
3′-UTR Polymorphisms in Thymidylate Synthase with Colorectal Cancer Prevalence and Prognosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Subjects
2.3. Phenotype Measurements
2.4. Genotyping
2.5. Quantitative Reverse Transcription-PCR
2.6. Statistical Analysis
3. Results
3.1. Patient Characteristics
3.2. Genotype Frequencies of TS 3′-UTR Variants
3.3. Effects of Combined Genotypes and Allelic Gene-Gene Interactions for TS 3′-UTR Variants on CRC Incident Rates
3.4. Stratified Effects of TS 3′-UTR Polymorphisms on CRC Incidence
3.5. Gene-Environment Combined Effects of TS 3′-UTR Polymorphisms on CRC Incidence
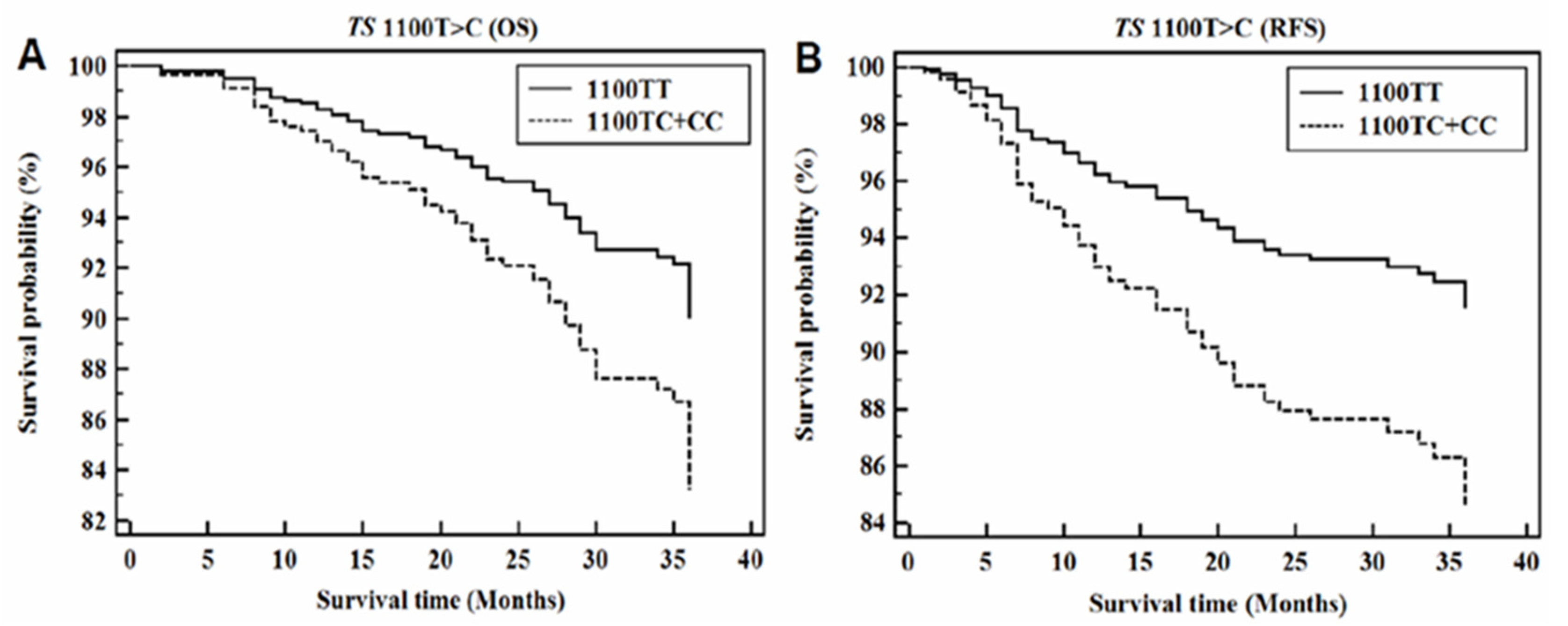
3.6. CRC Progression According to TS 3′-UTR Polymorphisms
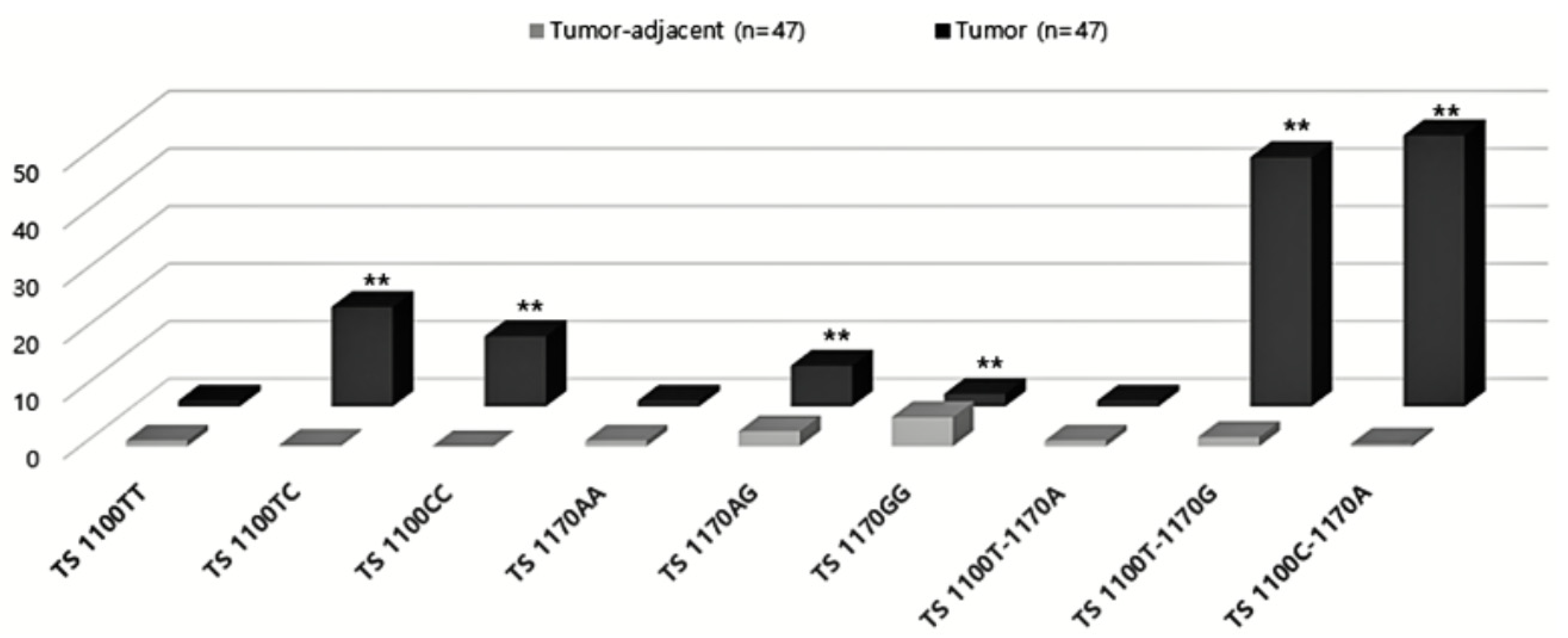
3.7. Expression of TS 3′-UTR Polymorphisms
3.8. Expression of Target miRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Incidence and Mortality Worldwide IARC CancerBase No. 11; International Agency for Research on Cancer: Lyon, France, 2018.
- Boyle, P.; Levin, B. Colorectal cancer. In World Cancer Report; International Agency for Research on Cancer: Lyon, France, 2008; pp. 374–378. [Google Scholar]
- Giovannucci, E.; Ascherio, A.; Rimm, E.B.; Colditz, G.; Stampfer, M.J.; Willett, W.C. Physical Activity, Obesity, and Risk for Colon Cancer and Adenoma in Men. Ann. Intern. Med. 1995, 122, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and Colon Cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef]
- Tsoi, K.K.; Pau, C.Y.; Wu, W.K.; Chan, F.K.; Griffiths, S.; Sung, J.J. Cigarette Smoking and the Risk of Colorectal Cancer: A Meta-analysis of Prospective Cohort Studies. Clin. Gastroenterol. Hepatol. 2009, 7, 682–688.e5. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Konno, M.; Koseki, J.; Taniguchi, M.; Vecchione, A.; Ishii, H. One-carbon metabolism for cancer diagnostic and therapeutic approaches. Cancer Lett. 2020, 470, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Omura, K.; Kanehira, E.; Watanabe, Y. Polymorphic tandem repeats in the thymidylate synthase gene is associated with its protein expression in human gastrointestinal cancers. Anticancer. Res. 2000, 19, 3249–3252. [Google Scholar]
- Mandola, M.V.; Stoehlmacher, J.; Zhang, W.; Groshen, S.; Yu, M.C.; Iqbal, S.; Lenz, H.-J.; Ladner, R.D. A 6 bp polymorphism in the thymidylate synthase gene causes message instability and is associated with decreased intratumoral TS mRNA levels. Pharmacogenetics 2004, 14, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Papamichael, D. The use of thymidylate synthase inhibitors in the treatment of advanced colorectal cancer: Current status. Oncology 1999, 4, 478–487. [Google Scholar]
- Berger, F.G.; Berger, S.H. Thymidylate synthase as a chemotherapeutic drug target: Where are we after fifty years? Cancer Biol. Ther. 2006, 5, 1238–1241. [Google Scholar] [CrossRef] [PubMed]
- Leichman, L.; Lenz, H.-J.; Leichman, C.; Groshen, S.; Danenberg, K.; Baranda, J.; Spears, C.; Boswell, W.; Silberman, H.; Ortega, A.; et al. Quantitation of intratumoral thymidylate synthase expression predicts for resistance to protracted infusion of 5-fluorouracil and weekly leucovorin in disseminated colorectal cancers: Preliminary report from an ongoing trial. Eur. J. Cancer 1995, 31, 1306–1310. [Google Scholar] [CrossRef]
- Johnston, P.G.; Fisher, E.R.; Rockette, H.E.; Fisher, B.; Wolmark, N.; Drake, J.C.; Chabner, B.A.; Allegra, C.J. The role of thymidylate synthase expression in prognosis and outcome of adjuvant chemotherapy in patients with rectal cancer. J. Clin. Oncol. 1994, 12, 2640–2647. [Google Scholar] [CrossRef]
- Lenz, H.J.; Hayashi, K.; Salonga, D.; Danenberg, K.D.; Danenberg, P.V.; Metzger, R.; Banerjee, D.; Bertino, J.R.; Groshen, S.; Leichman, L.P.; et al. p53 point mutations and thymidylate synthase messenger RNA levels in disseminated colorectal cancer: An analysis of response and survival. Clin. Cancer Res. 1998, 4, 1243–1250. [Google Scholar]
- Cheng, T.Y.; Makar, K.W.; Neuhouser, M.L.; Miller, J.W.; Song, X.; Brown, E.C.; Beresford, S.A.; Zheng, Y.; Poole, E.M.; Galbraith, R.L.; et al. Folate-mediated one-carbon metabolism genes and interactions with nutritional factors on colorectal cancer risk: Women’s Health Initiative Observational Study. Cancer 2015, 15, 3684–3691. [Google Scholar] [CrossRef]
- Haerian, B.S.; Haerian, M.S. Evaluation of association studies and meta-analyses of MTHFR gene polymorphisms in colorectal cancer. Pharmacogenomics 2015, 16, 413–425. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Kim, J.W.; Park, H.M.; Kim, J.O.; Jang, H.G.; Oh, J.; Hwang, S.G.; Kwon, S.W.; Oh, D.; Kim, N.K. Genetic variants in 3′-UTRs of methylenetetrahydrofolate reductase (MTHFR) predict olorectal cancer susceptibility in Koreans. Sci. Rep. 2015, 5, 11006. [Google Scholar] [CrossRef] [Green Version]
- Ose, J.; Botma, A.; Balavarca, Y.; Buck, K.; Scherer, D.; Habermann, N.; Beyerle, J.; Pfütze, K.; Seibold, P.; Kap, E.J.; et al. Pathway analysis of genetic variants in folate-mediated one-carbon metabolism-related genes and survival in a prospectively followed cohort of colorectal cancer patients. Cancer Med. 2018, 7, 2797–2807. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.; Carter, J.V.; Eichenberger, M.R.; Netz, U.; Galandiuk, S. Genetic polymorphisms in 5-Fluorouracil-related enzymes predict pathologic response after neoadjuvant chemoradiation for rectal cancer. Surgery 2016, 160, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Horie, N.; Aiba, H.; Oguro, K.; Hojo, H.; Takeishi, K. Functional Analysis and DNA Polymorphism of the Tandemly Repeated Sequences in the 5′-terminal Regulatory Region of the Human Gene for Thymidylate Synthase. Cell Struct. Funct. 1995, 20, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, S.; Collie-Duguid, E.S.; Li, T.; Liu, X.; McLeod, H.L. Ethnic Variation in the Thymidylate Synthase Enhancer Region Polymorphism among Caucasian and Asian Populations. Genomics 1999, 58, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, C.M.; Bigler, J.; Velicer, C.M.; Greene, E.A.; Farin, F.M.; Potter, J.D. Searching expressed sequence tag databases: Discovery and confirmation of a common polymorphism in the thymidylate synthase gene. Cancer Epidemiol. Biomark. Prev. 2000, 9, 1381–1385. [Google Scholar]
- Pullarkat, S.; Stoehlmacher, J.; Ghaderi, V.S.; Xiong, Y.-P.; Ingles, S.A.; Sherrod, A.; Warren, R.M.; Tsaowei, D.D.; Groshen, S.; Lenz, H.-J. Thymidylate synthase gene polymorphism determines response and toxicity of 5-FU chemotherapy. Pharmacogenomics J. 2001, 1, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Mauritz, R.; Giovannetti, E.; Beumer, I.J.; Smid, K.; Van Groeningen, C.J.; Pinedo, H.M.; Peters, G.J. Polymorphisms in the enhancer region of the thymidylate synthase gene are associated with thymidylate synthase levels in normal tissues but not in malignant tissues of patients with colorectal cancer. Clin. Colorectal Cancer 2009, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Dotor, E.; Cuatrecases, M.; Martínez-Iniesta, M.; Navarro, M.; Vilardell, F.; Guinó, E.; Pareja, L.; Figueras, A.; Mollevi, D.G.; Serrano, T.; et al. Tumor Thymidylate Synthase 1494del6 Genotype As a Prognostic Factor in Colorectal Cancer Patients Receiving Fluorouracil-Based Adjuvant Treatment. J. Clin. Oncol. 2006, 24, 1603–1611. [Google Scholar] [CrossRef]
- Figueiredo, J.C.; Levine, A.J.; Crott, J.W.; Baurley, J.; Haile, R.W. Folate-genetics and colorectal neoplasia: What we know and need to know next. Mol. Nutr. Food Res. 2013, 57, 607–627. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, Q.; Miao, Y.-R.; Yang, J.; Yang, W.; Yu, F.; Wang, D.; Guo, A.-Y.; Gong, J. SNP2APA: A database for evaluating effects of genetic variants on alternative polyadenylation in human cancers. Nucleic Acids Res. 2019, 48, D226–D232. [Google Scholar] [CrossRef] [Green Version]
- Charles, S.; Natarajan, J. Two way network construction and analysis of mRNA, miRNA and lncRNA reveals critical regulators and regulatory modules in cardiovascular diseases. Genes Genom. 2020, 42, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E. Metabolic syndrome, hyperinsulinemia, and colon cancer: A review. Am. J. Clin. Nutr. 2007, 86, 836S–842S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.D.; Hahn, L.W.; Roodi, N.; Bailey, L.R.; Dupont, W.D.; Parl, F.F.; Moore, J.H. Multifactor-Dimensionality Reduction Reveals High-Order Interactions among Estrogen-Metabolism Genes in Sporadic Breast Cancer. Am. J. Hum. Genet. 2001, 69, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.H.; Williams, S.M. New strategies for identifying gene-gene interactions in hypertension. Ann. Med. 2002, 34, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Chung, Y.; Elston, R.C.; Kim, Y.; Park, T. Log-linear model-based multifactor dimensionality reduction method to detect gene gene interactions. Bioinformatics 2007, 23, 2589–2595. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Sun, L.; Yang, J.; Li, Y.-Y. 3R variant of thymidylate synthase 5′-untranslated enhanced region contributes to colorectal cancer risk: A meta-analysis. Asian Pac. J. Cancer Prev. 2012, 13, 2605–2610. [Google Scholar] [CrossRef] [Green Version]
- Teng, Z.; Wang, L.; Cai, S.; Yu, P.; Wang, J.; Gong, J.; Liu, Y. The 677C>T (rs1801133) Polymorphism in the MTHFR Gene Contributes to Colorectal Cancer Risk: A Meta-Analysis Based on 71 Research Studies. PLoS ONE 2013, 8, e55332. [Google Scholar] [CrossRef] [Green Version]
- Subbarayan, P.R.; Hammad, H.; Sarkar, M.; Ardalan, B.; Gupta, N. The presence of three repeats in the 5′ UTR region of thymidylate synthase (TS) is associated with increased TS mRNA expression in cultured human cancer cell lines in vitro. Oncol. Rep. 2011, 27, 246–249. [Google Scholar] [CrossRef]
- Wang, S.-M.; Zeng, W.-X.; Wu, W.-S.; Sun, L.-L.; Yan, D. Genotype and allele frequencies of TYMS rs2790 A > G polymorphism in a Chinese paediatric population with acute lymphoblastic leukaemia. J. Clin. Pharm. Ther. 2018, 43, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.N.T.; Santos, A.C.S.; Nogueira, V.M.; Carneiro, C.M.; Lima, A.A. 3′UTR polymorphism of Thymidylate Synthase gene increased the risk of persistence of pre-neoplastic cervical lesions. BMC Cancer 2020, 20, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Tian, S.; Yin, Z.; Wu, S.; Liu, L.; Qian, Y.; Pei, D.; Gao, W.; Xu, J.; Yinet, Y.; et al. MicroRNA-binding site SNPs in deregulated genes are associated with clinical outcome of non-small cell lung cancer. Lung Cancer 2014, 85, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.-P.; Lin, C.-C.; Chen, T.-H.; Tsai, M.-C.; Huang, Y.-C. Higher Plasma Homocysteine Is Associated with Increased Risk of Developing Colorectal Polyps. Nutr. Cancer 2013, 65, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.W.; Beresford, S.A.A.; Neuhouser, M.L.; Cheng, T.-Y.D.; Song, X.; Brown, E.C.; Zheng, Y.; Rodriguez, B.; Green, R.; Ulrich, C.M. Homocysteine, cysteine, and risk of incident colorectal cancer in the Women’s Health Initiative observational cohort. Am. J. Clin. Nutr. 2013, 97, 827–834. [Google Scholar] [CrossRef]
- Brevik, A.; Joshi, A.D.; Corral, R.; Onland-Moret, N.C.; Siegmund, K.D.; Le Marchand, L.; Baron, J.A.; Martinez, M.E.; Haile, R.W.; Ahnen, D.J.; et al. Polymorphisms in Base Excision Repair Genes as Colorectal Cancer Risk Factors and Modifiers of the Effect of Diets High in Red Meat. Cancer Epidemiol. Biomark. Prev. 2010, 19, 3167–3173. [Google Scholar] [CrossRef] [Green Version]
- Branda, R.F.; Blickensderfer, D.B. Folate deficiency increases genetic damage caused by alkylating agents and gamma-irradiation in Chinese hamster ovary cells. Cancer Res. 1993, 53, 5401–5408. [Google Scholar]
- Choi, S.-W.; Kim, Y.-I.; Weitzel, J.N.; Mason, J.B. Folate depletion impairs DNA excision repair in the colon of the rat. Gut 1998, 43, 93–99. [Google Scholar] [CrossRef]
- Duthie, S.J.; Narayanan, S.; Blum, S.; Pirie, L.; Brand, G.M. Folate Deficiency In Vitro Induces Uracil Misincorporation and DNA Hypomethylation and Inhibits DNA Excision Repair in Immortalized Normal Human Colon Epithelial Cells. Nutr. Cancer 2000, 37, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Blount, B.C.; Mack, M.M.; Wehr, C.M.; MacGregor, J.T.; Hiatt, R.A.; Wang, G.; Wickramasinghe, S.N.; Everson, R.B.; Ames, B.N. Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: Implications for cancer and neuronal damage. Proc. Natl. Acad. Sci. USA 1997, 94, 3290–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasco, G.; Zannoni, U.; Paganelli, G.M.; Santucci, R.; Gionchetti, P.; Rivolta, G.; Miniero, R.; Pironi, L.; Calabrese, C.; Di Febo, G.; et al. Folic acid supplementation and cell kinetics of rectal mucosa in patients with ulcerative colitis. Cancer Epidemiol. Biomark. Prev. 1997, 6, 469–471. [Google Scholar]
- Akoglu, B.; Faust, D.; Milovic, V.; Stein, J. Folate and chemoprevention of colorectal cancer: Is 5-methyl-tetrahydrofolate an active antiproliferative agent in folate-treated colon-cancer cells? Nutriens 2001, 17, 652–653. [Google Scholar] [CrossRef]
- van der Put, N.M.; Gabreëls, F.; Stevens, E.M.; Smeitink, J.A.; Trijbels, F.J.; Eskes, T.K.; Heuvel, L.P.V.D.; Blom, H. A Second Common Mutation in the Methylenetetrahydrofolate Reductase Gene: An Additional Risk Factor for Neural-Tube Defects? Am. J. Hum. Genet. 1998, 62, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, S.J.; Albanes, D.; Selhub, J.; Graubard, B.; Lim, U.; Taylor, P.R.; Virtamo, J.; Stolzenberg-Solomon, R. One-Carbon Metabolism Biomarkers and Risk of Colon and Rectal Cancers. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3233–3240. [Google Scholar] [CrossRef] [Green Version]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gao, F.; Zhang, X.-P. miR-203 enhances chemosensitivity to 5-fluorouracil by targeting thymidylate synthase in colorectal cancer. Oncol. Rep. 2014, 33, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Boni, N.; Bitarte, I.; Cristobal, R.; Zarate, J.; Rodriguez, E.; Maiello, J.; Garcia-Foncillas, J.; Bandres, E. miR-192/miR-215 influence 5-fluorouracil resistance through cell cycle-mediated mechanisms complementary to its post-transcriptional thymidilate synthase regulation. Mol. Cancer. Ther. 2010, 9, 2265–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelton, P.M.; Duran, A.; Nakanishi, Y.; Reina-Campos, M.; Kasashima, H.; Llado, V.; Ma, L.; Campos, A.; Olmo, D.G.; García-Arranz, M.; et al. The Secretion of miR-200s by a PKCζ/ADAR2 Signaling Axis Promotes Liver Metastasis in Colorectal Cancer. Cell Rep. 2018, 23, 1178–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Characteristics | Control | CRC | p | Colon | p | Rectum | p |
|---|---|---|---|---|---|---|---|
| N | 400 | 450 | 260 | 186 | |||
| Age: years (mean ± SD) | 60.89 ± 11.72 | 62.05 ± 12.29 | 0.162 | 61.67 ± 12.86 | 0.426 | 62.33 ± 11.46 | 0.165 |
| Gender male: n (%) | 170 (42.5) | 212 (47.1) | 0.177 | 122 (46.9) | 0.299 | 88 (47.3) | 0.316 |
| Hypertension: n (%) | 157 (39.3) | 279 (62.0) | <0.001 | 155 (59.6) | <0.001 | 120 (64.5) | <0.001 |
| Diabetes mellitus: n (%) | 166 (41.5) | 253 (56.2) | <0.001 | 147 (56.5) | <0.001 | 104 (55.9) | 0.002 |
| BMI ≥ 25 kg/m2: n (%) | 93 (23.3) | 116 (25.8) | 0.393 | 63 (24.2) | 0.845 | 51 (27.4) | 0.323 |
| HDL-C < 40 (male) or 50 (female) mg/dL: n (%) | 78 (19.5) | 197 (43.8) | <0.001 | 111 (42.7) | <0.001 | 84 (45.2) | <0.001 |
| Homocysteine: μmol/L (n) | 9.80 ± 4.17 (395) | 10.51 ± 7.76 (383) | 0.115 | 10.26 ± 8.16 (220) | 0.362 | 10.77 ± 7.24 (159) | 0.050 |
| Folate: ng/mL (n) | 8.85 ± 8.06 (392) | 7.77 ± 6.65 (381) | 0.043 | 8.06 ± 7.19 (220) | 0.226 | 7.35 ± 5.85 (157) | 0.034 |
| Tumor size: n (%) | |||||||
| <5 cm | - | 181 (40.2) | NA | 92 (35.4) | NA | 87 (46.8) | NA |
| ≥5 cm | - | 269 (59.8) | NA | 168 (64.6) | NA | 99 (53.2) | NA |
| TNM stage: n (%) | |||||||
| I | - | 42 (9.3) | NA | 22 (8.5) | NA | 20 (10.8) | NA |
| II | - | 189 (42.0) | NA | 115 (44.2) | NA | 71 (38.2) | NA |
| III | - | 173 (38.4) | NA | 94 (36.2) | NA | 79 (42.5) | NA |
| IV | - | 46 (10.2) | NA | 29 (11.2) | NA | 16 (8.6) | NA |
| Genotypes | Control (n = 400) | CRC (n = 450) | AOR (95% CI) | p | Colon (n = 260) | AOR (95% CI) | p | Rectum (n = 186) | AOR (95% CI) | p |
|---|---|---|---|---|---|---|---|---|---|---|
| TS 1100T>C | ||||||||||
| TT | 195 (48.8) | 223 (49.6) | 1.00 (ref) | NA | 134 (51.5) | 1.00 (ref) | NA | 86 (46.2) | 1.00 (ref) | NA |
| TC | 177 (44.3) | 189 (42.0) | 0.99 (0.73–1.33) | 0.922 | 105 (40.4) | 0.88 (0.62–1.25) | 0.482 | 84 (45.2) | 1.21 (0.82–1.78) | 0.345 |
| CC | 28 (7.0) | 38 (8.4) | 1.27 (0.73–2.22) | 0.401 | 21 (8.1) | 1.25 (0.66–2.37) | 0.489 | 16 (8.6) | 1.28 (0.62–2.64) | 0.511 |
| TS 1170A>G | ||||||||||
| AA | 195 (48.8) | 155 (34.4) | 1.00 (ref) | NA | 82 (31.5) | 1.00 (ref) | NA | 71 (38.2) | 1.00 (ref) | NA |
| AG | 176 (44.0) | 224 (49.8) | 1.55 (1.13–2.11) | 0.006 | 132 (50.8) | 1.73 (1.20–2.50) | 0.003 | 90 (48.4) | 1.42 (0.95–2.11) | 0.085 |
| GG | 29 (7.3) | 71 (15.8) | 3.19 (1.91–5.34) | <0.001 | 46 (17.7) | 4.31 (2.42–7.66) | <0.001 | 25 (13.4) | 2.07 (1.08–3.95) | 0.028 |
| Genotypes | Control (n = 400) | CRC (n = 450) | AOR (95% CI) | p | Colon (n = 260) | AOR (95% CI) | p | Rectum (n = 186) | AOR (95% CI) | p |
|---|---|---|---|---|---|---|---|---|---|---|
| TS Genotype | ||||||||||
| 1100TT-1170AA | 63 (15.8) | 39 (8.7) | 1.00 (ref) | NA | 23 (8.8) | 1.00 (ref) | NA | 15 (8.1) | 1.00 (ref) | NA |
| 1100TT-1170AG | 103 (25.8) | 113 (25.1) | 1.81 (1.07–3.06) | 0.027 | 65 (25.0) | 1.75 (0.95–3.25) | 0.074 | 46 (24.7) | 1.94 (0.96–3.93) | 0.067 |
| 1100TT-1170GG | 29 (7.3) | 71 (15.8) | 5.24 (2.67–10.27) | <0.001 | 46 (17.7) | 6.10 (2.81–13.23) | <0.001 | 25 (13.4) | 4.04 (1.69–9.69) | 0.002 |
| 1100TC-1170AA | 104 (26.0) | 78 (17.3) | 1.40 (0.82–2.40) | 0.220 | 38 (14.6) | 1.10 (0.57–2.12) | 0.785 | 40 (21.5) | 1.85 (0.90–3.79) | 0.095 |
| 1100TC-1170AG | 73 (18.3) | 111 (24.7) | 3.06 (1.73–5.43) | <0.001 | 67 (25.8) | 3.09 (1.59–6.00) | <0.001 | 44 (23.7) | 3.43 (1.57–7.45) | 0.002 |
| 1100CC-1170AA | 28 (7.0) | 38 (8.4) | 3.00 (1.45–6.21) | 0.003 | 21 (8.1) | 3.12 (1.34–7.28) | 0.009 | 16 (8.6) | 2.98 (1.11–8.01) | 0.030 |
| TS haplotype | ||||||||||
| 1100T-1170A | 333 (41.6) | 269 (29.9) | 1.00 (ref) | NA | 149 (28.7) | 1.00 (ref) | NA | 116 (31.2) | 1.00 (ref) | NA |
| 1100T-1170G | 234 (29.3) | 366 (40.7) | 2.00 (1.56–2.55) | <0.001 | 224 (43.1) | 2.25 (1.69–3.00) | <0.001 | 140 (37.6) | 1.71 (1.24–2.34) | <0.001 |
| 1100C-1170A | 233 (29.1) | 265 (29.4) | 1.51 (1.17–1.96) | 0.002 | 147 (28.3) | 1.57 (1.16–2.13) | 0.004 | 116 (31.2) | 1.49 (1.08–2.07) | 0.017 |
| TS 1100TT-1170AG | TS 1100TT-1170GG | TS 1100TC-1170AA | TS 1100TC-1170AG | TS 1100CC-1170AA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Subsets | AOR (95% CI) | p | AOR (95% CI) | p | AOR (95% CI) | p | AOR (95% CI) | p | AOR (95% CI) | p |
| Age < 62 years | 2.08 (0.98–4.43) | 0.058 | 3.38 (1.34–8.52) | 0.010 | 1.07 (0.50–2.32) | 0.856 | 2.78 (1.24–6.24) | 0.013 | 1.48 (0.53–4.14) | 0.454 |
| ≥62 years | 1.79 (0.85–3.77) | 0.126 | 7.62 (2.68–21.68) | <0.001 | 1.87 (0.85–4.12) | 0.122 | 3.08 (1.35–7.04) | 0.008 | 6.63 (2.10–20.97) | 0.001 |
| Gender Male | 2.12 (0.90–5.01) | 0.086 | 4.61 (1.80–11.78) | 0.001 | 1.94 (0.80–4.67) | 0.142 | 3.57 (1.51–8.43) | 0.004 | 3.45 (1.16–10.29) | 0.026 |
| Female | 1.62 (0.82–3.18) | 0.162 | 6.23 (2.24–17.34) | <0.001 | 1.02 (0.50–2.07) | 0.953 | 2.70 (1.21–6.04) | 0.015 | 2.91 (1.06–8.01) | 0.039 |
| TS < 5 cm | 2.13 (1.06–4.27) | 0.034 | 5.83 (2.37–14.32) | <0.001 | 1.71 (0.82–3.57) | 0.154 | 3.88 (1.74–8.65) | <0.001 | 2.41 (0.83–7.00) | 0.106 |
| ≥5 cm | 1.65 (0.88–3.09) | 0.120 | 5.03 (2.35–10.80) | <0.001 | 1.24 (0.65–2.37) | 0.506 | 2.97 (1.53–5.77) | 0.001 | 3.71 (1.61–8.54) | 0.002 |
| TNM I + II | 2.20 (1.13–4.31) | 0.021 | 6.24 (2.74–14.18) | <0.001 | 1.63 (0.81–3.25) | 0.169 | 3.87 (1.89–7.90) | <0.001 | 3.35 (1.33–8.43) | 0.010 |
| III + IV | 1.53 (0.81–2.89) | 0.194 | 4.56 (2.01–10.36) | <0.001 | 1.22 (0.62–2.37) | 0.564 | 2.62 (1.28–5.34) | 0.008 | 3.00 (1.23–7.29) | 0.015 |
| HTN No | 1.40 (0.66–3.00) | 0.379 | 4.88 (1.87–12.78) | 0.001 | 1.13 (0.53–2.44) | 0.752 | 2.60 (1.17–5.79) | 0.019 | 2.07 (0.73–5.85) | 0.172 |
| Yes | 2.43 (1.16–5.11) | 0.019 | 5.61 (2.10–14.96) | <0.001 | 2.00 (0.91–4.36) | 0.083 | 3.86 (1.64–9.07) | 0.002 | 4.59 (1.58–13.31) | 0.005 |
| DM No | 3.29 (1.38–7.84) | 0.007 | 5.93 (2.27–15.50) | <0.001 | 2.44 (0.96–6.20) | 0.061 | 3.56 (1.43–8.86) | 0.006 | 2.09 (0.74–5.95) | 0.167 |
| Yes | 1.11 (0.55–2.23) | 0.773 | 4.60 (1.70–12.43) | 0.003 | 0.93 (0.45–1.92) | 0.840 | 2.62 (1.22–5.65) | 0.014 | 3.85 (1.28–11.54) | 0.016 |
| BMI < 25 kg/m2 | 2.13 (1.13–3.99) | 0.019 | 4.96 (2.27–10.84) | <0.001 | 1.27 (0.67–2.41) | 0.470 | 2.67 (1.39–5.12) | 0.003 | 3.96 (1.67–9.39) | 0.002 |
| ≥25 kg/m2 | 1.26 (0.48–3.33) | 0.643 | 6.41 (1.41–29.18) | 0.016 | 1.85 (0.66–5.17) | 0.240 | 5.07 (1.41–18.22) | 0.013 | 1.57 (0.35–7.08) | 0.558 |
| HDL-C < 40 (M)/50 (F) | 1.37 (0.57–3.26) | 0.479 | 2.82 (0.90–8.89) | 0.077 | 0.96 (0.38–2.45) | 0.934 | 3.71 (1.33–10.39) | 0.013 | 1.44 (0.35–5.98) | 0.618 |
| ≥40 (M)/50 (F) | 2.12 (1.07–4.20) | 0.031 | 7.69 (3.21–18.44) | <0.001 | 1.64 (0.82–3.29) | 0.163 | 3.02 (1.48–6.16) | 0.002 | 3.60 (1.50–8.66) | 0.004 |
| Hcy < 11.7 μmol/L | 2.11 (1.10–4.07) | 0.025 | 5.45 (2.40–12.35) | <0.001 | 1.41 (0.73–2.72) | 0.308 | 2.88 (1.40–5.96) | 0.004 | 3.14 (1.25–7.89) | 0.015 |
| ≥11.7 μmol/L | 0.97 (0.29–3.20) | 0.957 | 4.14 (0.96–17.85) | 0.057 | 1.87 (0.53–6.55) | 0.327 | 2.88 (0.82–10.14) | 0.100 | 4.12 (0.73–23.18) | 0.108 |
| FA < 4.58 ng/mL | 1.52 (0.47–4.86) | 0.485 | 3.31 (0.59–18.52) | 0.173 | 0.92 (0.27–3.08) | 0.888 | 2.44 (0.71–8.31) | 0.156 | 3.47 (0.55–21.80) | 0.184 |
| ≥4.58 ng/mL | 2.00 (1.02–3.92) | 0.043 | 8.36 (3.48–20.08) | <0.001 | 1.53 (0.77–3.04) | 0.228 | 3.43 (1.62–7.27) | 0.001 | 3.84 (1.43–10.35) | 0.008 |
| 1100T-1170G | 1100C-1170A | |||
|---|---|---|---|---|
| Subsets | AOR (95% CI) | p | AOR (95% CI) | p |
| Age < 62 years | 1.91 (1.35–2.72) | <0.001 | 1.22 (0.85–1.76) | 0.274 |
| ≥62 years | 1.95 (1.37–2.79) | <0.001 | 1.75 (1.20–2.53) | 0.003 |
| Gender Male | 2.08 (1.43–3.02) | <0.001 | 1.72 (1.16–2.56) | 0.007 |
| Female | 1.91 (1.37–2.68) | <0.001 | 1.37 (0.97–1.93) | 0.074 |
| Tumor size < 5 cm | 1.86 (1.36–2.53) | <0.001 | 1.32 (0.95–1.84) | 0.103 |
| ≥5 cm | 2.10 (1.57–2.81) | <0.001 | 1.70 (1.26–2.30) | <0.001 |
| TNM stage I + II | 2.15 (1.60–2.90) | <0.001 | 1.57 (1.15–2.15) | 0.005 |
| III + IV | 1.82 (1.35–2.46) | <0.001 | 1.47 (1.07–2.02) | 0.017 |
| HTN No | 1.99 (1.40–2.83) | <0.001 | 1.43 (0.99–2.07) | 0.059 |
| Yes | 1.97 (1.39–2.80) | <0.001 | 1.64 (1.14–2.35) | 0.008 |
| DM No | 2.04 (1.44–2.90) | <0.001 | 1.27 (0.87–1.84) | 0.214 |
| Yes | 1.90 (1.34–2.70) | <0.001 | 1.77 (1.23–2.54) | 0.002 |
| BMI < 25 kg/m2 | 2.00 (1.51–2.67) | <0.001 | 1.48 (1.10–1.99) | 0.009 |
| ≥25 kg/m2 | 1.97 (1.20–3.23) | 0.008 | 1.66 (0.98–2.82) | 0.061 |
| HDL-C < 40 (M)/50 (F) mg/dL | 1.84 (1.17–2.89) | 0.008 | 1.35 (0.83–2.19) | 0.221 |
| ≥40 (M)/50 (F) mg/dL | 2.09 (1.55–2.81) | <0.001 | 1.56 (1.15–2.12) | 0.005 |
| Hcy < 11.7 μmol/L | 1.97 (1.46–2.66) | <0.001 | 1.35 (0.99–1.83) | 0.060 |
| ≥11.7 μmol/L | 1.80 (1.05–3.08) | 0.031 | 1.92 (1.08–3.43) | 0.027 |
| FA < 4.58 ng/mL | 1.75 (1.00–3.07) | 0.049 | 1.52 (0.87–2.65) | 0.141 |
| ≥4.58 ng/mL | 2.12 (1.57–2.86) | <0.001 | 1.50 (1.09–2.06) | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, Y.-J.; Cho, S.-H.; Kim, E.-J.; Ryu, C.-S.; Park, H.-S.; Kim, J.-W.; Lee, J.-Y.; An, H.-J.; Kim, N.-K. 3′-UTR Polymorphisms in Thymidylate Synthase with Colorectal Cancer Prevalence and Prognosis. J. Pers. Med. 2021, 11, 537. https://doi.org/10.3390/jpm11060537
Jeon Y-J, Cho S-H, Kim E-J, Ryu C-S, Park H-S, Kim J-W, Lee J-Y, An H-J, Kim N-K. 3′-UTR Polymorphisms in Thymidylate Synthase with Colorectal Cancer Prevalence and Prognosis. Journal of Personalized Medicine. 2021; 11(6):537. https://doi.org/10.3390/jpm11060537
Chicago/Turabian StyleJeon, Young-Joo, Sung-Hwan Cho, Eo-Jin Kim, Chang-Soo Ryu, Han-Sung Park, Jong-Woo Kim, Jeong-Yong Lee, Hui-Jeong An, and Nam-Keun Kim. 2021. "3′-UTR Polymorphisms in Thymidylate Synthase with Colorectal Cancer Prevalence and Prognosis" Journal of Personalized Medicine 11, no. 6: 537. https://doi.org/10.3390/jpm11060537