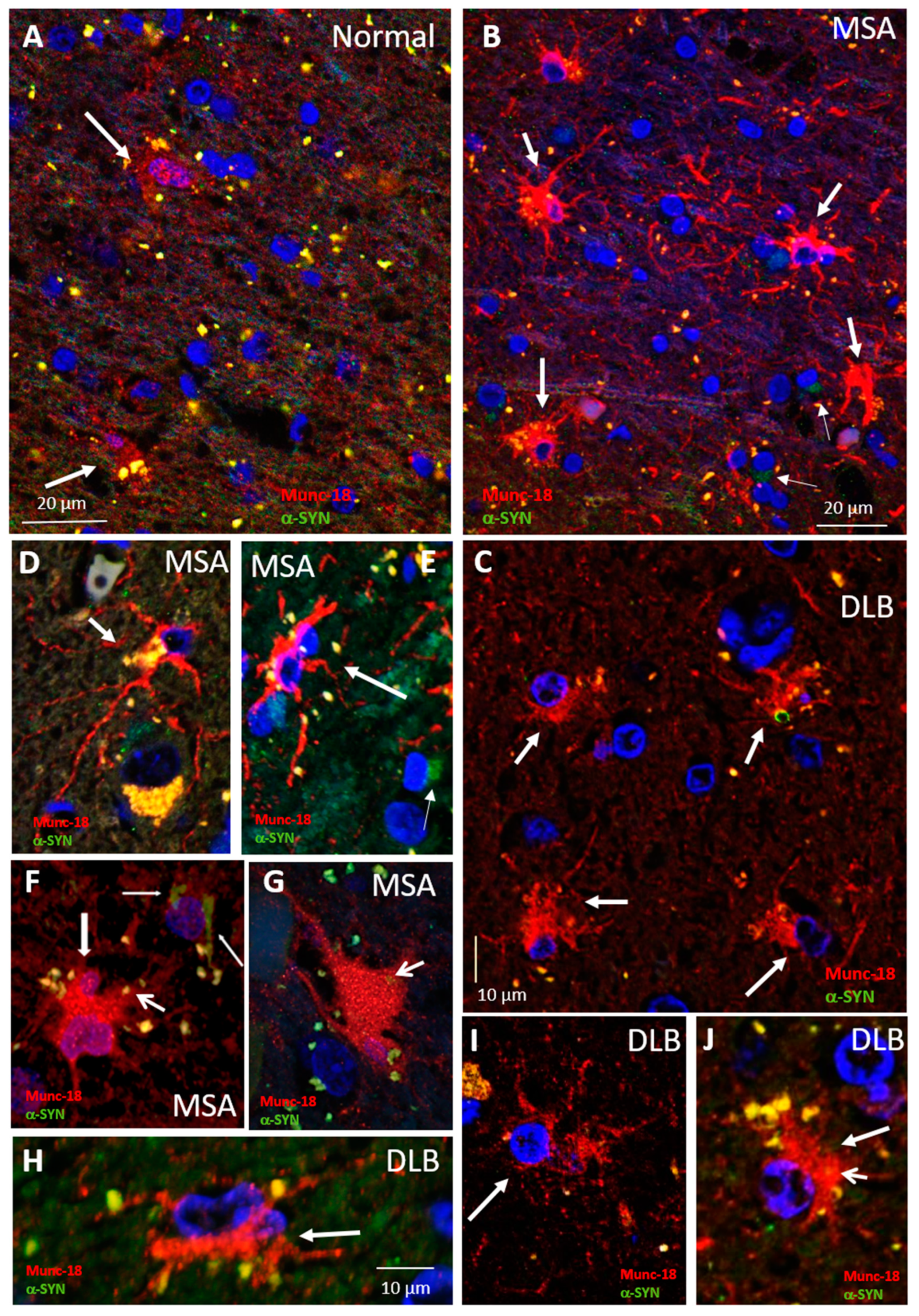
3.2. Munc18-Positive Vesicles Are Increased in MSA and DLB Astrocytes
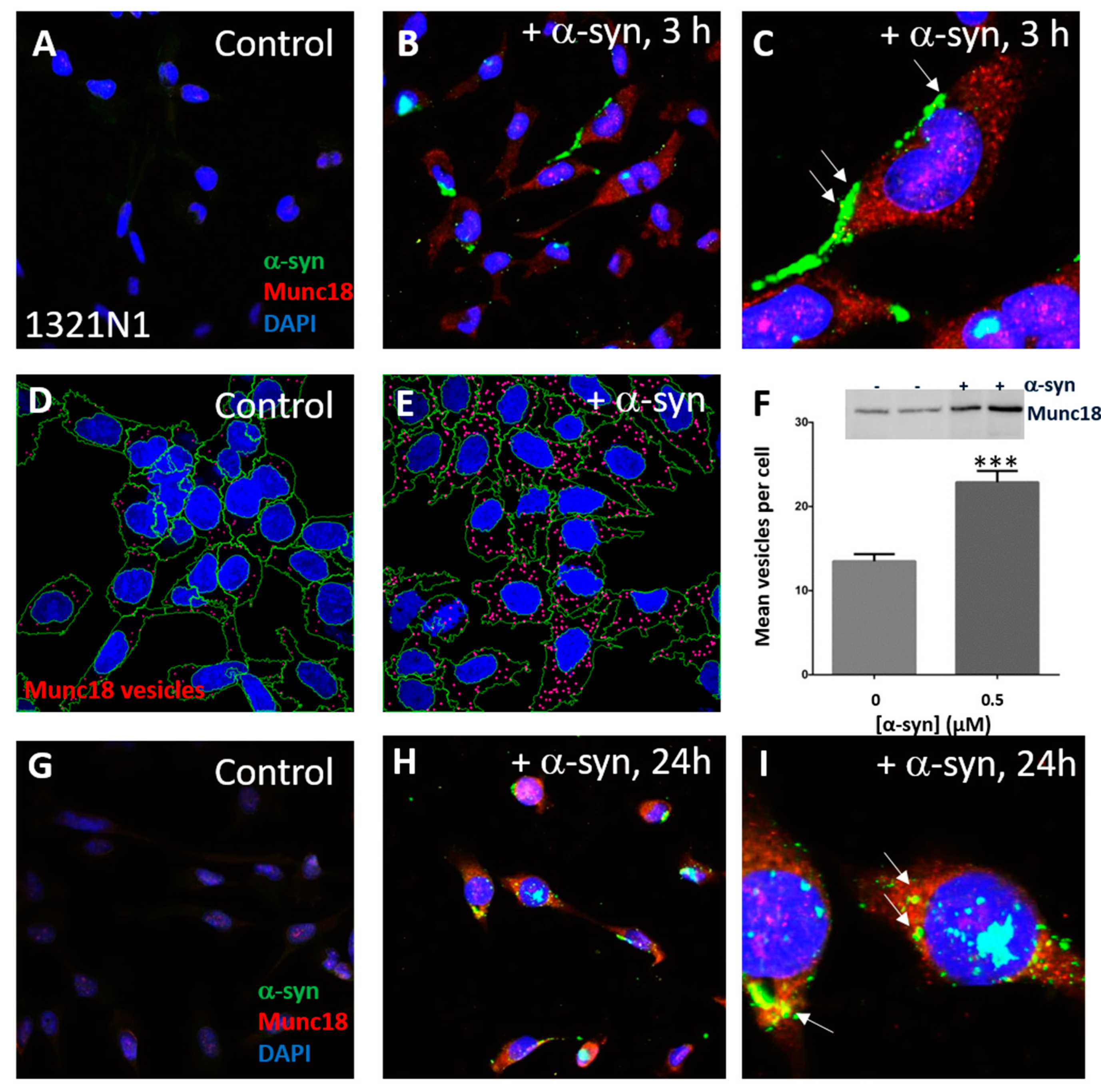
3.3. α-Syn Treatment Promotes Astroglial Accumulation of Munc18-Positive Vesicles
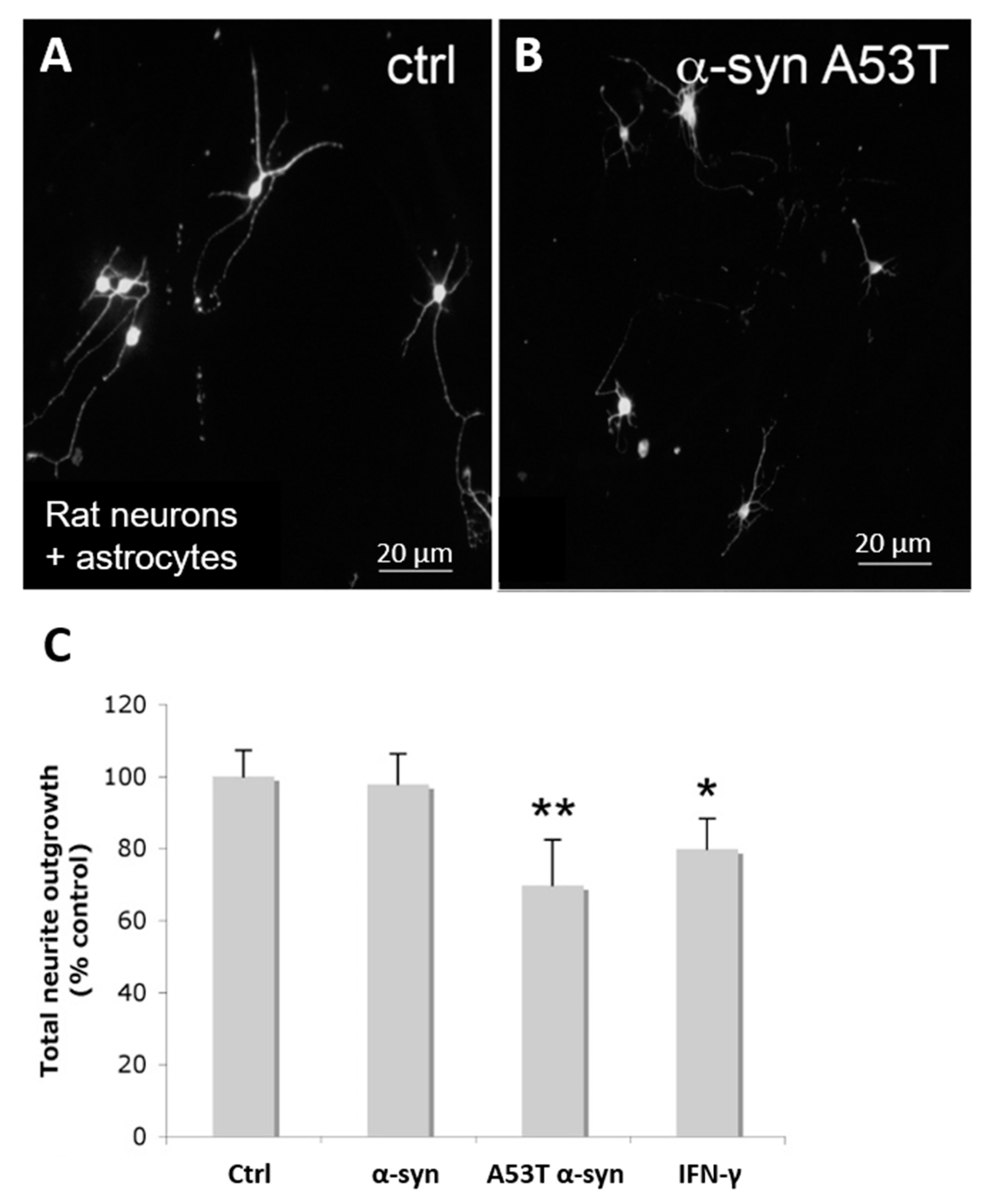
3.4. α-Syn Treatment Results in a Neuroinhibitory Astroglial Phenotype
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| α-syn | Alpha-synuclein |
| CSPG | Chondroitin-sulphate proteoglycan |
| JNK | c-Jun N-terminal kinases |
| DLB | Dementia with Lewy bodies |
| GCI | Glial cytoplasmic inclusions |
| GFAP | Glial fibrillary acidic protein |
| iNOS | inducible NO synthase |
| Interferon gamma | IFNγ |
| IL-6 | Interleukin-6 |
| MSA | Multiple system atrophy |
| NF-κB | Nuclear factor of kappa light polypeptide gene enhancer in B-cells |
| P38 MAPK | p38 mitogen-activated protein kinase |
| PD | Parkinson’s disease |
| SNAP | Soluble NSF Attachment Protein |
| SNARE | SNAP receptor |
| TLR | Toll-like receptor |
References
- Zaccai, J.; McCracken, C.; Brayne, C. A systematic review of prevalence and incidence studies of dementia with Lewy bodies. Age Ageing 2005, 34, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Braak, H.; Tredici, K.D.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Radford, R.; Wong, M.; Pountney, D.L. Neurodegenerative Aspects of Multiple System Atrophy. In Handbook of Neurotoxicity; Springer: New York, NY, USA, 2014; pp. 2157–2180. [Google Scholar]
- Gazulla, J.; Berciano, J.; Peyronnet, B.; Manunta, A.; Gamé, X.; Fanciulli, A.; Wenning, G.; Babu, D.S. Multiple-System Atrophy. N. Engl. J. Med. 2015, 372, 249–263. [Google Scholar]
- Jellinger, K.A. Formation and development of Lewy pathology: A critical update. J. Neurol. 2009, 256, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N. Neuropathology, biochemistry, and biophysics of α-synuclein aggregation. J. Neurochem. 2007, 103, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Trojanowski, J.Q.; Lee, V.M.Y. Parkinson’s Disease and Related α-Synucleinopathies Are Brain Amyloidoses. Ann. N. Y. Acad. Sci. 2003, 991, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.F.; Rey, N.L.; Bousset, L.; Melki, R.; Brundin, P.; Angot, E. α-synuclein transfers from neurons to oligodendrocytes. Glia 2014, 62, 387–398. [Google Scholar] [CrossRef]
- Hansen, C.; Angot, E.; Bergström, A.-L.; Steiner, J.A.; Pieri, L.; Paul, G.; Outeiro, T.F.; Melki, R.; Kallunki, P.; Fog, K.; et al. α-Synuclein propagates from mouse brain to grafted dopaminergic neurons and seeds aggregation in cultured human cells. J. Clin. Investig. 2011, 121, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Kordower, J.H. Lewy body pathology in fetal grafts. Ann. N. Y. Acad. Sci. 2009, 1184, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Visanji, N.P.; Brooks, P.L.; Hazrati, L.-N.; Lang, A. The prion hypothesis in Parkinson’s disease: Braak to the future. Acta Neuropathol. Commun. 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Vieira, B.D.M.; Radford, R.A.; Chung, R.S.; Guillemin, G.J.; Pountney, D.L. Neuroinflammation in Multiple System Atrophy: Response to and Cause of α-Synuclein Aggregation. Front. Cell. Neurosci. 2015, 9, 437. [Google Scholar] [CrossRef] [PubMed]
- Valdinocci, D.; Radford, R.; Goulding, M.; Hayashi, J.; Chung, R.S.; Pountney, D.L. Extracellular Interactions of Alpha-Synuclein in Multiple System Atrophy. Int. J. Mol. Sci. 2018, 19, 4129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostenbach, S.; Cambron, M.; D’Haeseleer, M.; Kooijman, R.; De Keyser, J. Astrocyte loss and astrogliosis in neuroinflammatory disorders. Neurosci. Lett. 2014, 565, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Ho, D.H.; Suk, J.-E.; You, S.; Michael, S.; Kang, J.; Lee, S.J.; Masliah, E.; Hwang, D.; Lee, H.-J.; et al. Neuron-released oligomeric α-synuclein is an endogenous agonist of TLR2 for paracrine activation of microglia. Nat. Commun. 2013, 4, 1562. [Google Scholar] [CrossRef] [Green Version]
- Fellner, L.; Irschick, R.B.; Schanda, K.; Reindl, M.; Klimaschewski, L.; Poewe, W.; Wenning, G.; Stefanova, N. Toll-like receptor 4 is required for α-synuclein dependent activation of microglia and astroglia. Glia 2013, 61, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.K.; Jha, N.K.; Kar, R.; Ambasta, R.K.; Kumar, P. p38 MAPK and PI3K/AKT Signalling Cascades in Parkinson’s Disease. Int. J. Mol. Cell Med. 2015, 4, 67–86. [Google Scholar]
- Teismann, P.; Tieu, K.; Choi, D.-K.; Wu, D.-C.; Naini, A.; Hunot, S.; Przedborski, S. Cyclooxygenase-2 is instrumental in Parkinson’s disease neurodegeneration. Proc. Natl. Acad. Sci. USA 2003, 100, 5473–5478. [Google Scholar] [CrossRef] [Green Version]
- Klegeris, A.; Giasson, B.I.; Zhang, H.; Maguire, J.; Pelech, S.; McGeer, P.L. Alpha-synuclein and its disease-causing mutants induce ICAM-1 and IL-6 in human astrocytes and astrocytoma cells. FASEB 2006, 20, 2000–2008. [Google Scholar] [CrossRef]
- Lee, H.-J.; Suk, J.-E.; Patrick, C.; Bae, E.-J.; Cho, J.-H.; Rho, S.; Hwang, D.; Masliah, E.; Lee, S.-J. Direct Transfer of α-Synuclein from Neuron to Astroglia Causes Inflammatory Responses in Synucleinopathies. J. Boil. Chem. 2010, 285, 9262–9272. [Google Scholar] [CrossRef] [Green Version]
- Escartin, C.; Rouach, N. Astroglial networking contributes to neurometabolic coupling. Front. Neuroenergetics 2013, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Rodriguez, J.J.; Parpura, V. Neurotransmitters and Integration in Neuronal-Astroglial Networks. Neurochem. Res. 2012, 37, 2326–2338. [Google Scholar] [CrossRef] [PubMed]
- Petzold, G.C.; Murthy, V.N. Role of Astrocytes in Neurovascular Coupling. Neuron 2011, 71, 782–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belanger, M.; Allaman, I.; Magistretti, P.J. Brain Energy Metabolism: Focus on Astrocyte-Neuron Metabolic Cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Farina, C.; Aloisi, F.; Meinl, E. Astrocytes are active players in cerebral innate immunity. Trends Immunol. 2007, 28, 138–145. [Google Scholar] [CrossRef]
- Brown, A.M.; Ransom, B.R. Astrocyte glycogen and brain energy metabolism. Glia 2007, 55, 1263–1271. [Google Scholar] [CrossRef]
- Burgoyne, R.D.; Morgan, A. Membrane Trafficking: Three Steps to Fusion. Curr. Boil. 2007, 17, R255–R258. [Google Scholar] [CrossRef] [Green Version]
- Rizo, J.; Südhof, T.C. Snares and munc18 in synaptic vesicle fusion. Nat. Rev. Neurosci. 2002, 3, 641–653. [Google Scholar] [CrossRef]
- Jahn, R.; Fasshauer, D. Molecular machines governing exocytosis of synaptic vesicles. Nature 2012, 490, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Brandhorst, D.; Zwilling, D.; Rizzoli, S.O.; Lippert, U.; Lang, T.; Jahn, R. Homotypic Fusion of Early Endosomes: SNAREs Do Not Determine Fusion Specificity. Proc. Natl. Acad. Sci. USA 2006, 103, 2701–2706. [Google Scholar] [CrossRef] [Green Version]
- Toonen, R.J.; Kochubey, O.; De Wit, H.; Gulyas-Kovacs, A.; Konijnenburg, B.; Sørensen, J.B.; Klingauf, J.; Verhage, M. Dissecting docking and tethering of secretory vesicles at the target membrane. EMBO J. 2006, 25, 3725–3737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toonen, R.J.; Verhage, M. Munc18-1 in secretion: Lonely Munc joins SNARE team and takes control. Trends Neurosci. 2007, 30, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Zilly, F.E.; Sørensen, J.B.; Jahn, R.; Lang, T. Munc18-bound syntaxin readily forms SNARE complexes with synaptobrevin in native plasma membranes. PLoS Boil. 2006, 4, e330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radford, R.; Rcom-H’Cheo-Gauthier, A.; Wong, M.; Eaton, E.D.; Quilty, M.; Blizzard, C.A.; Norazit, A.; Meedeniya, A.; Vickers, J.C.; Gai, W.P.; et al. The degree of astrocyte activation in multiple system atrophy is inversely proportional to the distance to α-synuclein inclusions. Mol. Cell. Neurosci. 2015, 65, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Honma, S.; Nakahata, N.; Kobayashi, H.; Ikeda, S.; Takeda, N.; Ohizumi, Y. Decrease in thromboxane A2 receptor expression by differentiation with dibutyryl cyclic AMP in 1321N1 human astrocytoma cells. Prostaglandins Other Lipid Mediat. 1999, 58, 51–62. [Google Scholar] [CrossRef]
- Goodwin, J.; Nath, S.; Engelborghs, Y.; Pountney, D.L. Raised calcium and oxidative stress cooperatively promote alpha-synuclein aggregate formation. Neurochem. Int. 2013, 62, 703–711. [Google Scholar] [CrossRef]
- Leung, Y.K.; Pankhurst, M.W.; Dunlop, S.A.; Ray, S.; Dittmann, J.; Eaton, E.; Palumaa, P.; Sillard, R.; Chuah, M.; West, A.; et al. Metallothionein induces a regenerative reactive astrocyte phenotype via JAK/STAT and RhoA signalling pathways. Exp. Neurol. 2010, 221, 98–106. [Google Scholar] [CrossRef]
- Yiu, G.; He, Z. Glial inhibition of CNS axon regeneration. Nat. Rev. Neurosci. 2006, 7, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Rcom-H’Cheo-Gauthier, A.; Meedeniya, A.C.B.; Pountney, D.L. Calcipotriol inhibits α-synuclein aggregation in SH-SY5Y neuroblastoma cells by a Calbindin-D28k-dependent mechanism. J. Neurochem. 2017, 141, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Emmanouilidou, E.; Melachroinou, K.; Roumeliotis, T.; Garbis, S.D.; Ntzouni, M.; Margaritis, L.H.; Stefanis, L.; Vekrellis, K. Cell-Produced -Synuclein Is Secreted in a Calcium-Dependent Manner by Exosomes and Impacts Neuronal Survival. J. Neurosci. 2010, 30, 6838–6851. [Google Scholar] [CrossRef] [Green Version]
- Fellner, L.; Stefanova, N. The Role of Glia in Alpha-Synucleinopathies. Mol. Neurobiol. 2013, 47, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Han, G.A.; Malintan, N.T.; Collins, B.M.; Meunier, F.A.; Sugita, S. Munc18-1 as a key regulator of neurosecretion. J. Neurochem. 2010, 115, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, N.J.; Benveniste, E.N. Interleukin-6 expression and regulation in astrocytes. J. Neuroimmunol. 1999, 100, 124–139. [Google Scholar] [CrossRef]
- Campbell, I.L.; Abraham, C.; Masliah, E.; Kemper, P.; Inglis, J.D.; Oldstone, M.B.; Mucke, L. Neurologic disease induced in transgenic mice by cerebral overexpression of interleukin 6. Proc. Natl. Acad. Sci. USA 1993, 90, 10061–10065. [Google Scholar] [CrossRef] [Green Version]
- Woiciechowsky, C.; Schöning, B.; Stoltenburg-Didinger, G.; Stockhammer, F.; Volk, H.D. Brain-IL-1β triggers astrogliosis through induction of IL-6: Inhibition by propranolol and IL-10. Med. Sci. Monit. 2004, 10, 330. [Google Scholar]
- Chiang, C.-S.; Stalder, A.; Samimi, A.; Campbell, I.L. Reactive Gliosis as a Consequence of Interleukin-6 Expression in the Brain: Studies in Transgenic Mice. Dev. Neurosci. 1994, 16, 212–221. [Google Scholar] [CrossRef]
- Eulenfeld, R.; Dittrich, A.; Khouri, C.; Müller, P.J.; Mütze, B.; Wolf, A.; Schaper, F. Interleukin-6 signalling: More than Jaks and STATs. Eur. J. Cell Boil. 2012, 91, 486–495. [Google Scholar] [CrossRef]
- Yamasaki, R.; Yamaguchi, H.; Matsushita, T.; Fujii, T.; Hiwatashi, A.; Kira, J. Early strong intrathecal inflammation in cerebellar type multiple system atrophy by cerebrospinal fluidcytokine/chemokine profiles: A case control study. J. Neuroinflamm. 2017, 14, 89. [Google Scholar] [CrossRef] [Green Version]
- Soubannier, V.; Maussion, G.; Chaineau, M.; Sigutova, V.; Rouleau, G.; Durcan, T.M.; Stifani, S. Characterization of human iPSC-derived astrocytes with potential for disease modeling and drug discovery. Neurosci. Lett. 2020, 731, 135028. [Google Scholar] [CrossRef]
- Yun, S.P.; Kam, T.-I.; Panicker, N.; Kim, S.; Oh, Y.; Park, J.-S.; Kwon, S.-H.; Park, Y.J.; Karuppagounder, S.S.; Park, H.; et al. Block of A1 astrocyte conversion by microglia is neuroprotective in models of Parkinson’s disease. Nat. Med. 2018, 24, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Muqit, M.M. Parkinson’s: A Disease of Aberrant Vesicle Trafficking. Annu. Rev. Cell Dev. Boil. 2020, 36. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Marco Vieira, B.; Radford, R.A.W.; Hayashi, J.; Eaton, E.D.; Greenaway, B.; Jambas, M.; Petcu, E.B.; Chung, R.S.; Pountney, D.L. Extracellular Alpha-Synuclein Promotes a Neuroinhibitory Secretory Phenotype in Astrocytes. Life 2020, 10, 183. https://doi.org/10.3390/life10090183
Di Marco Vieira B, Radford RAW, Hayashi J, Eaton ED, Greenaway B, Jambas M, Petcu EB, Chung RS, Pountney DL. Extracellular Alpha-Synuclein Promotes a Neuroinhibitory Secretory Phenotype in Astrocytes. Life. 2020; 10(9):183. https://doi.org/10.3390/life10090183
Chicago/Turabian StyleDi Marco Vieira, Bruno, Rowan A. W. Radford, Junna Hayashi, Emma D. Eaton, Ben Greenaway, Mark Jambas, Eugen B. Petcu, Roger S. Chung, and Dean L. Pountney. 2020. "Extracellular Alpha-Synuclein Promotes a Neuroinhibitory Secretory Phenotype in Astrocytes" Life 10, no. 9: 183. https://doi.org/10.3390/life10090183