
Identification of Novel Biomarkers and Candidate Drug in Ovarian Cancer

 ,
,  ,
,  , , , and
, , , and 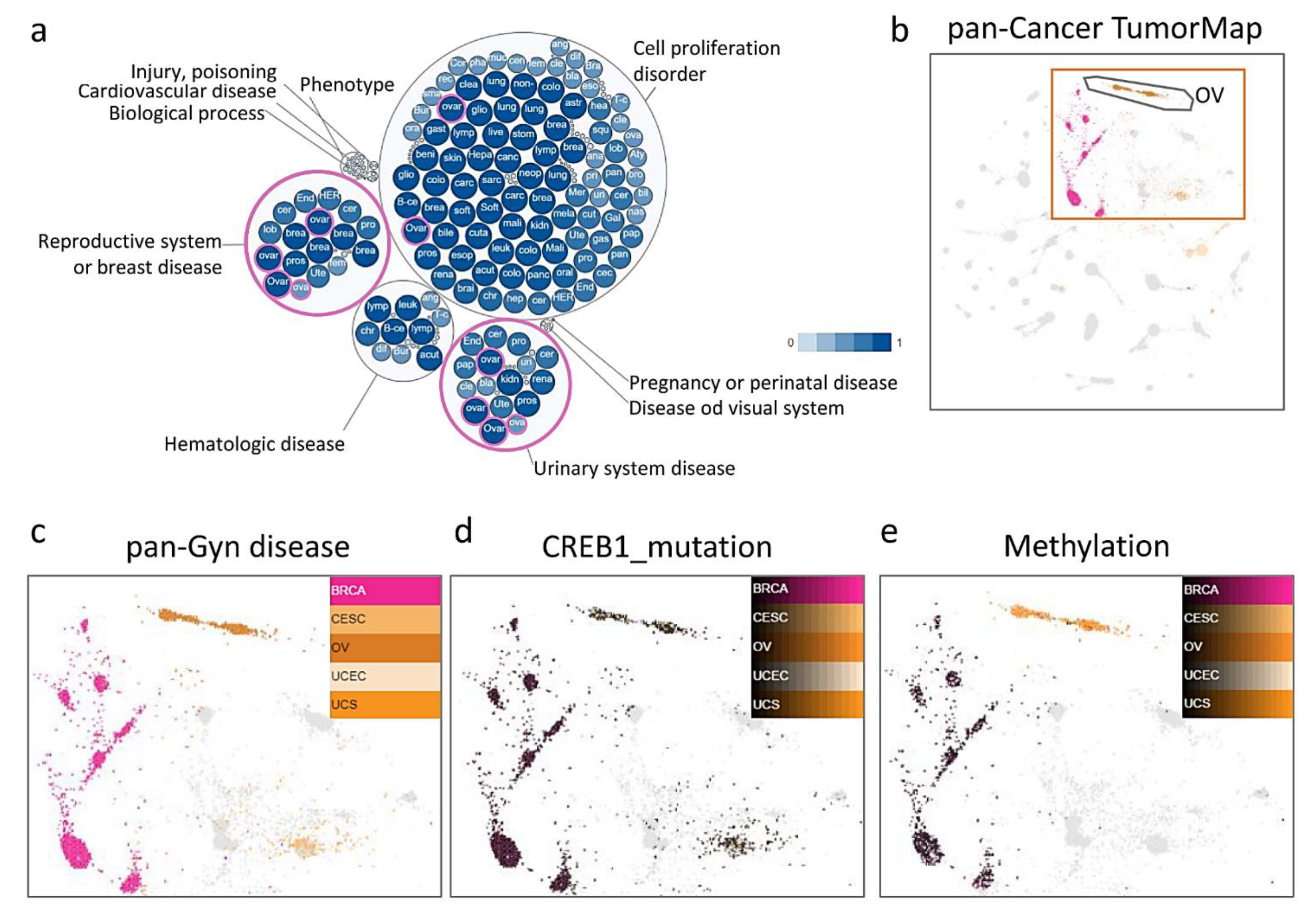{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. RNA Extraction and Real-Time PCR
2.3. Tissue microarrays (TMA) and Immunohistochemistry (IHC) Analysis
2.4. Multi-Omics Analysis
2.5. Statistical Analyses
3. Results
3.1. TumorMap and Integrated Cluster Identify Significant Features Distinguishing OV among PanCancer-33 Tumors
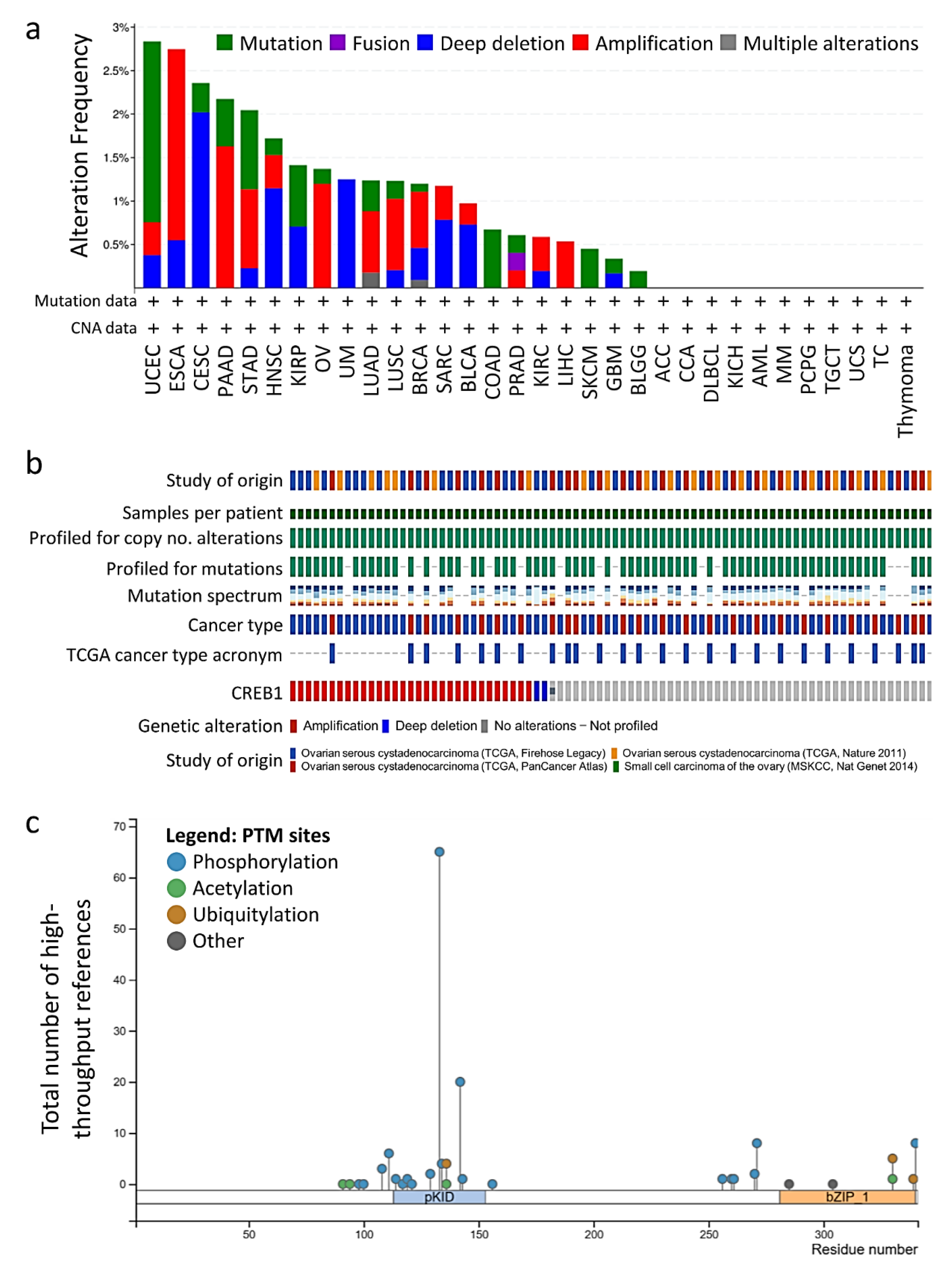
3.2. CREB1 Gene Mutation Predicts A Poorer Disease-Free Survival in OV Patients
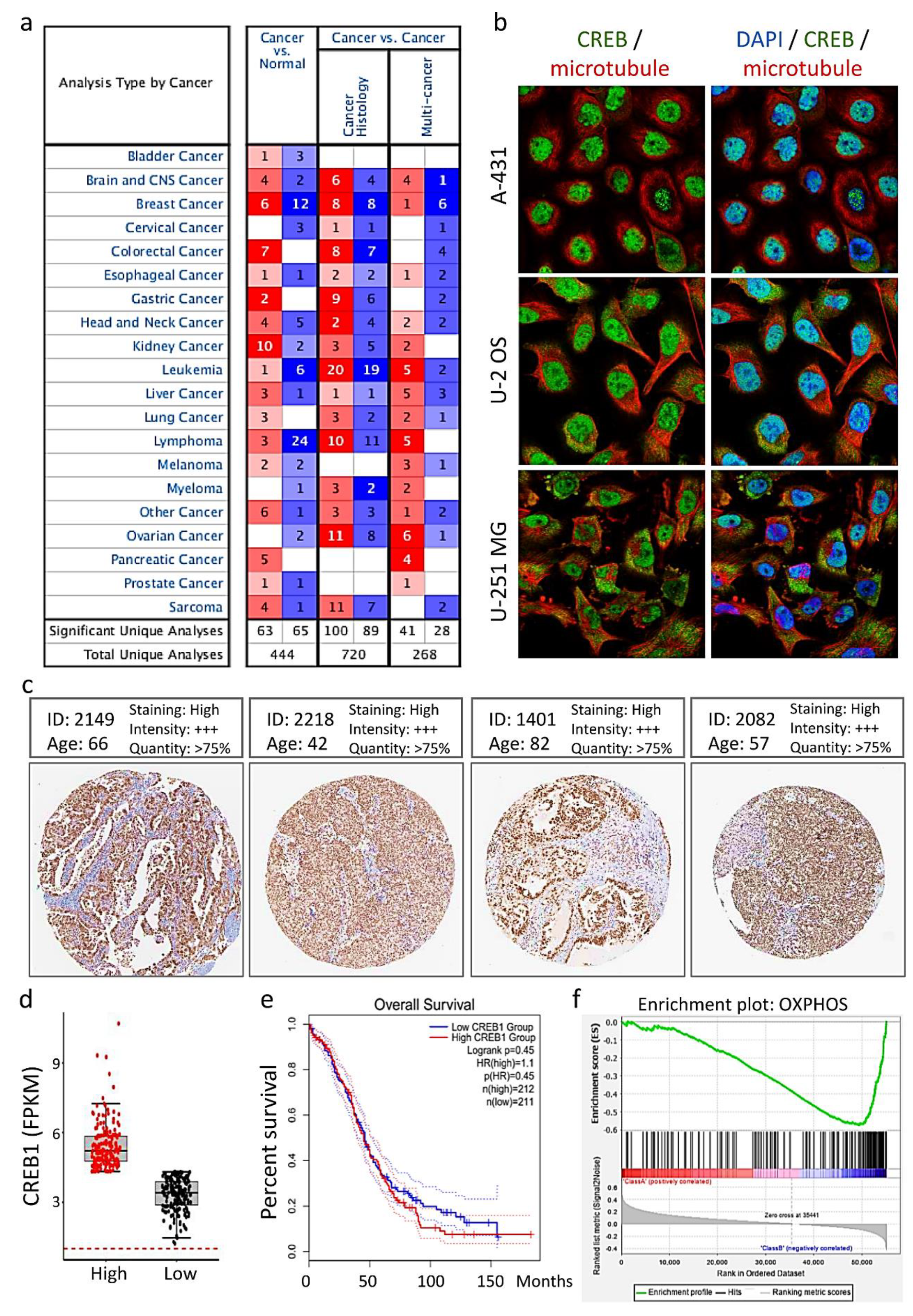
3.3. Distribution and Expression of CREB1 in Cancer Tissues of Patients with OV
3.4. Tissue Microarray Analysis of CREB1 Expression
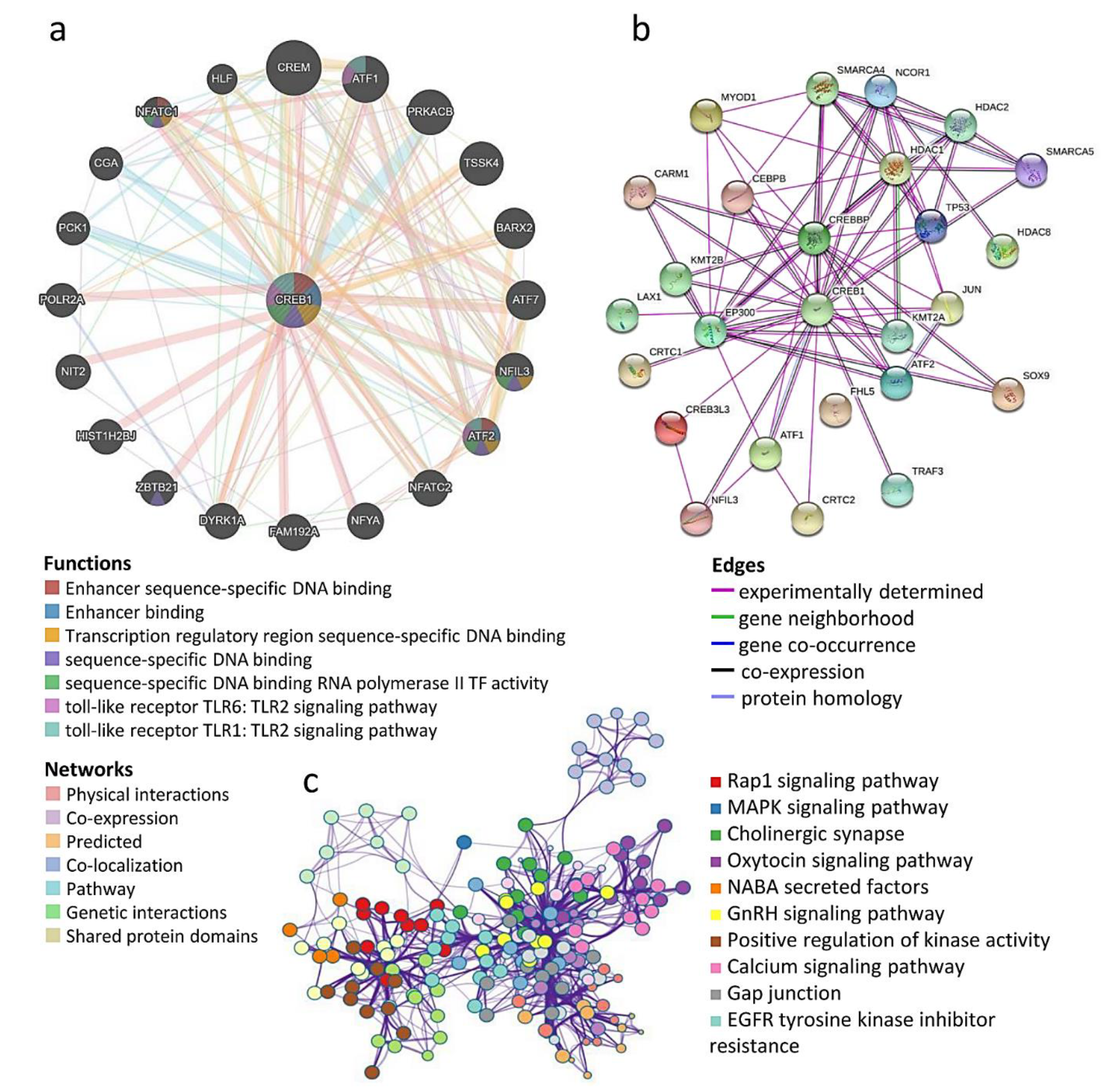
3.5. Prediction of Protein–Protein Interaction of CREB1 Mutations and Copy Number Alterations
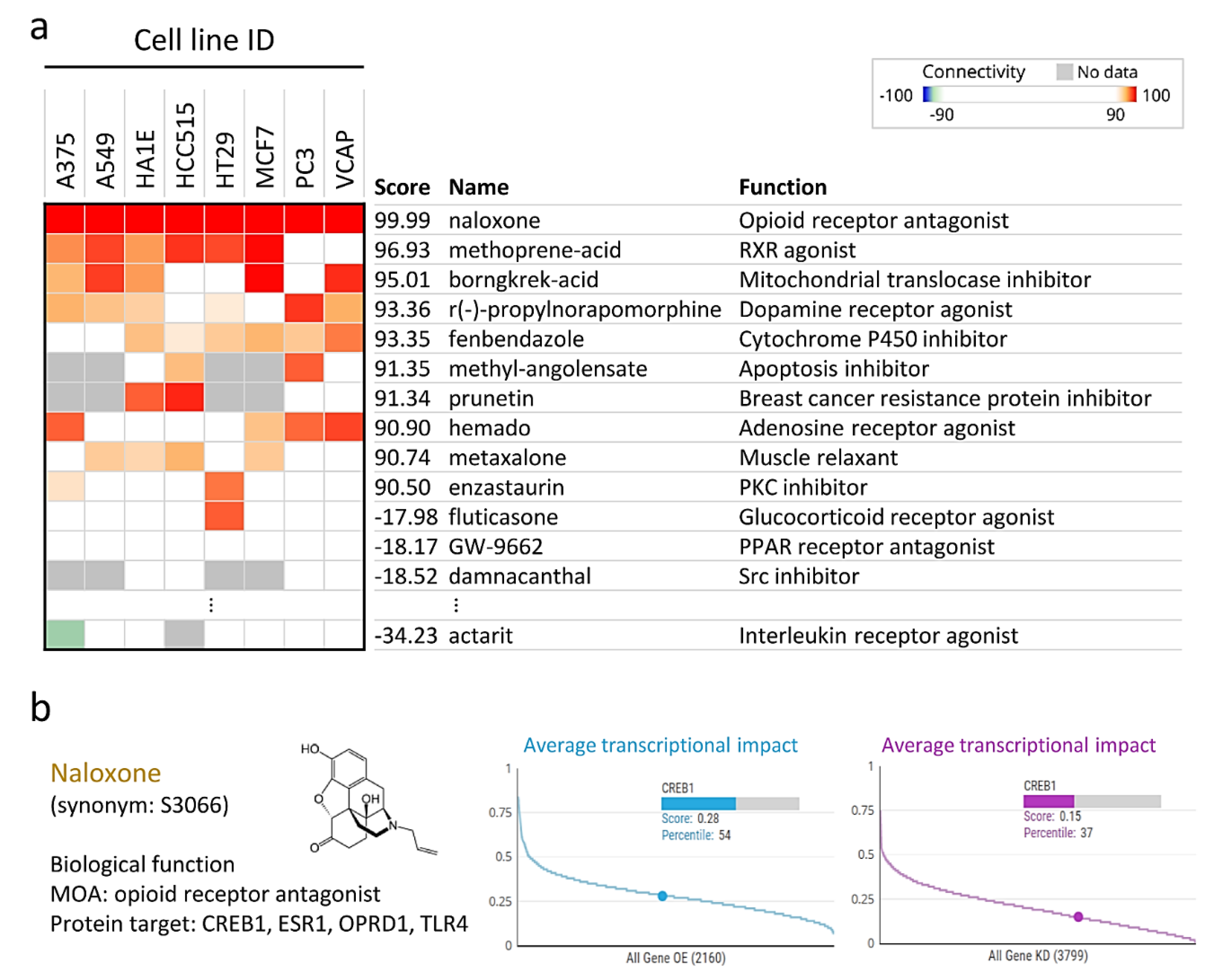
3.6. Naloxone Treatment Mimics the Gene Expression Profile of CREB1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Yang, Y.; Zhou, N.; Tang, K.; Lau, W.B.; Lau, B.; Wang, W.; Xu, L.; Yang, Z.; Huang, S.; et al. Epigenetics in ovarian cancer: Premise, properties, and perspectives. Mol. Cancer 2018, 17, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, S.; Kim, S.-W.; Ryu, S.-H.; Chung, W.-C.; Koo, J.S. Growth Suppression of Lung Cancer Cells by Targeting Cyclic AMP Response Element-Binding Protein. Cancer Res. 2008, 68, 981–988. [Google Scholar] [CrossRef] [Green Version]
- Montminy, M.R.; Bilezikjian, L.M. Binding of a nuclear protein to the cyclic-AMP response element of the somatostatin gene. Nat. Cell Biol. 1987, 328, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Antony, N.; McDougall, A.R.; Mantamadiotis, T.; Cole, T.J.; Bird, A.D. Creb1 regulates late stage mammalian lung development via respiratory epithelial and mesenchymal-independent mechanisms. Sci. Rep. 2016, 6, 25569. [Google Scholar] [CrossRef] [Green Version]
- Comuzzi, B.; Lambrinidis, L.; Rogatsch, H.; Godoy-Tundidor, S.; Knezevic, N.; Krhen, I.; Mareković, Z.; Bartsch, G.; Klocker, H.; Hobisch, A.; et al. The Transcriptional Co-Activator cAMP Response Element-Binding Protein-Binding Protein Is Expressed in Prostate Cancer and Enhances Androgen- and Anti-Androgen-Induced Androgen Receptor Function. Am. J. Pathol. 2003, 162, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Gubbay, O.; Rae, M.T.; McNeilly, A.S.; Donadeu, F.X.; Zeleznik, A.J.; Hillier, S.G. cAMP response element-binding (CREB) signalling and ovarian surface epithelial cell survival. J. Endocrinol. 2006, 191, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Lee, H.Y.; Crawley, S.; Hokari, R.; Kwon, S. Kim Bile acid regulates MUC2 transcription in colon cancer cells via positive EGFR/PKC/Ras/ERK/CREB, PI3K/Akt/IκB/NF-κB and p38/MSK1/CREB pathways and negative JNK/c-Jun/AP-1 pathway. Int. J. Oncol. 2010, 36, 941–953. [Google Scholar] [CrossRef] [Green Version]
- Son, J.; Lee, J.; Kim, H.; Ha, H.; Lee, Z. Camp-response-element-binding protein positively regulates breast cancer metastasis and subsequent bone destruction. Bone 2011, 48, S34. [Google Scholar] [CrossRef]
- Lin, Y.; Miao, Z.; Wu, Y.; Ge, F.-F.; Wen, Q.-P. Effect of low dose naloxone on the immune system function of a patient undergoing video-assisted thoracoscopic resection of lung cancer with sufentanil controlled analgesia—A randomized controlled trial. BMC Anesthesiol. 2019, 19, 236. [Google Scholar] [CrossRef] [Green Version]
- Chiang, A.-J.; Li, C.-J.; Tsui, K.-H.; Chang, C.; Chang, Y.-C.I.; Chen, L.-W.; Chang, T.-H.; Sheu, J.J.-C. UBE2C Drives Human Cervical Cancer Progression and Is Positively Modulated by mTOR. Biomolecules 2020, 11, 37. [Google Scholar] [CrossRef]
- Newton, Y.; Novak, A.M.; Swatloski, T.; McColl, D.C.; Chopra, S.; Graim, K.; Weinstein, A.S.; Baertsch, R.; Salama, S.R.; Ellrott, K.; et al. TumorMap: Exploring the Molecular Similarities of Cancer Samples in an Interactive Portal. Cancer Res. 2017, 77, e111–e114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Caudy, M.; Garapati, P.; Gillespie, M.; Kamdar, M.R.; et al. The Reactome pathway knowledgebase. Nucleic Acids Res. 2014, 42, D472–D477. [Google Scholar] [CrossRef]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F.; McKay, S.; et al. The Reactome pathway Knowledgebase. Nucleic Acids Res. 2016, 44, D481–D487. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452. [Google Scholar] [CrossRef]
- Chen, S.-N.; Chang, R.; Lin, L.-T.; Chern, C.-U.; Tsai, H.-W.; Wen, Z.-H.; Li, Y.-H.; Li, C.-J.; Tsui, K.-H. MicroRNA in Ovarian Cancer: Biology, Pathogenesis, and Therapeutic Opportunities. Int. J. Environ. Res. Public Health 2019, 16, 1510. [Google Scholar] [CrossRef] [Green Version]
- Graffeo, R.; Livraghi, L.; Pagani, O.; Goldhirsch, A.; Partridge, A.H.; Garber, J.E. Time to incorporate germline multigene panel testing into breast and ovarian cancer patient care. Breast Cancer Res. Treat. 2016, 160, 393–410. [Google Scholar] [CrossRef]
- Hsu, H.-C.; Tsai, S.-Y.; Wu, S.-L.; Jeang, S.-R.; Ho, M.-Y.; Liou, W.-S.; Chiang, A.-J.; Chang, T.-H. Longitudinal perceptions of the side effects of chemotherapy in patients with gynecological cancer. Support. Care Cancer 2017, 25, 3457–3464. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-H.; Lin, L.-T.; Li, C.-J.; Kao, P.-G.; Tsai, H.-W.; Chen, S.-N.; Wen, Z.-H.; Wang, P.-H.; Tsui, K.-H. Combining Bioinformatics and Experiments to Identify CREB1 as a Key Regulator in Senescent Granulosa Cells. Diagnostics 2020, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Somers, J.P.; DeLoia, J.A.; Zeleznik, A.J. Adenovirus-directed expression of a nonphosphorylatable mutant of CREB (cAMP response element-binding protein) adversely affects the survival, but not the differentiation, of rat granulosa cells. Mol. Endocrinol. 1999, 13, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Alper, Ö.; Bergmann-Leitner, E.S.; Abrams, S.; Cho-Chung, Y.S. Apoptosis, growth arrest and suppression of invasiveness by CRE-decoy oligonucleotide in ovarian cancer cells: Protein kinase A downregulation and cytoplasmic export of CRE-binding proteins. Mol. Cell. Biochem. 2001, 218, 55–63. [Google Scholar] [CrossRef]
- Li, J.-Y.; Li, C.-J.; Lin, L.-T.; Tsui, K.-H. Multi-Omics Analysis Identifying Key Biomarkers in Ovarian Cancer. Cancer Control. 2020, 27, 1073274820976671. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yin, M.; Wang, Y. CREB1, a direct target of miR-122, promotes cell proliferation and invasion in bladder cancer. Oncol. Lett. 2018, 16, 3842–3848. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Tucker-Burden, C.; Kaissi, E.; Newsam, A.; Duggireddy, H.; Chau, M.; Zhang, C.; Diwedi, B.; Rupji, M.; Seby, S.; et al. CDK5 Inhibition Resolves PKA/cAMP-Independent Activation of CREB1 Signaling in Glioma Stem Cells. Cell Rep. 2018, 23, 1651–1664. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.; Zhu, Y.; Cong, X.; Li, Q. Knockdown of CREB1 inhibits tumor growth of human gastric cancer in vitro and in vivo. Oncol. Rep. 2017, 37, 3361–3368. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.W.; Huang, Z.H.; Liao, L.; Zhang, Q.H.; Li, J.Q.; Zheng, C.C.; He, Y.; Luo, T.T.; Wang, Y.; Hu, H.F.; et al. Direct Targeting of CREB1 with Imperatorin Inhibits TGFbeta2-ERK Signaling to Suppress Esophageal Cancer Metastasis. Adv. Sci. 2020, 7, 2000925. [Google Scholar]
- Li, B.X.; Gardner, R.; Xue, C.; Qian, D.Z.; Xie, F.; Thomas, G.; Kazmierczak, S.C.; Habecker, B.A.; Xiao, X. Systemic Inhibition of CREB is Well-tolerated in vivo. Sci. Rep. 2016, 6, 34513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhabra, A.; Fernando, H.; Watkins, G.; Mansel, R.E.; Jiang, W.G. Expression of transcription factor CREB1 in human breast cancer and its correlation with prognosis. Oncol. Rep. 2007, 18, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Wang, S.; Zhu, L.; Wu, C.; Yin, B.; Zhao, J.; Yuan, J.; Qiang, B.; Peng, X. cAMP response element-binding protein promotes gliomagenesis by modulating the expression of oncogenic microRNA-23a. Proc. Natl. Acad. Sci. USA 2012, 109, 15805–15810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, C.; Sklan, E.H.; Soreq, H. CREB regulates AChE-R-induced proliferation of human glioblastoma cells. Neoplasia 2004, 6, 279–286. [Google Scholar] [CrossRef]
- Sandoval, S.; Pigazzi, M.; Sakamoto, K.M. CREB: A Key Regulator of Normal and Neoplastic Hematopoiesis. Adv. Hematol. 2009, 2009, 634292. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-W.; Chen, X.; Gao, J.-W.; Zhang, H.; Ma, R.-R.; Gao, Z.-H.; Gao, P. High expression of cAMP responsive element binding protein 1 (CREB1) is associated with metastasis, tumor stage and poor outcome in gastric cancer. Oncotarget 2015, 6, 10646–10657. [Google Scholar] [CrossRef]
- Tian, T.; Chen, Z.-H.; Zheng, Z.; Liu, Y.; Zhao, Q.; Liu, Y.; Qiu, H.; Long, Q.; Chen, M.; Li, L.; et al. Investigation of the role and mechanism of ARHGAP5-mediated colorectal cancer metastasis. Theranostics 2020, 10, 5998–6010. [Google Scholar] [CrossRef]
- Li, C.-J.; Lin, H.-Y.; Ko, C.-J.; Lai, J.-C.; Chu, P.-Y. A Novel Biomarker Driving Poor-Prognosis Liver Cancer: Overexpression of the Mitochondrial Calcium Gatekeepers. Biomedicines 2020, 8, 451. [Google Scholar] [CrossRef]
- Tsui, K.-H.; Wu, M.-Y.; Lin, L.-T.; Wen, Z.-H.; Li, Y.-H.; Chu, P.-Y.; Li, C.-J. Disruption of mitochondrial homeostasis with artemisinin unravels anti-angiogenesis effects via auto-paracrine mechanisms. Theranostics 2019, 9, 6631–6645. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Jiang, Q.; Ma, R.; Zhang, H.; Tong, D.; Tang, K.; Wang, X.; Ni, L.; Miao, J.; Duan, B.; et al. Norepinephrine-CREB1-miR-373 axis promotes progression of colon cancer. Mol. Oncol. 2020, 14, 1059–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.-J.; Lin, L.-T.; Chu, P.-Y.; Chiang, A.-J.; Tsai, H.-W.; Chiu, Y.-H.; Huang, M.-S.; Wen, Z.-H.; Tsui, K.-H. Identification of Novel Biomarkers and Candidate Drug in Ovarian Cancer. J. Pers. Med. 2021, 11, 316. https://doi.org/10.3390/jpm11040316
Li C-J, Lin L-T, Chu P-Y, Chiang A-J, Tsai H-W, Chiu Y-H, Huang M-S, Wen Z-H, Tsui K-H. Identification of Novel Biomarkers and Candidate Drug in Ovarian Cancer. Journal of Personalized Medicine. 2021; 11(4):316. https://doi.org/10.3390/jpm11040316
Chicago/Turabian StyleLi, Chia-Jung, Li-Te Lin, Pei-Yi Chu, An-Jen Chiang, Hsiao-Wen Tsai, Yi-Han Chiu, Mei-Shu Huang, Zhi-Hong Wen, and Kuan-Hao Tsui. 2021. "Identification of Novel Biomarkers and Candidate Drug in Ovarian Cancer" Journal of Personalized Medicine 11, no. 4: 316. https://doi.org/10.3390/jpm11040316